Ergogenic Effects of β-Alanine and Carnosine: Proposed Future Research to Quantify Their Efficacy

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Experimental Section
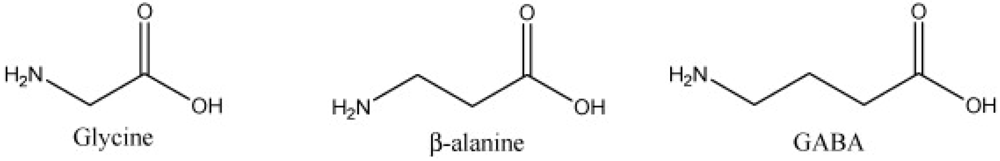
2.1. β-Alanine

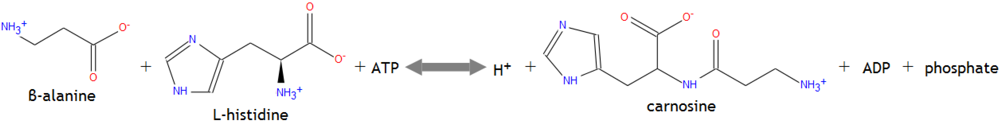
2.2. Carnosine
Carnosine and Exercise-Induced Fatigue
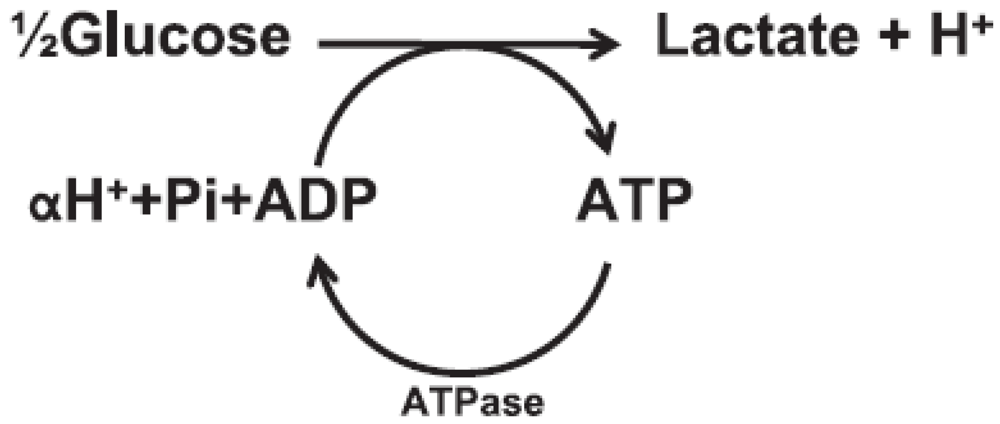
 (2)
(2)2.3. The Impact of Exercise and Dietary Interventions on Carnosine Concentrations
Oral β-alanine as an Ergogenic Aid to Exercise Performance
3. Recommended β-Alanine Dosages
4. Precautions
5. Implications/Future Recommendations
Conflict of Interest
References
- Artoli, G.G.; Gualano, B.; Smith, A.; Stout, J.; Lancha, A.H. Role of β-alanine supplementation on muscle carnosine and exercise performance. Med. Sci. Sports Exerc. 2009, 42, 1162–1173. [Google Scholar]
- Derave, W.; Everaert, I.; Beckman, S.; Baguet, A. Muscle carnosine metabolism and β-alanine supplementation in relation to exercise and training. Sports Med. 2010, 40, 247–263. [Google Scholar] [CrossRef]
- Hobson, R.M.; Saunders, B.; Ball, G.; Harris, R.C.; Sale, C. Effects of β-alanine supplementation on exercise performance: A meta-analysis. Amino Acids 2012. [Google Scholar] [CrossRef]
- Sale, C.; Saunders, B.; Harris, R.C. Effect of beta-alanine supplementation on muscle carnosine concentrations and exercise performance. Amino Acids 2010, 39, 321–333. [Google Scholar] [CrossRef]
- Culbertson, J.Y.; Kreider, R.B.; Greenwood, M.; Cooke, M. Effects ofβ-alanine on muscle carnsoine and exercise performance: A review of the current literature. Nutrients 2010, 2, 75–98. [Google Scholar] [CrossRef] [Green Version]
- Stout, J.R.; Graves, B.S.; Smith, A.E.; Hartman, M.J.; Cramer, J.T.; Beck, T.W.; Harris, R.C. The effect of beta-alanine supplementation on neuromuscular fatigue in elderly (55-92 Years): A double-blind randomized study. J. Int. Soc. Sports Nutr. 2008, 5. [Google Scholar] [CrossRef]
- Gojkovic, Z.; Sandrini, M.P.; Piskur, J. Eukaryotic β-alanine synthases are functionally related but have a high degree of structural diversity. Genetics 2001, 158, 999–1011. [Google Scholar]
- Suzuki, Y.; Nakao, T.; Maemura, H.; Sato, M.; Kamahara, K.; Morimatsu, F.; Takamatsu, K. Carnosine and anserine ingestion enhances contribution of nonbicarbonate buffering. Med. Sci. Sports Exerc. 2006, 38, 334–338. [Google Scholar]
- Suzuki, Y.; Osamu, I.; Mukai, N.; Takahashi, H.; Takamatsu, K. High level of skeletal muscle carnosine contributes to the letter half of exercise performance during 30-s maximal cycle ergometer sprinting. Jpn. J. Physiol. 2002, 52, 199–205. [Google Scholar] [CrossRef]
- Tiedje, K.E.; Stevens, K.; Barnes, S.; Weaver, D.F. Alanine as a small molecule neurotransmitter. Neurochem. Int. 2010, 57, 177–188. [Google Scholar] [CrossRef]
- Sterlingwerff, T.; Decombaz, J.; Harris, R.C.; Boesch, C. Optimizing human in vivo dosing and delivery of β-alanine supplements for muscle carnosine synthesis. Amino Acids 2012. [Google Scholar] [CrossRef]
