Impact of Polyphenol Antioxidants on Cycling Performance and Cardiovascular Function

Abstract
:1. Introduction
2. Experimental Section
2.1. Subjects
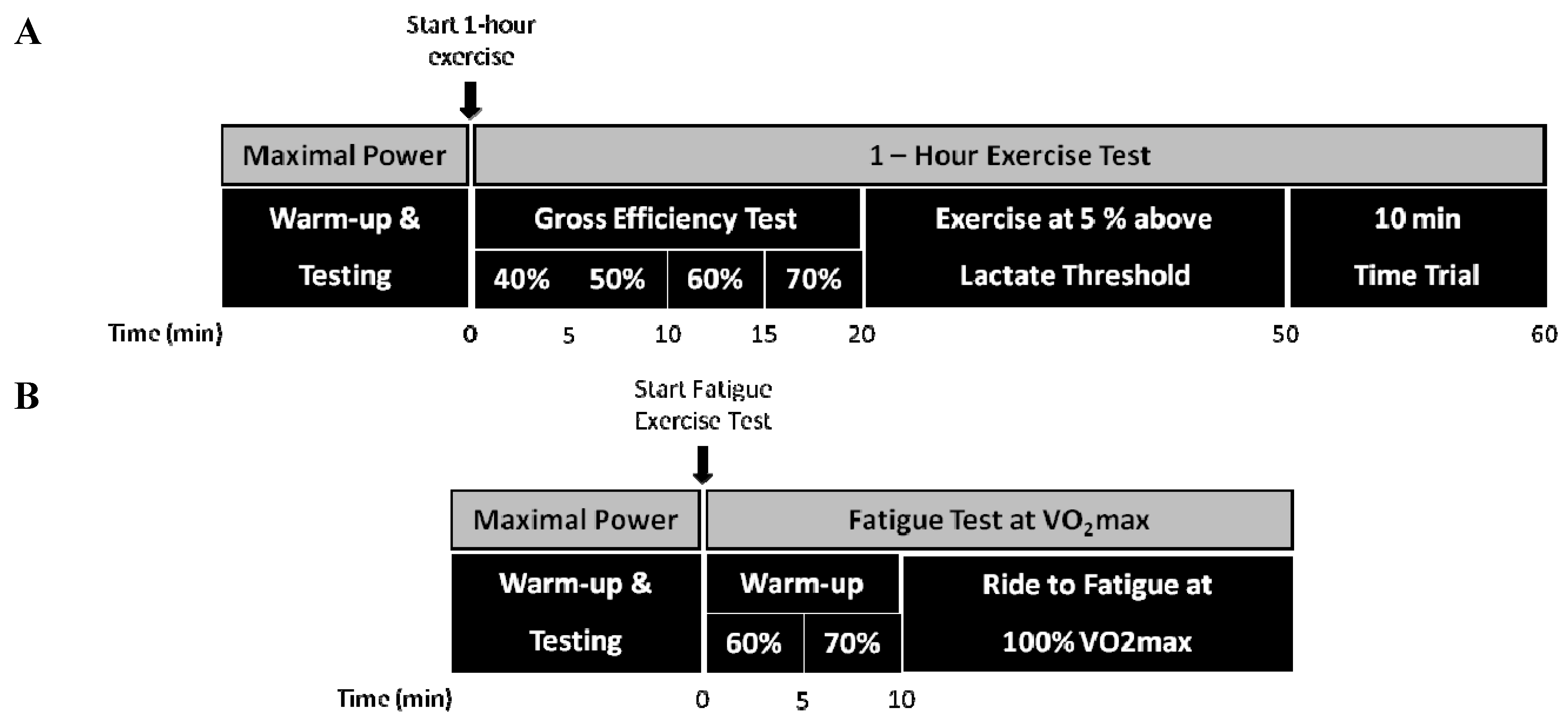
2.2. Experimental Design

2.3. Supplementation
2.4. Respiratory Measurements
2.5. Cardiovascular Measurements
2.6. Body Temperatures and Rating of Perceived Exertion
2.7. Maximal Neuromuscular Power
2.8. Statistics
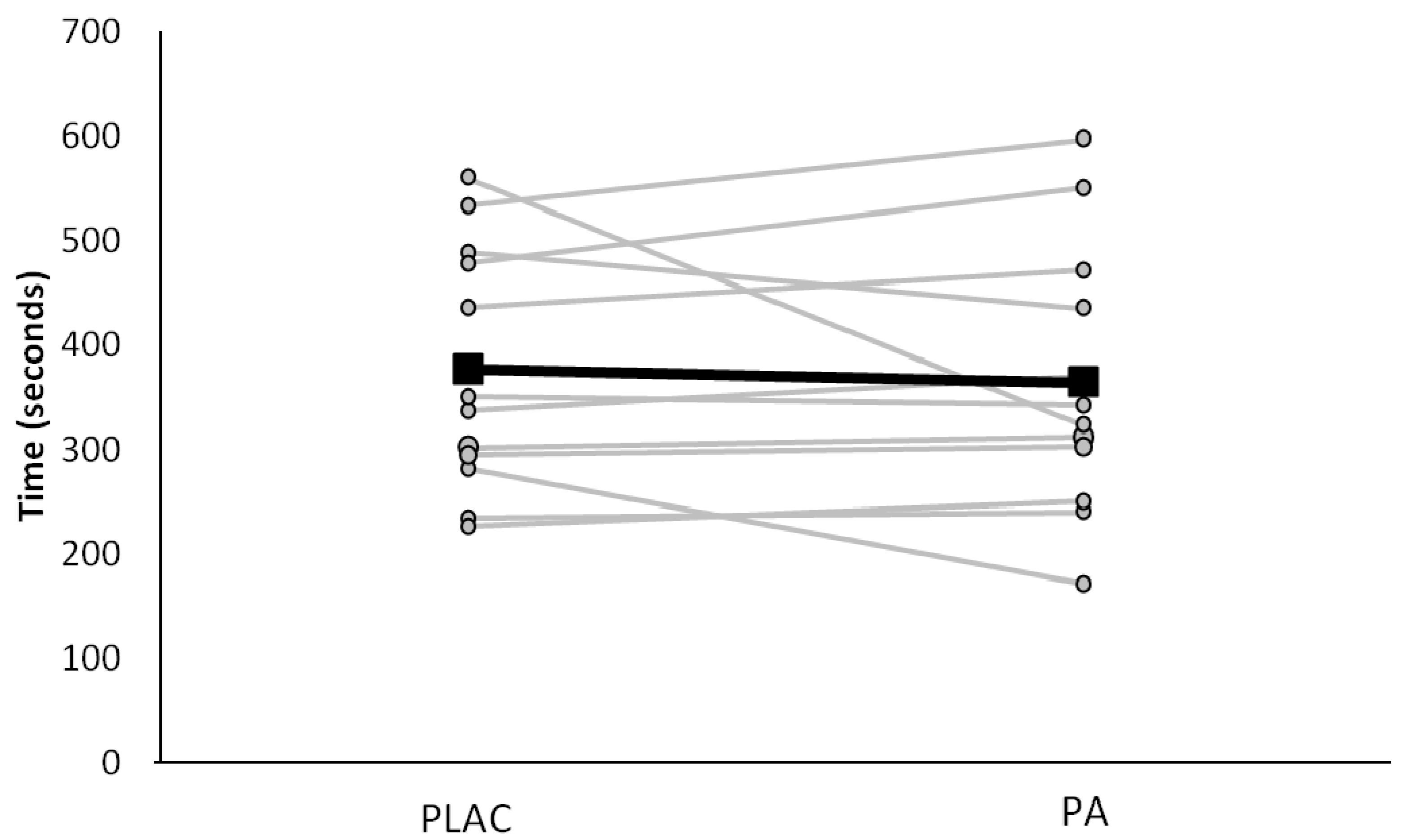
3. Results
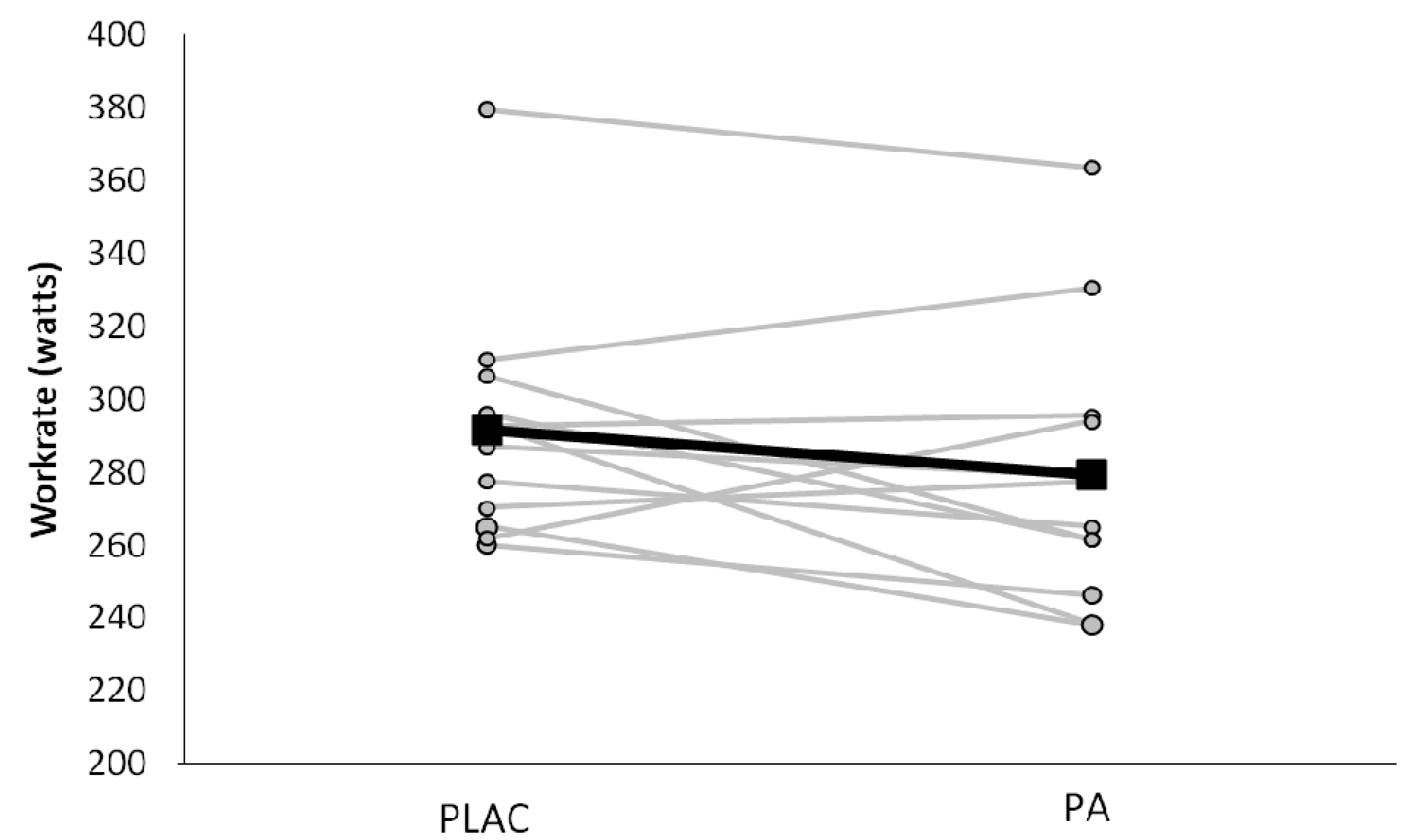
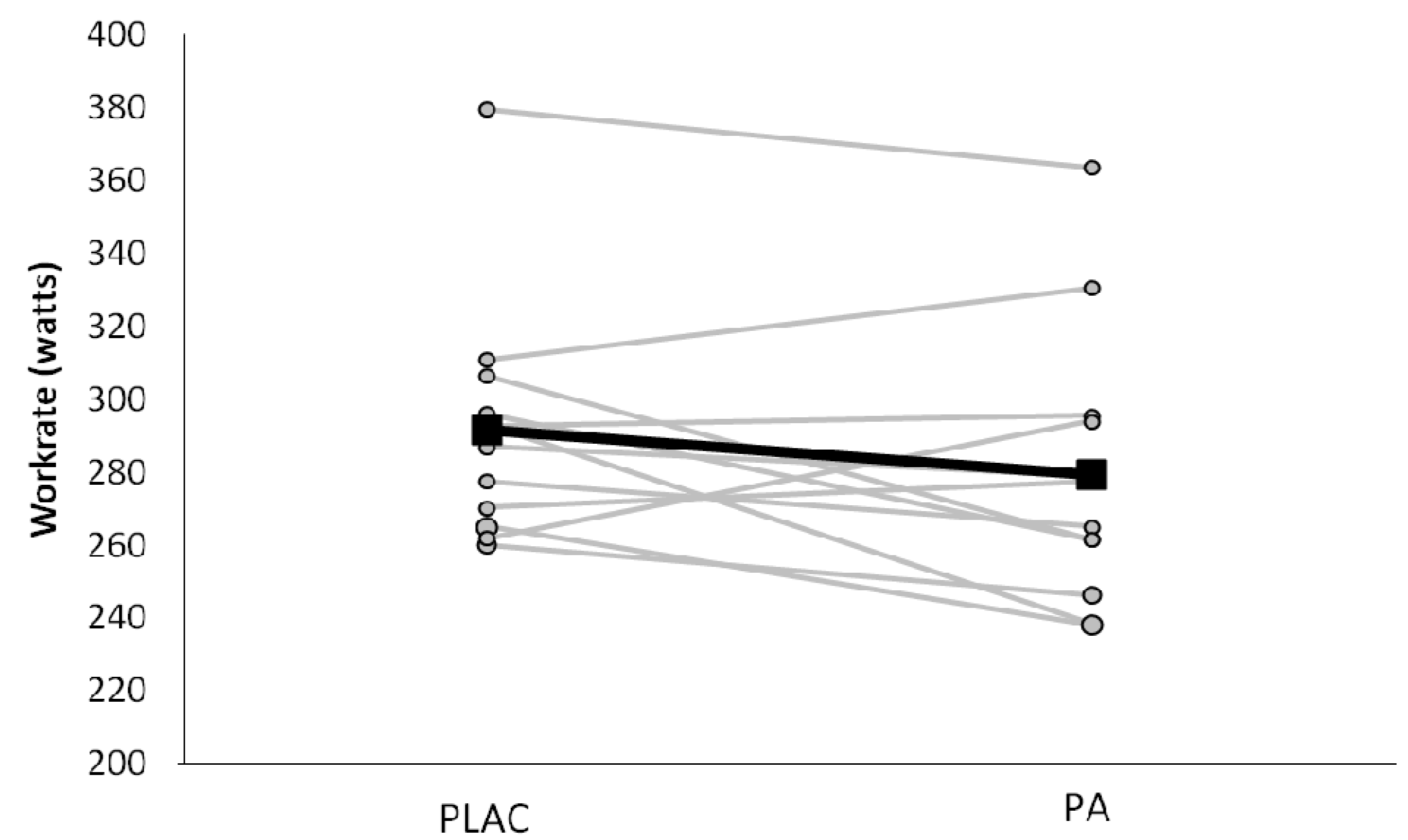
3.1. Subject Characteristics and Environmental Factors
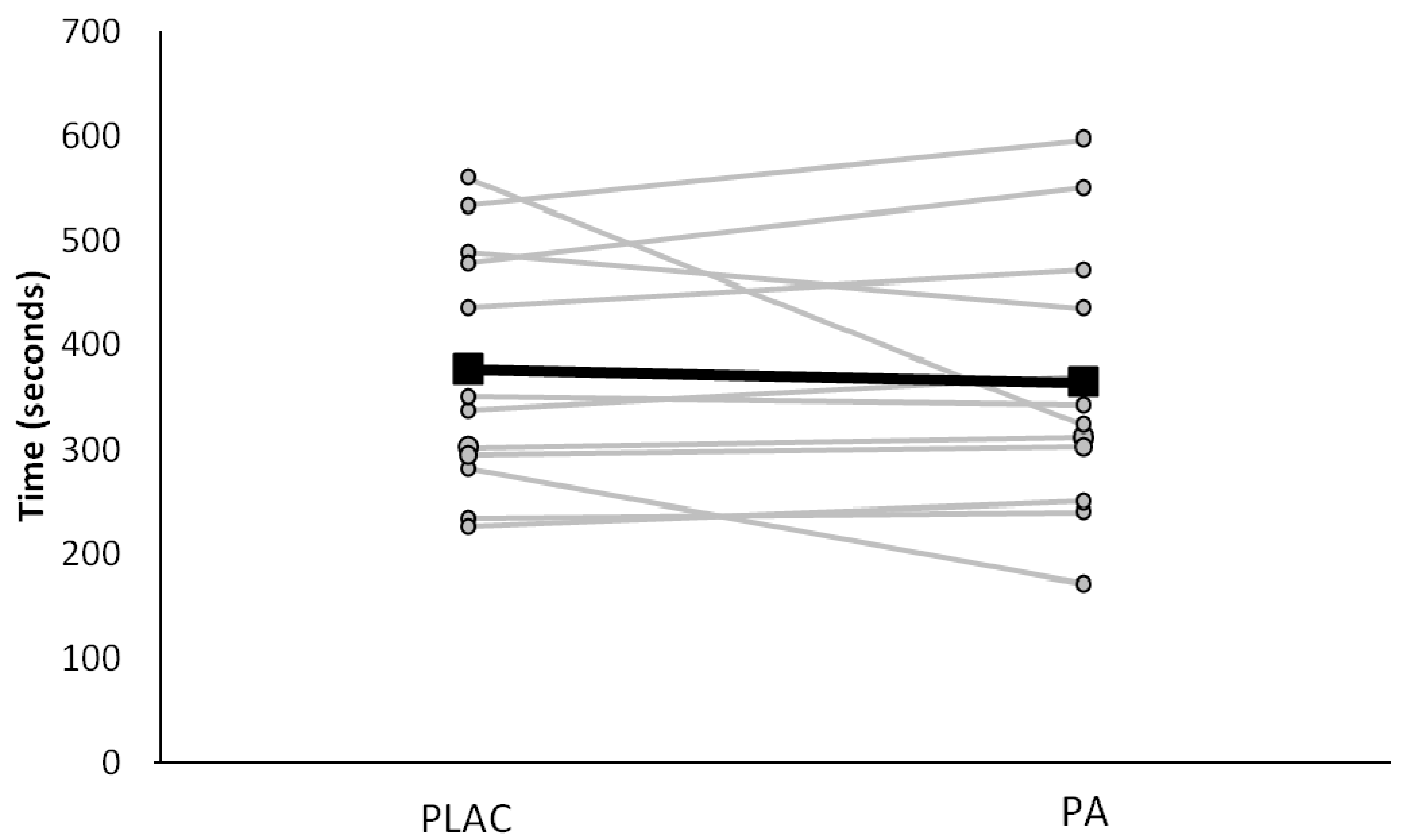
3.2. Exercise Intensity and Respiratory Responses

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time, min | 5 | 10 | 15 | 20 |
|---|---|---|---|---|
| VO2, mL/min | ||||
| PLAC | 1807 ± 147 | 2347 ± 240 * | 2760 ± 218 * | 3271 ± 224 * |
| PA | 1882 ± 171 | 2354 ± 192 * | 2839 ± 235 * | 3324 ± 284 * |
| RER | ||||
| PLAC | 0.84 ± 0.03 | 0.87 ± 0.03 * | 0.87 ± 0.03 | 0.88 ± 0.04 * |
| PA | 0.84 ± 0.03 | 0.86 ± 0.03 * | 0.87 ± 0.03 * | 0.89 ± 0.04 * |
| GE, % | ||||
| PLAC | 16.2 ± 1.5 | 18.1 ± 1.4 * | 20.1 ± 1.0 * | 20.9 ± 1.2 * |
| PA | 15.6 ± 1.5 | 18.1 ± 1.3 * | 19.6 ± 1.1 * | 20.6 ± 0.9 * |
| Parameters | Placebo (Day 1) | PA (Day 1) | Placebo (Day 2) | PA (Day 2) |
|---|---|---|---|---|
| Pmax, watts | 1250 ± 231 | 1240 ± 229 | 1240 ± 214 | 1243 ± 258 |
| IP, watts | 2027 ± 384 | 2029 ± 362 | 2027 ± 336 | 2046 ± 415 |
| RPM | 121 ± 9 | 121 ± 9 | 120 ± 9 | 117 ± 8 |
| Pmax, W/kg | 16.8 ± 1.8 | 16.6 ± 1.9 | 16.7 ± 1.6 | 16.6 ± 2.0 |
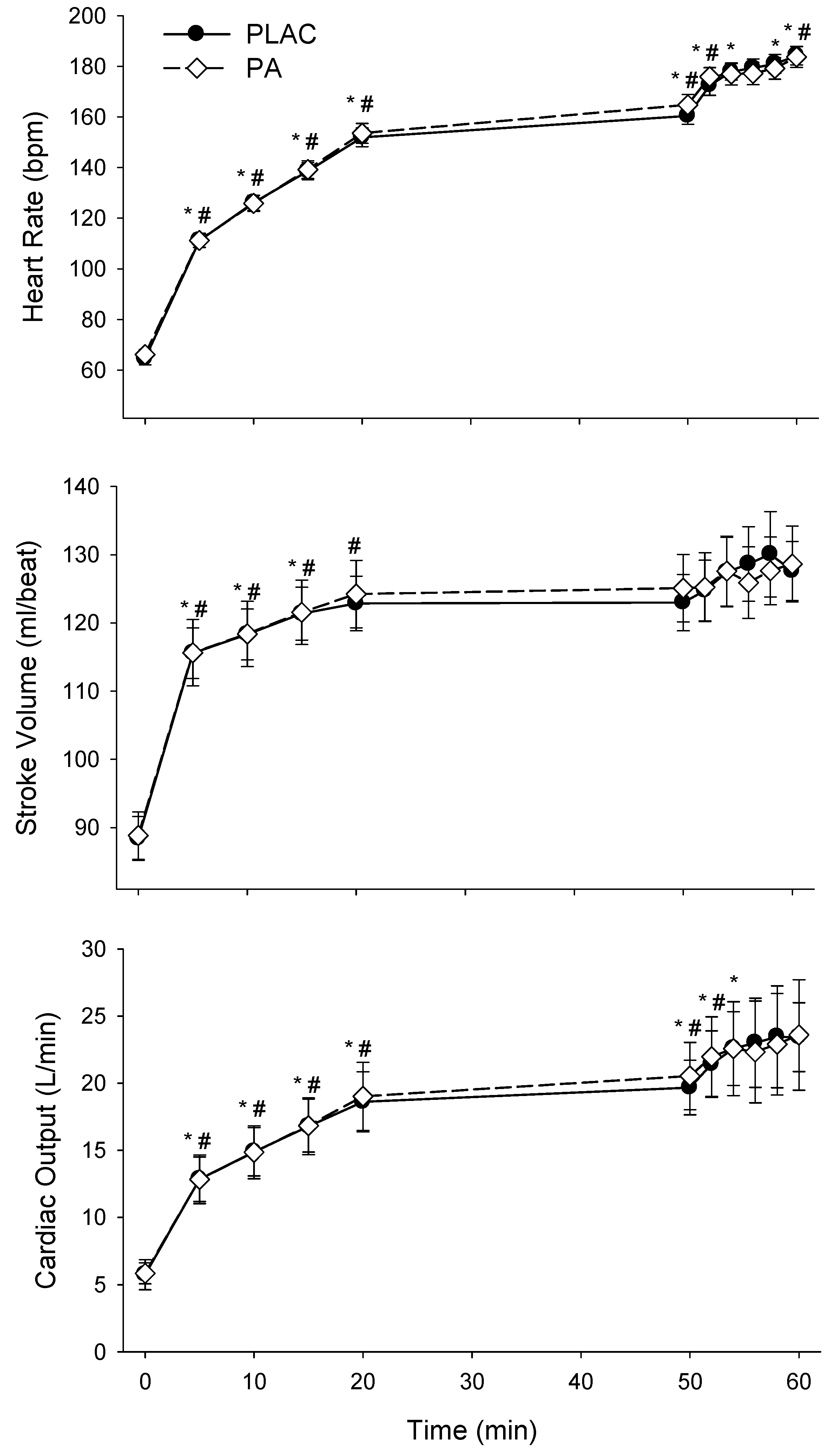
3.3. Cardiovascular Responses
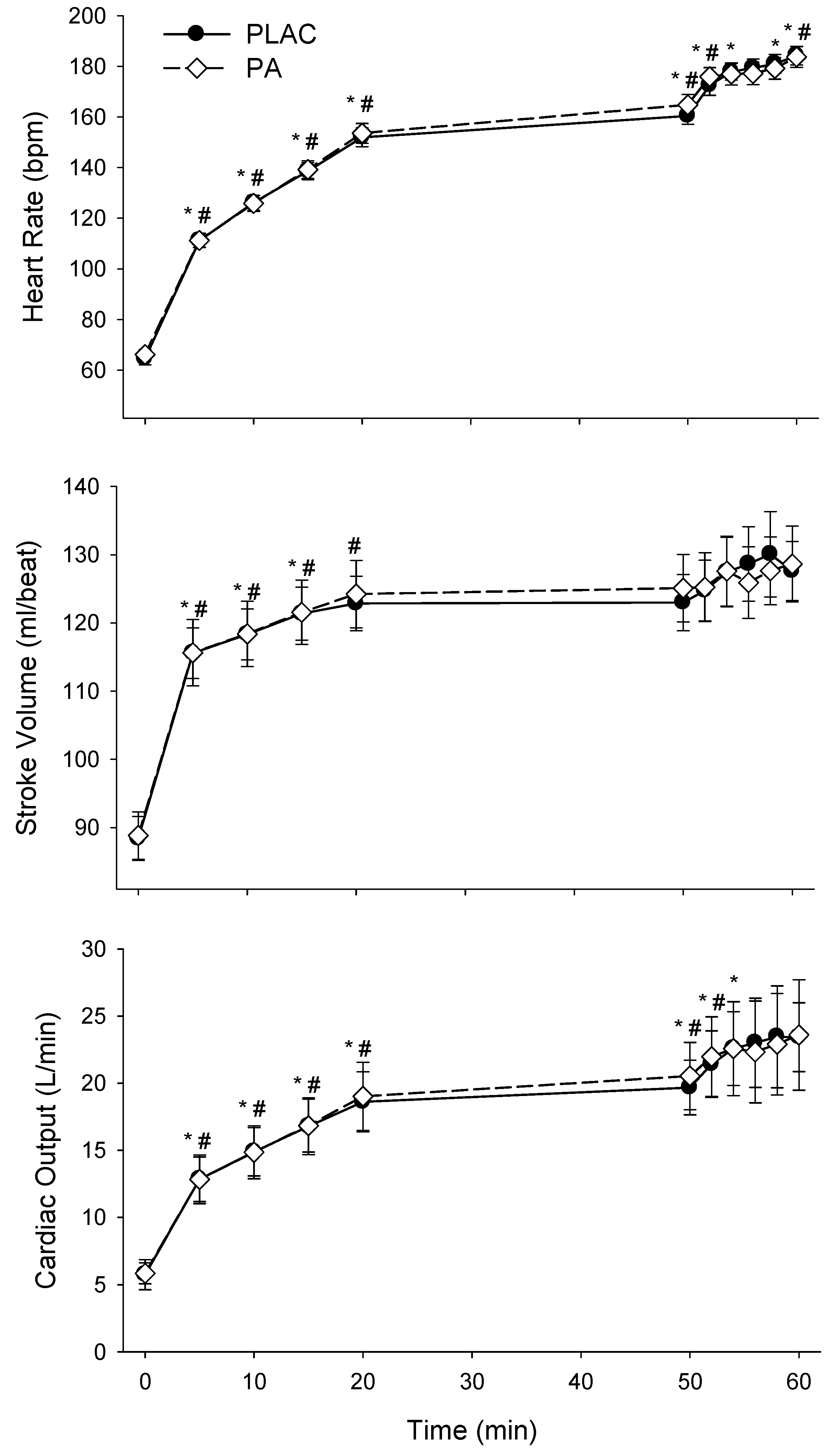
| Time, min | rest | 5 | 10 | 15 | 20 | 50 | 52 | 54 | 56 | 58 | 60 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| MAP, mmHg | |||||||||||
| PLAC | 96 ± 5 | 108 ± 11 | 110 ± 10 | 110 ± 11 | 112 ± 10 | 114 ± 8 | 120 ± 7 | 124 ± 9 | 117 ± 10 | 122 ± 11 | 117 ± 10 |
| PA | 98 ± 6 | 106 ± 11 | 110 ± 11 | 110 ± 10 | 113 ± 9 | 116 ± 12 | 114 ± 10 | 118 ± 11 | 119 ± 10 | 116 ± 12 | 120 ± 11 |
| Tcore, °C | |||||||||||
| PLAC | 37.3 ± 0.3 | 37.4 ± 0.3 * | 37.5 ± 0.3 * | 37.7 ± 0.3 * | 37.8 ± 0.3 * | 38.5 ± 0.3 * | 38.5 ± 0.3 * | 38.6 ± 0.3 * | 38.7 ± 0.3 * | 38.8 ± 0.3 * | 38.9 ± 0.3 * |
| PA | 37.3 ± 0.2 | 37.3 ± 0.2 | 37.4 ± 0.2 * | 37.6 ± 0.2 * | 37.8 ± 0.2 * | 38.5 ± 0.3 * | 38.6 ± 0.3 * | 38.7 ± 0.3 * | 38.7 ± 0.3 * | 38.8 ± 0.3 * | 38.9 ± 0.3 * |
| Tskin, °C | |||||||||||
