Human Cytomegalovirus US28 Facilitates Cell-to-Cell Viral Dissemination

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
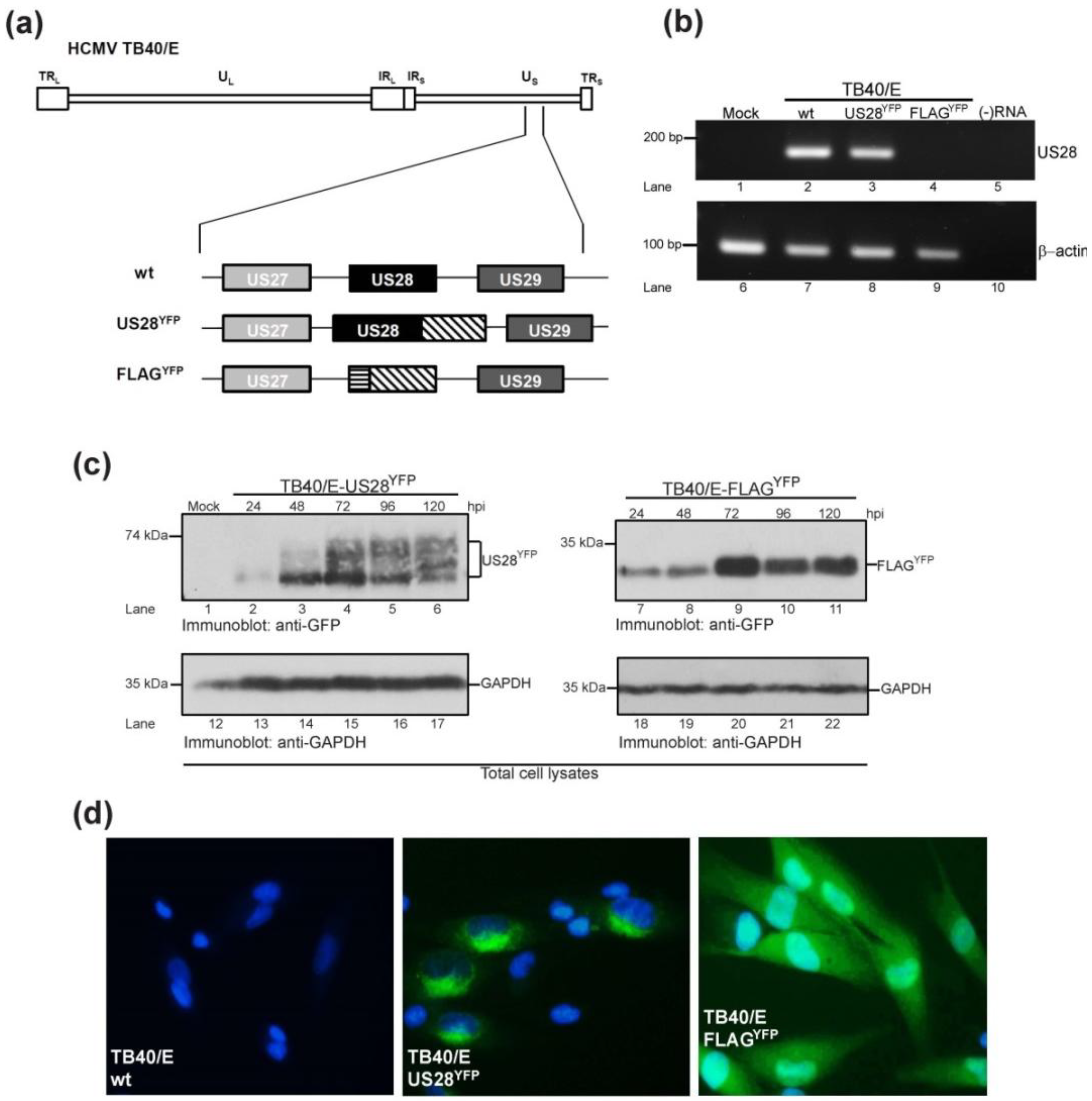
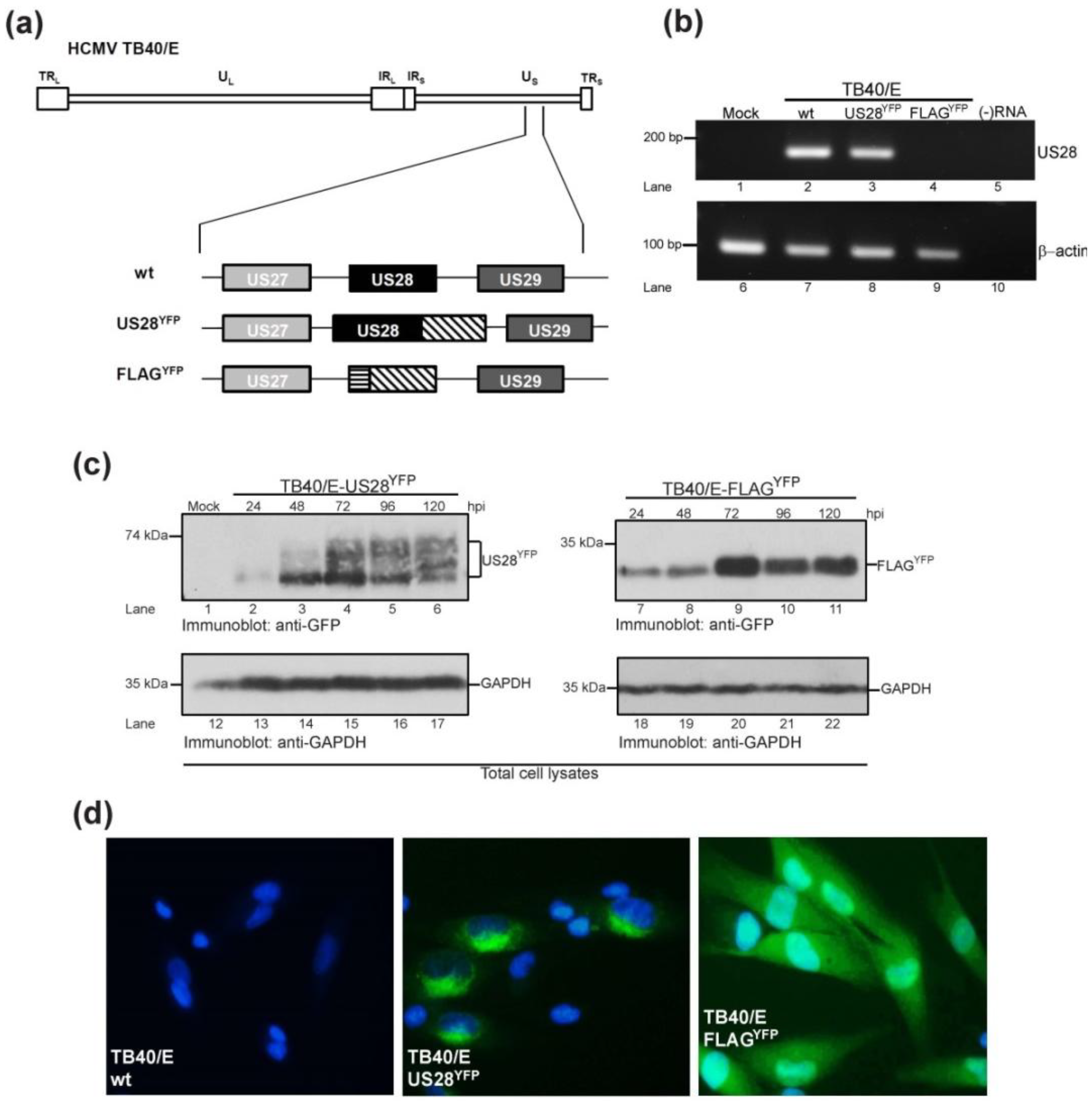
2.1. Generation of HCMV TB40/E US28 Variants
