Calcium Nutrition and Extracellular Calcium Sensing: Relevance for the Pathogenesis of Osteoporosis, Cancer and Cardiovascular Diseases


Abstract
:1. Introduction

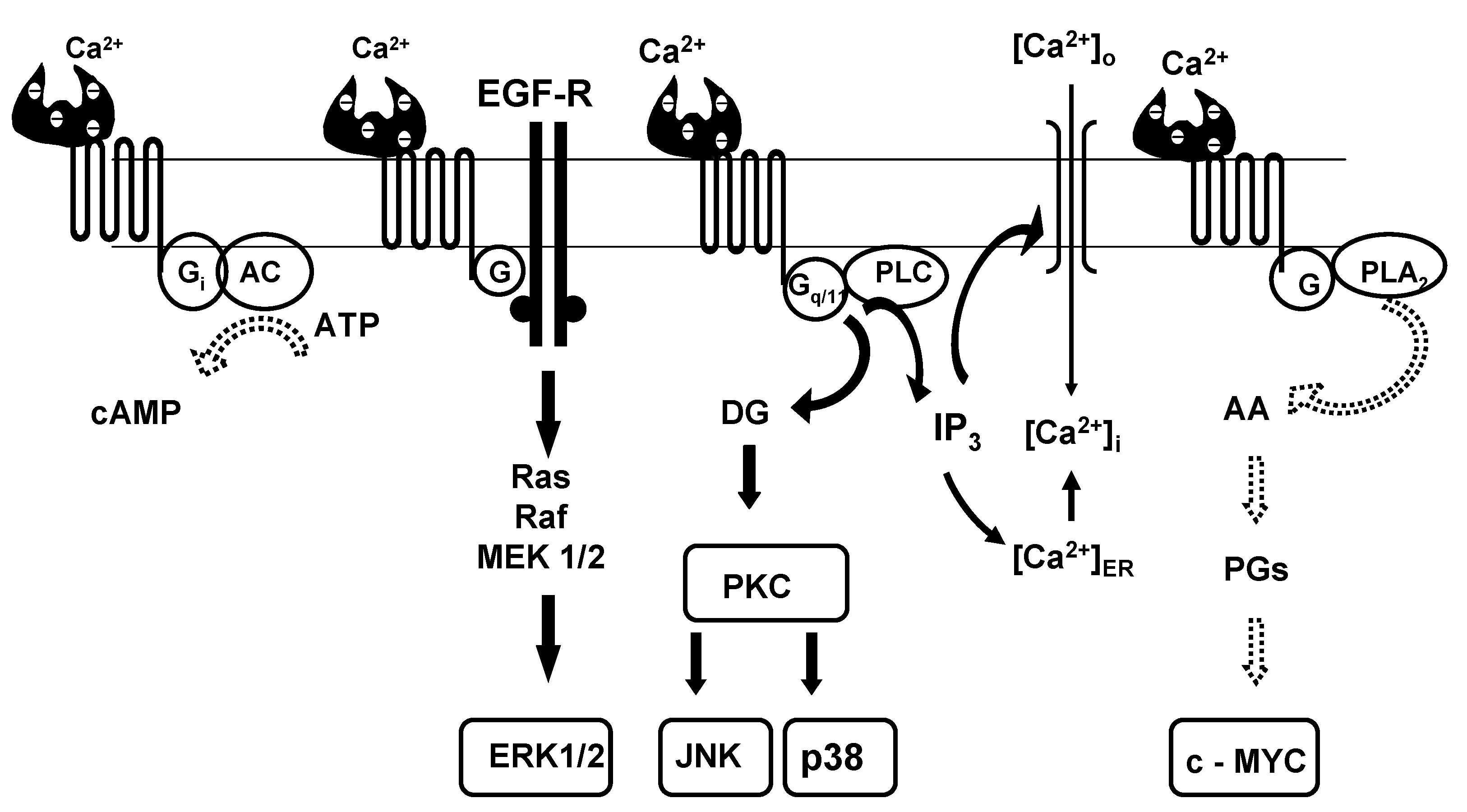{kind=link}
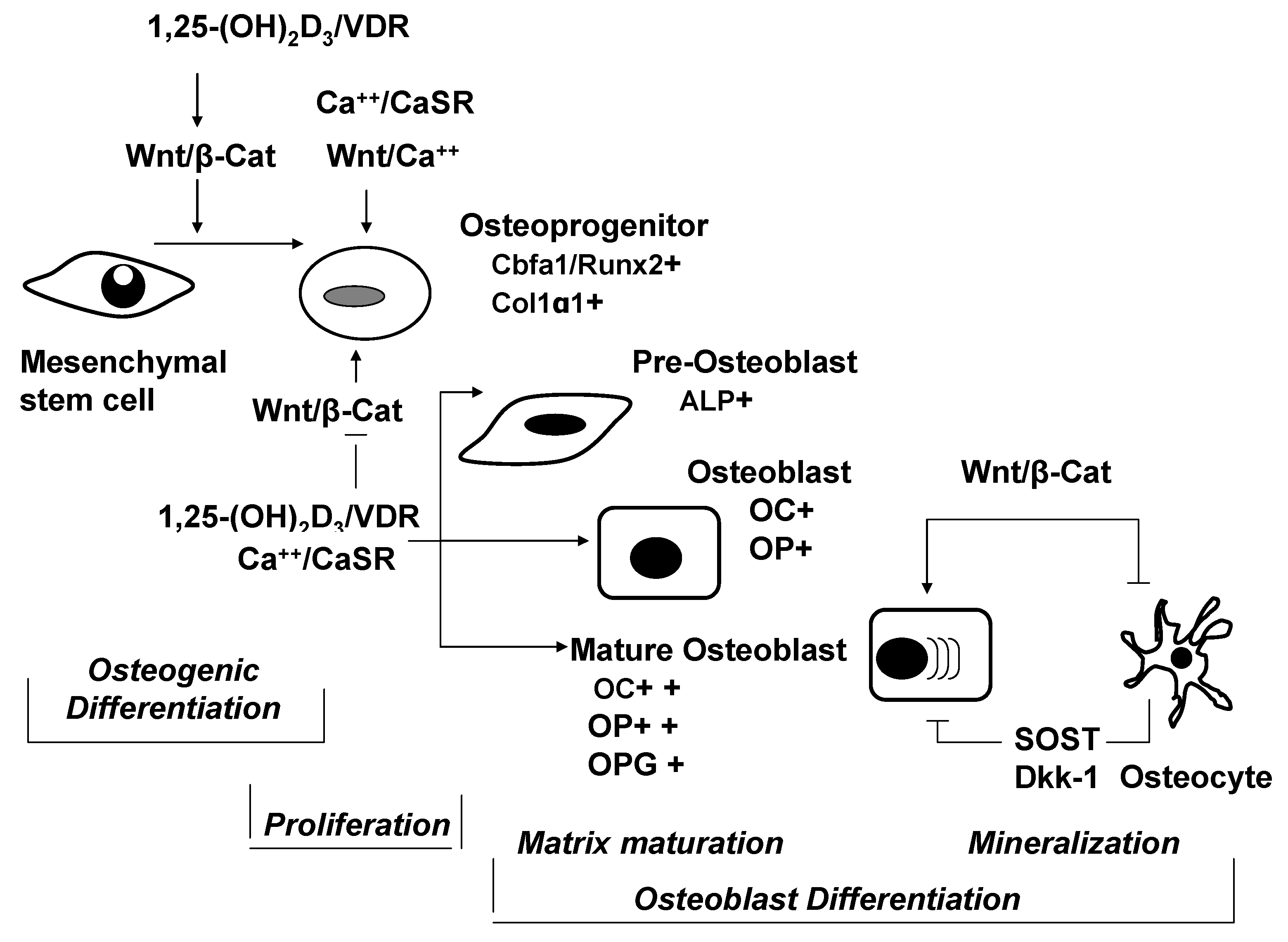{kind=link}
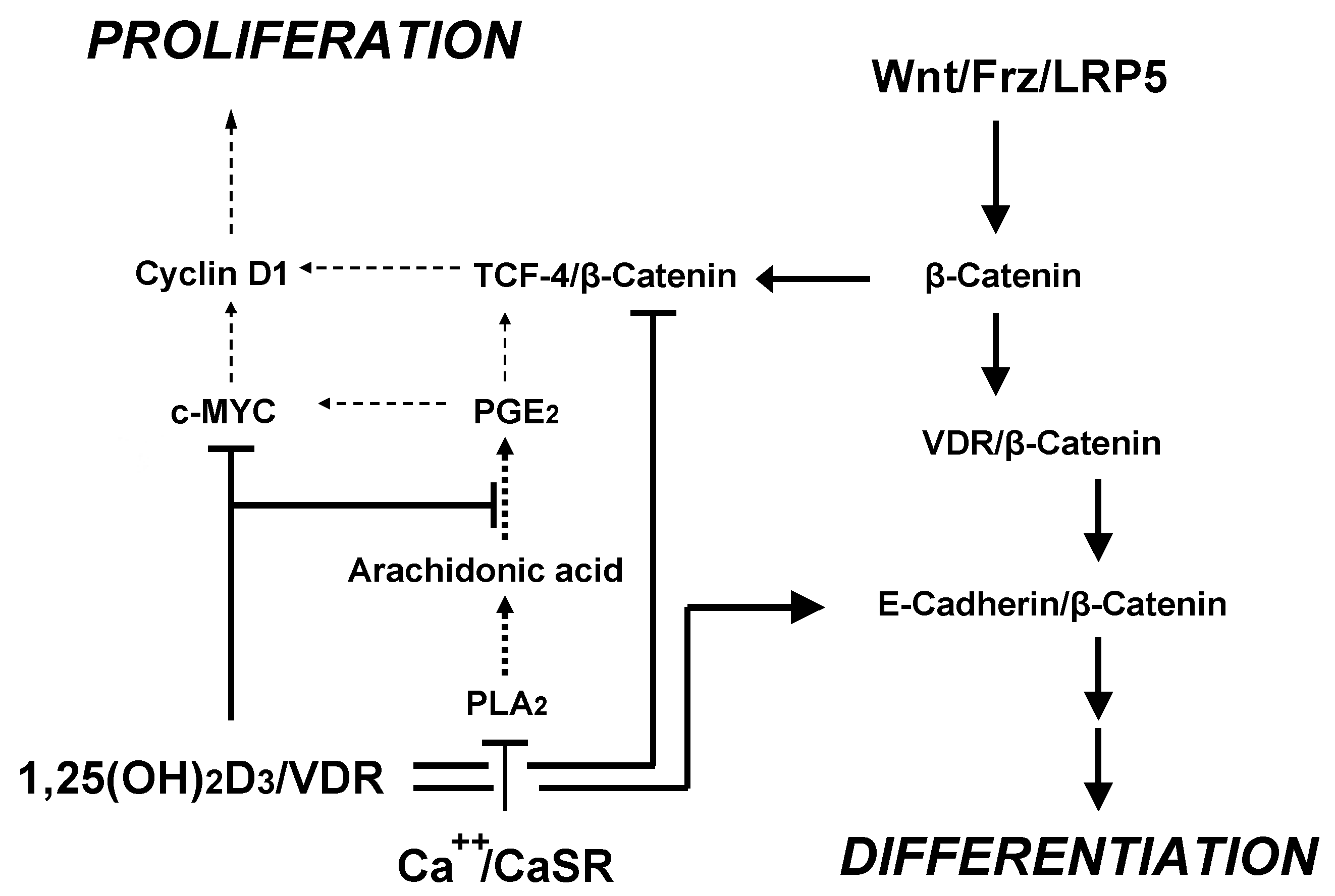{kind=link}
| Country | Age (years) | RDA a (mg/day) | Calcium intake (mg/day) | Study | |
|---|---|---|---|---|---|
| Gender | |||||
| Male | Female | ||||
| Europe | |||||
| Austria | 19–79 | 1000–1300 | 561 (±290) b | 576 (±309) b | Kudlacek et al. [8] |
| Belgium | 75–80 | 1300 | 748 (324–1166) e | 676 (287–1101) e | Amorim Cruz et al. [9] |
| Denmark | 70–75 | 1300 | 544 (127–1812) e | Andersen et al. [10] | |
| France | 75–80 | 1300 | 620 (402–1010) e | 635 (428–944) e | Amorim Cruz et al. [9] |
| Germany | 18–79 | 1000–1300 | 1181 (902–1535) e | 1082 (849–1379) e | Hintzpeter et al. [11] |
| Netherlands | 75–80 | 1300 | 1036 (725–1447) e | 1010 (612–1616) e | Amorim Cruz et al. [9] |
| Poland | 70–75 | 1300 | 325 (86–851) e | Andersen et al. [10] | |
| Middle East | |||||
| Lebanon | 10–16 | 1300 | 873 (793–952) d | 673 (595–750) d | Salamoun et al. [12] |
| America | |||||
| Canada | 18–35 | 1000 | 562 (0–2630) e | Rubin et al. [13] | |
| USA | 31–50 | 1000 | 1118 (±25) b | 864 (±20) b | Bailey et al. [14] |
| USA | >55 | 1300 | 611 (381–892) e | Lappe et al. [15] | |
| Brazil | 16–20 | 1300 | 659 (596–721) d | 881 (730–1032) d | Peters et al. [16] |
| SE Asia | |||||
| Bangladesh | 16–40 | 1000 | 180 c | Islam et al. [17] | |
| Indonesia | 18–40 | 1000 | 270 (239–302) d | Green et al. [18] | |
| Malaysia | 18–40 | 1000 | 386 (353–420) d | Green et al. [18] | |
| China | >55 | 1300 | 485 (±253) b | Kruger et al. [19] | |
| Japan | 65–75 | 1300 | 527 (±195) b | Nakamura et al. [20] | |
| Oceania | |||||
| Australia | 20–94 | 1000–1300 | 643 (±340) b | Pasco et al. [21] | |
2. Calcium Malnutrition and Disease Incidence: Epidemiological Evidence
2.1. Bone Diseases
2.1.1. Calcium Deficiency and Rickets