| PLAC | 32.0 ± 0.9 | 32.2 ± 0.5 | 32.2 ± 0.5 | 32.3 ± 0.5 | 32.1 ± 0.7 | 31.0 ± 0.8 * | 31.1 ± 0.8 | 31.1 ± 0.7 | 30.9 ± 0.7 | 31.0 ± 0.7 | 31.1 ± 1.0 |
| PA | 32.6 ± 0.7 † | 32.4 ± 0.6 | 32.2 ± 0.5 | 32.0 ± 0.4 | 32.1 ± 0.4 | 31.2 ± 1.0 * | 31.1 ± 1.1 | 31.0 ± 1.1 | 30.9 ± 0.9 | 30.8 ± 1.0 | 30.6 ± 1.2 |
| SkBF, L/min | |||||||||||
| PLAC | 1.3 ± 0.2 | 1.7 ± 0.3 * | 2.0 ± 0.4 * | 2.2 ± 0.4 * | 1.7 ± 0.3 * | ||||||
| PA | 1.5 ± 0.2 | 1.7 ± 0.2 * | 2.0 ± 0.2 * | 2.2 ± 0.3 * | 1.8 ± 0.3 * | ||||||
| Time (% of completion) | 0% | 20% | 40% | 60% | 80% | 100% |
|---|---|---|---|---|---|---|
| HR, bpm | ||||||
| PLA | 154 ± 15 | 166 ± 13 * | 173 ± 13 * | 176 ± 13 * | 179 ± 12 * | 181 ± 12 * |
| POM | 156 ± 14 | 166 ± 14 * | 173 ± 13 * | 176 ± 13 * | 179 ± 12 * | 181 ± 13 |
| SV, mL/beat | ||||||
| PLA | 127 ± 13 | 129 ± 13 | 130 ± 16 | 130 ± 15 | 130 ± 15 | 130 ± 16 |
| POM | 130 ± 13 | 132 ± 17 | 133 ± 17 | 133 ± 15 | 133 ± 15 | 133 ± 17 |
| CO, L/min | ||||||
| PLA | 19.6 ± 1.9 | 21.4 ± 2.0 * | 22.4 ± 2.4 * | 22.9 ± 2.2 | 23.2 ± 2.6 | 23.5 ± 2.9 |
| POM | 20.1 ± 1.9 | 21.8 ± 2.7 * | 22.9 ± 2.7 * | 23.4 ± 2.6 * | 23.7 ± 2.6 | 24.0 ± 3.1 |
| Tcore, °C | ||||||
| PLA | 37.7 ± 0.2 | 37.7 ± 0.2 * | 37.8 ± 0.2 * | 38.0 ± 0.2 * | 38.1 ± 0.3 * | 38.2 ± 0.3 * |
| POM | 37.7 ± 0.2 | 37.8 ± 0.2 | 37.9 ± 0.2 * | 38.0 ± 0.3 * | 38.2 ± 0.3 * | 38.3 ± 0.4 * |
| Tskin, °C | ||||||
| PLA | 32.3 ± 0.4 | 32.3 ± 0.4 | 32.1 ± 0.41 * | 31.9 ± 0.42 * | 31.9 ± 0.5 | 32.0 ± 0.4 |
| POM | 32.1 ± 0.7 | 32.2 ± 0.7 | 31.9 ± 0.7 | 31.9 ± 0.7 | 31.9 ± 0.7 | 31.7 ± 0.7 |
3.4. Thermoregulatory Responses
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Davies, K.; Quintanilha, A.; Brooks, G.; Packer, L. Free radicals and tissue damage produced by exercise. Biochem. Biophys. Res. Commun. 1982, 107, 1198–1205. [Google Scholar] [CrossRef]
- Powers, S.K.; Jackson, M.J. Exercise-Induced Oxidative Stress: Cellular Mechanisms and Impact on Muscle Force Production. Physiol. Rev. 2008, 88, 1243–1276. [Google Scholar] [CrossRef]
- Bailey, D.M.; Young, I.S.; McEneny, J.; Lawrenson, L.; Kim, J.; Barden, J.; Richardson, R.S. Regulation of free radical outflow from an isolated muscle bed in exercising humans. Am. J. Physiol. Heart Circ. Physiol. 2004, 287, H1689–H1699. [Google Scholar] [CrossRef]
- Flanagan, S.; Moseley, P.; Buettner, G. Increased flux of free radicals in cells subjected to hyperthermia: Detection by electron paramagnetic resonance spin trapping. FEBS Lett. 1998, 431, 285–286. [Google Scholar] [CrossRef]
- McAnulty, S.; McAnulty, L.; Pascoe, D.; Gropper, S.; Keith, R.; Morrow, J.; Gladden, L. Hyperthermia increases exercise-induced oxidative stress. Int. J. Sports Med. 2005, 26, 188–192. [Google Scholar] [CrossRef]