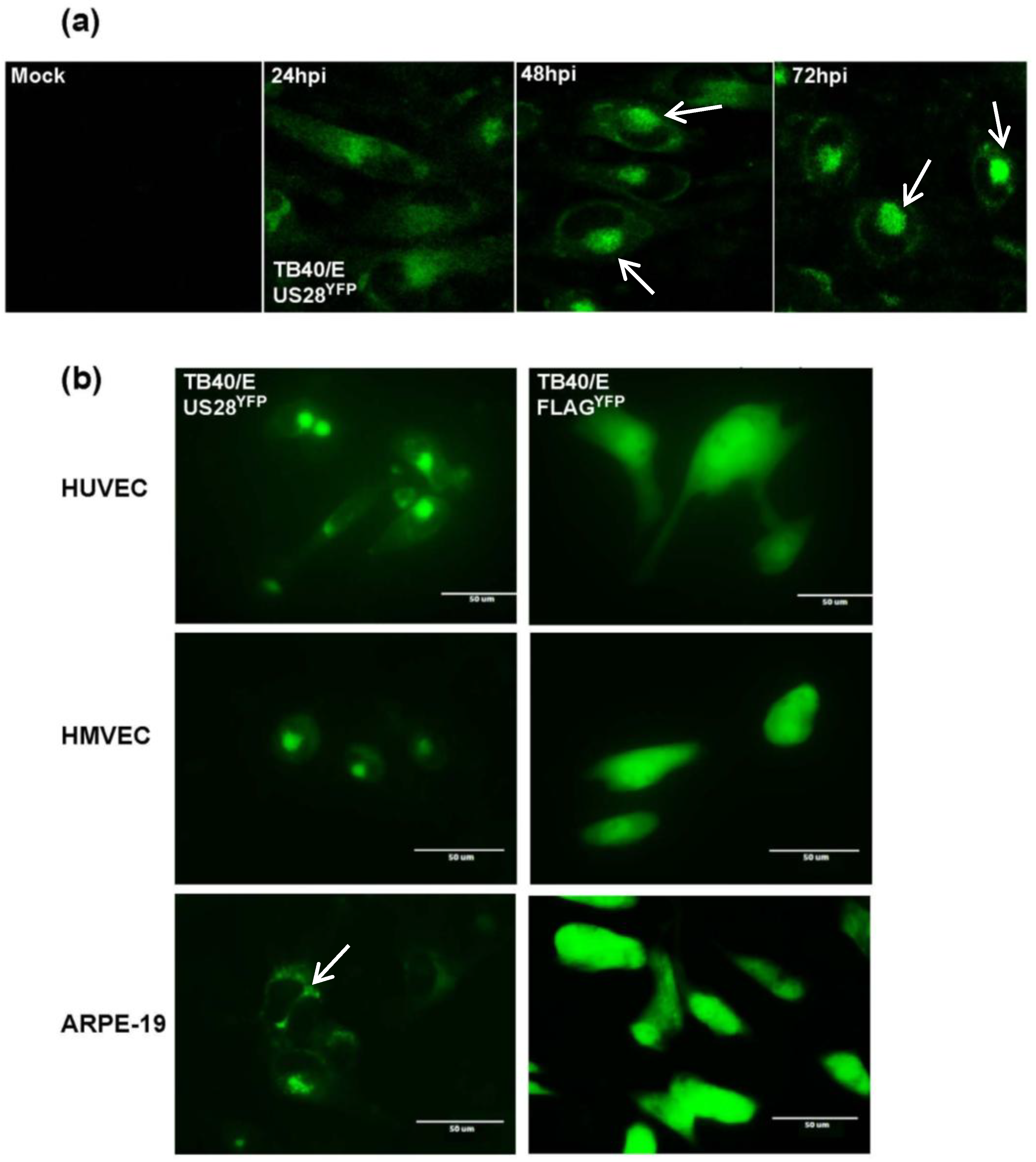
2.2. HCMV US28YFP Localizes as Large Vesicular Structures at Late Times of Infection
2.3. Characterization of US28YFP