2.1.2. Osteoporosis, Falls and Fractures
2.2. Cancer
2.2.1. Colorectal Cancer
2.2.2. Breast Cancer
2.2.3. Prostate Cancer
2.3. Cardiovascular Disease
2.3.1. Cardiovascular Risk Factors
2.3.2. Cardiovascular Disease: Incidence and Mortality
3. Principles of Extracellular Calcium Sensing

3.1. The Extracellular Calcium-Sensing Receptor (CaSR): General Properties and Function
3.2. The Extracellular Calcium-Sensing Receptor (CaSR): Molecular Properties
3.3. The Extracellular Calcium-Sensing Receptor (CaSR): Cell-Specific Actions
3.3.1. Osteoblasts

3.3.2. Cancer Cells

3.3.3. Cardiac, Vascular, Renal Juxtaglomerular and Epithelial Cells
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Peterlik, M.; Cross, H.S. Vitamin D and calcium deficits predispose for multiple chronic diseases. Eur. J. Clin. Investig. 2005, 35, 290–304. [Google Scholar] [CrossRef]
- Peterlik, M.; Cross, H.S. Vitamin D and calcium insufficiency-related chronic diseases: Molecular and cellular pathophysiology. Eur. J. Clin. Nutr. 2009, 63, 1377–1386. [Google Scholar] [CrossRef]
- Peterlik, M.; Grant, W.B.; Cross, H.S. Calcium, vitamin D and cancer. Anticancer Res. 2009, 29, 3687–3698. [Google Scholar]
- Ward, B.K.; Magno, A.L.; Walsh, J.P.; Ratajczak, T. The role of the calcium-sensing receptor in human disease. Clin. Biochem. 2012, 45, 943–953. [Google Scholar] [CrossRef]
- Peterlik, M.; Boonen, S.; Cross, H.S.; Lamberg-Allardt, C. Vitamin D and calcium insufficiency-related chronic diseases: An emerging world-wide public health problem. Int. J. Environ. Res. Public Health 2009, 6, 2585–2607. [Google Scholar] [CrossRef]
- Ross, A.C.; Manson, J.E.; Abrams, S.A.; Aloia, J.F.; Brannon, P.M.; Clinton, S.K.; Durazo-Arvizu, R.A.; Gallagher, J.C.; Gallo, R.L.; Jones, G.; et al. The 2011 report on dietary reference intakes for calcium and vitamin D from the Institute of Medicine: What clinicians need to know. J. Clin. Endocrinol. Metab. 2011, 96, 53–58. [Google Scholar] [CrossRef]
- FAO/WHO, Human Vitamin and Mineral Requirements; FAO/WHO non-series publication: Rome, Italy, 2002.
