European Bats as Carriers of Viruses with Zoonotic Potential

Centre for Biological Threats and Special Pathogens, Robert Koch Institute, Nordufer 20, 13353 Berlin, Germany
*
Author to whom correspondence should be addressed.
Viruses 2014, 6(8), 3110-3128; https://doi.org/10.3390/v6083110
Submission received: 17 April 2014
/
Revised: 29 July 2014
/
Accepted: 30 July 2014
/
Published: 13 August 2014
(This article belongs to the Special Issue Viruses and Bats)
Abstract
:Bats are being increasingly recognized as reservoir hosts of highly pathogenic and zoonotic emerging viruses (Marburg virus, Nipah virus, Hendra virus, Rabies virus, and coronaviruses). While numerous studies have focused on the mentioned highly human-pathogenic bat viruses in tropical regions, little is known on similar human-pathogenic viruses that may be present in European bats. Although novel viruses are being detected, their zoonotic potential remains unclear unless further studies are conducted. At present, it is assumed that the risk posed by bats to the general public is rather low. In this review, selected viruses detected and isolated in Europe are discussed from our point of view in regard to their human-pathogenic potential. All European bat species and their roosts are legally protected and some European species are even endangered. Nevertheless, the increasing public fear of bats and their viruses is an obstacle to their protection. Educating the public regarding bat lyssaviruses might result in reduced threats to both the public and the bats.
1. Introduction
The European continent is inhabited by 52 bat species; some are threatened with extinction on the population level and are hence protected under the International Union for Conservation of Nature (IUCN) Red List of Threatened Species and the Convention on the Conservation of Migratory Species of Wild Animals [1,2]. The European bat species that inhabit colder regions use torpor and hibernation. Many bat species migrate over vast distances while others are rather territorial. All bats in Europe utilize echolocation to navigate. Contrary to the worldwide efforts in protecting bats, they have been increasingly gaining attention as potential reservoir hosts of some of the most virulent viruses we know. Various publications reviewed bats globally as carriers and potential reservoir hosts of human-pathogenic and zoonotic viruses [3,4,5,6,7,8,9,10], while hardly anything is known about human-pathogenicity of European bat viruses apart from lyssaviruses. In this review, we discuss a selection of viruses as possible threats posed by European bats to the public from our point of view. A summary of viruses that have been detected in European bats is given in Table 1 at the end of the manuscript. A more comprehensive and up-to-date list of bat-associated viruses can be found online at the Database of Bat-associated Viruses (DBatVir) [11].
European bat lyssaviruses (family Rhabdoviridae) are the most important zoonotic bat-borne viruses in Europe and have been comprehensively reviewed by Banyard et al. in this special issue on bat viruses (Title: Lyssavirus Infections of Bats: Emergence and Zoonotic Threat) [12]. Therefore, we will provide a short overview.

{kind=link}
{kind=link}
| Virus Family | Genus | Bat Species | Origin | Detection | Refs. |
|---|---|---|---|---|---|
| Adenoviridae | Mastadenovirus | Pipistrellus nathusii Pipistrellus pipistrellus | Germany | Isolation PCR | [13,14] |
| Nyctalus noctula Rhinolophus ferrumequinum | Hungary | PCR | [15] | ||
| Myotis myotis | Germany | PCR | [16] | ||
| Astroviridae | Mamastrovirus | Myotis myotis | Germany | PCR | [16] |
| Mamastrovirus | Myotis daubentonii Myotis bechsteinii Plecotus auritus | Hungary | PCR | [17] | |
| Borna-viridae | unclassified | Myotis nattereri Pipistrellus pipistrellus | France | Metagenomics | [18] |
| Bunyaviridae | Phlebovirus Toscana virus | Pipistrellus kuhlii | Italy | Isolation | [19] |
| Nairovirus | Myotis mystacinus | France | Metagenomics | [18] | |
| Coronaviridae | Alphacoronavirus | Myotis bechsteinii Myotis dasycneme Myotis daubentonii Pipistrellus nathusii Pipistrellus pygmaeus | Germany | PCR | [20] |
| Myotis blythii Myotis daubentonii Myotis myotis Mineropterus schreibersii Nyctalus lasiopterus Pipistrellus kuhlii Pipistrellus sp. | Spain | PCR | [21] | ||
| Hypsugo savii Nyctalus noctula Pipistrellus kuhlii Pipistrellus spp. Rhinolophus hipposideros | Italy | PCR | [22] | ||
| Miniopterus schreibersii Nyctalus leisleri Rhinolophus euryale Rhinolophus blasii Rhinolophus ferrumequinum Rhinolophus mehelyi | Germany | PCR | [16] | ||
| Myotis daubentonii Myotis nattereri | United Kingdom | PCR | [23] | ||
| Betacoronavirus | Miniopterus schreibersii Nyctalus leisleri Myotis daubentonii Rhinolophus euryale Rhinolophus blasii Rhinolophus ferrumequinumv Rhinolophus mehelyi Rhinolophus hipposideros | Bulgaria Germany | PCR | [24] | |
| Rhinolophus hipposideros | Slovenia | PCR | [25] | ||
| Pipistrellus nathusii | Ukraine | PCR | [26] | ||
| Pipistrellus pipistrellus | Netherlands | PCR | [27] | ||
| Eptesicus isabellinus Hypsugo savii | Spain | PCR | [21] | ||
| Eptesicus serotinus Hypsugo savii Nyctalus noctula Pipistrellus kuhlii Pipistrellus sp. Rhinolophus hipposideros | Italy | PCR | [22,28] | ||
| Filoviridae | Cuevovirus | Miniopterus schreibersii | Spain | PCR | [29] |
| Hepeviruses | Hep-E-related viruses | Eptesicus serotinus Myotis bechsteinii Myotis daubentonii | Germany Bulgaria | PCR | [30] |
| Herpesviridae | Betaherpesvirus Gammaherpesvirus | Myotis myotis Myotis nattereri Nyctalus noctula Pipistrellus pipistrellus Plecotus auritus | Germany | PCR | [31] |
| Betaherpesvirus Alphaherpesvirus | Rousettus aegyptiacus | Hungary | PCR | [15] | |
| Gammaherpesvirus | Eptesicus serotinus | Hungary | PCR | [32] | |
| Papillomavirus | Papillomavirus | Eptesicus serotinus Rhinolophus ferrumequinum | Spain | PCR | [33] |
| Paramyxoviridae | Unassigned | Myotis mystacinus Nyctalus noctula Pipistrellus pipistrellus | Germany | PCR | [34] |
| Morbillivirus | Myotis bechsteinii Myotis daubentonii Myotis myotis Myotis mystacinus Myotis alcathoe Myotis capaccinii | Bulgaria Germany Romania | PCR | [35] | |
| Reoviridae | Orthoreovirus | Myotis mystacinus Nyctalus noctula Pipistrellus pipistrellus Pipistrellus nathusii Pipistrellus kuhlii Plecotus auritus | Germany | IsolationPCR | [36] |
| Pipistrellus kuhlii Rhinolophus hipposideros Nyctalus noctula Tadarida teniotis Nyctalus noctula | Italy | Isolation PCR | [37] | ||
| Rotavirus | Myotis mystacinus | France | Metagenomics | [18] | |
| Retrovirus | Gammaretrovirus | Eptesicus serotinus | France | Metagenomics | [18] |
| Rhabdoviridae | Various European bat lyssaviruses | Eptesicus serotinus Eptesicus isabellinus Hypsugo savii Minopterus schreibersii Myotis myotis Myotis daubentonii Myotis dasycneme Myotis nattereri Plectorus auritus Pipistrellus pipistrellus Rhinolophus ferrumequinum Rousettus aegyptiacus Vespertilio murinus unclassified Chiroptera | Denmark
France Finland Germany Hungary Netherlands Poland Slovakia Spain Switzerland Ukraine United Kingdom | Microscopy isolation PCR | [33,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54] |
2. Molecular Detection of European Bat Viruses
The postulates drafted by Jacob Henle and Robert Koch in the late 19th century constitute a framework regarding the principles of cause-and-effect in microbiology [55]. Back then, it was comparatively straightforward to limit cause-and-effect to four postulates, although viruses had not yet been discovered nor was molecular biology developed (Table 1). All of the postulates are hard to fulfill for viruses, as they do not grow on nutrient media, but require living cells for replication. When looking for viruses on a molecular level, it is necessary to consider that only the first postulate can be accomplished. Studies identifying a host-pathogen relationship solely at the molecular level do not take into consideration that detection does not equal etiology. Even though polymerase chain reaction (PCR) screening and metagenomic studies are indispensable and valuable tools, virologists should stay close to the Henle-Koch postulates when assuming a possible virulence of viruses detected in bat hosts.
2.1. European Bat Coronaviruses
A plethora of coronaviruses has been detected in bats, mostly belonging to the alpha- and betacoronaviruses [11,56]. The genus alphacoronavirus hosts human-pathogenic strains (i.e., Human CoV 229E and NL63); however, in this review, we will focus on selected highly human-pathogenic betacoronaviruses and their European bat virus relatives [56].
From November 2002 until July 2003 the world was confronted with the first pandemic of the new millennium, caused by a novel coronavirus (CoV) inducing the Severe Acute Respiratory Syndrome in humans (SARS) [57,58,59]. The pandemic spread from its origin, a wet-market in the Guangdong province in China, through 33 countries on five continents resulted in more than 8000 infected humans of whom more than 700 eventually died [60,61]. The search for the animal reservoir began, identifying masked palm civets and bats as possible sources. Subsequently, a plethora of diverse coronaviruses of distinct groups have been detected in various bat species around the world via molecular-biological techniques. In 2012, another human-pathogenic coronavirus, called Middle East respiratory syndrome coronavirus (MERS-CoV), began spreading from the Arabian Peninsula, so far resulting in globally 707 laboratory-confirmed cases of infection with MERS-CoV, including at least 252 deaths [62]. Dromedaries and bats are suspected as reservoirs for MERS-CoV [63]. Recent findings support the plausibility of dromedaries as reservoir species [64].