in HCMV-Infected Cells
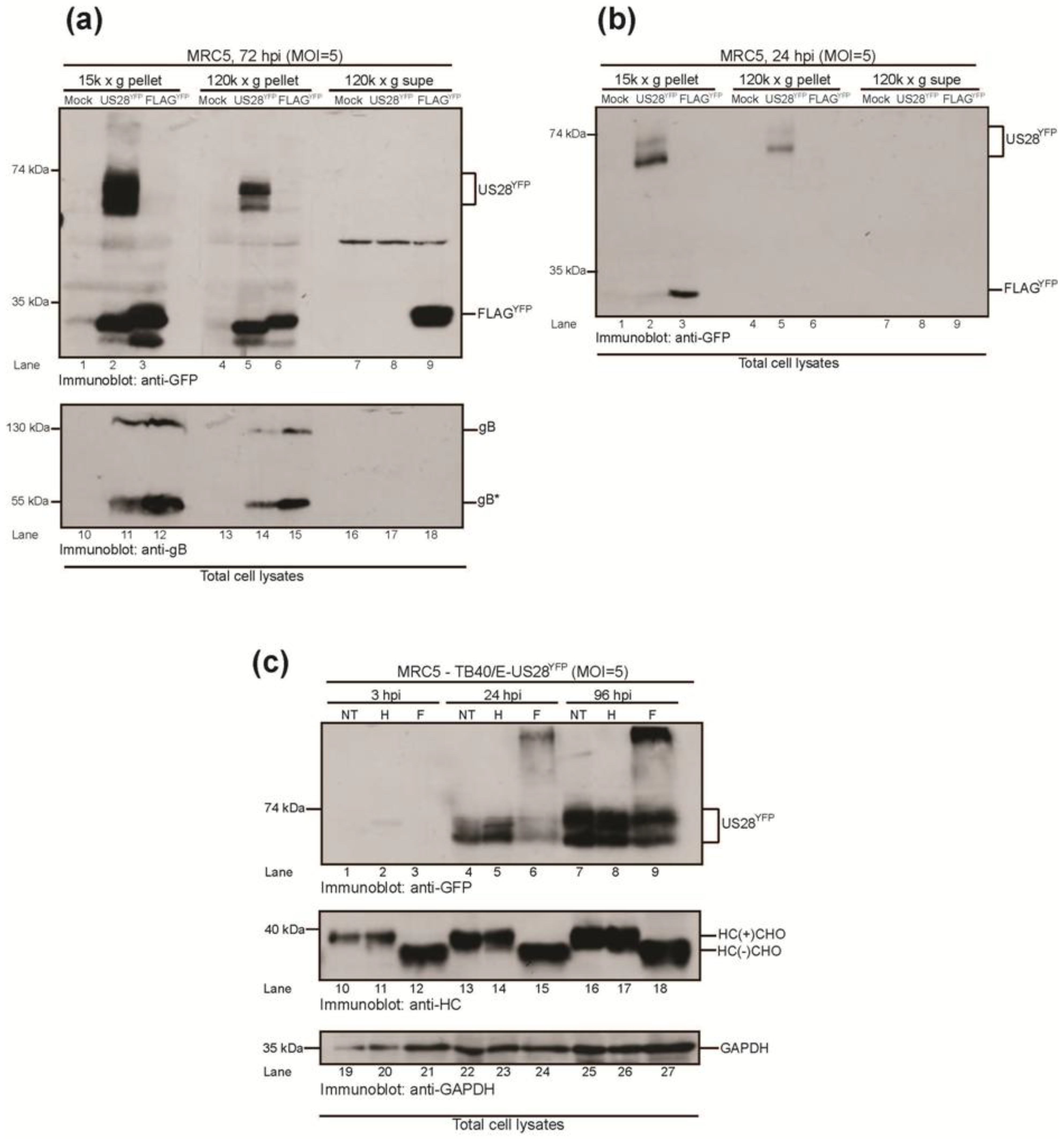
2.3.1. HCMV US28YFP Is Integrated into Dense Vesicular Bodies