- Davis, J.M.; Murphy, E.A.; Carmichael, M.D.; Davis, B. Quercetin increases brain and muscle mitochondrial biogenesis and exercise tolerance. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2009, 296, R1071–R1077. [Google Scholar] [CrossRef]
- MacRae, H.; Mefferd, K. Dietary antioxidant supplementation combined with quercetin improves cycling time trial performance. Int. J. Sport Nutr. Exerc. Metab. 2006, 16, 405–419. [Google Scholar]
- McKenna, M.J.; Medved, I.; Goodman, C.A.; Brown, M.J.; Bjorksten, A.R.; Murphy, K.T.; Petersen, A.C.; Sostaric, S.; Gong, X. N-Acetylcysteine attenuates the decline in muscle Na+, K+-pump activity and delays fatigue during prolonged exercise in humans. J. Physiol. 2006, 576, 279–288. [Google Scholar] [CrossRef]
- McKenna, M.J.; Bangsbo, J.; Renaud, J.-M. Muscle K+, Na+, and Cl− disturbances and Na+-K+ pump inactivation: Implications for fatigue. J. Appl. Physiol. 2008, 104, 288–295. [Google Scholar] [CrossRef]
- Medved, I.; Brown, M.J.; Bjorksten, A.R.; Murphy, K.T.; Petersen, A.C.; Sostaric, S.; Gong, X.; McKenna, M.J. N-Acetylcysteine enhances muscle cysteine and glutathione availability and attenuates fatigue during prolonged exercise in endurance-trained individuals. J. Appl. Physiol. 2004, 97, 1477–1485. [Google Scholar] [CrossRef]
- Medved, I.; Brown, M.J.; Bjorksten, A.R.; McKenna, M.J. Effects of intravenous N-acetylcysteine infusion on time to fatigue and potassium regulation during prolonged cycling exercise. J. Appl. Physiol. 2004, 96, 211–217. [Google Scholar]
- McKenna, M.J.; Hargreaves, M. Resolving fatigue mechanisms determining exercise performance: Integrative physiology at its finest! J. Appl. Physiol. 2008, 104, 286–287. [Google Scholar] [CrossRef]
- Cheuvront, S.N.; Ely, B.R.; Kenefick, R.W.; Michniak-Kohn, B.B.; Rood, J.C.; Sawka, M.N. No effect of nutritional adenosine receptor antagonists on exercise performance in the heat. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2009, 296, R394–R401. [Google Scholar]
- Nieman, D.; Henson, D.; Maxwell, K.; Williams, A.; McAnulty, S.; Jin, F.; Shanely, R.; Lines, T. Effects of quercetin and EGCG on mitochondrial biogenesis and immunity. Med. Sci. Sports Exerc. 2009, 41, 1467–1475. [Google Scholar] [CrossRef]
- Utter, A.; Nieman, D.; Kang, J.; Dumke, C.; Quindry, J.; McAnulty, S.; McAnulty, L. Quercetin does not affect rating of perceived exertion in athletes during the Western States endurance run. Res. Sports Med. 2009, 17, 71–83. [Google Scholar] [CrossRef]
