Iron, Human Growth, and the Global Epidemic of Obesity

Abstract
:1. Introduction
2. Iron Fortification and Supplementation

{kind=link}
| Intervention | Date |
|---|---|
| Fortification of food | 1941 |
| Fortification of infant formula | 1959 |
| Iron supplements to pregnant women | 1970s and 1980s |
| Iron supplements to general population | Recent |
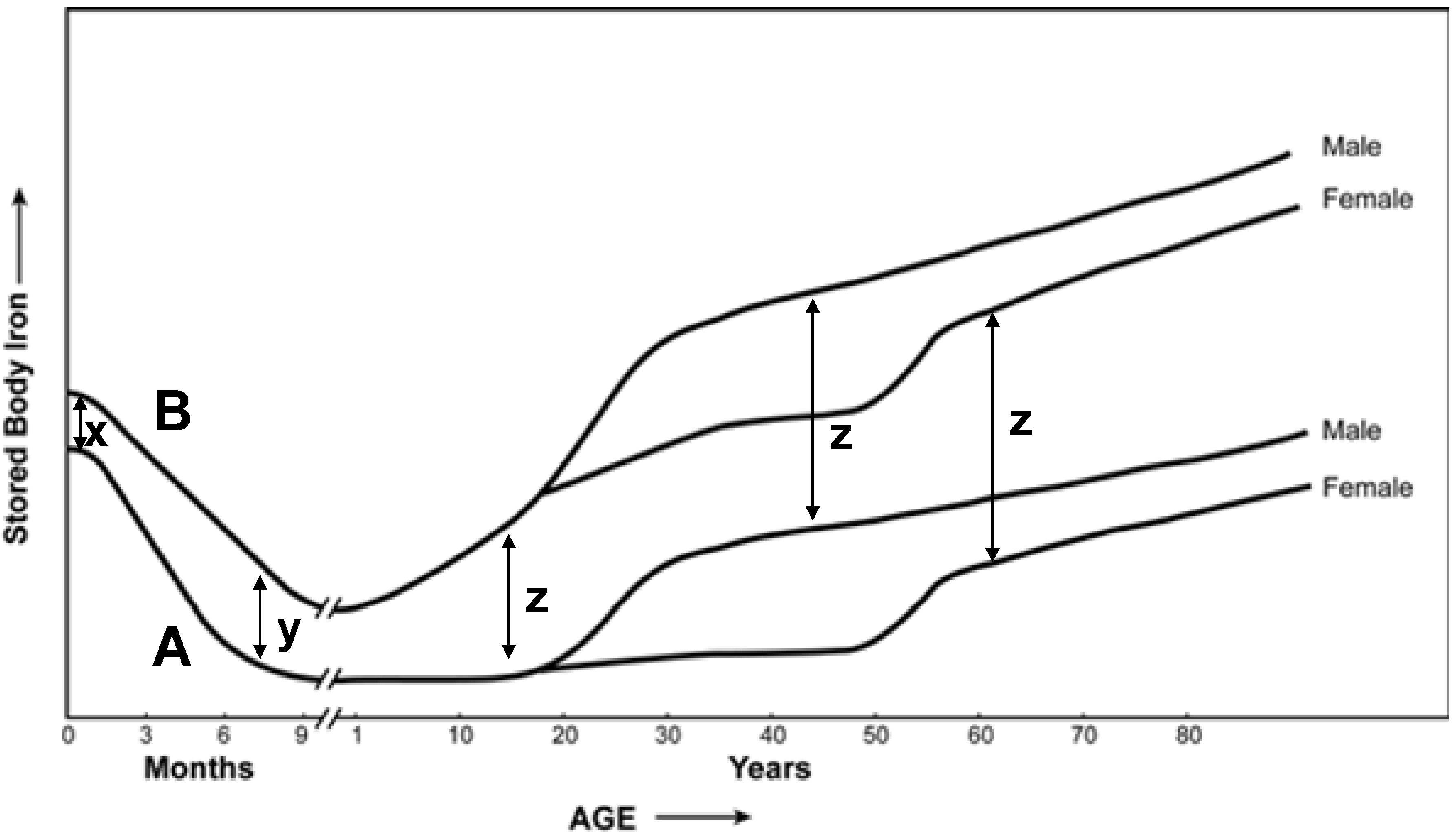
3. Body Iron after Fortification and Supplementation

4. Iron, Growth and Obesity
5. Diseases of Obesity and Iron
6. Conclusions
Conflicts of Interest
References
- Crichton, R.R.; Ward, R.J. An overview of iron metabolism: Molecular and cellular criteria for the selection of iron chelators. Curr. Med. Chem. 2003, 10, 997–1004. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC). Trends in wheat-flour fortification with folic acid and iron—Worldwide, 2004 and 2007. Morb. Mortal. Wkly. Rep. 2008, 57, 8–10.