Although numerous studies in European bats report the presence of SARS-like-CoV and MERS-like-CoV sequences [21,24,25,26,65], no final conclusion can be drawn regarding their zoonotic potential. A related virus detected in bats cannot necessarily be considered as zoonotic. A few alterations in the SARS-CoV spike protein enabled its binding to the host receptor ACE-2, thus SARS-CoV became capable of infecting humans [66]. So far, the SARS-like CoV detected in European bats lack these alterations and thus are not predicted to be capable of infecting humans. Although virus strains might be similar or related on a nucleic acid level, the distinct function of proteins is crucial when determining the host range. Therefore, mere similarity is not sufficient to examine the potential of viruses to infect humans or even predict their virulence. It took ten years from the emergence of SARS-CoV for the first bat CoV to be isolated from Rhinolophus bats in China, that displayed the human ACE-2 receptor, which enabled the virus to infect human cells [67]. These findings provide evidence for the reservoir theory. From the European perspective, nevertheless, no SARS-like CoV or MERS-like CoV has been isolated from any European bat, nor has any transmission of SARS-like CoV or MERS-like CoV to humans been reported. The case of MERS-CoV is slightly different, as a sequence of 190 base pairs with 100% identity to MERS-CoV was detected in a bat (Taphozous perforates—the species identification performed was not beyond doubt, as it was based on exclusion criteria (no Cytochrome b sequence of Taphozous perforates is available in GenBank [68])) in Saudi Arabia [8]. This finding initiated a controversy among leading CoV experts, as the journal Nature recently reported [69]. They discussed that the complete genome sequence of MERS-CoV obtained from the bat should confirm that the virus was indeed identical and not coincidentally just a short conserved region of the virus genome. Furthermore, a prevalence study might provide insights into the distribution of MERS-CoV in bat populations. Although Taphozous perforates are not abundant in Europe, climate change and environmental factors may have an effect on the future distribution of this bat species (Figure 1) [70]. The case of MERS-CoV emergence impressively demonstrates the necessity of virus discovery and prevalence studies. With the first sequence of MERS-CoV that became available, bats were suspected as reservoir hosts, not only because MERS-CoV is a SARS-CoV relative, but also because previous bat virus discovery studies had provided eligible sequences of bat CoV to GenBank, allowing for correlations with the novel MERS-CoV. Recently, a quasi-species of MERS CoVs was recovered from nasal swabs of dromedaries of the Kingdom of Saudi Arabia [64]. The MERS CoV consensus genome variants from dromedaries and humans are indistinguishable, supporting the plausibility of dromedaries in the role of transmission [64].
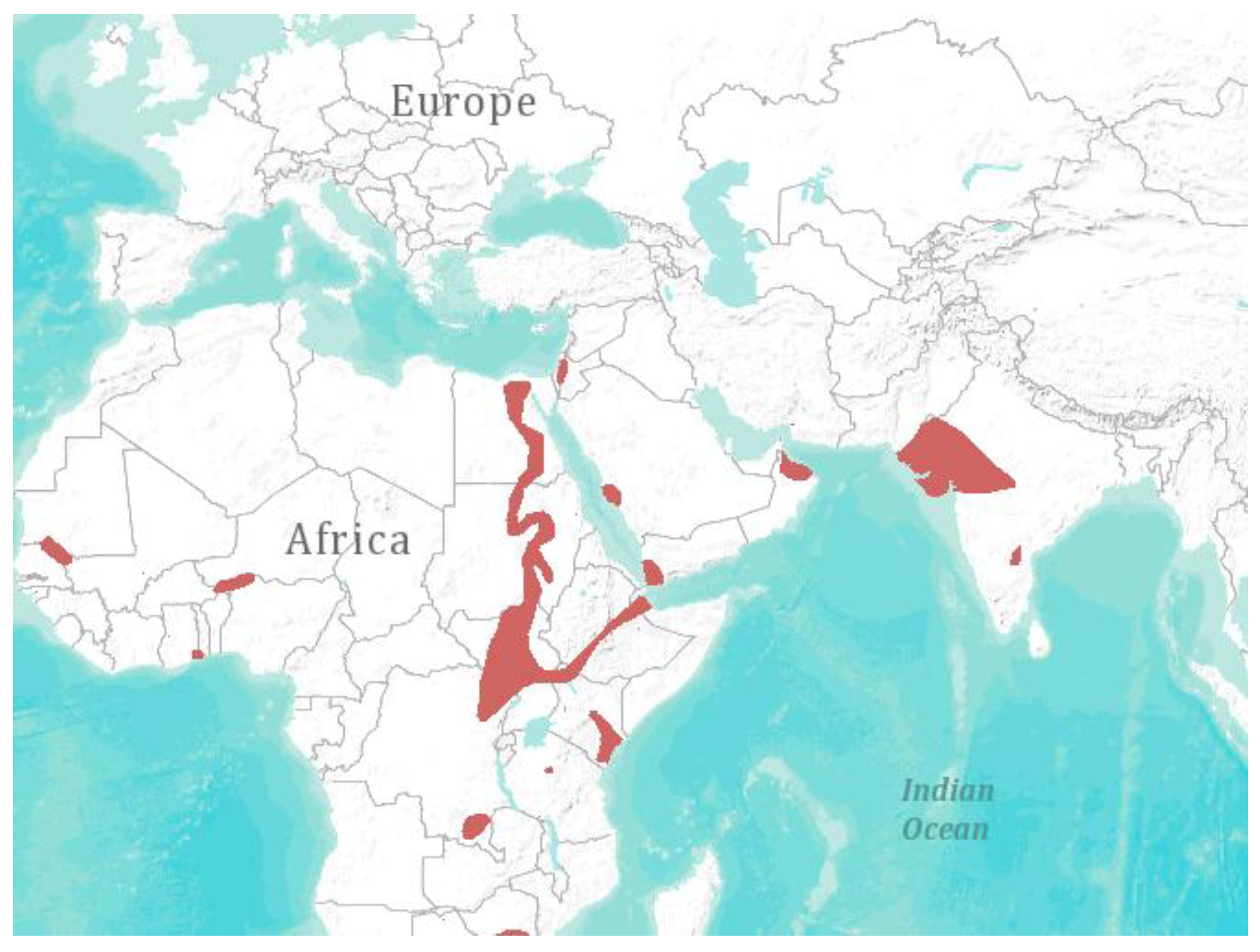
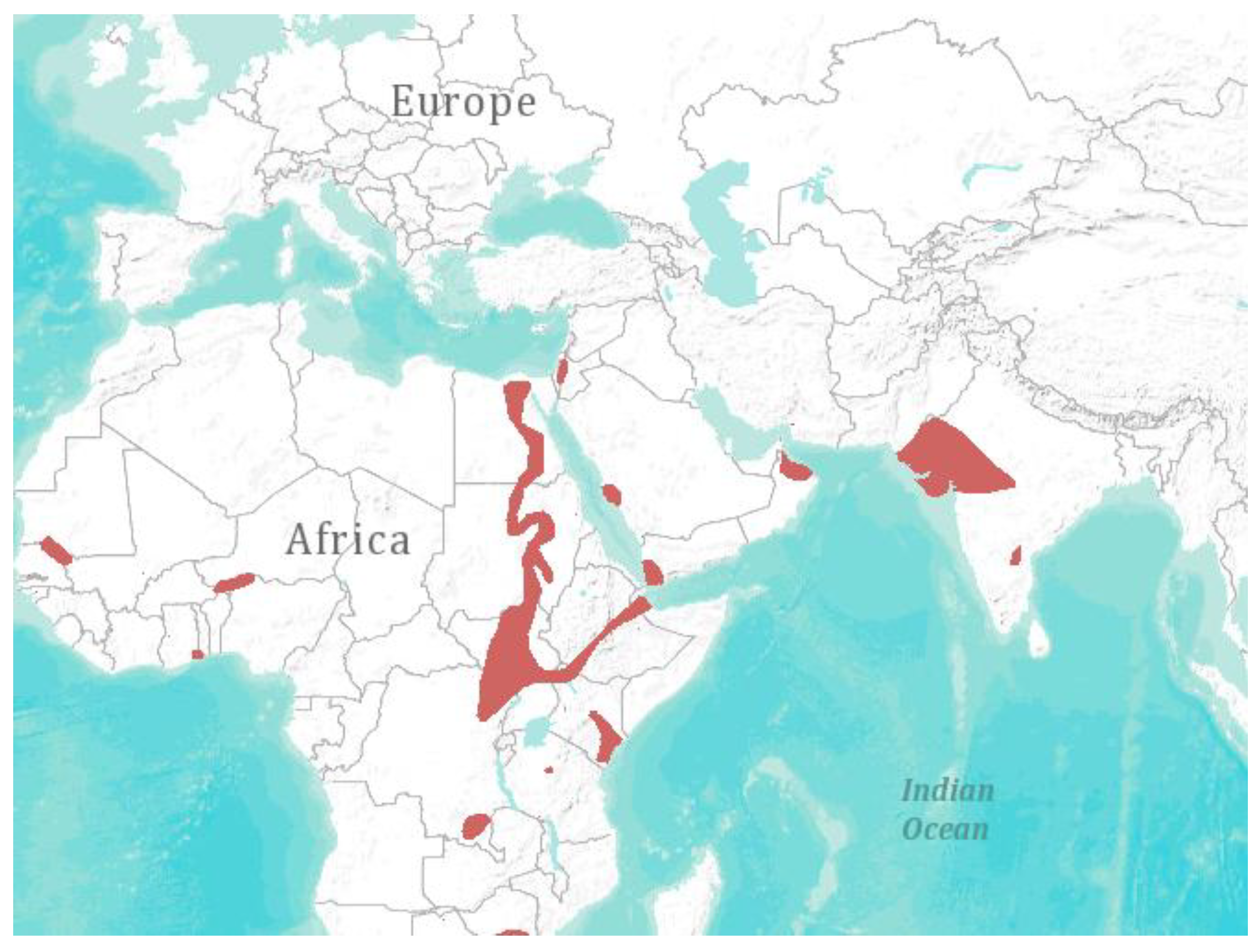
Figure 1.