- Kudlacek, S.; Schneider, B.; Peterlik, M.; Leb, G.; Klaushofer, K.; Weber, K.; Woloszczuk, W.; Willvonseder, R. Assessment of vitamin D and calcium status in healthy adult Austrians. Eur. J. Clin. Investig. 2003, 33, 323–331. [Google Scholar] [CrossRef]
- Amorim Cruz, J.A.; Moreiras, O.; Brzozowska, A. Longitudinal changes in the intake of vitamins and minerals of elderly Europeans. Eur. J. Clin. Nutr. 1996, 50, S77–S85. [Google Scholar]
- Andersen, R.; Molgaard, C.; Skovgaard, L.T.; Brot, C.; Cashman, K.D.; Chabros, E.; Charzewska, J.; Flynn, A.; Jakobsen, J.; Karkkainen, M.; et al. Teenage girls and elderly women living in northern Europe have low winter vitamin D status. Eur. J. Clin. Nutr. 2005, 59, 533–541. [Google Scholar] [CrossRef]
- Hintzpeter, B.; Mensink, G.B.; Thierfelder, W.; Muller, M.J.; Scheidt-Nave, C. Vitamin D status and health correlates among German adults. Eur. J. Clin. Nutr. 2008, 62, 1079–1089. [Google Scholar] [CrossRef]
- Salamoun, M.M.; Kizirian, A.S.; Tannous, R.I.; Nabulsi, M.M.; Choucair, M.K.; Deeb, M.E.; El-Hajj Fuleihan, G.A. Low calcium and vitamin D intake in healthy children and adolescents and their correlates. Eur. J. Clin. Nutr. 2005, 59, 177–184. [Google Scholar] [CrossRef]
- Rubin, L.A.; Hawker, G.A.; Peltekova, V.D.; Fielding, L.J.; Ridout, R.; Cole, D.E. Determinants of peak bone mass: Clinical and genetic analyses in a young female Canadian cohort. J. Bone Miner. Res. 1999, 14, 633–643. [Google Scholar] [CrossRef]
- Bailey, R.L.; Dodd, K.W.; Goldman, J.A.; Gahche, J.J.; Dwyer, J.T.; Moshfegh, A.J.; Sempos, C.T.; Picciano, M.F. Estimation of total usual calcium and vitamin D intakes in the United States. J. Nutr. 2010, 140, 817–822. [Google Scholar] [CrossRef]
- Lappe, J.M.; Davies, K.M.; Travers-Gustafson, D.; Heaney, R.P. Vitamin D status in a rural postmenopausal female population. J. Am. Coll. Nutr. 2006, 25, 395–402. [Google Scholar]
- Peters, B.S.; dos Santos, L.C.; Fisberg, M.; Wood, R.J.; Martini, L.A. Prevalence of vitamin D insufficiency in Brazilian adolescents. Ann. Nutr. Metab. 2009, 54, 15–21. [Google Scholar]
- Islam, M.Z.; Lamberg-Allardt, C.; Karkkainen, M.; Ali, S.M. Dietary calcium intake in premenopausal Bangladeshi women: Do socio-economic or physiological factors play a role? Eur. J. Clin. Nutr. 2003, 57, 674–680. [Google Scholar] [CrossRef]
- Green, T.J.; Skeaff, C.M.; Rockell, J.E.; Venn, B.J.; Lambert, A.; Todd, J.; Khor, G.L.; Loh, S.P.; Muslimatun, S.; Agustina, R.; et al. Vitamin D status and its association with parathyroid hormone concentrations in women of child-bearing age living in Jakarta and Kuala Lumpur. Eur. J. Clin. Nutr. 2008, 62, 373–378. [Google Scholar] [CrossRef]
- Kruger, M.C.; Ha, P.C.; Todd, J.M.; Kuhn-Sherlock, B.; Schollum, L.M.; Ma, J.; Qin, G.; Lau, E. High-calcium, vitamin D fortified milk is effective in improving bone turnover markers and vitamin D status in healthy postmenopausal Chinese women. Eur. J. Clin. Nutr. 2012, 66, 856–861. [Google Scholar]
- Nakamura, K.; Saito, T.; Yoshihara, A.; Ishikawa, M.; Tsuchiya, Y.; Oshiki, R.; Kobayashi, R.; Maruyama, K.; Hyodo, K.; Nashimoto, M.; et al. Low calcium intake is associated with increased bone resorption in postmenopausal Japanese women: Yokogoshi study. Public Health Nutr. 2009, 12, 2366–2370. [Google Scholar] [CrossRef]
- Pasco, J.A.; Henry, M.J.; Nicholson, G.C.; Brennan, S.L.; Kotowicz, M.A. Behavioural and physical characteristics associated with vitamin D status in women. Bone 2009, 44, 1085–1091. [Google Scholar] [CrossRef]
- Rejnmark, L.; Avenell, A.; Masud, T.; Anderson, F.; Meyer, H.E.; Sanders, K.M.; Salovaara, K.; Cooper, C.; Smith, H.E.; Jacobs, E.T.; et al. Vitamin D with calcium reduces mortality: Patient level pooled analysis of 70,528 patients from eight major vitamin D trials. J. Clin. Endocrinol. Metab. 2012, 97, 2670–2681. [Google Scholar] [CrossRef]
- Zhu, Y.; Mahon, B.D.; Froicu, M.; Cantorna, M.T. Calcium and 1α,25-dihydroxyvitamin D3 target the TNF-α pathway to suppress experimental inflammatory bowel disease. Eur. J. Immunol. 2005, 35, 217–224. [Google Scholar] [CrossRef]
- Cantorna, M.T.; Humpal-Winter, J.; DeLuca, H.F. Dietary calcium is a major factor in 1,25-dihydroxycholecalciferol suppression of experimental autoimmune encephalomyelitis in mice. J. Nutr. 1999, 129, 1966–1971. [Google Scholar]
- Mathieu, C.; van Etten, E.; Gysemans, C.; Decallonne, B.; Kato, S.; Laureys, J.; Depovere, J.; Valckx, D.; Verstuyf, A.; Bouillon, R. In vitro and in vivo analysis of the immune system of vitamin D receptor knockout mice. J. Bone Miner. Res. 2001, 16, 2057–2065. [Google Scholar] [CrossRef]
- Olszak, I.T.; Poznansky, M.C.; Evans, R.H.; Olson, D.; Kos, C.; Pollak, M.R.; Brown, E.M.; Scadden, D.T. Extracellular calcium elicits a chemokinetic response from monocytes in vitro and in vivo. J. Clin. Investig. 2000, 105, 1299–1305. [Google Scholar] [CrossRef]
- Tiosano, D.; Hochberg, Z. Hypophosphatemia: The common denominator of all rickets. J. Bone Miner. Metab. 2009, 27, 392–401. [Google Scholar] [CrossRef]
- Pettifor, J.M. Nutritional rickets: Deficiency of vitamin D, calcium, or both? Am. J. Clin. Nutr. 2004, 80, 1725S–1729S. [Google Scholar]
- Thacher, T.D.; Fischer, P.R.; Pettifor, J.M.; Lawson, J.O.; Isichei, C.O.; Reading, J.C.; Chan, G.M. A comparison of calcium, vitamin D, or both for nutritional rickets in Nigerian children. N. Engl. J. Med. 1999, 341, 563–568. [Google Scholar] [CrossRef]
- Balasubramanian, K.; Rajeswari, J.; Gulab; Govil, Y.C.; Agarwal, A.K.; Kumar, A.; Bhatia, V. Varying role of vitamin D deficiency in the etiology of rickets in young children vs. adolescents in northern India. J. Trop. Pediatr. 2003, 49, 201–206. [Google Scholar] [CrossRef]