- Baguet, A.; Bourgois, J.; Vanhee, L.; Achten, E.; Derave, W. Important role of muscle carnosine in rowing performance. J. Appl. Physiol. 2010, 109, 1096–1101. [Google Scholar] [CrossRef]
- Harris, R.C.; Wise, J.A.; Price, K.A.; Kim, H.J.; Kim, C.K.; Sale, C. Determinants of muscle carnosine content. Amino Acids 2012, 43, 5–12. [Google Scholar] [CrossRef]
- Kendrick, I.P.; Kim, H.J.; Harris, R.C.; Kim, C.K.; Dang, V.H.; Lam, T.Q.; Bui, T.T.; Wise, J.A. The effect of 4 weeks β-alanine supplementation and isokinetic training on carnosine concentrations in type I and II human skeletal muscle fibres. Eur. J. Appl. Physiol. 2009, 106, 131–138. [Google Scholar] [CrossRef]
- Bakardjiev, A.; Bauer, K. Transport of beta-alanine and biosynthesis of carnoisne by skeletal cells in primary culture. Eur. J. Biochem. 1994, 225, 617–623. [Google Scholar] [CrossRef]
- Bhardwaj, R.K.; Herrera-Ruiz, D.; Eltoukhy, N.; Saad, M.; Knipp, G.T. The functional evaluation of human peptide/histidine transports 1 (hPHT1) in transiently transfected COS-7 cells. Eur. J. Pharm. Sci. 2006, 27, 533–542. [Google Scholar] [CrossRef]
- Jappar, D.; Hu, Y.; Keep, R.F.; Smith, D.E. Transport mechanisms of carnosine in SKPT cells contribution of apical and basolateral membrane transporters. Pharm. Res. 2009, 26, 172–181. [Google Scholar] [CrossRef]
- Harris, R.C.; Marlin, D.J.; Dunnett, M. Muscle buffering capacity and dipepetide content in the thoroughbred horse, greyhound dog and man. Comp. Biochem. Physiol. A 1990, 97, 249–251. [Google Scholar]
- Baguet, A.; Reyngoudt, H.; Pottier, A.; Everaert, I.; Callens, S.; Achtem, E.; Derave, W. Carnosine loading and washout in human skeletal muscles. J. Appl. Physiol. 2009, 106, 837–842. [Google Scholar] [CrossRef]
- Derave, W.; Özdemir, M.S.; Harris, R.C.; Pottier, A.; Reyngoudt, H.; Koppo, K.; Wise, J.A.; Achten, E. β-Alanine supplementation augments muscle carnosine content and attenuates fatigue during repeated isokinetic contraction bouts in trained sprinters. J. Appl. Physiol. 2007, 103, 1736–1743. [Google Scholar] [CrossRef]
- Dunnett, M.; Harris, R.C.; Soliman, M.Z.; Surwar, A.A. Carnosine, anserine and taurine contents in individual fibres from the middle gluteal muscle of the camel. Res. Vet. Sci. 1997, 62, 213–216. [Google Scholar] [CrossRef]
- Tallon, M.J.; Harris, R.C.; Boobis, L.H.; Fallowfield, J.L.; Wise, J.A. The carnosine content of vastus laterials is elevated in resistance-trained bodybuilders. J. Strength Cond. Res. 2005, 19, 725–729. [Google Scholar]
- Penafiel, R.; Ruzafa, C.; Monserrat, F.; Cremades, A. Gender-related differences in carnosine, anserine and lysine content of murine skeletal muscle. Amino Acids 2004, 26, 53–58. [Google Scholar] [CrossRef]
- Boldyrev, A.A. Does carnosine possess direct anti-oxidant activity? Int. J. Biochem. 1993, 25, 1101–1107. [Google Scholar] [CrossRef]