Geographical distribution of Taphozous perforates [1]. Visualization: ArcGIS Explorer.1:50.000.000 [71] (Map sources: Esri, U.S. Geological Survey (USGS), National Oceanic and Atmospheric Administration (NOAA). Sources: Esri, DeLorme, USGS, NPS. Spatial data source: International Union for Conservation of Nature (IUCN)).
Figure 1.
Geographical distribution of Taphozous perforates [1]. Visualization: ArcGIS Explorer.1:50.000.000 [71] (Map sources: Esri, U.S. Geological Survey (USGS), National Oceanic and Atmospheric Administration (NOAA). Sources: Esri, DeLorme, USGS, NPS. Spatial data source: International Union for Conservation of Nature (IUCN)).
2.2. European Bat Filoviruses
In 2002, the first reported outbreak of filovirus, named Lloviu virus (LLOV), in a European bat population occurred in France, Spain, and Portugal [29]. Several colonies of Schreiber’s bats (Miniopterus schreibersii) suddenly declined due to an unknown disease. LLOV was found in animals that showed signs of viral infection, but not in healthy bats co-roosting in the caves (Myotis myotis). LLOV is distinctly related to filoviruses found in African bats and was classified in 2013 as type species of the novel genus Cuevavirus [56]. Unfortunately, the lack of successful isolation of LLOV prohibits the experimental infection of Schreiber’s bats to clarify whether LLOV is the first filovirus capable of inducing disease in bats. This would challenge the hypothesis of bats as potential reservoir hosts for other filoviruses like Ebola and Marburg virus. Schreiber’s bats are distributed in distinct lineages throughout Oceania, Africa, Southern Europe, and South-East Asia (Figure 2) [72]. They are thought to transmit and maintain LLOV across different lineages throughout their habitats, although no studies are available to prove this hypothesis.
Consequently, the sole demonstration of a novel filovirus sequence does not provide evidence of a possible public health threat. Following the Henle-Koch postulates, the virus should be isolated and further characterized to draw conclusions on the evolution of filoviruses in their respective bat host. As most filoviruses are described as highly pathogenic for humans, the occurrence of LLOV should be carefully monitored by prevalence studies in the highly abundant Miniopterus schreibersii (Figure 2).
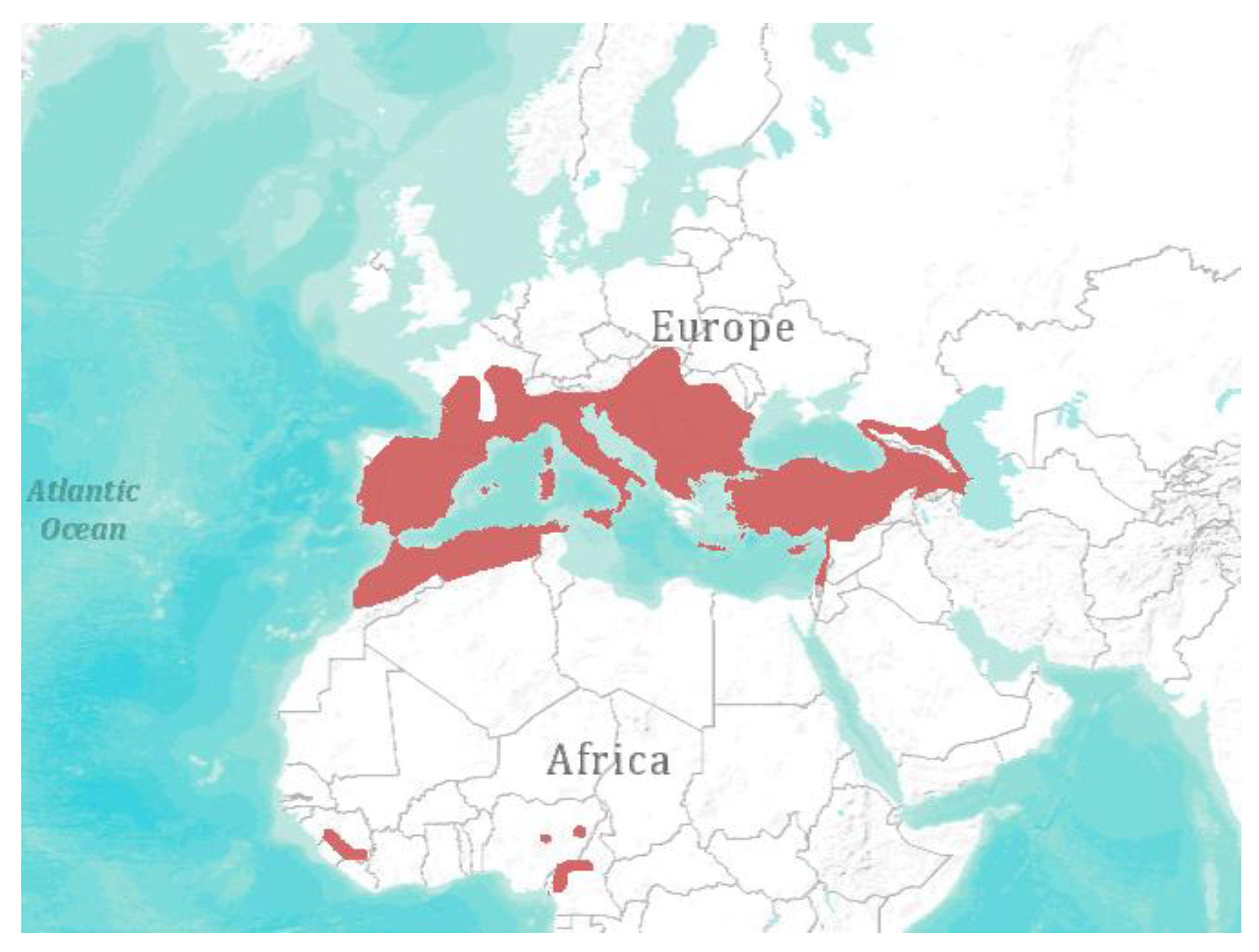
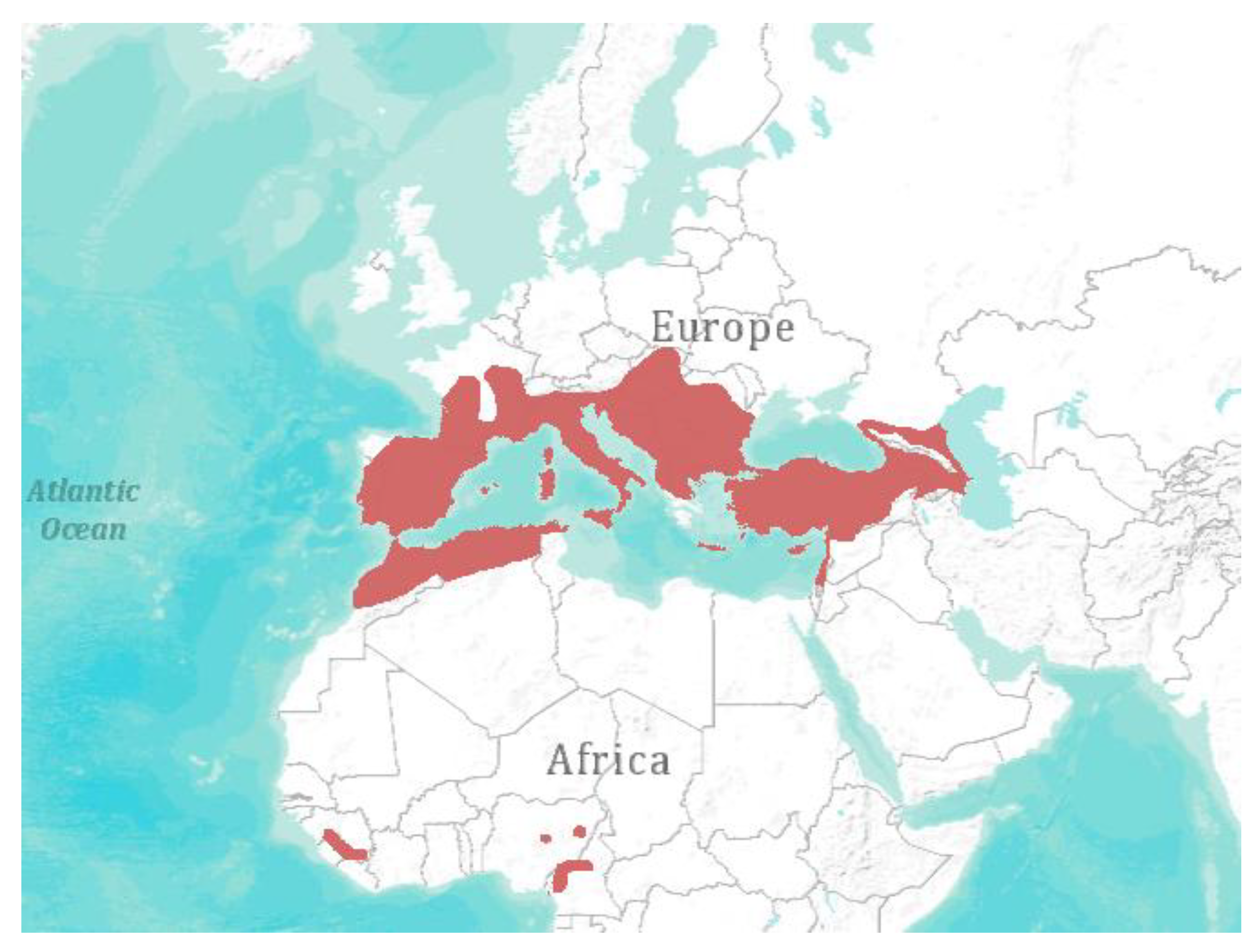
Figure 2.