- DeLucia, M.C.; Mitnick, M.E.; Carpenter, T.O. Nutritional rickets with normal circulating 25-hydroxyvitamin D: A call for reexamining the role of dietary calcium intake in North American infants. J. Clin. Endocrinol. Metab. 2003, 88, 3539–3545. [Google Scholar] [CrossRef]
- Riggs, B.L.; Khosla, S.; Melton, L.J., III. A unitary model for involutional osteoporosis: Estrogen deficiency causes both type I and type II osteoporosis in postmenopausal women and contributes to bone loss in aging men. J. Bone Miner. Res. 1998, 13, 763–773. [Google Scholar] [CrossRef]
- Heaney, R.P. Calcium in the prevention and treatment of osteoporosis. J. Intern. Med. 1992, 231, 169–180. [Google Scholar] [CrossRef]
- Dawson-Hughes, B.; Dallal, G.E.; Krall, E.A.; Sadowski, L.; Sahyoun, N.; Tannenbaum, S. A controlled trial of the effect of calcium supplementation on bone density in postmenopausal women. N. Engl. J. Med. 1990, 323, 878–883. [Google Scholar] [CrossRef]
- Elders, P.J.; Lips, P.; Netelenbos, J.C.; van Ginkel, F.C.; Khoe, E.; van der Vijgh, W.J.; van der Stelt, P.F. Long-term effect of calcium supplementation on bone loss in perimenopausal women. J. Bone Miner. Res. 1994, 9, 963–970. [Google Scholar]
- Recker, R.R.; Hinders, S.; Davies, K.M.; Heaney, R.P.; Stegman, M.R.; Lappe, J.M.; Kimmel, D.B. Correcting calcium nutritional deficiency prevents spine fractures in elderly women. J. Bone Miner. Res. 1996, 11, 1961–1966. [Google Scholar]
- Dawson-Hughes, B.; Harris, S.S.; Krall, E.A.; Dallal, G.E. Effect of calcium and vitamin D supplementation on bone density in men and women 65 years of age or older. N. Engl. J. Med. 1997, 337, 670–676. [Google Scholar]
- Chapuy, M.C.; Arlot, M.E.; Duboeuf, F.; Brun, J.; Crouzet, B.; Arnaud, S.; Delmas, P.D.; Meunier, P.J. Vitamin D3 and calcium to prevent hip fractures in the elderly women. N. Engl. J. Med. 1992, 327, 1637–1642. [Google Scholar] [CrossRef]
- Larsen, E.R.; Mosekilde, L.; Foldspang, A. Vitamin D and calcium supplementation prevents osteoporotic fractures in elderly community dwelling residents: A pragmatic population-based 3-year intervention study. J. Bone Miner. Res. 2004, 19, 370–378. [Google Scholar]
- Boonen, S.; Lips, P.; Bouillon, R.; Bischoff-Ferrari, H.A.; Vanderschueren, D.; Haentjens, P. Need for additional calcium to reduce the risk of hip fracture with vitamin D supplementation: Evidence from a comparative metaanalysis of randomized controlled trials. J. Clin. Endocrinol. Metab. 2007, 92, 1415–1423. [Google Scholar] [CrossRef]
- Abrahamsen, B.; Masud, T.; Avenell, A.; Anderson, F.; Meyer, H.E.; Cooper, J.C.; Smith, H.; LaCroix, A.Z.; Torgerson, D.; Johansen, A.; et al. Patient level pooled analysis of 68,500 patients from seven major vitamin D fracture trials in US and Europe. BMJ 2010, 340, b5463. [Google Scholar]
- Tang, B.M.; Eslick, G.D.; Nowson, C.; Smith, C.; Bensoussan, A. Use of calcium or calcium in combination with vitamin D supplementation to prevent fractures and bone loss in people aged 50 years and older: A meta-analysis. Lancet 2007, 370, 657–666. [Google Scholar] [CrossRef]
- Aloia, J.F. African Americans, 25-hydroxyvitamin D, and osteoporosis: A paradox. Am. J. Clin. Nutr. 2008, 88, 545S–550S. [Google Scholar]
- Pfeifer, M.; Begerow, B.; Minne, H.W.; Abrams, C.; Nachtigall, D.; Hansen, C. Effects of a short-term vitamin D and calcium supplementation on body sway and secondary hyperparathyroidism in elderly women. J. Bone Miner. Res. 2000, 15, 1113–1118. [Google Scholar] [CrossRef]
- Pfeifer, M.; Begerow, B.; Minne, H.W.; Suppan, K.; Fahrleitner-Pammer, A.; Dobnig, H. Effects of a long-term vitamin D and calcium supplementation on falls and parameters of muscle function in community-dwelling older individuals. Osteoporos. Int. 2009, 20, 315–322. [Google Scholar] [CrossRef]
- Lips, P.; Bouillon, R.; van Schoor, N.M.; Vanderschueren, D.; Verschueren, S.; Kuchuk, N.; Milisen, K.; Boonen, S. Reducing fracture risk with calcium and vitamin D. Clin. Endocrinol. 2010, 73, 277–285. [Google Scholar] [CrossRef]
- Park, Y.; Leitzmann, M.F.; Subar, A.F.; Hollenbeck, A.; Schatzkin, A. Dairy food, calcium, and risk of cancer in the NIH-AARP diet and health study. Arch. Intern. Med. 2009, 169, 391–401. [Google Scholar] [CrossRef]
- Garland, C.; Shekelle, R.B.; Barrett-Connor, E.; Criqui, M.H.; Rossof, A.H.; Paul, O. Dietary vitamin D and calcium and risk of colorectal cancer: A 19-year prospective study in men. Lancet 1985, 1, 307–309. [Google Scholar]
- Garland, C.F.; Garland, F.C.; Gorham, E.D. Can colon cancer incidence and death rates be reduced with calcium and vitamin D? Am. J. Clin. Nutr. 1991, 54, 193S–201S. [Google Scholar]
- Huncharek, M.; Muscat, J.; Kupelnick, B. Colorectal cancer risk and dietary intake of calcium, vitamin D, and dairy products: A meta-analysis of 26,335 cases from 60 observational studies. Nutr. Cancer 2009, 61, 47–69. [Google Scholar] [CrossRef]
- Wu, K.; Willett, W.C.; Fuchs, C.S.; Colditz, G.A.; Giovannucci, E.L. Calcium intake and risk of colon cancer in women and men. J. Natl. Cancer Inst. 2002, 94, 437–446. [Google Scholar] [CrossRef]
- Cho, E.; Smith-Warner, S.A.; Spiegelman, D.; Beeson, W.L.; van den Brandt, P.A.; Colditz, G.A.; Folsom, A.R.; Fraser, G.E.; Freudenheim, J.L.; Giovannucci, E.; et al. Dairy foods, calcium, and colorectal cancer: A pooled analysis of 10 cohort studies. J. Natl. Cancer Inst. 2004, 96, 1015–1022. [Google Scholar] [CrossRef]
- Jenab, M.; Bueno-de-Mesquita, H.B.; Ferrari, P.; van Duijnhoven, F.J.; Norat, T.; Pischon, T.; Jansen, E.H.; Slimani, N.; Byrnes, G.; Rinaldi, S.; et al. Association between pre-diagnostic circulating vitamin D concentration and risk of colorectal cancer in European populations:A nested case-control study. BMJ 2010, 340, b5500. [Google Scholar]
- Mizoue, T.; Kimura, Y.; Toyomura, K.; Nagano, J.; Kono, S.; Mibu, R.; Tanaka, M.; Kakeji, Y.; Maehara, Y.; Okamura, T.; et al. Calcium, dairy foods, vitamin D, and colorectal cancer risk: The Fukuoka colorectal cancer study. Cancer Epidemiol. Biomarkers Prev. 2008, 17, 2800–2807. [Google Scholar] [CrossRef]
