Magnesium and Osteoporosis: Current State of Knowledge and Future Research Directions

Abstract
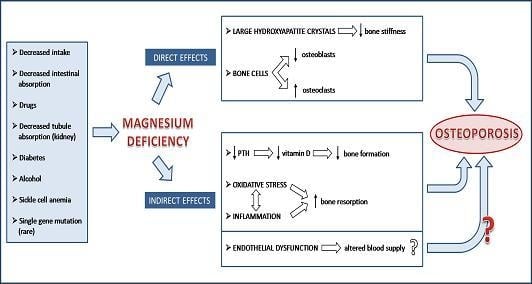
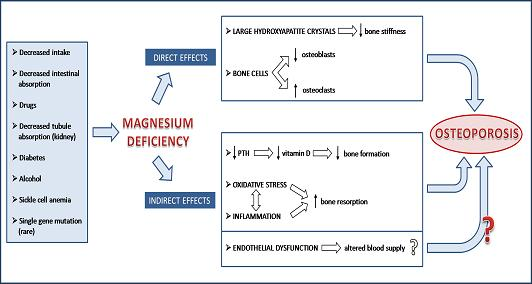
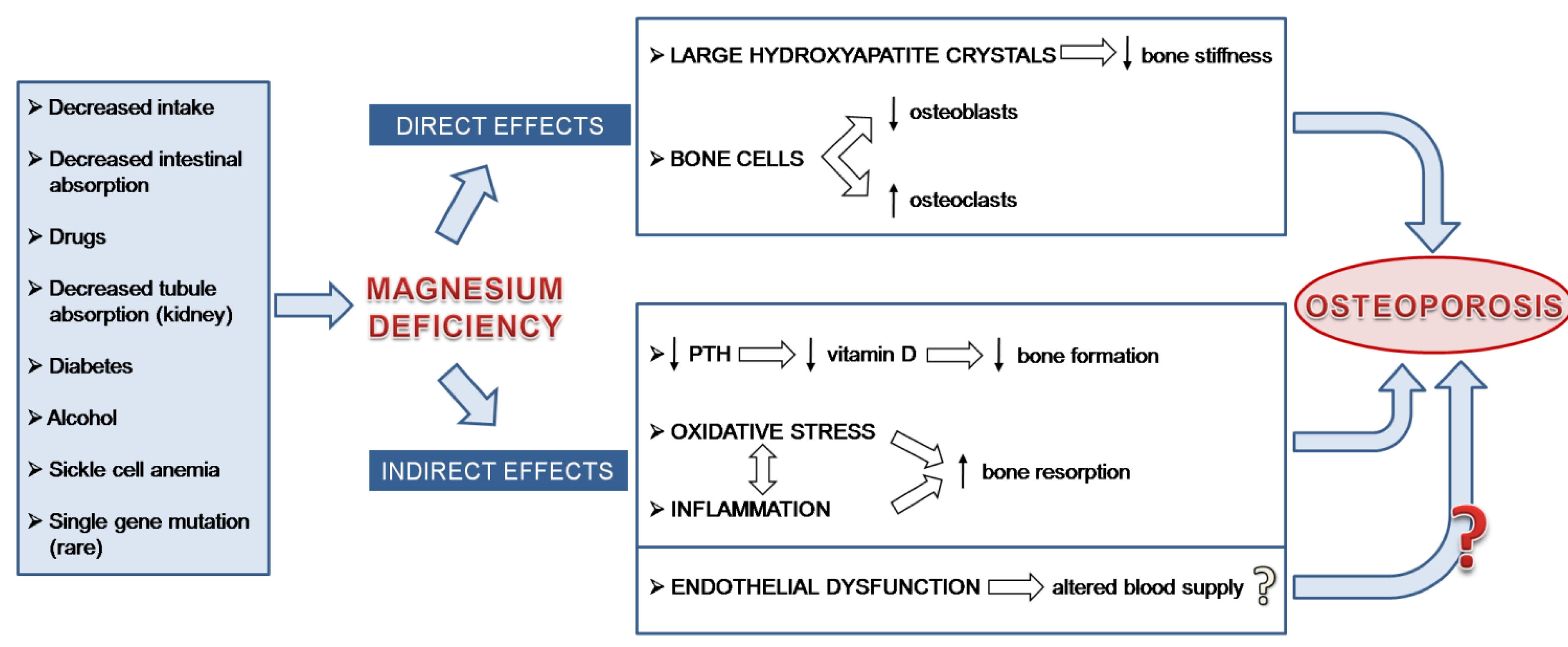
:
1. Introduction
2. Magnesium and the Bone: Molecular, Biochemical and Cellular Insights
3. Low Magnesium and Osteoporosis: Experimental Evidence
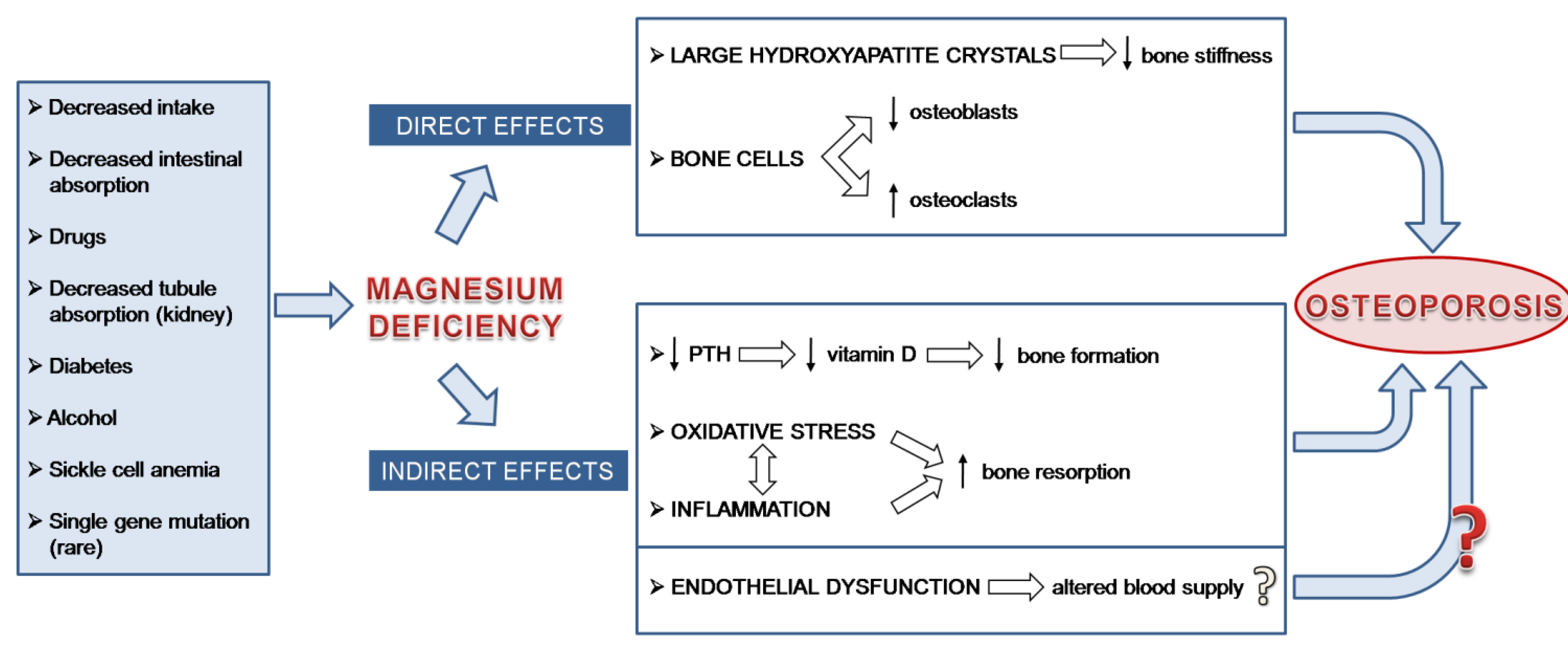
4. Low Magnesium and Osteoporosis: Studies in Humans

{kind=link}
{kind=link}
| Disease | * Omim® Reference | Gene | Protein |
|---|---|---|---|
| recessive familial hypomagnesemia with hypercalciuria and nephrocalcinosis | 248250 248190 | CLDN16 CLDN19 | Claudin-16 and Claudin-19 Function: tight junction proteins Localization: thick ascending limb of Henle’s loop and convolute distal tubule in the kidney Involved in paracellular epithelial transport |
| recessive hypomagnesemia with secondary hypolcalcemia | 602074 | TRPM6 | TRPM6 Function: cation channel and α-kinase Localization: intestine and distal convolute tubule in the kidney Involved in transcellular Mg reabsorption in epithelial cells |
| dominant renal hypomagnesemia | 154020 | FXYD2 | FXYD2 Function: γ-subunit of the Na+/K+ ATPase Localization: basolateral membrane of proximal and distal tubules in the kidney Involved in transcellular Mg reabsorption |
| recessive renal hypomagnesemia | 131530 | EGF | EGF Function: growth factor and magnesiotropic hormone Involved in stimulating magnesium reabsorption in the renal distal convoluted tubule via activation of TRPM6 |
| dominant hypomagnesemia | 607803 | CNNM2 | CNNM2 Function: metal trasporter Localization: ubiquitous; thick ascending limb of Henle’s loop and basolateral membrane of distal tubule in the kidney Involved in renal Mg reabsorption |
| autosomal dominant myokymia with hypomagnesemia | 176260 | KCNA1 | Kv1.1 Function: voltage-gated K+ channel Localization: ubiquitous; distal convolute tubule in the kidney Involved in renal Mg reabsorption |
5. Critical Issues and Future Perspective
6. Conclusions
Conflict of Interest
References
- Rachner, T.D.; Khosla, S.; Hofbauer, L.C. Osteoporosis: Now and the future. Lancet 2011, 377, 1276–1287. [Google Scholar] [CrossRef]
- Nieves, J.W. Osteoporosis: The role of micronutrients. Am. J. Clin. Nutr. 2005, 81, 1232S–1239S. [Google Scholar]