Geographical distribution of Miniopterus schreibersii [1]. Visualization: ArcGIS Explorer.1:50.000.000 [71] (Map sources: Esri, U.S. Geological Survey (USGS), National Oceanic and Atmospheric Administration (NOAA). Sources: Esri, DeLorme, USGS, NPS. Spatial data source: International Union for Conservation of Nature (IUCN)).
Figure 2.
Geographical distribution of Miniopterus schreibersii [1]. Visualization: ArcGIS Explorer.1:50.000.000 [71] (Map sources: Esri, U.S. Geological Survey (USGS), National Oceanic and Atmospheric Administration (NOAA). Sources: Esri, DeLorme, USGS, NPS. Spatial data source: International Union for Conservation of Nature (IUCN)).
2.3. European Bat Paramyxoviruses
In 2012, three distinct paramyxoviruses were detected in German bats, two of which were related to the proposed genus Jeilongvirus (Myotis mystacinus, Pipistrellus pipistrellus) and one was related to the genus Rubulavirus (Nyctalus noctula) [34]. Another study published in the same year described another 12 different paramyxoviruses in bats from Germany (Myotis bechsteinii, M. daubentonii, M. myotis, and M. mystacinus) and Bulgaria (Myotis alcathoe and M. capaccinii), all of which belong to the genus Morbillivirus [35].
None of the novel bat paramyxoviruses are closely related to viruses of the highly pathogenic genus Henipavirus or other human-pathogenic paramyxoviruses [34,35]. There is no evidence to suggest that any of these novel paramyxoviruses are capable of infecting humans. Similar to the case of the LLOV filovirus, virus isolates and prevalence studies in both humans and bats could improve knowledge and clarify their zoonotic potential.
2.4. Other Human-Pathogenic Viruses
Few studies have documented the negative results from PCR testing of European bats for other human-pathogenic viruses. For instance following generic PCR screening for flavi-, hanta- and influenza-A viruses in 210 European bats in 2011 [73], testing of another 1369 Central European bats for influenza-A viruses [74] and testing 42 European bats for hepadnaviruses in 2013 did not lead to the detection of any viral nucleic acids [75]. PCR screening of 468 European bats for orthopoxviruses has not revealed any known or novel virus sequences [76].
3. European Bat Virus Isolates
So far, the only virus isolates (beside lyssaviruses) obtained from European bats are one bunyavirus, one adenovirus and 22 orthoreoviruses [13,19,36,37]. These represent the only isolates that would allow for further characterization and potential clarification of their zoonotic potential. Nevertheless, recombinant viruses, constructed on sequence information, are also valuable tools to study prevalence and pathogenicity in vitro.
3.1. European Bat Bunyavirus Isolate
Toscana virus (TOSV) was isolated from a bat’s brain in 1988, while simultaneously TOSV was isolated from sandflies in the laboratory [19]. As TOSV has never been reported in bats afterwards and no hemagglutination-inhibiting antibodies has been initially found in the bat, there is a reasonable chance that this TOSV isolation may have been a cross-contamination [77].
3.2. European Bat Adenovirus Isolate
Bat adenovirus 2 (Bat AdV-2) was isolated from a bat’s intestine in 2009 [13], and the whole genome was obtained and circumstantially analyzed [14,78]. Bat AdV-2 displays a monophyletic relationship to the adenoviruses of canids (CAdV). Moreover, open trading frames (ORF) in the Bat AdV-2 genome and the CAdV are identical and not present in other members of the mastadenoviruses. The closely related canine AdV contribute to the severe kennel cough syndrome in canids and show an unusually broad host range [79]. This provides evidence suggesting an ancestral inter-species transmission of mastadenoviruses between bats and canids. Like in the case of rabies virus, which is prevalent in both bats and terrestrial mammals (e.g., dogs, raccoons, skunks, and foxes) of the Americas, a continuing exchange and transmission between bats and canids or other terrestrial animals might be possible [80]. There is no evidence of a zoonotic potential of Bat AdV-2.
3.3. European Bat Reovirus Isolates
In 2012, three novel orthoreoviruses were isolated from Plecotus auritus and Myotis mystacinus in Germany [36]. A subsequent PCR screening obtained identical viral sequences also in other bat species: Pipistrellus pipistrellus, Pipistrellus nathusii, Pipistrellus kuhlii, and Nyctalus noctula. At the same time, a group in Italy detected further 19 orthoreoviruses in Myotis kuhlii, Rhinolophus hyposideros, Tadarida teniotis, and Vespertilio murinus [37].
Summing up the data for the reovirus isolates from Germany and Italy, a close relationship was revealed to the genus Mammalian Orthoreovirus (MRV), in particular to an orthoreovirus obtained from a dog (strain T3/D04) with hemorrhagic enteritis in Italy [36,37,81]. No ancestral relationship was assumed here, but rather an opportunistic “behavior” of the novel closely related MRVs, as they were detected in various different bat species. Moreover, the newly isolated MRVs are phylogenetically related to viruses capable of inducing severe meningitis in humans [82]. Recently, a study published by Steyer et al. described the detection of an MRV from a child hospitalized with acute gastroenteritis in Slovenia [83]. The causative agent was determined to be an MRV with the highest similarity of 98.4%–99.0% in the respective segments to a bat MRV (T3/Bat/Germany/342/08) [83]. This might indicate a human-pathogenic potential of strain T3/Bat/Germany/342/08. As the case of SARS-CoV has shown that even small changes in the genome are important for determining the host range, this has to be determined for the bat MRVs in further studies. Interestingly, no contact was reported between the infected child and bats, but contact to a domestic dog was assumed [83]. The isolated viruses will allow for a seroprevalence study (cross-reactivity and cross-neutralization with other strains) in humans, which shall be initiated to examine the prevalence of specific antibodies to Bat MRVs in Germany and Italy (where these viruses have been found) to clarify their zoonotic potential. This is especially interesting as Asian bat orthoreoviruses of the genus Pteropine Orthoreovirus have already been linked to potentially zoonotic respiratory diseases in humans [84,85].
3.4. European Bat Lyssaviruses
Rhabdoviruses of the genus Lyssavirus that have been detected in Europe are considerably harmful and truly zoonotic agents, inevitably causing the death of unvaccinated humans if not treated in time before onset of the rabies disease [86]. Even though bat-transmitted lyssaviruses have a fatality rate of virtually 100% and are suspected to be transmissible by bat biting and scratching, the reported total number of human fatalities in Europe is low (n = 2–5 since 1963) [86,87,88]. All described hosts of European bat lyssaviruses (EBLV-1 and EBLV-2) are synanthropic, hence sharing their habitats with humans [87]. EBLV-1 has been predominantly detected in Eptesicus serotinus and E. isabellinus in Europe, both living in buildings, roofs, and attics usually in the southern regions of Europe (E. serotinus until 55° North, E. isabellinus in southern Portugal—E. isabellinus is a North African population of E. serotinus that is controversially but not concludingly discussed as a novel species [1]), and male bats are reported to co-roost with multiple bat species [90]. EBLV-1 was also detected in V. murinus, M. schreibersii, M. myotis, M. nattereri, R. ferrumequinum, and T. teniotis. Whether these bat species constitute accidental hosts infected by spillover from co-roosting E. serotinus species, or whether they are additional reservoirs, has not yet been determined [38,39,40,41,91].
Two human cases described by Johnson et al. were confirmed as infected with EBLV-2, which is prevalent in European M. daubentonii and M. dasycneme [40,86]. M. daubentonii is prevalent in north-eastern Europe and is frequently found co-roosting with P. pipistrellus and M. nattereri, whereas M. dasycneme is found throughout Europe and in the Mediterranean, co-roosting with M. capaccinii. So far, none of the co-roosting bats have been reported to carry EBLV-2 [90]. However, spillover transmission to other animals (stone-marten, sheep, and cat) was described for EBLV-1 [92,93,94].
Overall, lyssaviruses prevalent in European bats pose a risk to public health, and preventive measures have already been implemented by many European countries for decades (e.g., surveillance, vaccination plans, and post exposure prophylaxis) [87]. Especially the high-risk occupational groups (i.e., bat workers, bat carers in bat bat hospitals) are at increased risk. However, lyssavirus prevalence in European bats is very difficult to determine and results are very heterogenic [40]. The lyssavirus prevalences are considerably low, but changes of behavior as a result of a lyssavirus infection may be more likely to bring bats into contact with humans. However, it is necessary to balance the risk with the total number of fatal human cases during the last 35 years (five cases in 590 million people living in greater Europe) [87]. Accordingly, the risk is relatively low and would probably fall to zero if people were educated appropriately. Direct contact (bites and scratches) with certain bat species might be risky and require post exposure prophylaxis. Only few of the European bat species are known to be reservoirs of EBLV-1 and EBLV-2, but all of the European species are endangered or close to extinction. Relocation or culling of bat colonies, in spite of being an obvious solution from the viewpoint of the general public, increases the risk of lyssavirus exposure and transmission and should not be considered [95]. Only education can channel public fear to avoid further threats to the bats and the general public.