- Ishihara, J.; Inoue, M.; Iwasaki, M.; Sasazuki, S.; Tsugane, S. Dietary calcium, vitamin D, and the risk of colorectal cancer. Am. J. Clin. Nutr. 2008, 88, 1576–1583. [Google Scholar] [CrossRef]
- Fedirko, V.; Riboli, E.; Tjonneland, A.; Ferrari, P.; Olsen, A.; Bueno-de-Mesquita, H.B.; van Duijnhoven, F.J.; Norat, T.; Jansen, E.H.; Dahm, C.C.; et al. Prediagnostic 25-hydroxyvitamin D, VDR and CaSR polymorphisms, and survival in patients with colorectal cancer in western European populations. Cancer Epidemiol. Biomarkers Prev. 2012, 21, 582–593. [Google Scholar] [CrossRef]
- Lipkin, M.; Newmark, H.L. Vitamin D, calcium and prevention of breast cancer: A review. J. Am. Coll. Nutr. 1999, 18, 392S–397S. [Google Scholar]
- Lin, J.; Manson, J.E.; Lee, I.M.; Cook, N.R.; Buring, J.E.; Zhang, S.M. Intakes of calcium and vitamin D and breast cancer risk in women. Arch. Intern. Med. 2007, 167, 1050–1059. [Google Scholar]
- Shin, M.H.; Holmes, M.D.; Hankinson, S.E.; Wu, K.; Colditz, G.A.; Willett, W.C. Intake of dairy products, calcium, and vitamin D and risk of breast cancer. J. Natl. Cancer. Inst. 2002, 94, 1301–1311. [Google Scholar] [CrossRef]
- McCullough, M.L.; Rodriguez, C.; Diver, W.R.; Feigelson, H.S.; Stevens, V.L.; Thun, M.J.; Calle, E.E. Dairy, calcium, and vitamin D intake and postmenopausal breast cancer risk in the cancer prevention study II nutrition cohort. Cancer Epidemiol. Biomarkers Prev. 2005, 14, 2898–2904. [Google Scholar] [CrossRef]
- Zhang, C.X.; Ho, S.C.; Fu, J.H.; Cheng, S.Z.; Chen, Y.M.; Lin, F.Y. Dairy products, calcium intake, and breast cancer risk: A case-control study in China. Nutr. Cancer 2011, 63, 12–20. [Google Scholar]
- Chen, P.; Hu, P.; Xie, D.; Qin, Y.; Wang, F.; Wang, H. Meta-analysis of vitamin D, calcium and the prevention of breast cancer. Breast Cancer Res. Treat. 2010, 121, 469–477. [Google Scholar] [CrossRef]
- Berube, S.; Diorio, C.; Masse, B.; Hebert-Croteau, N.; Byrne, C.; Cote, G.; Pollak, M.; Yaffe, M.; Brisson, J. Vitamin D and calcium intakes from food or supplements and mammographic breast density. Cancer Epidemiol. Biomarkers Prev. 2005, 14, 1653–1659. [Google Scholar]
- Abbas, S.; Linseisen, J.; Chang-Claude, J. Dietary vitamin D and calcium intake and premenopausal breast cancer risk in a German case-control study. Nutr. Cancer 2007, 59, 54–61. [Google Scholar] [CrossRef]
- Giovannucci, E.; Rimm, E.B.; Wolk, A.; Ascherio, A.; Stampfer, M.J.; Colditz, G.A.; Willett, W.C. Calcium and fructose intake in relation to risk of prostate cancer. Cancer Res. 1998, 58, 442–447. [Google Scholar]
- Gao, X.; LaValley, M.P.; Tucker, K.L. Prospective studies of dairy product and calcium intakes and prostate cancer risk: A meta-analysis. J. Natl. Cancer Inst. 2005, 97, 1768–1777. [Google Scholar] [CrossRef]
- Allen, N.E.; Key, T.J.; Appleby, P.N.; Travis, R.C.; Roddam, A.W.; Tjonneland, A.; Johnsen, N.F.; Overvad, K.; Linseisen, J.; Rohrmann, S.; et al. Animal foods, protein, calcium and prostate cancer risk: The European prospective investigation into cancer and nutrition. Br. J. Cancer 2008, 98, 1574–1581. [Google Scholar] [CrossRef]
- Huncharek, M.; Muscat, J.; Kupelnick, B. Dairy products, dietary calcium and vitamin D intake as risk factors for prostate cancer: A meta-analysis of 26,769 cases from 45 observational studies. Nutr. Cancer 2008, 60, 421–441. [Google Scholar]
- Kristal, A.R.; Arnold, K.B.; Neuhouser, M.L.; Goodman, P.; Platz, E.A.; Albanes, D.; Thompson, I.M. Diet, supplement use, and prostate cancer risk: Results from the prostate cancer prevention trial. Am. J. Epidemiol. 2010, 172, 566–577. [Google Scholar] [CrossRef]
- Williams, C.D.; Whitley, B.M.; Hoyo, C.; Grant, D.J.; Schwartz, G.G.; Presti, J.C., Jr.; Iraggi, J.D.; Newman, K.A.; Gerber, L.; Taylor, L.A.; et al. Dietary calcium and risk for prostate cancer: A case-control study among US veterans. Prev. Chronic Dis. 2012, 9, E39. [Google Scholar]
- Wang, L.; Manson, J.E.; Song, Y.; Sesso, H.D. Systematic review: Vitamin D and calcium supplementation in prevention of cardiovascular events. Ann. Intern. Med. 2010, 152, 315–323. [Google Scholar]
- Wang, L.; Manson, J.E.; Sesso, H.D. Calcium intake and risk of cardiovascular disease: A review of prospective studies and randomized clinical trials. Am. J. Cardiovasc. Drugs 2012, 12, 105–116. [Google Scholar] [CrossRef]
- Davies, K.M.; Heaney, R.P.; Recker, R.R.; Lappe, J.M.; Barger-Lux, M.J.; Rafferty, K.; Hinders, S. Calcium intake and body weight. J. Clin. Endocrinol. Metab. 2000, 85, 4635–4638. [Google Scholar] [CrossRef]
- Zemel, M.B.; Shi, H.; Greer, B.; Dirienzo, D.; Zemel, P.C. Regulation of adiposity by dietary calcium. FASEB J. 2000, 14, 1132–1138. [Google Scholar]
- Puntus, T.; Schneider, B.; Meran, J.; Peterlik, M.; Kudlacek, S. Influence of age and gender on associations of body mass index with bone mineral density, bone turnover markers and circulating calcium-regulating and bone-active sex hormones. Bone 2011, 49, 824–829. [Google Scholar] [CrossRef]
- Pittas, A.G.; Lau, J.; Hu, F.B.; Dawson-Hughes, B. The role of vitamin D and calcium in type 2 diabetes. A systematic review and meta-analysis. J. Clin. Endocrinol. Metab. 2007, 92, 2017–2029. [Google Scholar] [CrossRef]
- McCarron, D.A.; Reusser, M.E. Finding consensus in the dietary calcium-blood pressure debate. J. Am. Coll. Nutr. 1999, 18, 398S–405S. [Google Scholar]
- Dobnig, H.; Pfeifer, M.; Begerow, B.; Suppan, K. Calcium, Not Vitamin D Decreases Blood Pressure Effectively in Elderly Subjects with Low Vitamin D Levels: A Randomized, Double-blind, Multi-center Study. In Abstracts of the 87th Annual Meeting of the Endocrine Society, San Diego, California, USA, 4–7 June 2005.