- Dallman, P.R. Upper limits of iron in infant formulas. J. Nutr. 1989, 119, 1852–1855. [Google Scholar]
- Looker, A.C.; Dallman, P.R.; Carroll, M.D.; Gunter, E.W.; Johnson, C.L. Prevalence of iron deficiency in the united states. JAMA 1997, 277, 973–976. [Google Scholar] [CrossRef]
- Fleming, D.J.; Tucker, K.L.; Jacques, P.F.; Dallal, G.E.; Wilson, P.W.; Wood, R.J. Dietary factors associated with the risk of high iron stores in the elderly framingham heart study cohort. Am. J. Clin. Nutr. 2002, 76, 1375–1384. [Google Scholar]
- Wang, J.L.; Shaw, N.S. Iron status of the taiwanese elderly: The prevalence of iron deficiency and elevated iron stores. Asia Pac. J. Clin. Nutr. 2005, 14, 278–284. [Google Scholar]
- Pan, Y.; Jackson, R.T. Ethnic difference in the relationship between acute inflammation and serum ferritin in us adult males. Epidemiol. Infect. 2008, 136, 421–431. [Google Scholar]
- Leibold, E.A.; Munro, H.N. Cytoplasmic protein binds in vitro to a highly conserved sequence in the 5′ untranslated region of ferritin heavy- and light-subunit mrnas. Proc. Natl. Acad. Sci. USA 1988, 85, 2171–2175. [Google Scholar] [CrossRef]
- Hallberg, L.; Hulthen, L. Prediction of dietary iron absorption: An algorithm for calculating absorption and bioavailability of dietary iron. Am. J. Clin. Nutr. 2000, 71, 1147–1160. [Google Scholar]
- Kaneshige, E. Serum ferritin as an assessment of iron stores and other hematologic parameters during pregnancy. Obstet. Gynecol. 1981, 57, 238–242. [Google Scholar]
- Kelly, A.M.; MacDonald, D.J.; McDougall, A.N. Observations on maternal and fetal ferritin concentrations at term. Br. J. Obstet. Gynaecol. 1978, 85, 338–343. [Google Scholar] [CrossRef]
- Milman, N.; Agger, A.O.; Nielsen, O.J. Iron supplementation during pregnancy. Effect on iron status markers, serum erythropoietin and human placental lactogen. A placebo controlled study in 207 danish women. Dan. Med. Bull. 1991, 38, 471–476. [Google Scholar]
- Milman, N.; Ibsen, K.K.; Christensen, J.M. Serum ferritin and iron status in mothers and newborn infants. Acta Obstet. Gynecol. Scand. 1987, 66, 205–211. [Google Scholar] [CrossRef]
- Puolakka, J.; Janne, O.; Vihko, R. Evaluation by serum ferritin assay of the influence of maternal iron stores on the iron status of newborns and infants. Acta Obstet. Gynecol. Scand. Suppl. 1980, 95, 53–56. [Google Scholar] [CrossRef]
- Allen, L.H. Pregnancy and iron deficiency: Unresolved issues. Nutr. Rev. 1997, 55, 91–101. [Google Scholar] [CrossRef]
- Choi, J.W.; Kim, C.S.; Pai, S.H. Erythropoietic activity and soluble transferrin receptor level in neonates and maternal blood. Acta Paediatr. 2000, 89, 675–679. [Google Scholar] [CrossRef]
- Georgieff, M.K.; Mills, M.M.; Gordon, K.; Wobken, J.D. Reduced neonatal liver iron concentrations after uteroplacental insufficiency. J. Pediatr. 1995, 127, 308–311. [Google Scholar] [CrossRef]
- Hay, G.; Refsum, H.; Whitelaw, A.; Melbye, E.L.; Haug, E.; Borch-Iohnsen, B. Predictors of serum ferritin and serum soluble transferrin receptor in newborns and their associations with iron status during the first 2 y of life. Am. J. Clin. Nutr. 2007, 86, 64–73. [Google Scholar]
- Gill, D.G.; Vincent, S.; Segal, D.S. Follow-on formula in the prevention of iron deficiency: A multicentre study. Acta Paediatr. 1997, 86, 683–689. [Google Scholar] [CrossRef]
- Thane, C.W.; Bates, C.J.; Prentice, A. Risk factors for low iron intake and poor iron status in a national sample of british young people aged 4–18 years. Public Health Nutr. 2003, 6, 485–496. [Google Scholar]
- Milman, N.; Backer, V.; Laursen, E.M.; Graudal, N.; Ibsen, K.K.; Jordal, R. Serum ferritin in children and adolescents. Results from population surveys in 1979 and 1986 comprising 1312 individuals. Eur. J. Haematol. 1994, 53, 16–20. [Google Scholar]
- Gibson, S.; Ashwell, M. The association between red and processed meat consumption and iron intakes and status among british adults. Public Health Nutr. 2003, 6, 341–350. [Google Scholar]
- Milman, N.; Mathiassen, B.; Bohm, J.; Hansen, J.C. Serum ferritin in a greenlandic inuit hunter population from the thule district. Arct. Med. Res. 1992, 51, 10–15. [Google Scholar]
- Moyo, V.M.; Mvundura, E.; Khumalo, H.; Gangaidzo, I.T.; Saungweme, T.; Nouraie, M.; Rouault, T.A.; Gomo, Z.A.; Gordeuk, V.R. Serum ferritin concentrations in africans with low dietary iron. Ann. Hematol. 2009, 88, 1131–1136. [Google Scholar] [CrossRef]