- Ström, O.; Borgström, F.; Kanis, J.A.; Compston, J.; Cooper, C.; McCloskey, E.V.; Jönsson, B. Osteoporosis: Burden, health care provision and opportunities in the EU: A report prepared in collaboration with the International Osteoporosis Foundation (IOF) and the European Federation of Pharmaceutical Industry Associations (EFPIA). Arch.Osteoporos. 2011, 6, 59–155. [Google Scholar] [CrossRef]
- Song, Y.; Li, T.Y.; van Dam, R.M.; Manson, J.E.; Hu, F.B. Magnesium intake and plasma concentrations of markers of systemic inflammation and endothelial dysfunction in women. Am. J. Clin. Nutr. 2007, 85, 1068–1074. [Google Scholar]
- Nieves, J.W. Skeletal effects of nutrients and nutraceuticals, beyond calcium and vitamin D. Osteoporos. Int. 2013, 24, 771–786. [Google Scholar] [CrossRef]
- Jahnen-Dechent, W.; Ketteler, M. Magnesium basics. Clin. Kidney J. 2012, 5, i3–i14. [Google Scholar] [CrossRef]
- Alfrey, A.C.; Miller, N.L. Bone magnesium pools in uremia. J. Clin. Investig. 1973, 52, 3019–3027. [Google Scholar] [CrossRef]
- Cunningham, J.; Rodriguez, M.; Messa, P. Magnesium in chronic kidney disease Stages 3 and 4 and in dialysis patients. Clin. Kidney J. 2012, 5, i39–i51. [Google Scholar] [CrossRef]
- Iseri, L.T.; French, J.H. Magnesium: Nature’s physiologic calcium blocker. Am. Heart J. 1984, 108, 188–193. [Google Scholar] [CrossRef]
- Li, F.Y.; Chaigne-Delalande, B.; Kanellopoulou, C.; Davis, J.C.; Matthews, H.F.; Douek, D.C.; Cohen, J.I.; Uzel, G.; Su, H.C.; Lenardo, M.J. Second messenger role for Mg2+ revealed by human T-cell immunodeficiency. Nature 2011, 475, 471–476. [Google Scholar] [CrossRef]
- Rude, R.K.; Singer, F.R.; Gruber, H.E. Skeletal and hormonal effects of magnesium deficiency. J. Am. Coll.Nutr. 2009, 28, 131–141. [Google Scholar] [CrossRef]
- Boskey, A.L.; Rimnac, C.M.; Bansal, M.; Federman, M.; Lian, J.; Boyan, B.D. Effect of short-term hypomagnesemia on the chemical and mechanical properties of rat bone. J. Orthop. Res. 1992, 10, 774–783. [Google Scholar] [CrossRef]
- Belluci, M.M.; Giro, G.; del Barrio, R.A.; Pereira, R.M.; Marcantonio, E.; Orrico, S.R. Effects of magnesium intake deficiency on bone metabolism and bone tissue around osseointegrated implants. Clin. Oral Implants Res. 2011, 22, 716–721. [Google Scholar] [CrossRef]