4. Spatial Abundance and Biodiversity
Alexander von Humboldt discovered the latitudinal gradient in species diversity as early as 1799 [96]: The richness of species is subject to a global diversity gradient, abating from the species-rich tropics toward the higher latitudes [97]. Bats influence this gradient significantly. More than 1100 bat species have been described worldwide. Although they are abundant worldwide except for the polar regions, a steep diversity gradient is present from the tropics towards the poles [97,98,99,100]. Are fewer viruses prevalent in European bats because of the lower abundance of species in the more temperate Europe? And is the zoonotic risk posed by bats decreased accordingly?
Only few studies on the biogeography of microorganisms are available. These studies indicate that the latitudinal diversity gradient has either no or a top-down effect on microbial diversity [101,102,103,104,105]. Two studies hypothesized that the local diversity and dispersal of viruses is very high, though overall, the viral diversity is limited on the global scale [106,107]. Therefore, no assumptions can be made regarding the viral diversity in species abundant in temperate climates. As the total number of abundant species might not be essential, the change in biodiversity might play a role.
The effect of decline in biodiversity on the emergence of diseases is subject of numerous publications [108,109,110,111,112,113,114]. Basically, there are arguments in favor of two controversial theories; reduced biodiversity could either increase (dilution effect) or decrease the risk of disease transmission. For almost half of the zoonotic diseases that have newly emerged by spillover since 1940, a preceding change in land-use, agriculture, and wildlife hunting was reported [108]. All of the above-mentioned effects contribute to changes in biodiversity and increased contact situations between human and animal hosts, also in Europe. Once spillover in novel hosts has occurred, a high density of the novel host population eventually facilitates the establishment in the novel niche. Thus, human overpopulation and a decreased biodiversity might be mutual factors promoting the establishment of emerging infectious diseases.
In conclusion, the Baas Becking hypothesis from 1932 might still be appropriate: “Everything is everywhere, but the environment selects” [115].
5. Conclusive Remarks
Until now, lyssaviruses have been the only proven zoonotic viruses in European bats and may cause rabies in humans. However, only few bat species are known to transmit lyssaviruses in Europe, and the number of human cases is rather low. Nevertheless, education of the general public should be intensified to avoid easily preventable infections. Although viruses with zoonotic potential have been detected in European bats, no clear assumption can be made without further studies. Sero-prevalence studies should be conducted on the orthoreoviruses isolated from European bats, especially as a closely related virus was detected in a diseased child in Slovenia [83]. Other bat viruses detected by using molecular techniques should be isolated (e.g., MERS-like CoV or Bat Bunyavirus) to allow for characterization and follow-up sero-prevalence studies.
In general, bats are special reservoir hosts because of their biological features, long-time co-evolution and high diversity of viruses that can be found. Furthermore, there is neither a clearly decreased risk in the emergence of zoonotic viruses in temperate climates compared to the tropics nor a decreased risk in regions of lower biodiversity.
In conclusion, many drivers of emergence in the tropics also have validity in Europe. However, European bats are endangered species, and some of them are threatened by extinction. Although lyssaviruses are prevalent in European bats, and some viruses might have a zoonotic potential, the overall hazard for humans is comparably low. Moreover, the protection of bats (and any wildlife) will consecutively also protect the general public.
Acknowledgments
The authors are grateful to Ursula Erikli for copy-editing and the two anonymous reviewers for their valuable comments.
Author Contributions
Both authors reviewed the literature and wrote the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References and Notes
- International Union for Conservation of Nature and Natural Resources (IUCN). 2013 The IUCN Red List of Threatened Species. Version 2013.2. Available online: http://www.iucnredlist.org/ (accessed on 15 April 2014).
- CMS. Convention on the Conservation of Migratory Species of Wild Animals. Available online: http://www.cms.int/ (accessed on 15 April 2014).
- Calisher, C.H.; Childs, J.E.; Field, H.E.; Holmes, K.V.; Schountz, T. Bats: Important reservoir hosts of emerging viruses. Clin. Microbiol. Rev. 2006, 19, 531–545. [Google Scholar] [CrossRef] [PubMed]
- Calisher, C.H.; Holmes, K.V.; Dominguez, S.R.; Schountz, T.; Cryan, P. Bats prove to be rich reservoirs for emerging viruses. Microbe 2008, 3, 521–528. [Google Scholar]
- Kuzmin, I.V.; Bozick, B.; Guagliardo, S.A.; Kunkel, R.; Shak, J.R.; Tong, S.; Rupprecht, C.E. Bats, emerging infectious diseases, and the rabies paradigm revisited. Emerg. Health Threats J. 2011, 4, 7159. [Google Scholar] [PubMed]
- Smith, I.; Wang, L.-F. Bats and their virome: An important source of emerging viruses capable of infecting humans. Curr. Opin. Virol. 2013, 3, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Rupprecht, C.; Wang, L.F.; Real, L.A. Bat zoonoses: The realities. In Food Security in a Global Economy Veterinary Medicine and Public Health; Smith, G., Kelly, A.M., Eds.; University of Pennsylvania Press: Philadelphia, PA, USA, 2008; Chapter 13; p. 145. [Google Scholar]
- Wang, L.-F. Bats and viruses: A brief review. Virol. Sin. 2009, 24, 93–99. [Google Scholar] [CrossRef]
- Wang, L.-F.; Walker, P.J.; Poon, L.L.M. Mass extinctions, biodiversity and mitochondrial function: Are bats “special” as reservoirs for emerging viruses? Curr. Opin. Virol. 2011, 1, 649–657. [Google Scholar]
- Wong, S.; Lau, S.; Woo, P.; Yuen, K.Y. Bats as a continuing source of emerging infections in humans. Rev. Med. Virol. 2007, 17, 67–91. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Liu, B.; Yang, J.; Jin, Q. DBatVir: The database of bat-associated viruses. Database Oxford 2014, 2014. [Google Scholar] [CrossRef]
- Banyard, A.C.; Evans, J.S.; Luo, T.R.; Fooks, A.R. Lyssaviruses and Bats: Emergence and Zoonotic Threat. Viruses 2014, 6, 2974–2990. [Google Scholar]
- Sonntag, M.; Mühldorfer, K.; Speck, S.; Wibbelt, G.; Kurth, A. New adenovirus in bats, Germany. Emerg. Infect. Dis. 2009, 15, 2052. [Google Scholar] [CrossRef] [PubMed]
- Kohl, C.; Vidovszky, M.Z.; Mühldorfer, K.; Dabrowski, P.W.; Radonić, A.; Nitsche, A.; Wibbelt, G.; Kurth, A.; Harrach, B. Genome analysis of bat adenovirus 2: Indications of interspecies transmission. J. Virol. 2012, 86, 1888–1892. [Google Scholar] [CrossRef] [PubMed]
- Jánoska, M.; Vidovszky, M.; Molnár, V.; Liptovszky, M.; Harrach, B.; Benko, M. Novel adenoviruses and herpesviruses detected in bats. Vet. J. 2011, 189, 118–121. [Google Scholar] [CrossRef] [PubMed]
- Drexler, J.F.; Corman, V.M.; Wegner, T.; Tateno, A.F.; Zerbinati, R.M.; Gloza-Rausch, F.; Seebens, A.; Müller, M.A.; Drosten, C. Amplification of emerging viruses in a bat colony. Emerg. Infect. Dis. 2011, 17, 449–456. [Google Scholar] [CrossRef] [PubMed]
- Kemenesi, G.; Dallos, B.; Görföl, T.; Boldogh, S.; Estók, P.; Kurucz, K.; Oldal, M.; Németh, V.; Madai, M.; Bányai, K.; et al. Novel European lineages of bat astroviruses identified in Hungary. Acta Virol. 2014, 58, 95–98. [Google Scholar] [CrossRef] [PubMed]
- Dacheux, L.; Cervantes-Gonzalez, M.; Guigon, G.; Thiberge, J.-M.; Vandenbogaert, M.; Maufrais, C.; Caro, V.; Bourhy, H. A preliminary study of viral metagenomics of French bat species in contact with humans: Identification of new mammalian viruses. PLoS One 2014, 9, e87194. [Google Scholar] [CrossRef] [PubMed]
- Verani, P.; Ciufolini, M.G.; Caciolli, S.; Renzi, A.; Nicoletti, L.; Sabatinelli, G.; Bartolozzi, D.; Volpi, G.; Amaducci, L.; Coluzzi, M. Ecology of viruses isolated from sand flies in Italy and characterized of a new Phlebovirus (Arabia virus). Am. J. Trop. Med. Hyg. 1988, 38, 433–439. [Google Scholar] [PubMed]
- Gloza-Rausch, F.; Ipsen, A.; Seebens, A.; Göttsche, M.; Panning, M.; Drexler, J.F.; Petersen, N.; Annan, A.; Grywna, K.; Müller, M.; et al. Detection and prevalence patterns of group I coronaviruses in bats, northern Germany. Emerg. Infect. Dis. 2008, 14, 626–631. [Google Scholar] [CrossRef] [PubMed]