- Hotz, C.; Porcayo, M.; Onofre, G.; Garcia-Guerra, A.; Elliott, T.; Jankowski, S.; Greiner, T. Efficacy of iron-fortified ultra rice in improving the iron status of women in Mexico. Food Nutr. Bull. 2008, 29, 140–149. [Google Scholar]
- Whittaker, P.; Tufaro, P.R.; Rader, J.I. Iron and folate in fortified cereals. J. Am. Coll. Nutr. 2001, 20, 247–254. [Google Scholar] [CrossRef]
- Mallory, M.A.; Sthapanachai, C.; Kowdley, K.V. Iron overload related to excessive vitamin c intake. Ann. Intern. Med. 2003, 139, 532–533. [Google Scholar]
- Lederman, H.M.; Cohen, A.; Lee, J.W.; Freedman, M.H.; Gelfand, E.W. Deferoxamine: A reversible s-phase inhibitor of human lymphocyte proliferation. Blood 1984, 64, 748–753. [Google Scholar]
- Yu, Y.; Kovacevic, Z.; Richardson, D.R. Tuning cell cycle regulation with an iron key. Cell Cycle 2007, 6, 1982–1994. [Google Scholar] [CrossRef]
- Hileti, D.; Panayiotidis, P.; Hoffbrand, A.V. Iron chelators induce apoptosis in proliferating cells. Br. J. Haematol. 1995, 89, 181–187. [Google Scholar] [CrossRef]
- Hershko, C. Control of disease by selective iron depletion: A novel therapeutic strategy utilizing iron chelators. Baillieres Clin. Haematol. 1994, 7, 965–1000. [Google Scholar] [CrossRef]
- Thelander, L.; Reichard, P. Reduction of ribonucleotides. Annu. Rev. Biochem. 1979, 48, 133–158. [Google Scholar] [CrossRef]
- Jordan, A.; Reichard, P. Ribonucleotide reductases. Annu. Rev. Biochem. 1998, 67, 71–98. [Google Scholar] [CrossRef]
- Terada, N.; Lucas, J.J.; Gelfand, E.W. Differential regulation of the tumor suppressor molecules, retinoblastoma susceptibility gene product (rb) and p53, during cell cycle progression of normal human t cells. J. Immunol. 1991, 147, 698–704. [Google Scholar]
- Becker, E.M.; Lovejoy, D.B.; Greer, J.M.; Watts, R.; Richardson, D.R. Identification of the di-pyridyl ketone isonicotinoyl hydrazone (pkih) analogues as potent iron chelators and anti-tumour agents. Br. J. Pharmacol. 2003, 138, 819–830. [Google Scholar] [CrossRef]
- Chaston, T.B.; Watts, R.N.; Yuan, J.; Richardson, D.R. Potent antitumor activity of novel iron chelators derived from di-2-pyridylketone isonicotinoyl hydrazone involves fenton-derived free radical generation. Clin. Cancer Res. 2004, 10, 7365–7374. [Google Scholar] [CrossRef]
- Kalinowski, D.S.; Sharpe, P.C.; Bernhardt, P.V.; Richardson, D.R. Design, synthesis, and characterization of new iron chelators with anti-proliferative activity: Structure-activity relationships of novel thiohydrazone analogues. J. Med. Chem. 2007, 50, 6212–6225. [Google Scholar] [CrossRef]
- Ponka, P.; Borova, J.; Neuwirt, J.; Fuchs, O. Mobilization of iron from reticulocytes. Identification of pyridoxal isonicotinoyl hydrazone as a new iron chelating agent. FEBS Lett. 1979, 97, 317–321. [Google Scholar] [CrossRef]
- Ponka, P.; Borova, J.; Neuwirt, J.; Fuchs, O.; Necas, E. A study of intracellular iron metabolism using pyridoxal isonicotinoyl hydrazone and other synthetic chelating agents. Biochim. Biophys. Acta 1979, 586, 278–297. [Google Scholar] [CrossRef]
- Richardson, D.R.; Tran, E.H.; Ponka, P. The potential of iron chelators of the pyridoxal isonicotinoyl hydrazone class as effective antiproliferative agents. Blood 1995, 86, 4295–4306. [Google Scholar]
- Rudland, P.S.; Durbin, H.; Clingan, D.; de Asua, L.J. Iron salts and transferrin are specifically required for cell division of cultured 3T6 cells. Biochem. Biophys. Res. Commun. 1977, 75, 556–562. [Google Scholar] [CrossRef]
- Oguchi, S.; Walker, W.A.; Sanderson, I.R. Iron saturation alters the effect of lactoferrin on the proliferation and differentiation of human enterocytes (caco-2 cells). Biol. Neonate 1995, 67, 330–339. [Google Scholar] [CrossRef]
- Keenan, J.; Clynes, M. Replacement of transferrin by simple iron compounds for mdck cells grown and subcultured in serum-free medium. In Vitro Cell. Dev. Biol. Anim. 1996, 32, 451–453. [Google Scholar] [CrossRef]
- Laskey, J.; Webb, I.; Schulman, H.M.; Ponka, P. Evidence that transferrin supports cell proliferation by supplying iron for DNA synthesis. Exp. Cell Res. 1988, 176, 87–95. [Google Scholar] [CrossRef]
- Fan, L.; Iyer, J.; Zhu, S.; Frick, K.K.; Wada, R.K.; Eskenazi, A.E.; Berg, P.E.; Ikegaki, N.; Kennett, R.H.; Frantz, C.N. Inhibition of N-myc expression and induction of apoptosis by iron chelation in human neuroblastoma cells. Cancer Res. 2001, 61, 1073–1079. [Google Scholar]
- Fukuchi, K.; Tomoyasu, S.; Tsuruoka, N.; Gomi, K. Iron deprivation-induced apoptosis in hl-60 cells. FEBS Lett. 1994, 350, 139–142. [Google Scholar] [CrossRef]