- Creedon, A.; Flynn, A.; Cashman, K. The effect of moderately and severely restricted dietary magnesium intakes on bone composition and bone metabolism in the rat. Br. J. Nutr. 1999, 82, 63–71. [Google Scholar]
- Schwartz, R.; Reddi, A.H. Influence of magnesium depletion on matrix-induced endochondral bone formation. Calcif. Tissue Int. 1979, 29, 15–20. [Google Scholar] [CrossRef]
- Rude, R.K.; Gruber, H.E. Magnesium deficiency and osteoporosis: Animal and human observations. J. Nutr. Biochem. 2004, 15, 710–716. [Google Scholar] [CrossRef]
- Rude, R.K.; Kirchen, M.E.; Gruber, H.E.; Meyer, M.H.; Luck, J.S.; Crawford, D.L. Magnesium deficiency-induced osteoporosis in the rat: Uncoupling of bone formation and bone resorption. Magnes. Res. 1999, 12, 257–267. [Google Scholar]
- Rude, R.K.; Gruber, H.E.; Wie, L.Y.; Frausto, A.; Mills, B.G. Magnesium deficiency: Effect on bone and mineral metabolism in the mouse. Calcif. Tissue Int. 2003, 72, 32–41. [Google Scholar] [CrossRef]
- Leidi, M.; Dellera, F.; Mariotti, M.; Banfi, G.; Crapanzano, C.; Albisetti, W.; Maier, J.A. Nitric oxide mediates low magnesium inhibition of osteoblast-like cell proliferation. J. Nutr. Biochem. 2012, 23, 1224–1229. [Google Scholar] [CrossRef]
- Belluci, M.M.; Schoenmaker, T.; Rossa-Junior, C.; Orrico, S.R.; de Vries, T.J.; Everts, V. Magnesium deficiency results in an increased formation of osteoclasts. J. Nutr. Biochem. 2013. [Google Scholar] [CrossRef]
- Pironi, L.; Malucelli, E.; Guidetti, M.; Lanzoni, E.; Farruggia, G.; Pinna, A.D.; Barbiroli, D.; Iotti, S. The complex relationship between magnesium and serum parathyroid hormone: A study in patients with chronic intestinal failure. Magnes. Res. 2009, 22, 37–43. [Google Scholar]
- Gray, R.W.; Omdahl, J.L.; Ghazarian, J.G.; DeLuca, H.F. 25-Hydroxycholecalciferol-1 hydroxylase. Subcellular location and properties. J. Biol. Chem. 1972, 247, 7528–7532. [Google Scholar]
- Mazur, A.; Maier, J.A.; Rock, E.; Gueux, E.; Nowacki, W.; Rayssiguier, Y. Magnesium and the inflammatory response: Potential physiopathological implications. Arch. Biochem. Biophys. 2007, 458, 48–56. [Google Scholar] [CrossRef]
- Baker-LePain, J.C.; Nakamura, M.C.; Lane, N.E. Effects of inflammation on bone: An update. Curr. Opin. Rheumatol. 2011, 23, 389–395. [Google Scholar] [CrossRef]