- Gomez-Cabrera, M.-C.; Domenech, E.; Romagnoli, M.; Arduini, A.; Borras, C.; Pallardo, F.V.; Sastre, J.; Vina, J. Oral administration of vitamin C decreases muscle mitochondrial biogenesis and hampers training-induced adaptations in endurance performance. Am. J. Clin. Nutr. 2008, 87, 142–149. [Google Scholar]
- Medved, I.; Brown, M.J.; Bjorksten, A.R.; Leppik, J.A.; Sostaric, S.; McKenna, M.J. N-Acetylcysteine infusion alters blood redox status but not time to fatigue during intense exercise in humans. J. Appl. Physiol. 2003, 94, 1572–1582. [Google Scholar]
- Traveline, J.M.; Sudarshan, S.; Roy, B.G.; Cordova, F.; Leyenson, V.; Criner, G.J. Effect of N-Acetylcysteine on human diaphragm strength and fatigability. Am. J. Respir. Crit. Care Med. 1997, 156, 1567–1571. [Google Scholar] [CrossRef]
- Matuszczak, Y.; Farid, M.; Jones, J.; Lansdowne, S.; Smith, M.; Taylor, A.; Reid, M. Effects of N-acetylcysteine on glutathione oxidation and fatigue during handgrip exercise. Muscle Nerve 2005, 32, 633–638. [Google Scholar] [CrossRef]
- Hernandez, A.; Cheng, A.; Westerblad, H. Antioxidants and Skeletal Muscle Performance: “Common Knowledge” vs. Experimental Evidence. Front Physiol. 2012, 3. [Google Scholar] [CrossRef]
- Seeram, N.P.; Adams, L.S.; Henning, S.M.; Niu, Y.; Zhang, Y.; Nair, M.G.; Heber, D. In vitro antiproliferative, apoptotic and antioxidant activities of punicalagin, ellagic acid and a total pomegranate tannin extract are enhanced in combination with other polyphenols as found in pomegranate juice. J. Nutr. Biochem. 2005, 16, 360–367. [Google Scholar] [CrossRef]
- Scalbert, A.; Johnson, I.T.; Saltmarsh, M. Polyphenols: Antioxidants and beyond. Am. J. Clin. Nutr. 2005, 81, 215S–217S. [Google Scholar]
- Vita, J.A. Polyphenols and cardiovascular disease: Effects on endothelial and platelet function. Am. J. Clin. Nutr. 2005, 81, 292S–297S. [Google Scholar]
- Rosenblat, M.; Hayek, T.; Aviram, M. Anti-oxidative effects of pomegranate juice (PJ) consumption by diabetic patients on serum and on macrophages. Atherosclerosis 2006, 187, 363–371. [Google Scholar] [CrossRef]
- Aviram, M.; Rosenblat, M.; Gaitini, D.; Nitecki, S.; Hoffman, A.; Dornfeld, L.; Volkova, N.; Presser, D.; Attias, J.; Liker, H.; et al. Pomegranate juice consumption for 3 years by patients with carotid artery stenosis reduces common carotid intima-media thickness, blood pressure and LDL oxidation. Clin. Nutr. 2004, 23, 423–433. [Google Scholar] [CrossRef]