- Hoyes, K.P.; Hider, R.C.; Porter, J.B. Cell cycle synchronization and growth inhibition by 3-hydroxypyridin-4-one iron chelators in leukemia cell lines. Cancer Res. 1992, 52, 4591–4599. [Google Scholar]
- Sun, X.; Ge, R.; Cai, Z.; Sun, H.; He, Q.Y. Iron depletion decreases proliferation and induces apoptosis in a human colonic adenocarcinoma cell line, caco2. J. Inorg. Biochem. 2009, 103, 1074–1081. [Google Scholar] [CrossRef]
- Kaltreider, D.F.; Johnson, J.W. Patients at high risk for low-birth-weight delivery. Am. J. Obstet. Gynecol. 1976, 124, 251–256. [Google Scholar]
- Lee, H.S.; Kim, M.S.; Kim, M.H.; Kim, Y.J.; Kim, W.Y. Iron status and its association with pregnancy outcome in korean pregnant women. Eur. J. Clin. Nutr. 2006, 60, 1130–1135. [Google Scholar] [CrossRef]
- Roszkowski, I.; Wojcicka, J.; Zaleska, K. Serum iron deficiency during the third trimester of pregnancy: Maternal complications and fate of the neonate. Obstet. Gynecol. 1966, 28, 820–825. [Google Scholar]
- VanNagell, J.; Koepke, J.; Dilts, P.V., Jr. Preventable anemia and pregnancy. Obstet. Gynecol. Surv. 1971, 26, 551–563. [Google Scholar] [CrossRef]
- Palma, S.; Perez-Iglesias, R.; Prieto, D.; Pardo, R.; Llorca, J.; Delgado-Rodriguez, M. Iron but not folic acid supplementation reduces the risk of low birthweight in pregnant women without anaemia: A case-control study. J. Epidemiol. Community Health 2008, 62, 120–124. [Google Scholar] [CrossRef]
- Aukett, M.A.; Parks, Y.A.; Scott, P.H.; Wharton, B.A. Treatment with iron increases weight gain and psychomotor development. Arch. Dis. Child. 1986, 61, 849–857. [Google Scholar] [CrossRef]
- Bandhu, R.; Shankar, N.; Tandon, O.P. Effect of iron on growth in iron deficient anemic school going children. Indian J. Physiol. Pharmacol. 2003, 47, 59–66. [Google Scholar]
- Chwang, L.C.; Soemantri, A.G.; Pollitt, E. Iron supplementation and physical growth of rural indonesian children. Am. J. Clin. Nutr. 1988, 47, 496–501. [Google Scholar]
- Latham, M.C.; Stephenson, L.S.; Kinoti, S.N.; Zaman, M.S.; Kurz, K.M. Improvements in growth following iron supplementation in young kenyan school children. Nutrition 1990, 6, 159–165. [Google Scholar]
- Morais, M.B.; Ferrari, A.A.; Fisberg, M. Effect of oral iron therapy on physical growth. Rev. Paul. Med. 1993, 111, 439–444. [Google Scholar]
- Stoltzfus, R.J.; Heidkamp, R.; Kenkel, D.; Habicht, J.P. Iron supplementation of young children: Learning from the new evidence. Food Nutr. Bull. 2007, 28, S572–S584. [Google Scholar]
- Gunnarsson, B.S.; Thorsdottir, I.; Palsson, G. Iron status in 6-y-old children: Associations with growth and earlier iron status. Eur. J. Clin. Nutr. 2005, 59, 761–767. [Google Scholar] [CrossRef] [Green Version]
- Bougle, D.; Laroche, D.; Bureau, F. Zinc and iron status and growth in healthy infants. Eur. J. Clin. Nutr. 2000, 54, 764–767. [Google Scholar] [CrossRef]
- Bruner, A.B.; Joffe, A.; Duggan, A.K.; Casella, J.F.; Brandt, J. Randomised study of cognitive effects of iron supplementation in non-anaemic iron-deficient adolescent girls. Lancet 1996, 348, 992–996. [Google Scholar] [CrossRef]
- Beasley, N.M.; Tomkins, A.M.; Hall, A.; Lorri, W.; Kihamia, C.M.; Bundy, D.A. The impact of weekly iron supplementation on the iron status and growth of adolescent girls in tanzania. Trop. Med. Int. Health 2000, 5, 794–799. [Google Scholar] [CrossRef]
- De Sanctis, V.; Katz, M.; Vullo, C.; Bagni, B.; Ughi, M.; Wonke, B. Effect of different treatment regimes on linear growth and final height in beta-thalassaemia major. Endocrinol. (Oxf.) 1994, 40, 791–798. [Google Scholar] [CrossRef]
- Iwasaki, T.; Nakajima, A.; Yoneda, M.; Yamada, Y.; Mukasa, K.; Fujita, K.; Fujisawa, N.; Wada, K.; Terauchi, Y. Serum ferritin is associated with visceral fat area and subcutaneous fat area. Diabetes Care 2005, 28, 2486–2491. [Google Scholar] [CrossRef]
- Gillum, R.F. Association of serum ferritin and indices of body fat distribution and obesity in mexican american men—The third national health and nutrition examination survey. Int. J. Obes. Relat. Metab. Disord. 2001, 25, 639–645. [Google Scholar] [CrossRef]
- Fernandez-Real, J.M.; Vendrell, J.; Baiget, M.; Gimferrer, E.; Ricart, W. C282y and h63d mutations of the hemochromatosis candidate gene in type 2 diabetes. Diabetes Care 1999, 22, 525–526. [Google Scholar] [CrossRef]
- Jehn, M.; Clark, J.M.; Guallar, E. Serum ferritin and risk of the metabolic syndrome in U.S. Adults. Diabetes Care 2004, 27, 2422–2428. [Google Scholar] [CrossRef]