- Falcón, A.; Vázquez-Morón, S.; Casas, I.; Aznar, C.; Ruiz, G.; Pozo, F.; Perez-Breña, P.; Juste, J.; Ibáñez, C.; Garin, I.; et al. Detection of alpha and betacoronaviruses in multiple Iberian bat species. Arch. Virol. 2011, 156, 1883–1890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lelli, D.; Papetti, A.; Sabelli, C.; Rosti, E.; Moreno, A.; Boniotti, M.B. Detection of coronaviruses in bats of various species in Italy. Viruses 2013, 5, 2679–2689. [Google Scholar] [CrossRef] [PubMed]
- August, T.A.; Mathews, F.; Nunn, M.A. Alphacoronavirus detected in bats in the United Kingdom. Vector Borne Zoonotic Dis. 2012, 12, 530–533. [Google Scholar] [CrossRef] [PubMed]
- Drexler, J.F.; Gloza-Rausch, F.; Glende, J.; Corman, V.M.; Muth, D.; Goettsche, M.; Seebens, A.; Niedrig, M.; Pfefferle, S.; Yordanov, S.; et al. Genomic characterization of severe acute respiratory syndrome-related coronavirus in European bats and classification of coronaviruses based on partial RNA-dependent RNA polymerase gene sequences. J. Virol. 2010, 84, 11336–11349. [Google Scholar]
- Rihtaric, D.; Hostnik, P.; Steyer, A.; Grom, J.; Toplak, I. Identification of SARS-like coronaviruses in horseshoe bats (Rhinolophus hipposideros) in Slovenia. Arch. Virol. 2010, 155, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Annan, A.; Baldwin, H.J.; Corman, V.M.; Klose, S.M.; Owusu, M.; Nkrumah, E.E.; Badu, E.K.; Anti, P.; Agbenyega, O.; Meyer, B.; et al. Human betacoronavirus 2c EMC/2012-related viruses in bats, Ghana and Europe. Emerg. Infect. Dis. 2013, 19, 456–459. [Google Scholar] [CrossRef] [PubMed]
- Reusken, C.B.E.M.; Lina, P.H.C.; Pielaat, A.; de Vries, A.; Dam-Deisz, C.; Adema, J.; Drexler, J.F.; Drosten, C.; Kooi, E.A. Circulation of group 2 coronaviruses in a bat species common to urban areas in Western Europe. Vector Borne Zoonotic Dis. 2010, 10, 785–791. [Google Scholar] [CrossRef] [PubMed]
- De Benedictis, P.; Marciano, S.; Scaravelli, D.; Priori, P.; Zecchin, B.; Capua, I.; Monne, I.; Cattoli, G. Alpha and lineage C betaCoV infections in Italian bats. Virus Genes 2014, 48, 366–371. [Google Scholar] [CrossRef] [PubMed]
- Negredo, A.; Palacios, G.; Vázquez-Morón, S.; González, F.; Dopazo, H.; Molero, F.; Juste, J.; Quetglas, J.; Savji, N.; de la Cruz Martínez, M.; et al. Discovery of an Ebolavirus-like filovirus in Europe. PLoS Pathog. 2011, 7, e1002304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drexler, J.F.; Seelen, A.; Corman, V.M.; Fumie Tateno, A.; Cottontail, V.; Melim Zerbinati, R.; Gloza-Rausch, F.; Klose, S.M.; Adu-Sarkodie, Y.; Oppong, S.K.; et al. Bats worldwide carry hepatitis E-related viruses that form a putative novel genus within the family Hepeviridae. J. Virol. 2012, 86, 9134–9147. [Google Scholar] [CrossRef] [PubMed]
- Wibbelt, G.; Kurth, A.; Yasmum, N.; Bannert, M.; Nagel, S.; Nitsche, A.; Ehlers, B. Discovery of herpesviruses in bats. J. Gen. Virol. 2007, 88, 2651–2655. [Google Scholar] [CrossRef] [PubMed]
- Molnár, V.; Jánoska, M.; Harrach, B.; Glávits, R.; Pálmai, N.; Rigó, D.; Sós, E.; Liptovszky, M. Detection of a novel bat gammaherpesvirus in Hungary. Acta Vet. Hung. 2008, 56, 529–538. [Google Scholar] [CrossRef] [PubMed]
- García-Pérez, R.; Ibáñez, C.; Godínez, J.M.; Aréchiga, N.; Garin, I.; Pérez-Suárez, G.; de Paz, O.; Juste, J.; Echevarría, J.E.; Bravo, I.G. Novel papillomaviruses in free-ranging Iberian bats: No virus-host co-evolution, no strict host specificity, and hints for recombination. Genome Biol. Evol. 2014, 6, 94–104. [Google Scholar] [CrossRef] [PubMed]
- Kurth, A.; Kohl, C.; Brinkmann, A.; Ebinger, A.; Harper, J.A.; Wang, L.-F.; Mühldorfer, K.; Wibbelt, G. Novel paramyxoviruses in free-ranging European bats. PLoS One 2012, 7, e38688. [Google Scholar] [CrossRef] [PubMed]
- Drexler, J.F.; Corman, V.M.; Müller, M.A.; Maganga, G.D.; Vallo, P.; Binger, T.; Gloza-Rausch, F.; Rasche, A.; Yordanov, S.; Seebens, A.; et al. Bats host major mammalian paramyxoviruses. Nat. Commun. 2012, 3, 796. [Google Scholar] [CrossRef] [PubMed]
- Kohl, C.; Lesnik, R.; Brinkmann, A.; Ebinger, A.; Radonić, A.; Nitsche, A.; Mühldorfer, K.; Wibbelt, G.; Kurth, A. Isolation and characterization of three mammalian orthoreoviruses from European bats. PLoS One 2012, 7, e43106. [Google Scholar] [CrossRef] [PubMed]
- Lelli, D.; Moreno, A.; Lavazza, A.; Bresaola, M.; Canelli, E.; Boniotti, M.B.; Cordioli, P. Identification of Mammalian Orthoreovirus Type 3 in Italian Bats. Zoonoses Public Health 2012, 60, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Bourhy, H.; Kissi, B.; Lafon, M.; Sacramento, D.; Tordo, N. Antigenic and molecular characterization of bat rabies virus in Europe. J. Clin. Microbiol. 1992, 30, 2419–2426. [Google Scholar] [PubMed]
- Fooks, A.R.; Brookes, S.M.; Johnson, N.; McElhinney, L.M.; Hutson, A.M. European bat lyssaviruses: An emerging zoonosis. Epidemiol. Infect. 2003, 131, 1029–1039. [Google Scholar] [CrossRef] [PubMed]
- Schatz, J.; Fooks, A.R.; McElhinney, L.; Horton, D.; Echevarria, J.; Vázquez-Moron, S.; Kooi, E.A.; Rasmussen, T.B.; Müller, T.; Freuling, C.M. Bat rabies surveillance in Europe. Zoonoses Public Health 2013, 60, 22–34. [Google Scholar] [CrossRef] [PubMed]
- Van Der Poel, W.H.M.; Van Der Heide, R.; Verstraten, E.R.A.M.; Takumi, K.; Lina, P.H.C.; Kramps, J.A. European bat lyssaviruses, The Netherlands. Emerg. Infect. Dis. 2005, 11, 1854–1859. [Google Scholar] [CrossRef]
- Delmas, O.; Holmes, E.C.; Talbi, C.; Larrous, F.; Dacheux, L.; Bouchier, C.; Bourhy, H. Genomic diversity and evolution of the lyssaviruses. PLoS One 2008, 3, e2057. [Google Scholar] [CrossRef] [PubMed]
- Dacheux, L.; Berthet, N.; Dissard, G.; Holmes, E.C.; Delmas, O.; Larrous, F.; Guigon, G.; Dickinson, P.; Faye, O.; Sall, A.A.; et al. Application of broad-spectrum resequencing microarray for genotyping rhabdoviruses. J. Virol. 2010, 84, 9557–9574. [Google Scholar] [CrossRef] [PubMed]
- Badrane, H.; Bahloul, C.; Perrin, P.; Tordo, N. Evidence of two Lyssavirus phylogroups with distinct pathogenicity and immunogenicity. J. Virol. 2001, 75, 3268–3276. [Google Scholar] [CrossRef] [PubMed]
- Müller, T.; Johnson, N.; Freuling, C.M.; Fooks, A.R.; Selhorst, T.; Vos, A. Epidemiology of bat rabies in Germany. Arch. Virol. 2007, 152, 273–288. [Google Scholar] [CrossRef] [PubMed]
- Picard-Meyer, E.; Barrat, J.; Tissot, E.; Barrat, M.J.; Bruyère, V.; Cliquet, F. Genetic analysis of European bat lyssavirus type 1 isolates from France. Vet. Rec. 2004, 154, 589–595. [Google Scholar] [CrossRef] [PubMed]
- Amengual, B.; Whitby, J.; King, A.; Cobo, J.; Bourhy, H. Evolution of European bat lyssaviruses. J. Gen. Virol. 1997, 78, 2319–2328. [Google Scholar] [PubMed]
- Freuling, C.M.; Beer, M.; Conraths, F.J.; Finke, S.; Hoffmann, B.; Keller, B.; Kliemt, J.; Mettenleiter, T.C.; Mühlbach, E.; Teifke, J.P.; et al. Novel lyssavirus in Natterer’s bat, Germany. Emerg. Infect. Dis. 2011, 17, 1519–1522. [Google Scholar] [PubMed]
- Jakava-Viljanen, M.; Lilley, T.; Kyheröinen, E.-M.; Huovilainen, A. First encounter of European bat lyssavirus type 2 (EBLV-2) in a bat in Finland. Epidemiol. Infect. 2010, 138, 1581–1585. [Google Scholar] [CrossRef] [PubMed]
- Vázquez-Moron, S.; Juste, J.; Ibáñez, C.; Berciano, J.M.; Echevarria, J.E. Phylogeny of European bat Lyssavirus 1 in Eptesicus isabellinus bats, Spain. Emerg. Infect. Dis. 2011, 17, 520–523. [Google Scholar] [CrossRef] [PubMed]
- Johnson, N.; Selden, D.; Parsons, G.; Healy, D.; Brookes, S.M.; McElhinney, L.M.; Hutson, A.M.; Fooks, A.R. Isolation of a European bat lyssavirus type 2 from a Daubenton’s bat in the United Kingdom. Vet. Rec. 2003, 152, 383–387. [Google Scholar] [CrossRef] [PubMed]
- Picard-Meyer, E.; Servat, A.; Robardet, E.; Moinet, M.; Borel, C.; Cliquet, F. Isolation of Bokeloh bat lyssavirus in Myotis nattereri in France. Arch. Virol. 2013, 158, 2333–2340. [Google Scholar] [CrossRef] [PubMed]
- Freuling, C.M.; Abendroth, B.; Beer, M.; Fischer, M.; Hanke, D.; Hoffmann, B.; Höper, D.; Just, F.; Mettenleiter, T.C.; Schatz, J.; et al. Molecular diagnostics for the detection of Bokeloh bat lyssavirus in a bat from Bavaria, Germany. Virus Res. 2013, 177, 201–204. [Google Scholar] [CrossRef] [PubMed]
- Aznar-Lopez, C.; Vazquez-Moron, S.; Marston, D.A.; Juste, J.; Ibáñez, C.; Berciano, J.M.; Salsamendi, E.; Aihartza, J.; Banyard, A.C.; McElhinney, L.; et al. Detection of rhabdovirus viral RNA in oropharyngeal swabs and ectoparasites of Spanish bats. J. Gen. Virol. 2013, 94, 69–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koch, R. Die aetiologie der tuberkulose. Klin. Wochenschr. 1932, 11, 490–492. [Google Scholar] [CrossRef]
- International Committee on Taxonomy of Viruses (ICTV) online 2013. Available online: http://www.ictvonline.org/virusTaxonomy.asp?version=2013 (accessed on 15 April 2014).