- Adams, L.; Seeram, N.; Aggarwal, B.; Takada, Y.; Sand, D.; Heber, D. Pomegranate juice, total pomegranate ellagitannins, and punicalagin suppress inflammatory cell signaling in colon cancer cells. J. Agric. Food Chem. 2006, 54, 980–985. [Google Scholar] [CrossRef]
- Shukla, M.; Gupta, K.; Rasheed, Z.; Khan, K.; Haqqi, T. Consumption of hydrolyzable tannins-rich pomegranate extract suppresses inflammation and joint damage in rheumatoid arthritis. Nutrition 2008, 24, 733–743. [Google Scholar] [CrossRef]
- Seeram, N.; Aviram, M.; Zhang, Y.; Henning, S.; Feng, L.; Dreher, M.; Heber, D. Comparison of antioxidant potency of commonly consumed polyphenol-rich beverages in the United States. J. Agric. Food Chem. 2008, 56, 1415–1422. [Google Scholar] [CrossRef]
- Seeram, N.; Zhang, Y.; McKeever, R.; Henning, S.; Lee, R.; Suchard, M.; Li, Z.; Chen, S.; Thames, G.; Zerlin, A.; et al. Pomegranate juice and extracts provide similar levels of plasma and urinary ellagitannin metabolites in human subjects. J. Med. Food 2008, 11, 390–394. [Google Scholar] [CrossRef]
- Seeram, N.P.; Henning, S.M.; Zhang, Y.; Suchard, M.; Li, Z.; Heber, D. Pomegranate Juice Ellagitannin Metabolites Are Present in Human Plasma and Some Persist in Urine for Up to 48 h. J. Nutr. 2006, 136, 2481–2485. [Google Scholar]
- Ignarro, L.; Byrns, R.; Sumi, D.; de Nigris, F.; Napoli, C. Pomegranate juice protects nitric oxide against oxidative destruction and enhances the biological actions of nitric oxide. Nitric Oxide 2006, 15, 93–102. [Google Scholar] [CrossRef]
- Kellogg, D.L., Jr.; Crandall, C.G.; Liu, Y.; Charkoudian, N.; Johnson, J.M. Nitric oxide and cutaneous active vasodilation during heat stress in humans. J. Appl. Physiol. 1998, 85, 824–829. [Google Scholar]
- Welch, G.; Foote, K.M.; Hansen, C.; Mack, G.W. Nonselective NOS inhibition blunts the sweat response to exercise in a warm environment. J. Appl. Physiol. 2009, 106, 796–803. [Google Scholar] [CrossRef]
- Boushel, R.; Langberg, H.; Gemmer, C.; Olesen, J.; Crameri, R.; Scheede, C.; Sander, M.; Kjaer, M. Combined inhibition of nitric oxide and prostaglandins reduces human skeletal muscle blood flow during exercise. J. Physiol. 2002, 543, 691–698. [Google Scholar] [CrossRef]
- Mortensen, S.P.; Gonzalez-Alonso, J.; Damsgaard, R.; Saltin, B.; Hellsten, Y. Inhibition of nitric oxide and prostaglandins, but not endothelial-derived hyperpolarizing factors, reduces blood flow and aerobic energy turnover in the exercising human leg. J. Physiol. 2007, 581, 853–861. [Google Scholar] [CrossRef]