- Vari, I.S.; Balkau, B.; Kettaneh, A.; Andre, P.; Tichet, J.; Fumeron, F.; Caces, E.; Marre, M.; Grandchamp, B.; Ducimetiere, P. Ferritin and transferrin are associated with metabolic syndrome abnormalities and their change over time in a general population: Data from an epidemiological study on the insulin resistance syndrome (desir). Diabetes Care 2007, 30, 1795–1801. [Google Scholar] [CrossRef]
- Alissa, E.M.; Ahmed, W.H.; Al-Ama, N.; Ferns, G.A. Relationship between indices of iron status and coronary risk factors including diabetes and the metabolic syndrome in saudi subjects without overt coronary disease. J. Trace Elem. Med. Biol. 2007, 21, 242–254. [Google Scholar] [CrossRef]
- Halle, M.; Konig, D.; Berg, A.; Keul, J.; Baumstark, M.W. Relationship of serum ferritin concentrations with metabolic cardiovascular risk factors in men without evidence for coronary artery disease. Atherosclerosis 1997, 128, 235–240. [Google Scholar] [CrossRef]
- Mainous, A.G., III; Wells, B.J.; Everett, C.J.; Gill, J.M.; King, D.E. Association of ferritin and lipids with c-reactive protein. Am. J. Cardiol. 2004, 93, 559–562. [Google Scholar] [CrossRef]
- Piperno, A.; Trombini, P.; Gelosa, M.; Mauri, V.; Pecci, V.; Vergani, A.; Salvioni, A.; Mariani, R.; Mancia, G. Increased serum ferritin is common in men with essential hypertension. J. Hypertens. 2002, 20, 1513–1518. [Google Scholar] [CrossRef]
- Sung, K.C.; Kang, J.H.; Shin, H.S. Relationship of cardiovascular risk factors and serum ferritin with c-reactive protein. Arch. Med. Res. 2007, 38, 121–125. [Google Scholar] [CrossRef]
- Williams, M.J.; Poulton, R.; Williams, S. Relationship of serum ferritin with cardiovascular risk factors and inflammation in young men and women. Atherosclerosis 2002, 165, 179–184. [Google Scholar] [CrossRef]
- Fernandez-Real, J.M.; Ricart-Engel, W.; Arroyo, E.; Balanca, R.; Casamitjana-Abella, R.; Cabrero, D.; Fernandez-Castaner, M.; Soler, J. Serum ferritin as a component of the insulin resistance syndrome. Diabetes Care 1998, 21, 62–68. [Google Scholar]
- Fumeron, F.; Pean, F.; Driss, F.; Balkau, B.; Tichet, J.; Marre, M.; Grandchamp, B. Ferritin and transferrin are both predictive of the onset of hyperglycemia in men and women over 3 years: The data from an epidemiological study on the insulin resistance syndrome (desir) study. Diabetes Care 2006, 29, 2090–2094. [Google Scholar] [CrossRef]
- Rajpathak, S.N.; Crandall, J.P.; Wylie-Rosett, J.; Kabat, G.C.; Rohan, T.E.; Hu, F.B. The role of iron in type 2 diabetes in humans. Biochim. Biophys. Acta 2009, 1790, 671–681. [Google Scholar]
- Lao, T.T.; Chan, P.L.; Tam, K.F. Gestational diabetes mellitus in the last trimester—A feature of maternal iron excess? Diabet. Med. 2001, 18, 218–223. [Google Scholar] [CrossRef]
- Lao, T.T.; Tam, K.F. Maternal serum ferritin and gestational impaired glucose tolerance. Diabetes Care 1997, 20, 1368–1369. [Google Scholar]
- Chen, X.; Scholl, T.O.; Stein, T.P. Association of elevated serum ferritin levels and the risk of gestational diabetes mellitus in pregnant women: The camden study. Diabetes Care 2006, 29, 1077–1082. [Google Scholar] [CrossRef]
- Tarim, E.; Kilicdag, E.; Bagis, T.; Ergin, T. High maternal hemoglobin and ferritin values as risk factors for gestational diabetes. Int. J. Gynaecol. Obstet. 2004, 84, 259–261. [Google Scholar] [CrossRef]
- Bo, S.; Menato, G.; Villois, P.; Gambino, R.; Cassader, M.; Cotrino, I.; Cavallo-Perin, P. Iron supplementation and gestational diabetes in midpregnancy. Am. J. Obstet. Gynecol. 201, 158.e151–158.e156.
- Lee, D.H.; Liu, D.Y.; Jacobs, D.R., Jr.; Shin, H.R.; Song, K.; Lee, I.K.; Kim, B.; Hider, R.C. Common presence of non-transferrin-bound iron among patients with type 2 diabetes. Diabetes Care 2006, 29, 1090–1095. [Google Scholar] [CrossRef]
- Salonen, J.T.; Tuomainen, T.P.; Nyyssonen, K.; Lakka, H.M.; Punnonen, K. Relation between iron stores and non-insulin dependent diabetes in men: Case-control study. BMJ 1998, 317, 727–730. [Google Scholar] [CrossRef]
- Jiang, R.; Ma, J.; Ascherio, A.; Stampfer, M.J.; Willett, W.C.; Hu, F.B. Dietary iron intake and blood donations in relation to risk of type 2 diabetes in men: A prospective cohort study. Am. J. Clin. Nutr. 2004, 79, 70–75. [Google Scholar]
- Lee, D.H.; Folsom, A.R.; Jacobs, D.R., Jr. Dietary iron intake and type 2 diabetes incidence in postmenopausal women: The iowa women’s health study. Diabetologia 2004, 47, 185–194. [Google Scholar] [CrossRef]
- Rajpathak, S.; Ma, J.; Manson, J.; Willett, W.C.; Hu, F.B. Iron intake and the risk of type 2 diabetes in women: A prospective cohort study. Diabetes Care 2006, 29, 1370–1376. [Google Scholar] [CrossRef]