- Bostick, R.M.; Kushi, L.H.; Wu, Y.; Meyer, K.A.; Sellers, T.A.; Folsom, A.R. Relation of calcium, vitamin D, and dairy food intake to ischemic heart disease mortality among postmenopausal women. Am. J. Epidemiol. 1999, 149, 151–161. [Google Scholar] [CrossRef]
- Umesawa, M.; Iso, H.; Ishihara, J.; Saito, I.; Kokubo, Y.; Inoue, M.; Tsugane, S. Dietary calcium intake and risks of stroke, its subtypes, and coronary heart disease in Japanese: The JPHC study cohort I. Stroke 2008, 39, 2449–2456. [Google Scholar] [CrossRef]
- Al-Delaimy, W.K.; Rimm, E.; Willett, W.C.; Stampfer, M.J.; Hu, F.B. A prospective study of calcium intake from diet and supplements and risk of ischemic heart disease among men. Am. J. Clin. Nutr. 2003, 77, 814–818. [Google Scholar]
- Lewis, J.R.; Calver, J.; Zhu, K.; Flicker, L.; Prince, R.L. Calcium supplementation and the risks of atherosclerotic vascular disease in older women: Results of a 5-year RCT and a 4.5-year follow-up. J. Bone Miner. Res. 2011, 26, 35–41. [Google Scholar] [CrossRef]
- Pentti, K.; Tuppurainen, M.T.; Honkanen, R.; Sandini, L.; Kroger, H.; Alhava, E.; Saarikoski, S. Use of calcium supplements and the risk of coronary heart disease in 52–62-year-old women: The Kuopio osteoporosis risk factor and prevention study. Maturitas 2009, 63, 73–78. [Google Scholar] [CrossRef]
- Bolland, M.J.; Barber, P.A.; Doughty, R.N.; Mason, B.; Horne, A.; Ames, R.; Gamble, G.D.; Grey, A.; Reid, I.R. Vascular events in healthy older women receiving calcium supplementation: Randomised controlled trial. BMJ 2008, 336, 262–266. [Google Scholar] [CrossRef]
- Bolland, M.J.; Bacon, C.J.; Horne, A.M.; Mason, B.H.; Ames, R.W.; Wang, T.K.; Grey, A.B.; Gamble, G.D.; Reid, I.R. Vitamin D insufficiency and health outcomes over 5 y in older women. Am. J. Clin. Nutr. 2010, 91, 82–89. [Google Scholar]
- Nordin, B.E.; Nakane, M.; Ma, J.; Metcalfe, A.V. Effects of a weak case. BMJ 2006, 186, 20–28. [Google Scholar]
- Burckhardt, P. Potential negative cardiovascular effects of calcium supplements. Osteoporos. Int. 2011, 22, 1645–1647. [Google Scholar] [CrossRef]
- Samelson, E.J.; Booth, S.L.; Fox, C.S.; Tucker, K.L.; Wang, T.J.; Hoffmann, U.; Cupples, L.A.; O’Donnell, C.J.; Kiel, D.P. Calcium intake is not associated with increased coronary artery calcification: The Framingham study. Am. J. Clin. Nutr. 2012, 96, 1274–1280. [Google Scholar] [CrossRef]
- De Boer, I.H.; Kestenbaum, B.; Shoben, A.B.; Michos, E.D.; Sarnak, M.J.; Siscovick, D.S. 25-hydroxyvitamin D levels inversely associate with risk for developing coronary artery calcification. J. Am. Soc. Nephrol. 2009, 20, 1805–1812. [Google Scholar] [CrossRef]
- Mathew, S.; Lund, R.J.; Chaudhary, L.R.; Geurs, T.; Hruska, K.A. Vitamin D receptor activators can protect against vascular calcification. J. Am. Soc. Nephrol. 2008, 19, 1509–1519. [Google Scholar] [CrossRef]
- Wu-Wong, J.R.; Nakane, M.; Ma, J.; Ruan, X.; Kroeger, P.E. Effects of vitamin D analogs on gene expression profiling in human coronary artery smooth muscle cells. Atherosclerosis 2006, 186, 20–28. [Google Scholar] [CrossRef]
- Watson, K.E.; Abrolat, M.L.; Malone, L.L.; Hoeg, J.M.; Doherty, T.; Detrano, R.; Demer, L.L. Active serum vitamin D levels are inversely correlated with coronary calcification. Circulation 1997, 96, 1755–1760. [Google Scholar] [CrossRef]
- Brown, E.M.; Gamba, G.; Riccardi, D.; Lombardi, M.; Butters, R.; Kifor, O.; Sun, A.; Hediger, M.A.; Lytton, J.; Hebert, S.C. Cloning and characterization of an extracellular Ca2+-sensing receptor from bovine parathyroid. Nature 1993, 366, 575–580. [Google Scholar]
- Tfelt-Hansen, J.; Brown, E.M. The calcium-sensing receptor in normal physiology and pathophysiology: A review. Crit. Rev. Clin. Lab. Sci. 2005, 42, 35–70. [Google Scholar] [CrossRef]
- Brown, E.M.; Pollak, M.; Hebert, S.C. Sensing of extracellular Ca2+ by parathyroid and kidney cells: Cloning and characterization of an extracellular Ca2+-sensing receptor. Am. J. Kidney Dis. 1995, 25, 506–513. [Google Scholar] [CrossRef]
- Shalhoub, V.; Grisanti, M.; Padagas, J.; Scully, S.; Rattan, A.; Qi, M.; Varnum, B.; Vezina, C.; Lacey, D.; Martin, D. In vitro studies with the calcimimetic, cinacalcet HCl, on normal human adult osteoblastic and osteoclastic cells. Crit. Rev. Eukaryot. Gene Expr. 2003, 13, 89–106. [Google Scholar] [CrossRef]
- Hebert, S.C.; Cheng, S.; Geibel, J. Functions and roles of the extracellular Ca2+-sensing receptor in the gastrointestinal tract. Cell Calcium 2004, 35, 239–247. [Google Scholar] [CrossRef]
- Kallay, E.; Kifor, O.; Chattopadhyay, N.; Brown, E.M.; Bischof, M.G.; Peterlik, M.; Cross, H.S. Calcium-dependent c-myc proto-oncogene expression and proliferation of Caco-2 cells: A role for a luminal extracellular calcium-sensing receptor. Biochem. Biophys. Res. Commun. 1997, 232, 80–83. [Google Scholar] [CrossRef]
- Cheng, I.; Klingensmith, M.E.; Chattopadhyay, N.; Kifor, O.; Butters, R.R.; Soybel, D.I.; Brown, E.M. Identification and localization of the extracellular calcium-sensing receptor in human breast. J. Clin. Endocrinol. Metab. 1998, 83, 703–707. [Google Scholar] [CrossRef]
- McNeil, L.; Hobson, S.; Nipper, V.; Rodland, K.D. Functional calcium-sensing receptor expression in ovarian surface epithelial cells. Am. J. Obstet. Gynecol. 1998, 178, 305–313. [Google Scholar] [CrossRef]
- Sanders, J.L.; Chattopadhyay, N.; Kifor, O.; Yamaguchi, T.; Brown, E.M. Ca2+-sensing receptor expression and PTHrP secretion in PC-3 human prostate cancer cells. Am. J. Physiol. Endocrinol. Metab. 2001, 281, E1267–E1274. [Google Scholar]
- Racz, G.Z.; Kittel, A.; Riccardi, D.; Case, R.M.; Elliott, A.C.; Varga, G. Extracellular calcium sensing receptor in human pancreatic cells. Gut 2002, 51, 705–711. [Google Scholar] [CrossRef]
- Squires, P.E.; Harris, T.E.; Persaud, S.J.; Curtis, S.B.; Buchan, A.M.; Jones, P.M. The extracellular calcium-sensing receptor on human beta-cells negatively modulates insulin secretion. Diabetes 2000, 49, 409–417. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Olozak, I.; Chattopadhyay, N.; Butters, R.R.; Kifor, O.; Scadden, D.T.; Brown, E.M. Expression of extracellular calcium (Ca2+)o-sensing receptor in human peripheral blood monocytes. Biochem. Biophys. Res. Commun. 1998, 246, 501–506. [Google Scholar] [CrossRef]
- Molostvov, G.; James, S.; Fletcher, S.; Bennett, J.; Lehnert, H.; Bland, R.; Zehnder, D. Extracellular calcium-sensing receptor is functionally expressed in human artery. Am. J. Physiol. Renal Physiol. 2007, 293, F946–F955. [Google Scholar] [CrossRef]
- Yano, S.; Brown, E.M.; Chattopadhyay, N. Calcium-sensing receptor in the brain. Cell Calcium 2004, 35, 257–264. [Google Scholar] [CrossRef]
- Chattopadhyay, N.; Brown, E.M. Calcium-Sensing Receptor; Kluwer Academic Publishers: Boston, MA, USA, 2003. [Google Scholar]
- Chakravarti, B.; Chattopadhyay, N.; Brown, E.M. Signaling through the extracellular calcium-sensing receptor (CaSR). Adv. Exp. Med. Biol. 2012, 740, 103–142. [Google Scholar] [CrossRef]
- Maiti, A.; Hait, N.C.; Beckman, M.J. Extracellular calcium-sensing receptor activation induces vitamin D receptor levels in proximal kidney HK-2G cells by a mechanism that requires phosphorylation of p38 alpha MAPK. J. Biol. Chem. 2008, 283, 175–183. [Google Scholar]