- Bailey, S.J.; Fulford, J.; Vanhatalo, A.; Winyard, P.G.; Blackwell, J.R.; DiMenna, F.J.; Wilkerson, D.P.; Benjamin, N.; Jones, A.M. Dietary nitrate supplementation enhances muscle contractile efficiency during knee-extensor exercise in humans. J. Appl. Physiol. 2010, 109, 135–148. [Google Scholar] [CrossRef]
- Bailey, S.J.; Winyard, P.; Vanhatalo, A.; Blackwell, J.R.; DiMenna, F.J.; Wilkerson, D.P.; Tarr, J.; Benjamin, N.; Jones, A.M. Dietary nitrate supplementation reduces the O2 cost of low-intensity exercise and enhances tolerance to high-intensity exercise in humans. J. Appl. Physiol. 2009, 107, 1144–1155. [Google Scholar] [CrossRef]
- Jones, A.M.; Bailey, S.J.; Vanhatalo, A. Dietary Nitrate and O2 Consumption during Exercise. Med. Sport Sci. 2013, 59, 29–35. [Google Scholar]
- Larsen, F.J.; Schiffer, T.A.; Borniquel, S.; Sahlin, K.; Ekblom, B.; Lundberg, J.O.; Weitzberg, E. Dietary inorganic nitrate improves mitochondrial efficiency in humans. Cell Metab. 2011, 13, 149–159. [Google Scholar] [CrossRef]
- Martin, J.; Wagner, B.; Coyle, E. Inertial-load method determines maximal cycling power in a single exercise bout. Med. Sci. Sports Exerc. 1997, 29, 1505–1512. [Google Scholar] [CrossRef]
- Coyle, E.; Sidossis, L.; Horowitz, J.; Beltz, J. Cycling efficiency is related to the percentage of type I muscle fibers. Med. Sci. Sports Exerc. 1992, 24, 782–788. [Google Scholar]
- Below, P.; Mora-Rodriguez, R.; Gonzalez-Alonso, J.; Coyle, E. Fluid and carbohydrate ingestion independently improve performance during 1 h of intense exercise. Med. Sci. Sports Exerc. 1995, 27, 200–210. [Google Scholar]
- Trinity, J.D.; Lee, J.F.; Pahnke, M.D.; Beck, K.C.; Coyle, E.F. Attenuated relationship between cardiac output and oxygen uptake during high-intensity exercise. Acta Physiol. 2012, 204, 362–370. [Google Scholar] [CrossRef]
- Dingwell, J.; Joubert, J.; Diefenthaeler, F.; Trinity, J. Changes in muscle activity and kinematics of highly trained cyclists during fatigue. IEEE Trans. Biomed. Eng. 2008, 55, 2666–2674. [Google Scholar] [CrossRef]
- Charloux, A.; Lonsdorfer-Wolf, E.; Richard, R.; Lampert, E.; Oswald-Mammosser, M.; Mettauer, B.; Geny, B.; Lonsdorfer, J. A new impedance cardiograph device for the non-invasive evaluation of cardiac output at rest and during exercise: Comparison with the “direct” Fick method. Eur. J. Appl. Physiol. 2000, 82, 313–320. [Google Scholar] [CrossRef]
- Richard, R.; Lonsdorfer-Wolf, E.; Charloux, A.; Doutreleau, S.; Buchheit, M.; Oswald-Mammosser, M.; Lampert, E.; Mettauer, B.; Geny, B.; Lonsdorfer, J. Non-invasive cardiac output evaluation during a maximal progressive exercise test, using a new impedance cardiograph device. Eur. J. Appl. Physiol. 2001, 85, 202–207. [Google Scholar] [CrossRef]