- Harris, R.C.; Tallon, M.J.; Dunnett, M.; Boobis, L.; Coakley, J.; Kim, H.J.; Fallowfield, J.L.; Hill, C.A.; Sale, C.; Wise, J.A. The absorption of orally supplied β-alanine and its effect on muscle carnosine synthesis in human vastus lateralis. Amino Acids 2006, 30, 279–289. [Google Scholar] [CrossRef]
- Hipkiss, A.R. On the enigma of carnosine’s anti-aging action. Exp. Gerontol. 2008, 44, 237–242. [Google Scholar] [CrossRef]
- Hipkiss, A.R.; Michaelis, J.; Syrris, P. Non-enzymatic glycosylation of the dipeptide L-carnosine, a potential anti-protein-cross-linking agent. FEBS Lett. 1995, 371, 81–85. [Google Scholar] [CrossRef]
- Nagasawa, T.; Yonekura, T.; Nishizawa, N.; Kitts, D.D. In vitro and in vivo inhibition of muscle lipid and protein oxidation by carnosine. Mol. Cell Biochem. 2001, 225, 29–34. [Google Scholar] [CrossRef]
- Trombley, P.Q.; Horning, M.S.; Blakemore, L.J. Interactions between carnosine and zinc and copper: Implications for neuromodulation and neuroprotection. Biochemistry (Mosc.) 2000, 65, 807–816. [Google Scholar]
- Hoffman, J.; Ratames, N.A.; Ross, R.; Kang, J.; Magrell, J.; Neese, K.; Faigenbaum, A.D.; Wise, J.A. Alanine and the hormonal response to exercise. Int. J. Sports Med. 2008, 29, 952–958. [Google Scholar] [CrossRef]
- Böning, D.; Strobel, G.; Beneke, R.; Maassen, N. Lactic acid still remains the real cause of exercise-induced metabolic acidosis. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 289, R902–R903. [Google Scholar] [CrossRef]
- Stallknecht, B.; Vissing, J.; Galbo, H. Lactate production and clearance in exercise. Effects of training. A mini-review. Scand. J. Med. Sci. Sports 1998, 8, 127–131. [Google Scholar]
- Green, H.J. Mechanisms of muscle fatigue in intense exercise. J. Sports Sci. 1997, 15, 247–256. [Google Scholar] [CrossRef]
- Cady, E.B.; Jones, D.A.; Lynn, J.; Newham, D.J. Changes in force and intracellular metabolites during fatigue of human skeletal muscle. J. Physiol. 1989, 418, 311–325. [Google Scholar]
- Cairns, S.P. Lactic acid and exercise performance: Culprit or friend? Sports Med. 2006, 36, 279–291. [Google Scholar] [CrossRef]
- Mannion, A.F.; Jakeman, P.M.; Dunnett, M.; Harris, R.C.; Willian, P.L. Carnosine and anserine concentrations in the quadriceps femoris muscle of healthy humans. Eur. J. Appl. Physiol. 1992, 68, 356–361. [Google Scholar]
- Robergs, R.A. Nothing “evil” and no “conundrum” about muscle lactate production. Exp. Physiol. 2011, 96, 1097–1098. [Google Scholar] [CrossRef]
- Fitts, R.H. Cellular mechanisms of muscle fatigue. Physiol. Rev. 1994, 74, 49–94. [Google Scholar] [CrossRef]
- Medbo, J.O.; Tabata, I. Anaerobic energy release in working muscles during 30 s to 3 min of exhaustive bicycling. J. Appl. Physiol. 1993, 75, 1654–1660. [Google Scholar]
- Sahlin, K.; Harris, R.C.; Nylind, B. Lactate content and pH in muscle samples obtained after dynamic exercise. Pflügers Arch. 1976, 367, 143–149. [Google Scholar] [CrossRef]
- Gladden, L.B. Lactate metabolism: A new paradigm for the third millennium. J. Physiol. 2004, 558, 5–30. [Google Scholar] [CrossRef]