- Drosten, C.; Günther, S.; Preiser, W.; van der Werf, S.; Brodt, H.-R.; Becker, S.; Rabenau, H.; Panning, M.; Kolesnikova, L.; Fouchier, R.A.M.; et al. Identification of a novel coronavirus in patients with severe acute respiratory syndrome. N. Engl. J. Med. 2003, 348, 1967–1976. [Google Scholar] [CrossRef] [PubMed]
- Fouchier, R.A.M.; Kuiken, T.; Schutten, M.; van Amerongen, G.; van Doornum, G.J.J.; van den Hoogen, B.G.; Peiris, M.; Lim, W.; Stöhr, K.; Osterhaus, A.D.M.E. Aetiology: Koch’s postulates fulfilled for SARS virus. Nature 2003, 423, 240. [Google Scholar] [CrossRef] [PubMed]
- Kuiken, T.; Fouchier, R.A.M.; Schutten, M.; Rimmelzwaan, G.F.; van Amerongen, G.; van Riel, D.; Laman, J.D.; de Jong, T.; van Doornum, G.; Lim, W.; et al. Newly discovered coronavirus as the primary cause of severe acute respiratory syndrome. Lancet 2003, 362, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Peiris, J.S.M.; Guan, Y.; Yuen, K.Y. Severe acute respiratory syndrome. Nat. Med. 2004, 10, S88–S97. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.-F.; Shi, Z.; Zhang, S.; Field, H.; Daszak, P.; Eaton, B.T. Review of bats and SARS. Emerg. Infect. Dis. 2006, 12, 1834–1840. [Google Scholar] [CrossRef] [PubMed]
- WHO. Available online: http://www.who.int/csr/don/2014_06_26_mers/en (accessed on 26 June 2014).
- The WHO MERS-CoV Research Group. State of knowledge and data gaps of Middle East respiratory syndrome coronavirus (MERS-CoV) in humans. PLOS Curr. 2013, 5. doi:10.1371/currents.outbreaks.0bf719e352e7478f8ad85fa30127ddb8. [Google Scholar]
- Briese, T.; Mishra, N.; Jain, K.; Zalmout, I.S.; Jabado, O.J.; Karesh, W.B.; Daszak, P.; Mohammed, O.B.; Alagaili, A.N.; Lipkin, W.I. Middle East respiratory syndrome coronavirus quasispecies that include homologues of human isolates revealed through whole-genome analysis and virus cultured from dromedary camels in Saudi Arabia. MBio 2014, 5, e01146–14. [Google Scholar] [PubMed]
- Ithete, N.L.; Stoffberg, S.; Corman, V.M.; Cottontail, V.M.; Richards, L.R.; Schoeman, M.C.; Drosten, C.; Drexler, J.F.; Preiser, W. Close relative of human Middle East respiratory syndrome coronavirus in bat, South Africa. Emerg. Infect. Dis. 2013, 19, 1697–1699. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Wong, S.-K.; Li, F.; Kuhn, J.H.; Huang, I.-C.; Choe, H.; Farzan, M. Animal origins of the severe acute respiratory syndrome coronavirus: Insight from ACE2-S-protein interactions. J. Virol. 2006, 80, 4211–4219. [Google Scholar] [CrossRef] [PubMed]
- Ge, X.-Y.; Li, J.-L.; Yang, X.-L.; Chmura, A.A.; Zhu, G.; Epstein, J.H.; Mazet, J.K.; Hu, B.; Zhang, W.; Peng, C.; et al. Isolation and characterization of a bat SARS-like coronavirus that uses the ACE2 receptor. Nature 2013, 503, 535–538. [Google Scholar] [CrossRef] [PubMed]
- Memish, Z.A.; Mishra, N.; Olival, K.J.; Fagbo, S.F.; Kapoor, V. Middle East respiratory syndrome coronavirus in bats, Saudi Arabia. Emerg. Infect. Dis. 2013, 19, 7–13. [Google Scholar] [CrossRef]
- Mole, B. Deadly coronavirus found in bats. Nat. News 2013. [Google Scholar] [CrossRef]
- Van der Putten, W.H.; Macel, M.; Visser, M.E. Predicting species distribution and abundance responses to climate change: Why it is essential to include biotic interactions across trophic levels. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2010, 365, 2025–2034. [Google Scholar] [CrossRef] [PubMed]
- ArcGIS Explorer; ESRI Inc: ESRI Inc, 1999.
- Appleton, B.R.; McKenzie, J.A.; Christidis, L. Molecular systematics and biogeography of the bent-wing bat complex Miniopterus schreibersii (Kuhl, 1817) (Chiroptera: Vespertilionidae). Mol. Phylogenet. Evol. 2004, 31, 431–439. [Google Scholar] [CrossRef] [PubMed]
- Mühldorfer, K.; Speck, S.; Kurth, A.; Lesnik, R.; Freuling, C.; Müller, T.; Kramer-Schadt, S.; Wibbelt, G. Diseases and causes of death in European bats: Dynamics in disease susceptibility and infection rates. PLoS One 2011, 6, e29773. [Google Scholar] [CrossRef] [PubMed]
- Fereidouni, S.; Kwasnitschka, L.; Balkema Buschmann, A.; Müller, T.; Freuling, C.; Schatz, J.; Pikula, J.; Bandouchova, H.; Hoffmann, R.; Ohlendorf, B.; et al. No Virological Evidence for an Influenza A—Like Virus in European Bats. Zoonoses Public Health 2014. [Google Scholar] [CrossRef]
- Drexler, J.F.; Geipel, A.; König, A.; Corman, V.M.; van Riel, D.; Leijten, L.M.; Bremer, C.M.; Rasche, A.; Cottontail, V.M.; Maganga, G.D.; et al. Bats carry pathogenic hepadnaviruses antigenically related to hepatitis B virus and capable of infecting human hepatocytes. Proc. Natl. Acad. Sci. USA 2013, 110, 16151–16156. [Google Scholar] [CrossRef]
- Kohl, C.; Kurth, A. Robert Koch Institute: Berlin, Germany, Unpublished work. 2013.
- Jung, Y.T.; Kim, G.R. Genomic characterization of M and S RNA segments of hantaviruses isolated from bats. Acta Virol. 1995, 39, 231–233. [Google Scholar] [PubMed]
- Kohl, C.; Vidovszky, M.Z.; Kurth, A. Create Two Species, Bat Adenovirus B and Murine Adenovirus B, in the Genus Mastadenovirus, Family Adenoviridae; In the report of the international committee on taxonomy of viruses. Elsevier Academic Press: San Diego, CA, USA; ICTV: 2011 ICTV: 2011, ICTV 2011.024aV. Available online: http://talk.ictvonline.org/ (accessed on 15 April 2014).