- Hardy, J.D.; Du Bois, E.F.; Soderstrom, G.F. The Technic of Measuring Radiation and Convection: One Figure. J. Nutr. 1938, 15, 461–475. [Google Scholar]
- Borg, G. Simple Rating Methods for Estimation of Perceived Exertion. In Physical Work and Effort; Borg, G., Ed.; Pergamon: New York, NY, USA, 1975; pp. 39–46. [Google Scholar]
- Sawka, M.N.; Wenger, C. Physiological Responses to Acute Exercise-heat Stress. In Human Performance Physiology and Environmental Medicine at Terrestrial Extremes; Pandolf, K.B., Sawka, M.N., Gonzalez, R.R., Eds.; Cooper Publishing: Traverse City, MI, USA, 1988; pp. 97–151. [Google Scholar]
- Trinity, J.; Pahnke, M.; Reese, E.; Coyle, E. Maximal mechanical power during a taper in elite swimmers. Med. Sci. Sports Exerc. 2006, 38, 1643–1649. [Google Scholar] [CrossRef]
- Trinity, J.; Pahnke, M.; Sterkel, J.; Coyle, E. Maximal power and performance during a swim taper. Int. J. Sports Med. 2008, 29, 500–506. [Google Scholar] [CrossRef]
- Lescaudron, L.; Peltekian, E.; Fontaine-Perus, J.; Paulin, D.; Zampieri, M.; Garcia, L.; Parrish, E. Blood borne macrophages are essential for the triggering of muscle regeneration following muscle transplant. Neuromuscul. Disord. 1999, 9, 72–80. [Google Scholar] [CrossRef]
- Richardson, R.S.; Donato, A.J.; Uberoi, A.; Wray, D.W.; Lawrenson, L.; Nishiyama, S.; Bailey, D.M. Exercise-induced brachial artery vasodilation: Role of free radicals. Am. J. Physiol. Heart Circ. Physiol. 2007, 292, H1516–H1522. [Google Scholar]
- Wray, D.; Uberoi, A.; Lawrenson, L.; Bailey, D.; Richardson, R. Oral antioxidants and cardiovascular health in the exercise-trained and untrained elderly: A radically different outcome. Clin. Sci. 2009, 116, 433–441. [Google Scholar] [CrossRef]
- Trombold, J.R.; Reinfeld, A.S.; Casler, J.R.; Coyle, E.F. The effect of pomegranate juice supplementation on strength and soreness after eccentric exercise. J. Strength Cond. Res. 2011, 25, 1782–1788. [Google Scholar] [CrossRef]
- Trombold, J.R.; Barnes, J.N.; Critchley, L.; Coyle, E.F. Ellagitannin comsumption improves strength recovery 2–3 days following eccentric exercise. Med. Sci. Sports Exerc. 2010, 42, 493–498. [Google Scholar]
- Murphy, K.T.; Medved, I.; Brown, M.J.; Cameron-Smith, D.; McKenna, M.J. Antioxidant treatment with N-acetylcysteine regulates mammalian skeletal muscle Na+-K+-ATPaseα gene expression during repeated contractions. Exp. Physiol. 2008, 93, 1239–1248. [Google Scholar] [CrossRef]