- Wolf, F.I.; Trapani, V.; Simonacci, M.; Boninsegna, A.; Mazur, A.; Maier, J.A. Magnesium deficiency affects mammary epithelial cell proliferation: Involvement of oxidative stress. Nutr. Cancer 2009, 61, 131–136. [Google Scholar] [CrossRef]
- Garrett, I.R.; Boyce, B.F.; Oreffo, R.O.; Bonewald, L.; Poser, J.; Mundy, G.R. Oxygen-derived free radicals stimulate osteoclastic bone resorption in rodent bone in vitro and in vivo. J. Clin. Investig. 1990, 85, 632–639. [Google Scholar] [CrossRef]
- Ding, W.G.; Yan, W.H.; Wei, Z.X.; Liu, J.B. Difference in intraosseous blood vessel volume and number in osteoporotic model mice induced by spinal cord injury and sciatic nerve resection. J. Bone Miner. Metab. 2012, 30, 400–407. [Google Scholar] [CrossRef]
- Prisby, R.D.; Ramsey, M.W.; Behnke, B.J.; Dominguez, J.M.; Donato, A.J.; Allen, M.R.; Delp, M.D. Aging reduces skeletal blood flow, endothelium-dependent vasodilation, and NO bioavailability in rats. J. Bone Miner. Res. 2007, 22, 1280–1288. [Google Scholar] [CrossRef]
- Rosanoff, A.; Weaver, C.M.; Rude, R.K. Suboptimal magnesium status in the United States: Are the health consequences underestimated? Nutr. Rev. 2012, 70, 153–164. [Google Scholar] [CrossRef]
- Naderi, A.S.; Reilly, R.F. Hereditary etiologies of hypomagnesemia. Nat.Clin. Pract. Nephrol. 2008, 4, 80–89. [Google Scholar] [CrossRef]
- Meyer, T.E.; Verwoert, G.C.; Hwang, S.J.; Glazer, N.L.; Smith, A.V.; van Rooij, F.J.; Ehret, G.B.; Boerwinkle, E.; Felix, J.F.; Leak, T.S.; et al. Genome-wide association studies of serum magnesium, potassium, and sodium concentrations identify six loci influencing serum magnesium levels. PLoS Genet. 2010, 6. [Google Scholar] [CrossRef]
- Runnels, L.W. TRPM6 and TRPM7: A Mul-TRP-PLIK-cation of channel functions. Curr. Pharm. Biotechnol. 2011, 12, 42–53. [Google Scholar] [CrossRef]
- Geiger, H.; Wanner, C. Magnesium in diseases. Clin. Kidney Dis. 2012, 5, i25–i38. [Google Scholar] [CrossRef]
- Rodriguez-Moran, M.; Simental Mendia, L.E.; Zambrano-Galvan, G.; Guerrero-Romero, F. The role of magnesium in type 2 diabetes: A brief clinical-based review. Magnes. Res. 2011, 24, 156–162. [Google Scholar]
- Chacko, S.A.; Song, Y.; Nathan, L.; Tinker, L.; de Boer, I.H.; Tylavsky, F.; Wallace, R.; Liu, S. Relations of dietary magnesium intake to biomarkers of inflammation and endothelial dysfunction in an ethnically diverse cohort of postmenopausal women. Diabetes Care 2010, 33, 304–310. [Google Scholar] [CrossRef]