- Liu, Q.; Sun, L.; Tan, Y.; Wang, G.; Lin, X.; Cai, L. Role of iron deficiency and overload in the pathogenesis of diabetes and diabetic complications. Curr. Med. Chem. 2009, 16, 113–129. [Google Scholar] [CrossRef]
- Swaminathan, S.; Fonseca, V.A.; Alam, M.G.; Shah, S.V. The role of iron in diabetes and its complications. Diabetes Care 2007, 30, 1926–1933. [Google Scholar] [CrossRef]
- Ascherio, A.; Rimm, E.B.; Giovannucci, E.; Willett, W.C.; Stampfer, M.J. Blood donations and risk of coronary heart disease in men. Circulation 2001, 103, 52–57. [Google Scholar] [CrossRef]
- Fernandez-Real, J.M.; Lopez-Bermejo, A.; Ricart, W. Iron stores, blood donation, and insulin sensitivity and secretion. Clin. Chem. 2005, 51, 1201–1205. [Google Scholar] [CrossRef]
- Cutler, P. Deferoxamine therapy in high-ferritin diabetes. Diabetes 1989, 38, 1207–1210. [Google Scholar] [CrossRef]
- Kannel, W.B.; Hjortland, M.C.; McNamara, P.M.; Gordon, T. Menopause and risk of cardiovascular disease: The framingham study. Ann. Intern. Med. 1976, 85, 447–452. [Google Scholar] [CrossRef]
- Tuomainen, T.P.; Punnonen, K.; Nyyssonen, K.; Salonen, J.T. Association between body iron stores and the risk of acute myocardial infarction in men. Circulation 1998, 97, 1461–1466. [Google Scholar] [CrossRef]
- Salonen, J.T.; Nyyssonen, K.; Korpela, H.; Tuomilehto, J.; Seppanen, R.; Salonen, R. High stored iron levels are associated with excess risk of myocardial infarction in eastern finnish men. Circulation 1992, 86, 803–811. [Google Scholar] [CrossRef]
- Van Lenten, B.J.; Prieve, J.; Navab, M.; Hama, S.; Lusis, A.J.; Fogelman, A.M. Lipid-induced changes in intracellular iron homeostasis in vitro and in vivo. J. Clin. Investig. 1995, 95, 2104–2110. [Google Scholar] [CrossRef]
- Stadler, N.; Lindner, R.A.; Davies, M.J. Direct detection and quantification of transition metal ions in human atherosclerotic plaques: Evidence for the presence of elevated levels of iron and copper. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 949–954. [Google Scholar] [CrossRef]
- De Valk, B.; Marx, J.J. Iron, atherosclerosis, and ischemic heart disease. Arch. Intern. Med. 1999, 159, 1542–1548. [Google Scholar]
- Asmis, R.; Begley, J.G. Oxidized ldl promotes peroxide-mediated mitochondrial dysfunction and cell death in human macrophages: A caspase-3-independent pathway. Circ. Res. 2003, 92, e20–e29. [Google Scholar] [CrossRef]
- Matthews, A.J.; Vercellotti, G.M.; Menchaca, H.J.; Bloch, P.H.; Michalek, V.N.; Marker, P.H.; Murar, J.; Buchwald, H. Iron and atherosclerosis: Inhibition by the iron chelator deferiprone (L1). J. Surg. Res. 1997, 73, 35–40. [Google Scholar] [CrossRef]
- Meyers, D.G. The iron hypothesis—Does iron cause atherosclerosis? Clin. Cardiol. 1996, 19, 925–929. [Google Scholar] [CrossRef]
- Sullivan, J.L. Iron in arterial plaque: Modifiable risk factor for atherosclerosis. Biochim. Biophys. Acta 2009, 1790, 718–723. [Google Scholar] [CrossRef]
- Qi, L.; van Dam, R.M.; Rexrode, K.; Hu, F.B. Heme iron from diet as a risk factor for coronary heart disease in women with type 2 diabetes. Diabetes Care 2007, 30, 101–106. [Google Scholar] [CrossRef]
- Ascherio, A.; Willett, W.C.; Rimm, E.B.; Giovannucci, E.L.; Stampfer, M.J. Dietary iron intake and risk of coronary disease among men. Circulation 1994, 89, 969–974. [Google Scholar] [CrossRef]
- Klipstein-Grobusch, K.; Grobbee, D.E.; den Breeijen, J.H.; Boeing, H.; Hofman, A.; Witteman, J.C. Dietary iron and risk of myocardial infarction in the rotterdam study. Am. J. Epidemiol. 1999, 149, 421–428. [Google Scholar] [CrossRef]
- Lee, D.H.; Folsom, A.R.; Jacobs, D.R., Jr. Iron, zinc, and alcohol consumption and mortality from cardiovascular diseases: The iowa women’s health study. Am. J. Clin. Nutr. 2005, 81, 787–791. [Google Scholar]
- Malaviarachchi, D.; Veugelers, P.J.; Yip, A.M.; MacLean, D.R. Dietary iron as a risk factor for myocardial infarction. Public health considerations for nova scotia. Can. J. Public Health 2002, 93, 267–270. [Google Scholar]
- Van der A, D.L.; Grobbee, D.E.; Roest, M.; Marx, J.J.; Voorbij, H.A.; van der Schouw, Y.T. Serum ferritin is a risk factor for stroke in postmenopausal women. Stroke 2005, 36, 1637–1641. [Google Scholar] [CrossRef]
- Kiechl, S.; Willeit, J.; Egger, G.; Poewe, W.; Oberhollenzer, F. Body iron stores and the risk of carotid atherosclerosis: Prospective results from the bruneck study. Circulation 1997, 96, 3300–3307. [Google Scholar] [CrossRef]