- Tyldum, G.A.; Schjerve, I.E.; Tjonna, A.E.; Kirkeby-Garstad, I.; Stolen, T.O.; Richardson, R.S.; Wisloff, U. Endothelial Dysfunction Induced by Post-Prandial Lipemia: Complete Protection Afforded by High-Intensity Aerobic Interval Exercise. J. Am. Coll. Cardiol. 2009, 53, 200–206. [Google Scholar] [CrossRef]
- Plotnick, G.D.; Corretti, M.C.; Vogel, R.A.; Hesslink, R., Jr.; Wise, J.A. Effect of supplemental phytonutrients on impairment of the flow-mediated brachialartery vasoactivity after a single high-fat meal. J. Am. Coll. Cardiol. 2003, 41, 1744–1749. [Google Scholar] [CrossRef]
- Barringer, T.A.; Hatcher, L.; Sasser, H.C. Potential Benefits on Impairment of Endothelial Function after a High-fat Meal of 4 weeks of Flavonoid Supplementation. Evid. Based Complement. Altern. Med. 2008, 2011. [Google Scholar] [CrossRef]
- Eskurza, I.; Monahan, K.D.; Robinson, J.A.; Seals, D.R. Effect of acute and chronic ascorbic acid on flow-mediated dilatation with sedentary and physically active human ageing. J. Physiol. 2004, 556, 315–324. [Google Scholar] [CrossRef]
- Kirby, B.S.; Voyles, W.F.; Simpson, C.B.; Carlson, R.E.; Schrage, W.G.; Dinenno, F.A. Endothelium-dependent vasodilatation and exercise hyperaemia in ageing humans: Impact of acute ascorbic acid administration. J. Physiol. 2009, 587, 1989–2003. [Google Scholar] [CrossRef]
- Jablonski, K.L.; Seals, D.R.; Eskurza, I.; Monahan, K.D.; Donato, A.J. High-dose ascorbic acid infusion abolishes chronic vasoconstriction and restores resting leg blood flow in healthy older men. J. Appl. Physiol. 2007, 103, 1715–1721. [Google Scholar] [CrossRef]
- Donato, A.J.; Uberoi, A.; Bailey, D.M.; Walter Wray, D.; Richardson, R.S. Exercise-induced brachial artery vasodilation: Effects of antioxidants and exercise training in elderly men. Am. J. Physiol. Heart Circ. Physiol. 2010, 298, H671–H678. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Trinity, J.D.; Pahnke, M.D.; Trombold, J.R.; Coyle, E.F. Impact of Polyphenol Antioxidants on Cycling Performance and Cardiovascular Function. Nutrients 2014, 6, 1273-1292. https://doi.org/10.3390/nu6031273
Trinity JD, Pahnke MD, Trombold JR, Coyle EF. Impact of Polyphenol Antioxidants on Cycling Performance and Cardiovascular Function. Nutrients. 2014; 6(3):1273-1292. https://doi.org/10.3390/nu6031273
Chicago/Turabian StyleTrinity, Joel D., Matthew D. Pahnke, Justin R. Trombold, and Edward F. Coyle. 2014. "Impact of Polyphenol Antioxidants on Cycling Performance and Cardiovascular Function" Nutrients 6, no. 3: 1273-1292. https://doi.org/10.3390/nu6031273