- Gollnick, P.D.; Korge, P.; Karpakka, J.; Saltin, B. Elongation of skeletal muscle relaxation during exercise is linked to reduced calcium uptake by the sarcoplasmic reticulum in man. Acta Physiol. Scand. 1991, 142, 135–136. [Google Scholar] [CrossRef]
- Bate Smith, E.C. The buffering of muscle in rigor; protein, phosphate and carnosine. J. Physiol. 1938, 92, 336–343. [Google Scholar]
- Sweeney, K.M.; Wright, G.A.; Brice, A.G.; Doberstein, S.T. The effect of β-alanine supplementation on power performance during repeated sprint activity. J. Strength Cond. Res. 2010, 24, 79–87. [Google Scholar] [CrossRef]
- Dutka, T.L.; Lamboley, C.R.; McKenna, M.J.; Murphy, R.M.; Lamb, G.D. Effects of carnosine on contractile apparatus Ca2+-sensitivity and sarcoplasmic reticulum Ca2+ release in human skeletal muscle fibers. J. Appl. Physiol. 2011, 112, 728–736. [Google Scholar]
- Suzuki, Y.; Ito, O.; Takahashi, H.; Takamasu, K. The effect of sprint training on skeletal muscle carnosine in humans. Int. J. Sports Health Sci. 2004, 2, 105–110. [Google Scholar] [CrossRef]
- Harris, R.C. Muscle carnosine elevation with supplementation and training, and the effects of elevation on exercise performance. In Presented at the 2nd Annual International Society of Sports Nutrition Conference, New Orleans, LA, USA, 16-18 June 2005; pp. 39–40.
- Hill, C.A.; Harris, R.C.; Kim, H.J.; Boobis, L.; Sale, C.; Wise, J.A. The effect of beta-alanine and creatine monohydrate supplementation on muscle composition and exercise performance. Med. Sci. Sports Exerc. 2005, 37, S348. [Google Scholar]
- Zoeller, R.F.; Stout, J.R.; O’Kroy, J.A.; Torok, D.J.; Mielke, M. Effects of 28 days of beta-alanine and creatine monohydrate supplementation on anaerobic power, ventilatory and lactate thresholds, and time to exhaustion. Amino Acids 2007, 33, 505–510. [Google Scholar] [CrossRef]
- Hill, C.A.; Harris, R.C.; Kim, H.J.; Harris, B.D.; Sale, C.; Boobis, L.H.; Kim, C.K.; Wise, J.A. Influence of β-alanine supplementation on skeletal muscle carnosine concentrations and high intensity cycling capacity. Amino Acids 2007, 32, 225–233. [Google Scholar] [CrossRef]
- Kendrick, I.P.; Kim, H.J.; Harris, R.C.; Kim, C.K.; Dang, V.H.; Lam, T.Q.; Bui, T.T.; Smith, M.; Wise, J.A. The effects of 10 weeks of resistance training combined with β-alanine supplementation on whole body strength, force production, muscular endurance and body composition. Amino Acids 2008, 34, 547–554. [Google Scholar] [CrossRef]
- Del Favero, S.; Roschel, H.; Solis, M.Y.; Hayashi, A.P.; Artoli, G.G.; Otadut, M.C.; Benatti, F.B.; Harris, R.C.; Wise, J.A.; Leite, C.C.; et al. Beta-alanine (Carnosyn™) supplementation in elderly subjects (60-80 years): Effects on muscle carnosine content and physical capacity. Amino Acids 2012, 43, 49–56. [Google Scholar] [CrossRef]
- Bellinger, P.M.; Howe, S.T.; Shing, C.M.; Fell, J.W. The effect of combined β-alanine and NaHCO3 supplementation on cycling performance. Med. Sci. Sports Exerc. 2012, in press. [Google Scholar]