- Garrett, J.E.; Tamir, H.; Kifor, O.; Simin, R.T.; Rogers, K.V.; Mithal, A.; Gagel, R.F.; Brown, E.M. Calcitonin-secreting cells of the thyroid express an extracellular calcium receptor gene. Endocrinology 1995, 136, 5202–5211. [Google Scholar] [CrossRef]
- Kapur, K.; Johnson, T.; Beckmann, N.D.; Sehmi, J.; Tanaka, T.; Kutalik, Z.; Styrkarsdottir, U.; Zhang, W.; Marek, D.; Gudbjartsson, D.F.; et al. Genome-wide meta-analysis for serum calcium identifies significantly associated SNPs near the calcium-sensing receptor (CaSR) gene. PLoS Genet. 2010, 6, e1001035. [Google Scholar] [CrossRef]
- Dvorak, M.M.; Siddiqua, A.; Ward, D.T.; Carter, D.H.; Dallas, S.L.; Nemeth, E.F.; Riccardi, D. Physiological changes in extracellular calcium concentration directly control osteoblast function in the absence of calciotropic hormones. Proc. Natl. Acad. Sci. USA 2004, 101, 5140–5145. [Google Scholar]
- Chang, W.; Tu, C.; Chen, T.H.; Bikle, D.; Shoback, D. The extracellular calcium-sensing receptor (CaSR) is a critical modulator of skeletal development. Sci. Signal. 2008, 1, ra1. [Google Scholar] [CrossRef]
- Brown, E.M.; Lian, J.B. New insights in bone biology: Unmasking skeletal effects of the extracellular calcium-sensing receptor. Sci. Signal. 2008, 1, pe40. [Google Scholar] [CrossRef]
- Dvorak-Ewell, M.M.; Chen, T.H.; Liang, N.; Garvey, C.; Liu, B.; Tu, C.; Chang, W.; Bikle, D.D.; Shoback, D.M. Osteoblast extracellular Ca2+-sensing receptor regulates bone development, mineralization, and turnover. J. Bone Miner. Res. 2011, 26, 2935–2947. [Google Scholar] [CrossRef]
- Rodland, K.D. The role of the calcium-sensing receptor in cancer. Cell Calcium 2004, 35, 291–295. [Google Scholar]
- Saidak, Z.; Mentaverri, R.; Brown, E.M. The role of the calcium-sensing receptor in the development and progression of cancer. Endocr. Rev. 2009, 30, 178–195. [Google Scholar] [CrossRef]
- Brown, E.M.; Yang, J.J. Biochemistry and biology of the extracellular calcium-sensing receptor. Bone 2009, 44, S201–S202. [Google Scholar] [CrossRef]
- Magno, A.L.; Ward, B.K.; Ratajczak, T. The calcium-sensing receptor: A molecular perspective. Endocr. Rev. 2011, 32, 3–30. [Google Scholar] [CrossRef]
- Dvorak, M.M.; Riccardi, D. Ca2+ as an extracellular signal in bone. Cell Calcium 2004, 35, 249–255. [Google Scholar]
- Yamaguchi, T.; Kifor, O.; Chattopadhyay, N.; Brown, E.M. Expression of extracellular calcium (Ca2+)o-sensing receptor in the clonal osteoblast-like cell lines, UMR-106 and SAOS-2. Biochem. Biophys. Res. Commun. 1998, 243, 753–757. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Chattopadhyay, N.; Kifor, O.; Butters, R.R., Jr.; Sugimoto, T.; Brown, E.M. Mouse osteoblastic cell line (MC3T3-E1) expresses extracellular calcium (Ca2+)o-sensing receptor and its agonists stimulate chemotaxis and proliferation of MC3T3-E1 cells. J. Bone Miner. Res. 1998, 13, 1530–1538. [Google Scholar]
- Yamaguchi, T.; Chattopadhyay, N.; Kifor, O.; Brown, E.M. Extracellular calcium (Ca2+)o-sensing receptor in a murine bone marrow-derived stromal cell line (ST2): Potential mediator of the actions of (Ca2+)o on the function of ST2 cells. Endocrinology 1998, 139, 3561–3568. [Google Scholar] [CrossRef]
- Baron, R.; Rawadi, G. Targeting the Wnt/beta-catenin pathway to regulate bone formation in the adult skeleton. Endocrinology 2007, 148, 2635–2643. [Google Scholar] [CrossRef]
- Krause, C.; de Gorter, D.J.J.; Karperien, M.; ten Dijke, P. Signal transduction cascades controlling osteoblast differentiation. In Primer of the Metabolic Bone Diseases, 7th ed; American Society of Bone Mineral Research: Washington, DC, USA, 2008; pp. 10–14. [Google Scholar]
- Kohn, A.D.; Moon, R.T. Wnt and calcium signaling: β-catenin-independent pathways. Cell Calcium 2005, 38, 439–446. [Google Scholar] [CrossRef]
- Owen, T.A.; Aronow, M.S.; Barone, L.M.; Bettencourt, B.; Stein, G.S.; Lian, J.B. Pleiotropic effects of vitamin D on osteoblast gene expression are related to the proliferative and differentiated state of the bone cell phenotype: Dependency upon basal levels of gene expression, duration of exposure, and bone matrix competency in normal rat osteoblast cultures. Endocrinology 1991, 128, 1496–1504. [Google Scholar] [CrossRef]
- Sheinin, Y.; Kallay, E.; Wrba, F.; Kriwanek, S.; Peterlik, M.; Cross, H.S. Immunocytochemical localization of the extracellular calcium-sensing receptor in normal and malignant human large intestinal mucosa. J. Histochem. Cytochem. 2000, 48, 595–602. [Google Scholar] [CrossRef]
- Whitfield, J.F. Calcium, calcium-sensing receptor and colon cancer. Cancer Lett. 2009, 275, 9–16. [Google Scholar] [CrossRef]
- Bhagavathula, N.; Kelley, E.A.; Reddy, M.; Nerusu, K.C.; Leonard, C.; Fay, K.; Chakrabarty, S.; Varani, J. Upregulation of calcium-sensing receptor and mitogen-activated protein kinase signalling in the regulation of growth and differentiation in colon carcinoma. Br. J. Cancer 2005, 93, 1364–1371. [Google Scholar] [CrossRef]
- Rey, O.; Chang, W.; Bikle, D.; Rozengurt, N.; Young, S.H.; Rozengurt, E. Negative cross-talk between calcium-sensing receptor and beta-catenin signaling systems in colonic epithelium. J. Biol. Chem. 2012, 287, 1158–1167. [Google Scholar]
- Gama, L.; Baxendale-Cox, L.M.; Breitwieser, G.E. Ca2+-sensing receptors in intestinal epithelium. Am. J. Physiol. 1997, 273, C1168–C1175. [Google Scholar]
- Kallay, E.; Bajna, E.; Wrba, F.; Kriwanek, S.; Peterlik, M.; Cross, H.S. Dietary calcium and growth modulation of human colon cancer cells: Role of the extracellular calcium-sensing receptor. Cancer Detect. Prev. 2000, 24, 127–136. [Google Scholar]
- Hizaki, K.; Yamamoto, H.; Taniguchi, H.; Adachi, Y.; Nakazawa, M.; Tanuma, T.; Kato, N.; Sukawa, Y.; Sanchez, J.V.; Suzuki, H.; et al. Epigenetic inactivation of calcium-sensing receptor in colorectal carcinogenesis. Mod. Pathol. 2011, 24, 876–884. [Google Scholar] [CrossRef]
- Kallay, E.; Bonner, E.; Wrba, F.; Thakker, R.V.; Peterlik, M.; Cross, H.S. Molecular and functional characterization of the extracellular calcium-sensing receptor in human colon cancer cells. Oncol. Res. 2003, 13, 551–559. [Google Scholar]
- Kallay, E.; Bises, G.; Bajna, E.; Bieglmayer, C.; Gerdenitsch, W.; Steffan, I.; Kato, S.; Armbrecht, H.J.; Cross, H.S. Colon-specific regulation of vitamin D hydroxylases—A possible approach for tumor prevention. Carcinogenesis 2005, 26, 1581–1589. [Google Scholar] [CrossRef]
- Chakrabarty, S.; Wang, H.; Canaff, L.; Hendy, G.N.; Appelman, H.; Varani, J. Calcium sensing receptor in human colon carcinoma: Interaction with Ca2+ and 1,25-dihydroxyvitamin D3. Cancer Res. 2005, 65, 493–498. [Google Scholar]
- MacLeod, R.J.; Hayes, M.; Pacheco, I. Wnt5a secretion stimulated by the extracellular calcium-sensing receptor inhibits defective Wnt signaling in colon cancer cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 293, G403–G411. [Google Scholar] [CrossRef]
- Bhagavathula, N.; Hanosh, A.W.; Nerusu, K.C.; Appelman, H.; Chakrabarty, S.; Varani, J. Regulation of E-cadherin and β-catenin by Ca2+ in colon carcinoma is dependent on calcium-sensing receptor expression and function. Int. J. Cancer 2007, 121, 1455–1462. [Google Scholar] [CrossRef]