- Elshal, M.F.; Bernawi, A.E.; Al-Ghamdy, M.A.; Jalal, J.A. The association of bone mineral density and parathyroid hormone with serum magnesium in adult patients with sickle-cell anaemia. Arch. Med. Sci. 2012, 8, 270–276. [Google Scholar]
- Saito, N.; Tabata, N.; Saito, S.; Andou, Y.; Onaga, Y.; Iwamitsu, A.; Sakamoto, M.; Hori, T.; Sayama, H.; Kawakita, T. Bone mineral density, serum albumin and serum magnesium. J. Am. Coll. Nutr. 2004, 23, 701S–703S. [Google Scholar] [CrossRef]
- Tucker, K.L.; Hannan, M.T.; Chen, H.; Cupples, L.A.; Wilson, P.W.; Kiel, D.P. Potassium, magnesium, and fruit and vegetable intakes are associated with greater bone mineral density in elderly men and women. Am. J. Clin. Nutr. 1999, 69, 727–736. [Google Scholar]
- Ryder, K.M.; Shorr, R.I.; Bush, A.J.; Kritchevsky, S.B.; Harris, T.; Stone, K.; Cauley, J.; Tylavsky, F.A. Magnesium intake from food and supplements is associated with bone mineral density in healthy older white subjects. J. Am. Geriatr. Soc. 2005, 53, 1875–1880. [Google Scholar] [CrossRef]
- Stendig-Lindberg, G.; Koeller, W.; Bauer, A.; Rob, P.M. Experimentally induced prolonged magnesium deficiency causes osteoporosis in the rat. Eur. J. Intern. Med. 2004, 15, 97–107. [Google Scholar] [CrossRef]
- Tranquilli, A.L.; Lucino, E.; Garzetti, G.G.; Romanini, C. Calcium, phosphorus and magnesium intakes correlate with bone mineral content in postmenopausal women. Gynecol. Endocrinol. 1994, 8, 55–58. [Google Scholar] [CrossRef]
- Wang, M.C.; Moore, E.C.; Crawford, P.B.; Hudes, M.; Sabry, Z.I.; Marcus, R.; Bachrach, L.K. Influence of pre-adolescent diet on quantitative ultrasound measurements of the calcaneus in young adult women. Osteoporos. Int. 1999, 9, 532–535. [Google Scholar]
- Carpenter, T.O.; DeLucia, M.C.; Zhang, J.H.; Bejnerowicz, G.; Tartamella, L.; Dziura, J.; Petersen, K.F.; Befroy, D.; Cohen, D. A randomized controlled study of effects of dietary magnesium oxide supplementation on bone mineral content in healthy girls. J. Clin. Endocrinol. Metab. 2006, 91, 4866–4872. [Google Scholar] [CrossRef]