- Meyers, D.G.; Strickland, D.; Maloley, P.A.; Seburg, J.K.; Wilson, J.E.; McManus, B.F. Possible association of a reduction in cardiovascular events with blood donation. Heart 1997, 78, 188–193. [Google Scholar]
- Tuomainen, T.P.; Salonen, R.; Nyyssonen, K.; Salonen, J.T. Cohort study of relation between donating blood and risk of myocardial infarction in 2682 men in eastern finland. BMJ 1997, 314, 793–794. [Google Scholar] [CrossRef]
- Castellanos, M.; Puig, N.; Carbonell, T.; Castillo, J.; Martinez, J.; Rama, R.; Davalos, A. Iron intake increases infarct volume after permanent middle cerebral artery occlusion in rats. Brain Res. 2002, 952, 1–6. [Google Scholar] [CrossRef]
- Mehta, S.H.; Webb, R.C.; Ergul, A.; Tawfik, A.; Dorrance, A.M. Neuroprotection by tempol in a model of iron-induced oxidative stress in acute ischemic stroke. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2004, 286, R283–R288. [Google Scholar]
- Kiechl, S.; Aichner, F.; Gerstenbrand, F.; Egger, G.; Mair, A.; Rungger, G.; Spogler, F.; Jarosch, E.; Oberhollenzer, F.; Willeit, J. Body iron stores and presence of carotid atherosclerosis. Results from the bruneck study. Arterioscler. Thromb. 1994, 14, 1625–1630. [Google Scholar] [CrossRef]
- Davalos, A.; Fernandez-Real, J.M.; Ricart, W.; Soler, S.; Molins, A.; Planas, E.; Genis, D. Iron-related damage in acute ischemic stroke. Stroke 1994, 25, 1543–1546. [Google Scholar] [CrossRef]
- Millan, M.; Sobrino, T.; Castellanos, M.; Nombela, F.; Arenillas, J.F.; Riva, E.; Cristobo, I.; Garcia, M.M.; Vivancos, J.; Serena, J.; et al. Increased body iron stores are associated with poor outcome after thrombolytic treatment in acute stroke. Stroke 2007, 38, 90–95. [Google Scholar] [CrossRef]
- Duffy, S.J.; Biegelsen, E.S.; Holbrook, M.; Russell, J.D.; Gokce, N.; Keaney, J.F., Jr.; Vita, J.A. Iron chelation improves endothelial function in patients with coronary artery disease. Circulation 2001, 103, 2799–2804. [Google Scholar] [CrossRef]
- Pratico, D.; Pasin, M.; Barry, O.P.; Ghiselli, A.; Sabatino, G.; Iuliano, L.; FitzGerald, G.A.; Violi, F. Iron-dependent human platelet activation and hydroxyl radical formation: Involvement of protein kinase c. Circulation 1999, 99, 3118–3124. [Google Scholar] [CrossRef]
- Selim, M.H.; Ratan, R.R. The role of iron neurotoxicity in ischemic stroke. Ageing Res. Rev. 2004, 3, 345–353. [Google Scholar] [CrossRef]
- Carbonell, T.; Rama, R. Iron, oxidative stress and early neurological deterioration in ischemic stroke. Curr. Med. Chem. 2007, 14, 857–874. [Google Scholar] [CrossRef]
- Zacharski, L.R.; Ornstein, D.L.; Woloshin, S.; Schwartz, L.M. Association of age, sex, and race with body iron stores in adults: Analysis of nhanes iii data. Am. Heart J. 2000, 140, 98–104. [Google Scholar] [CrossRef]
- Mainous, A.G., III; Gill, J.M.; Carek, P.J. Elevated serum transferrin saturation and mortality. Ann. Fam. Med. 2004, 2, 133–138. [Google Scholar] [CrossRef]
- Mainous, A.G., III; Wells, B.; Carek, P.J.; Gill, J.M.; Geesey, M.E. The mortality risk of elevated serum transferrin saturation and consumption of dietary iron. Ann. Fam. Med. 2004, 2, 139–144. [Google Scholar] [CrossRef]
- Mainous, A.G., III; Gill, J.M.; Everett, C.J. Transferrin saturation, dietary iron intake, and risk of cancer. Ann. Fam. Med. 2005, 3, 131–137. [Google Scholar] [CrossRef]
- Edgren, G.; Reilly, M.; Hjalgrim, H.; Tran, T.N.; Rostgaard, K.; Adami, J.; Titlestad, K.; Shanwell, A.; Melbye, M.; Nyren, O. Donation frequency, iron loss, and risk of cancer among blood donors. J. Natl. Cancer Inst. 2008, 100, 572–579. [Google Scholar] [CrossRef]
- Merk, K.; Mattsson, B.; Mattsson, A.; Holm, G.; Gullbring, B.; Bjorkholm, M. The incidence of cancer among blood donors. Int. J. Epidemiol. 1990, 19, 505–509. [Google Scholar] [CrossRef]
- Hjalgrim, H.; Edgren, G.; Rostgaard, K.; Reilly, M.; Tran, T.N.; Titlestad, K.E.; Shanwell, A.; Jersild, C.; Adami, J.; Wikman, A.; et al. Cancer incidence in blood transfusion recipients. J. Natl. Cancer Inst. 2007, 99, 1864–1874. [Google Scholar]
- Kato, J.; Miyanishi, K.; Kobune, M.; Nakamura, T.; Takada, K.; Takimoto, R.; Kawano, Y.; Takahashi, S.; Takahashi, M.; Sato, Y.; et al. Long-term phlebotomy with low-iron diet therapy lowers risk of development of hepatocellular carcinoma from chronic hepatitis c. J. Gastroenterol. 2007, 42, 830–836. [Google Scholar]
- Zacharski, L.R.; Chow, B.K.; Howes, P.S.; Shamayeva, G.; Baron, J.A.; Dalman, R.L.; Malenka, D.J.; Ozaki, C.K.; Lavori, P.W. Decreased cancer risk after iron reduction in patients with peripheral arterial disease: Results from a randomized trial. J. Natl. Cancer Inst. 2008, 100, 996–1002. [Google Scholar]