- Spradley, B.D.; Crowley, K.R.; Tai, C.Y.; Kendall, K.L.; Fukuda, D.H.; Esposito, E.N.; Moon, S.E.; Moon, J.R. Ingesting a pre-workout supplement containing caffeine, B-vitamins, amino acids, creatine, and beta-alanine before exercise delays fatigue while improving reaction time and muscular endurance. Nutr. Metab. 2012, 9. Available online: http://www.nutritionandmetabolism.com/content/9/1/28 (accessed on 1 June 2012). [CrossRef]
- Kern, B.D.; Robinson, T.L. Effects of β-alanine supplementation on performance and body composition in college wrestlers and football players. J. Strength Cond. Res. 2011, 25, 1804–1815. [Google Scholar] [CrossRef]
- Jordan, T.; Lukaszuk, J.; Misic, M.; Umoren, J. Effect of beta-alanine supplementation on the onset of blood lactate accumulation (OBLA) during treadmill running: Pre/post 2 treatment experimental design. Int. Soc. Sports Nutr. 2010, 7. Available online: http://www.jissn.com/content/7/1/20 (accessed on 1 June 2012). [CrossRef]
- Saunders, B.; Sale, C.; Harris, R.C.; Sunderland, C. Effect of beta-alanine supplementation on repeated sprint performance during the Loughborough Intermittent Shuttle Test. Amino Acids 2012. [Google Scholar] [CrossRef]
- Jagim, A.R.; Wright, G.A.; Brice, A.G.; Doberstein, S.T. Effects of beta-alanine supplementation on sprint endurance. J. Strength. Cond. Res. 2012, in press. [Google Scholar]
- Baguet, A.; Koppo, K.; Pottier, A.; Derave, W. Alanine supplementation reduces acidosis but not oxygen uptake response during high-intensity cycling exercise. Eur. J. Appl. Physiol. 2010, 108, 495–503. [Google Scholar] [CrossRef]
- Stout, J.R.; Cramer, J.T.; Mielke, M.; O’Kroy, J.; Torok, D.J.; Zoeller, R.F. Effects of twenty-eight days of beta-alanine and creatine monohydrate supplementation on the physical work capacity at neuromuscular fatigue threshold. J. Strength Cond. Res. 2006, 20, 928–931. [Google Scholar]
- Goto, K.; Maemura, H.; Takamatsu, K.; Ishii, N. Hormonal responses to resistance exercise after ingestion of carnosine and anserine. J. Strength Cond. Res. 2011, 25, 398–405. [Google Scholar] [CrossRef]
- Van Thienen, R.; van Proeyen, K.; Vanden Eynde, B.; Puype, J.; Lefere, T.; Hespel, P. Alanine improves sprint performance in endurance cycling. Med. Sci. Sports Exerc. 2009, 41, 898–903. [Google Scholar] [CrossRef]
- Stout, J.R.; Cramer, J.T.; Zoeller, R.F.; Torok, D.; Costa, P.; Hoffman, J.R.; Harris, R.C. Effects of beta-alanine supplementation on the onset of neuromuscular fatigue and ventilatory threshold in women. Amino Acids 2007, 32, 381–386. [Google Scholar] [CrossRef]
- Batrukova, M.A.; Rubtsov, A.M. Histidine-containing dipeptides as endogenous regulators of the activity of sarcoplasmic reticulum Ca-release channels. Biochim. Biophys. Acta 1324, 142–150. [Google Scholar]
- Dutka, T.L.; Lamb, G.D. Effect of carnosine on excitation-contraction coupling in mechanically-skinned rat skeletal muscle. J. Muscle Res. Cell Motil. 2004, 25, 203–213. [Google Scholar] [CrossRef]