- Matias, C.N.; Santos, D.A.; Monteiro, C.P.; Vasco, A.M.; Baptista, F.; Sardinha, L.B.; Laires, M.J.; Silva, A.M. Magnesium intake mediates the association between bone mineral density and lean soft tissue in elite swimmers. Magnes. Res. 2012, 25, 120–125. [Google Scholar]
- Cohen, L.; Kitzes, R. Infrared spectroscopy and magnesium content of bone mineral in osteoporotic women. Isr. J. Med. Sci. 1981, 17, 1123–1125. [Google Scholar]
- Rude, R.K.; Adams, J.S.; Ryzen, E.; Endres, D.B.; Niimi, H.; Horst, R.L.; Haddad, J.G.; Singer, F.R. Low serum concentrations of 1,25-dihydroxyvitamin D in human magnesium deficiency. J. Clin. Endocrinol. Metab. 1985, 61, 933–940. [Google Scholar] [CrossRef]
- Sahota, O.; Mundey, M.K.; San, P.; Godber, I.M.; Hosking, D.J. Vitamin D insufficiency and the blunted PTH response in established osteoporosis: The role of magnesium deficiency. Osteoporos. Int. 2006, 17, 1013–1021. [Google Scholar] [CrossRef]
- Saggese, G.; Federico, G.; Bertelloni, S.; Baroncelli, G.I.; Calisti, L. Hypomagnesemia and the parathyroid hormone-vitamin D endocrine system in children with insulin-dependent diabetes mellitus: Effects of Mg administration. J. Pediatr. 1991, 118, 220–225. [Google Scholar] [CrossRef]
- Maier, J.A. Endothelial cells and magnesium: Implications in atherosclerosis. Clin. Sci. (Lond.) 2012, 122, 397–407. [Google Scholar] [CrossRef]
- Warburton, D.E.; Nicol, C.W.; Gatto, S.N.; Bredin, S.S. Cardiovascular disease and osteoporosis: Balancing risk management. Vasc. Health Risk Manag. 2007, 3, 673–689. [Google Scholar]
- Arnett, T.R. Extracellular pH regulates bone cell function. J. Nutr. 2008, 138, 415S–418S. [Google Scholar]
- Jehle, S.; Hulter, H.N.; Krapf, R. Effect of potassium citrate on bone density, microarchitecture, and fracture risk in healthy older adults without osteoporosis: A randomized controlled trial. J. Clin. Endocrinol. Metab. 2013, 98, 207–217. [Google Scholar] [CrossRef]
- Weng, L.; Webster, T.J. Nanostructured magnesium has fewer detrimental effects on osteoblast function. Int. J. Nanomed. 2013, 8, 1773–1781. [Google Scholar] [CrossRef]
- Navarro-González, J.F.; Mora-Fernández, C.; García-Pérez, J. Clinical implications of disordered magnesium homeostasis in chronic renal failure and dialysis. Semin. Dial. 2009, 22, 37–44. [Google Scholar] [CrossRef]