- Buonavoglia, C.; Martella, V. Canine respiratory viruses. Vet. Res. 2007, 38, 355–373. [Google Scholar] [PubMed]
- Davis, P.L.; Bourhy, H.; Holmes, E.C. The evolutionary history and dynamics of bat rabies virus. Infect. Genet. Evol. 2006, 6, 464–473. [Google Scholar] [CrossRef] [PubMed]
- Decaro, N.; Campolo, M.; Desario, C.; Ricci, D.; Camero, M.; Lorusso, E.; Elia, G.; Lavazza, A.; Martella, V.; Buonavoglia, C. Virological and molecular characterization of a mammalian orthoreovirus type 3 strain isolated from a dog in Italy. Vet. Microbiol. 2005, 109, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Tyler, K.L.; Barton, E.S.; Ibach, M.L.; Robinson, C.; Campbell, J.A.; O’Donnell, S.M.; Valyi-Nagy, T.; Clarke, P.; Wetzel, J.D.; Dermody, T.S. Isolation and molecular characterization of a novel type 3 reovirus from a child with meningitis. J. Infect. Dis. 2004, 189, 1664–1675. [Google Scholar] [CrossRef] [PubMed]
- Steyer, A.; Gutiérrez-Aguire, I.; Kolenc, M.; Koren, S.; Kutnjak, D.; Pokorn, M.; Poljšak-Prijatelj, M.; Racki, N.; Ravnikar, M.; Sagadin, M.; et al. High similarity of novel orthoreovirus detected in a child hospitalized with acute gastroenteritis to mammalian orthoreoviruses found in bats in Europe. J. Clin. Microbiol. 2013, 51, 3818–3825. [Google Scholar] [CrossRef] [PubMed]
- Chua, K.B.; Crameri, G.; Hyatt, A.; Yu, M.; Tompang, M.R.; Rosli, J.; McEachern, J.; Crameri, S.; Kumarasamy, V.; Eaton, B.T.; et al. A previously unknown reovirus of bat origin is associated with an acute respiratory disease in humans. Proc. Natl. Acad. Sci. USA 2007, 104, 11424–11429. [Google Scholar] [CrossRef] [PubMed]
- Chua, K.B.; Voon, K.; Crameri, G.; Tan, H.S.; Rosli, J.; McEachern, J.A.; Suluraju, S.; Yu, M.; Wang, L.-F. Identification and characterization of a new orthoreovirus from patients with acute respiratory infections. PLoS One 2008, 3, e3803. [Google Scholar] [CrossRef] [PubMed]
- Johnson, N.; Vos, A.; Freuling, C.; Tordo, N.; Fooks, A.R.; Müller, T. Human rabies due to lyssavirus infection of bat origin. Vet. Microbiol. 2010, 142, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Racey, P.A.; Hutson, A.M.; Lina, P.H.C. Bat rabies, public health and European bat conservation. Zoonoses Public Health 2013, 60, 58–68. [Google Scholar] [CrossRef] [PubMed]
- Lumio, J.; Hillbom, M.; Roine, R.; Ketonen, L.; Haltia, M.; Valle, M.; Neuvonen, E.; Lahdevirta, J. Human rabies of bat origin in Europe. Lancet 1986, 1, 378. [Google Scholar] [CrossRef]
- Fooks, A.R.; Mcelhinney, L.M.; Pounder, D.J.; Finnegan, C.J.; Mansfield, K.; Johnson, N.; Brookes, S.M.; Parson, G.; White, K.; Mcintyre, P.G.; et al. Case report: Isolation of a European bat lyssavirus type 2a from a fatal human case of rabies encephalitis. J. Med. Virol. 2003, 71, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Dietz, C.; von Helversen, O.; Nill, D. Handbuch der Fledermäuse Europas und Nordwestafrikas; (in German). Kosmos: Stuttgart, Germany, 2007. [Google Scholar]
- Serra-Cobo, J.; Amengual, B.; Abellán, C.; Bourhy, H. European bat lyssavirus infection in Spanish bat populations. Emerg. Infect. Dis. 2002, 8, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Müller, T.; Cox, J.; Peter, W.; Schafer, R.; Johnson, N.; McElhinney, L.M.; Geue, J.L.; Tjornehoj, K.; Fooks, A.R. Spill-over of European bat lyssavirus type 1 into a stone marten (Martes foina) in Germany. J. Vet. Med. B 2004, 51, 49–54. [Google Scholar] [CrossRef]
- Tjornehoj, K.; Fooks, A.R.; Agerholm, J.S.; Ronsholt, L. Natural and experimental infection of sheep with European bat lyssavirus type-1 of Danish bat origin. J. Comp. Pathol. 2006, 134, 190–201. [Google Scholar] [CrossRef] [PubMed]
- Dacheux, L.; Larrous, F.; Mailles, A.; Boisseleau, D.; Delmas, O.; Biron, C.; Bouchier, C.; Capek, I.; Muller, M.; Ilari, F.; et al. European bat Lyssavirus transmission among cats, Europe. Emerg. Infect. Dis. 2009, 15, 280–284. [Google Scholar] [CrossRef] [PubMed]
- Streicker, D.G.; Recuenco, S.; Valderrama, W.; Gomez Benavides, J.; Vargas, I.; Pacheco, V.; Condori Condori, R.E.; Montgomery, J.; Rupprecht, C.E.; Rohani, P.; et al. Ecological and anthropogenic drivers of rabies exposure in vampire bats: Implications for transmission and control. Proc. Biol. Sci. 2012, 279, 3384–3392. [Google Scholar] [CrossRef] [PubMed]
- von Humboldt, A. Ansichten der Natur, mit wissenschaftlichen Erläuterungen(in German), 2nd ed.; Cottasche, J.G., Ed.; Buchhandlung: Stuttgart/Tübingen, Germany, 1826. [Google Scholar]
- Buckley, L.B.; Davies, T.J.; Ackerly, D.D.; Kraft, N.J.B.; Harrison, S.P.; Anacker, B.L.; Cornell, H.V.; Damschen, E.I.; Grytnes, J.-A.; Hawkins, B.A.; et al. Phylogeny, niche conservatism and the latitudinal diversity gradient in mammals. Proc. Biol. Sci. 2010, 277, 2131–2138. [Google Scholar] [CrossRef] [PubMed]
- Simmons, N.B. Taxonomy of chiroptera. In Mammal Species of the World: A Taxonomic and Geographic Reference, 3rd ed.; Wilson, D.E., Reeder, D.M., Eds.; Johns Hopkins University: Baltimore, MD, USA, 2005. [Google Scholar]
- Teeling, E.C.; Springer, M.S.; Madsen, O.; Bates, P.; O’Brien, S.J.; Murphy, W.J. A molecular phylogeny for bats illuminates biogeography and the fossil record. Science 2005, 307, 580–584. [Google Scholar] [PubMed]
- Willig, M.; Selcer, K. Bat species density gradients in the New World: A statistical assessment. J. Biogeogr. 1989, 16, 189–195. [Google Scholar] [CrossRef]
- Buckley, H.L.; Miller, T.E.; Ellison, A.M.; Gotelli, N.J. Reverse latitudinal trends in species richness of pitcher-plant food webs. Ecol. Lett. 2003, 6, 825–829. [Google Scholar] [CrossRef]
- Fierer, N.; Jackson, R.B. The diversity and biogeography of soil bacterial communities. Proc. Natl. Acad. Sci. USA 2006, 103, 626–631. [Google Scholar] [CrossRef] [PubMed]
- Hillebrand, H.; Azovsky, A.I. Body size determines the strength of the latitudinal diversity gradient. Ecography 2001, 24, 251–256. [Google Scholar] [CrossRef]
- Martiny, J.B.H.; Bohannan, B.J.M.; Brown, J.H.; Colwell, R.K.; Fuhrman, J.A.; Green, J.L.; Horner-Devine, M.C.; Kane, M.; Krumins, J.A.; Kuske, C.R.; et al. Microbial biogeography: Putting microorganisms on the map. Nat. Rev. Microbiol. 2006, 4, 102–112. [Google Scholar]
- Peterson, A.T. Biogeography of diseases: A framework for analysis. Naturwissenschaften 2008, 95, 483–491. [Google Scholar] [CrossRef] [PubMed]
- Breitbart, M.; Rohwer, F. Here a virus, there a virus, everywhere the same virus? Trends Microbiol. 2005, 13, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Dolan, J.R. Microbial biogeography? J. Biogeogr. 2006, 33, 199–200. [Google Scholar] [CrossRef]
- Keesing, F.; Belden, L.K.; Daszak, P.; Dobson, A.; Harvell, C.D.; Holt, R.D.; Hudson, P.; Jolles, A.; Jones, K.E.; Mitchell, C.E.; et al. Impacts of biodiversity on the emergence and transmission of infectious diseases. Nature 2010, 468, 647–652. [Google Scholar] [CrossRef] [PubMed]
- Mills, J.N. Biodiversity loss and emerging infectious disease: An example from the rodent-borne hemorrhagic fevers. Biodiversity 2006, 7, 9–17. [Google Scholar] [CrossRef]
- Ostfeld, R.S. Biodiversity loss and the rise of zoonotic pathogens. Clin. Microbiol. Infect. 2009, 15, 40–43. [Google Scholar] [CrossRef] [PubMed]
- Randolph, S.E.; Dobson, A.D.M. Pangloss revisited: A critique of the dilution effect and the biodiversity-buffers-disease paradigm. Parasitology 2012, 139, 847–863. [Google Scholar] [CrossRef] [PubMed]
- Salkeld, D.J.; Padgett, K.A.; Jones, J.H. A meta-analysis suggesting that the relationship between biodiversity and risk of zoonotic pathogen transmission is idiosyncratic. Ecol. Lett. 2013, 16, 679–686. [Google Scholar] [CrossRef] [PubMed]
- Woolhouse, M.E.J.; Gowtage-Sequeria, S. Host range and emerging and reemerging pathogens. Emerg. Infect. Dis. 2005, 11, 1842–1847. [Google Scholar] [CrossRef] [PubMed]
- Woolhouse, M.E.J.; Taylor, L.H.; Haydon, D.T. Population biology of multihost pathogens. Science 2001, 292, 1109–1112. [Google Scholar] [CrossRef] [PubMed]
- Baas Becking, L.G.M. Geobiologie of Inleiding tot de Milieukunde; (in Dutch). W.P. Van Stockum & Zoon: Gravenhage, The Netherlands, 1934. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
MDPI and ACS Style
Kohl, C.; Kurth, A. European Bats as Carriers of Viruses with Zoonotic Potential. Viruses 2014, 6, 3110-3128. https://doi.org/10.3390/v6083110
AMA Style
Kohl C, Kurth A. European Bats as Carriers of Viruses with Zoonotic Potential. Viruses. 2014; 6(8):3110-3128. https://doi.org/10.3390/v6083110
Chicago/Turabian StyleKohl, Claudia, and Andreas Kurth. 2014. "European Bats as Carriers of Viruses with Zoonotic Potential" Viruses 6, no. 8: 3110-3128. https://doi.org/10.3390/v6083110