- Wurzelmann, J.I.; Silver, A.; Schreinemachers, D.M.; Sandler, R.S.; Everson, R.B. Iron intake and the risk of colorectal cancer. Cancer Epidemiol. Biomark. Prev. 1996, 5, 503–507. [Google Scholar]
- Bird, C.L.; Witte, J.S.; Swendseid, M.E.; Shikany, J.M.; Hunt, I.F.; Frankl, H.D.; Lee, E.R.; Longnecker, M.P.; Haile, R.W. Plasma ferritin, iron intake, and the risk of colorectal polyps. Am. J. Epidemiol. 1996, 144, 34–41. [Google Scholar] [CrossRef]
- Nelson, R.L.; Davis, F.G.; Sutter, E.; Sobin, L.H.; Kikendall, J.W.; Bowen, P. Body iron stores and risk of colonic neoplasia. J. Natl. Cancer Inst. 1994, 86, 455–460. [Google Scholar]
- Graf, E.; Eaton, J.W. Dietary suppression of colonic cancer. Fiber or phytate? Cancer 1985, 56, 717–718. [Google Scholar]
- Campbell, S.; Ghosh, S. Ulcerative colitis and colon cancer: Strategies for cancer prevention. Dig. Dis. 2002, 20, 38–48. [Google Scholar] [CrossRef]
- Deugnier, Y. Iron and liver cancer. Alcohol 2003, 30, 145–150. [Google Scholar] [CrossRef]
- Toyokuni, S. Role of iron in carcinogenesis: Cancer as a ferrotoxic disease. Cancer Sci. 2009, 100, 9–16. [Google Scholar] [CrossRef]
- Huang, X. Iron overload and its association with cancer risk in humans: Evidence for iron as a carcinogenic metal. Mutat. Res. 2003, 533, 153–171. [Google Scholar] [CrossRef]
- Hann, H.W.; Kim, C.Y.; London, W.T.; Blumberg, B.S. Increased serum ferritin in chronic liver disease: A risk factor for primary hepatocellular carcinoma. Int. J. Cancer 1989, 43, 376–379. [Google Scholar] [CrossRef]
- Linet, M.S.; Gridley, G.; Nyren, O.; Mellemkjaer, L.; Olsen, J.H.; Keehn, S.; Adami, H.O.; Fraumeni, J.F., Jr. Primary liver cancer, other malignancies, and mortality risks following porphyria: A cohort study in denmark and sweden. Am. J. Epidemiol. 1999, 149, 1010–1015. [Google Scholar] [CrossRef]
- Grando-Lemaire, V.; Guettier, C.; Chevret, S.; Beaugrand, M.; Trinchet, J.C. Hepatocellular carcinoma without cirrhosis in the west: Epidemiological factors and histopathology of the non-tumorous liver. Groupe d’etude et de traitement du carcinome hepatocellulaire. J. Hepatol. 1999, 31, 508–513. [Google Scholar]
- Turlin, B.; Juguet, F.; Moirand, R.; Le Quilleuc, D.; Loreal, O.; Campion, J.P.; Launois, B.; Ramee, M.P.; Brissot, P.; Deugnier, Y. Increased liver iron stores in patients with hepatocellular carcinoma developed on a noncirrhotic liver. Hepatology 1995, 22, 446–450. [Google Scholar]
- Bhasin, G.; Kauser, H.; Athar, M. Low iron state is associated with reduced tumor promotion in a two-stage mouse skin carcinogenesis model. Food Chem. Toxicol. 2002, 40, 1105–1111. [Google Scholar]
- Goldstein, S.R.; Yang, G.Y.; Chen, X.; Curtis, S.K.; Yang, C.S. Studies of iron deposits, inducible nitric oxide synthase and nitrotyrosine in a rat model for esophageal adenocarcinoma. Carcinogenesis 1998, 19, 1445–1449. [Google Scholar] [CrossRef]
- Milman, N.; Byg, K.E.; Ovesen, L. Iron status in danes 1994. Ii: Prevalence of iron deficiency and iron overload in 1319 danish women aged 40–70 years. Influence of blood donation, alcohol intake and iron supplementation. Ann. Hematol. 2000, 79, 612–621. [Google Scholar] [CrossRef]
- Milman, N.; Byg, K.E.; Ovesen, L.; Kirchhoff, M.; Jurgensen, K.S. Iron status in danish women, 1984–1994: A cohort comparison of changes in iron stores and the prevalence of iron deficiency and iron overload. Eur. J. Haematol. 2003, 71, 51–61. [Google Scholar] [CrossRef]
- Milman, N.; Byg, K.E.; Ovesen, L.; Kirchhoff, M.; Jurgensen, K.S. Iron status in danish men 1984–94: A cohort comparison of changes in iron stores and the prevalence of iron deficiency and iron overload. Eur. J. Haematol. 2002, 68, 332–340. [Google Scholar] [CrossRef]
- German, A.J. The growing problem of obesity in dogs and cats. J. Nutr. 2006, 136, 1940S–1946S. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Sangani, R.G.; Ghio, A.J. Iron, Human Growth, and the Global Epidemic of Obesity. Nutrients 2013, 5, 4231-4249. https://doi.org/10.3390/nu5104231
Sangani RG, Ghio AJ. Iron, Human Growth, and the Global Epidemic of Obesity. Nutrients. 2013; 5(10):4231-4249. https://doi.org/10.3390/nu5104231
Chicago/Turabian StyleSangani, Rahul G., and Andrew J. Ghio. 2013. "Iron, Human Growth, and the Global Epidemic of Obesity" Nutrients 5, no. 10: 4231-4249. https://doi.org/10.3390/nu5104231
APA StyleSangani, R. G., & Ghio, A. J. (2013). Iron, Human Growth, and the Global Epidemic of Obesity. Nutrients, 5(10), 4231-4249. https://doi.org/10.3390/nu5104231