- Yokoyama, K.; Takahashi, N.; Yada, Y.; Yasunori, K.; Kawamata, R.; Uehara, R.; Kono, Y.; Honma, Y.; Momoi, M.Y. Prolonged maternal magnesium administration and bone metabolism in neonates. Early Hum. Dev. 2010, 86, 187–191. [Google Scholar] [CrossRef]
- Kaplan, W.; Haymond, M.W.; Karaviti, L.P. Osteopenic effects of MgSO4 in multiple pregnancies. J. Pediatr. Endocrinol. Metab. 2006, 19, 1225–1230. [Google Scholar]
- Leidi, M.; Dellera, F.; Mariotti, M.; Maier, J.A. High magnesium inhibits human osteoblast differentiation in vitro. Magnes. Res. 2011, 24, 1–6. [Google Scholar]
- Song, C.H.; Barrett-Connor, E.; Chung, J.H.; Kim, S.H.; Kim, K.S. Associations of calcium and magnesium in serum and hair with bone mineral density in premenopausal women. Biol. Trace Elem. Res. 2007, 118, 1–9. [Google Scholar] [CrossRef]
- Clevers, H. Wnt/beta-catenin signaling in development and disease. Cell 2006, 127, 469–480. [Google Scholar] [CrossRef]
- Albisetti, W.; Giarratana, L.S.; Viganò, C.; Castiglioni, S.; Maier, J.A. Sclerostin: A novel player regulating bone mass in inflammation? Eur. J. Inflamm. 2013. in press.
- Yang, C.; Yuan, G.; Zhang, J.; Tang, Z.; Zhang, X.; Dai, K. Effects of magnesium alloys extracts on adult human bone marrow-derived stromal cell viability and osteogenic differentiation. Biomed. Mater. 2010, 5. [Google Scholar] [CrossRef]
- Bonewald, L.F. The amazing osteocyte. J. BoneMiner. Res. 2011, 26, 229–238. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Castiglioni, S.; Cazzaniga, A.; Albisetti, W.; Maier, J.A.M. Magnesium and Osteoporosis: Current State of Knowledge and Future Research Directions. Nutrients 2013, 5, 3022-3033. https://doi.org/10.3390/nu5083022
Castiglioni S, Cazzaniga A, Albisetti W, Maier JAM. Magnesium and Osteoporosis: Current State of Knowledge and Future Research Directions. Nutrients. 2013; 5(8):3022-3033. https://doi.org/10.3390/nu5083022
Chicago/Turabian StyleCastiglioni, Sara, Alessandra Cazzaniga, Walter Albisetti, and Jeanette A. M. Maier. 2013. "Magnesium and Osteoporosis: Current State of Knowledge and Future Research Directions" Nutrients 5, no. 8: 3022-3033. https://doi.org/10.3390/nu5083022
APA StyleCastiglioni, S., Cazzaniga, A., Albisetti, W., & Maier, J. A. M. (2013). Magnesium and Osteoporosis: Current State of Knowledge and Future Research Directions. Nutrients, 5(8), 3022-3033. https://doi.org/10.3390/nu5083022