- Boldyrev, A.A.; Dupin, A.M.; Pindel, E.V.; Severin, S.E. Antioxidative properties of histidine-containing dipeptides from skeletal muscles of vertebrates. Comp. Biochem. Physiol. B 1988, 89, 245–250. [Google Scholar] [CrossRef]
- Ririe, D.G.; Roberts, P.R.; Shouse, M.N.; Zaloga, G.P. Vasodilatory actions of the dietary peptide carnosine. Nutrition 2000, 16, 168–172. [Google Scholar] [CrossRef]
- Walter, A.A.; Smith, A.E.; Kendall, K.L.; Stout, J.R.; Cramer, J.T. Six weeks of high-intensity interval training with and without β-alanine supplementation for improving cardiovascular fitness in women. J. Strength Cond. Res. 2010, 24, 1199–1207. [Google Scholar] [CrossRef]
- Freund, H.; Gendry, P. Lactate kinetics after short strenuous exercise in man. Eur. J. Appl. Physiol. 1978, 39, 123–135. [Google Scholar] [CrossRef]
- Freund, H.; Oyono-Enguelle, S.; Heitz, A.; Marbach, J.; Ott, C.; Gartner, M. Effect of exercise duration on lactate kinetics after short muscular exercise. Eur. J. Appl. Physiol. 1989, 58, 534–542. [Google Scholar] [CrossRef]
- Oyono-Enguelle, S.; Gartner, M.; Marbach, J.; Heitz, A.; Ott, C.; Freund, H. Comparison of arterial and venous blood lactate kinetics after short exercise. Int. J. Sports Med. 1989, 10, 16–24. [Google Scholar] [CrossRef]
- Beneke, R.; Hütler, M.; Jung, M.; Leithäuser, R.M. Modeling the blood lactate kinetics at maximal short-term exercise conditions in children, adolescents, and adults. J. Appl. Physiol. 2005, 99, 499–504. [Google Scholar] [CrossRef]
- Beneke, R.; Jumah, M.D.; Leithäuser, R.M. Modelling the lactate response to short-term all out exercise. Dyn. Med. 2007, 6. [Google Scholar] [CrossRef]
- Severin, S.E.; Kirzon, M.V.; Kaftanova, T.M. Effect of carnosine and anserine on action of isolated frog muscles. Dokl. Akad. Nauk. SSR 1953, 91, 691–701. [Google Scholar]
- Crozier, R.A.; Ajit, S.K.; Kaftan, E.J.; Pausch, M.H. MrgD activation inhibits KCNQ/M-currents and contributes to enhanced neuronal excitability. J. Neurosci. 2007, 27, 4492–4496. [Google Scholar] [CrossRef]
- Decombaz, J.; Beaumont, M.; Vuichoud, J.; Bouisset, F.; Sterlingwerff, T. Effect of slow-release β-alanine tablets on absorption kinetics and paraesthesia. Amino Acids 2011. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Caruso, J.; Charles, J.; Unruh, K.; Giebel, R.; Learmonth, L.; Potter, W. Ergogenic Effects of β-Alanine and Carnosine: Proposed Future Research to Quantify Their Efficacy. Nutrients 2012, 4, 585-601. https://doi.org/10.3390/nu4070585
Caruso J, Charles J, Unruh K, Giebel R, Learmonth L, Potter W. Ergogenic Effects of β-Alanine and Carnosine: Proposed Future Research to Quantify Their Efficacy. Nutrients. 2012; 4(7):585-601. https://doi.org/10.3390/nu4070585
Chicago/Turabian StyleCaruso, John, Jessica Charles, Kayla Unruh, Rachel Giebel, Lexis Learmonth, and William Potter. 2012. "Ergogenic Effects of β-Alanine and Carnosine: Proposed Future Research to Quantify Their Efficacy" Nutrients 4, no. 7: 585-601. https://doi.org/10.3390/nu4070585