- Palmer, H.G.; Gonzalez-Sancho, J.M.; Espada, J.; Berciano, M.T.; Puig, I.; Baulida, J.; Quintanilla, M.; Cano, A.; de Herreros, A.G.; Lafarga, M.; et al. Vitamin D3 promotes the differentiation of colon carcinoma cells by the induction of E-cadherin and the inhibition of β-catenin signaling. J. Cell. Biol. 2001, 154, 369–387. [Google Scholar] [CrossRef]
- Almquist, M.; Manjer, J.; Bondeson, L.; Bondeson, A.G. Serum calcium and breast cancer risk: Results from a prospective cohort study of 7847 women. Cancer Causes Control 2007, 18, 595–602. [Google Scholar] [CrossRef]
- El Hiani, Y.; Ahidouch, A.; Roudbaraki, M.; Guenin, S.; Brule, G.; Ouadid-Ahidouch, H. Calcium-sensing receptor stimulation induces nonselective cation channel activation in breast cancer cells. J. Membr. Biol. 2006, 211, 127–137. [Google Scholar] [CrossRef]
- Roderick, H.L.; Cook, S.J. Ca2+ signalling checkpoints in cancer: Remodelling Ca2+ for cancer cell proliferation and survival. Nat. Rev. Cancer 2008, 8, 361–375. [Google Scholar] [CrossRef]
- Curry, M.C.; Luk, N.A.; Kenny, P.A.; Roberts-Thomson, S.J.; Monteith, G.R. Distinct regulation of cytoplasmic calcium signals and cell death pathways by different plasma membrane calcium ATPase isoforms in MDA-MB-231 breast cancer cells. J. Biol. Chem. 2012, 287, 28598–28608. [Google Scholar]
- Promkan, M.; Liu, G.; Patmasiriwat, P.; Chakrabarty, S. BRCA1 suppresses the expression of survivin and promotes sensitivity to paclitaxel through the calcium sensing receptor (CaSR) in human breast cancer cells. Cell Calcium 2011, 49, 79–88. [Google Scholar] [CrossRef]
- Mathiasen, I.S.; Sergeev, I.N.; Bastholm, L.; Elling, F.; Norman, A.W.; Jaattela, M. Calcium and calpain as key mediators of apoptosis-like death induced by vitamin D compounds in breast cancer cells. J. Biol. Chem. 2002, 277, 30738–30745. [Google Scholar]
- Leclercq, G. Calcium-induced activation of estrogen receptor α—New insight. Steroids 2012, 77, 924–927. [Google Scholar] [CrossRef]
- Sanders, J.L.; Chattopadhyay, N.; Kifor, O.; Yamaguchi, T.; Butters, R.R.; Brown, E.M. Extracellular calcium-sensing receptor expression and its potential role in regulating parathyroid hormone-related peptide secretion in human breast cancer cell lines. Endocrinology 2000, 141, 4357–4364. [Google Scholar] [CrossRef]
- Yano, S.; Macleod, R.J.; Chattopadhyay, N.; Tfelt-Hansen, J.; Kifor, O.; Butters, R.R.; Brown, E.M. Calcium-sensing receptor activation stimulates parathyroid hormone-related protein secretion in prostate cancer cells: Role of epidermal growth factor receptor transactivation. Bone 2004, 35, 664–672. [Google Scholar] [CrossRef]
- Liao, J.; Schneider, A.; Datta, N.S.; McCauley, L.K. Extracellular calcium as a candidate mediator of prostate cancer skeletal metastasis. Cancer Res. 2006, 66, 9065–9073. [Google Scholar] [CrossRef]
- Shui, I.M.; Mucci, L.A.; Wilson, K.M.; Kraft, P.; Penney, K.L.; Stampfer, M.J.; Giovannucci, E. Common genetic variation of the calcium sensing receptor and lethal prostate cancer risk. Cancer Epidemiol. Biomarkers Prev. 2012, 22, 118–126. [Google Scholar]
- Smajilovic, S.; Yano, S.; Jabbari, R.; Tfelt-Hansen, J. The calcium-sensing receptor and calcimimetics in blood pressure modulation. Br. J. Pharmacol. 2011, 164, 884–893. [Google Scholar] [CrossRef]
- Torres, P.A.; de Broe, M. Calcium-sensing receptor, calcimimetics, and cardiovascular calcifications in chronic kidney disease. Kidney Int. 2012, 82, 19–25. [Google Scholar] [CrossRef]
- Tfelt-Hansen, J.; Hansen, J.L.; Smajilovic, S.; Terwilliger, E.F.; Haunso, S.; Sheikh, S.P. Calcium receptor is functionally expressed in rat neonatal ventricular cardiomyocytes. Am. J. Physiol. Heart Circ. Physiol. 2006, 290, H1165–H1171. [Google Scholar]
- Weston, A.H.; Geraghty, A.; Egner, I.; Edwards, G. The vascular extracellular calcium-sensing receptor: An update. Acta Physiol. 2011, 203, 127–137. [Google Scholar] [CrossRef]
- Wang, R.; Xu, C.; Zhao, W.; Zhang, J.; Cao, K.; Yang, B.; Wu, L. Calcium and polyamine regulated calcium-sensing receptors in cardiac tissue. Eur. J. Biochem. 2003, 270, 2680–2688. [Google Scholar] [CrossRef]
- Zhong, X.; Liu, J.; Lu, F.; Wang, Y.; Zhao, Y.; Dong, S.; Leng, X.; Jia, J.; Ren, H.; Xu, C.; et al. Calcium sensing receptor regulates cardiomyocyte function through nuclear calcium. Cell Biol. Int. 2012, 36, 937–943. [Google Scholar] [CrossRef]
- Hofer, A.M.; Curci, S.; Doble, M.A.; Brown, E.M.; Soybel, D.I. Intercellular communication mediated by the extracellular calcium-sensing receptor. Nat. Cell Biol. 2000, 2, 392–398. [Google Scholar] [CrossRef]
- Molostvov, G.; Bland, R.; Zehnder, D. Expression and role of the calcium-sensing receptor in the blood vessel wall. Curr. Pharm. Biotechnol. 2009, 10, 282–288. [Google Scholar] [CrossRef]
- Alam, M.U.; Kirton, J.P.; Wilkinson, F.L.; Towers, E.; Sinha, S.; Rouhi, M.; Vizard, T.N.; Sage, A.P.; Martin, D.; Ward, D.T.; et al. Calcification is associated with loss of functional calcium-sensing receptor in vascular smooth muscle cells. Cardiovasc. Res. 2009, 81, 260–268. [Google Scholar]
- Atchison, D.K.; Beierwaltes, W.H. The influence of extracellular and intracellular calcium on the secretion of renin. Pflugers Arch. 2012. [Google Scholar] [CrossRef]
- Wang, D.; An, S.J.; Wang, W.H.; McGiff, J.C.; Ferreri, N.R. CaR-mediated COX-2 expression in primary cultured MTAl cells. Am. J. Physiol. Renal Physiol. 2001, 281, F658–F664. [Google Scholar]
- Abdullah, H.I.; Pedraza, P.L.; McGiff, J.C.; Ferreri, N.R. Calcium-sensing receptor signaling pathways in medullary thick ascending limb cells mediate COX-2-derived PGE2 production: Functional significance. Am. J. Physiol. Renal Physiol. 2008, 295, F1082–F1089. [Google Scholar] [CrossRef]
- Gamba, G.; Friedman, P.A. Thick ascending limb: The Na+:K+:2Cl− co-transporter, NKCC2, and the calcium-sensing receptor, CasR. Pflugers Arch. 2009, 458, 61–76. [Google Scholar] [CrossRef]
- Vargas-Poussou, R.; Huang, C.; Hulin, P.; Houillier, P.; Jeunemaitre, X.; Paillard, M.; Planelles, G.; Dechaux, M.; Miller, R.T.; Antignac, C. Functional characterization of a calcium-sensing receptor mutation in severe autosomal dominant hypocalcemia with a Bartter-like syndrome. J. Am. Soc. Nephrol. 2002, 13, 2259–2266. [Google Scholar] [CrossRef]
- Peterlik, M. Vitamin D insufficiency and chronic diseases: Hype and reality. Food Funct. 2012, 3, 784–794. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Peterlik, M.; Kállay, E.; Cross, H.S. Calcium Nutrition and Extracellular Calcium Sensing: Relevance for the Pathogenesis of Osteoporosis, Cancer and Cardiovascular Diseases. Nutrients 2013, 5, 302-327. https://doi.org/10.3390/nu5010302
Peterlik M, Kállay E, Cross HS. Calcium Nutrition and Extracellular Calcium Sensing: Relevance for the Pathogenesis of Osteoporosis, Cancer and Cardiovascular Diseases. Nutrients. 2013; 5(1):302-327. https://doi.org/10.3390/nu5010302
Chicago/Turabian StylePeterlik, Meinrad, Enikoe Kállay, and Heide S. Cross. 2013. "Calcium Nutrition and Extracellular Calcium Sensing: Relevance for the Pathogenesis of Osteoporosis, Cancer and Cardiovascular Diseases" Nutrients 5, no. 1: 302-327. https://doi.org/10.3390/nu5010302