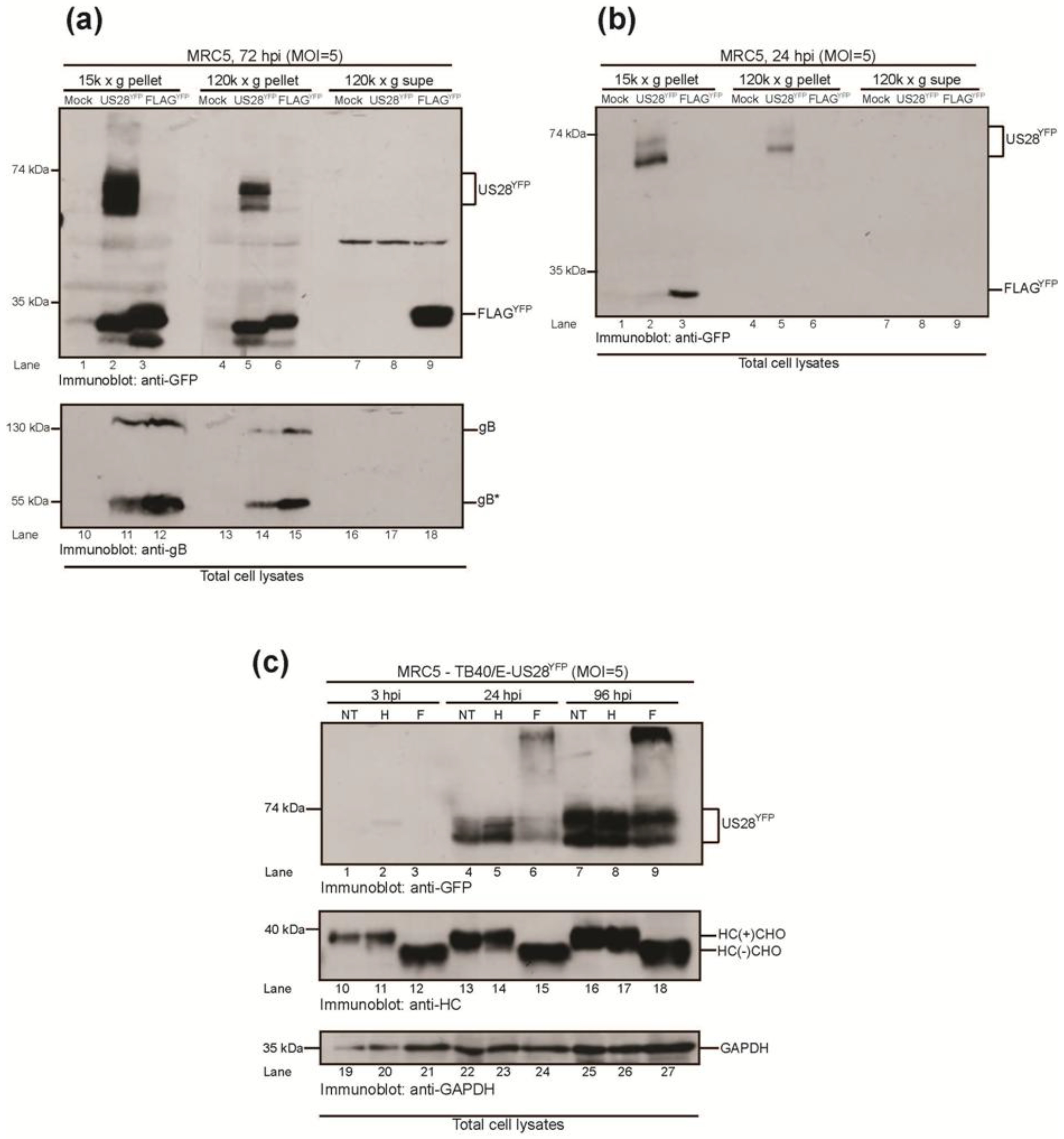
2.3.2. HMCV US28 Is a Non-Glycosylated Membrane Protein
2.4. Functional Analysis of TB40/E US28 Variants
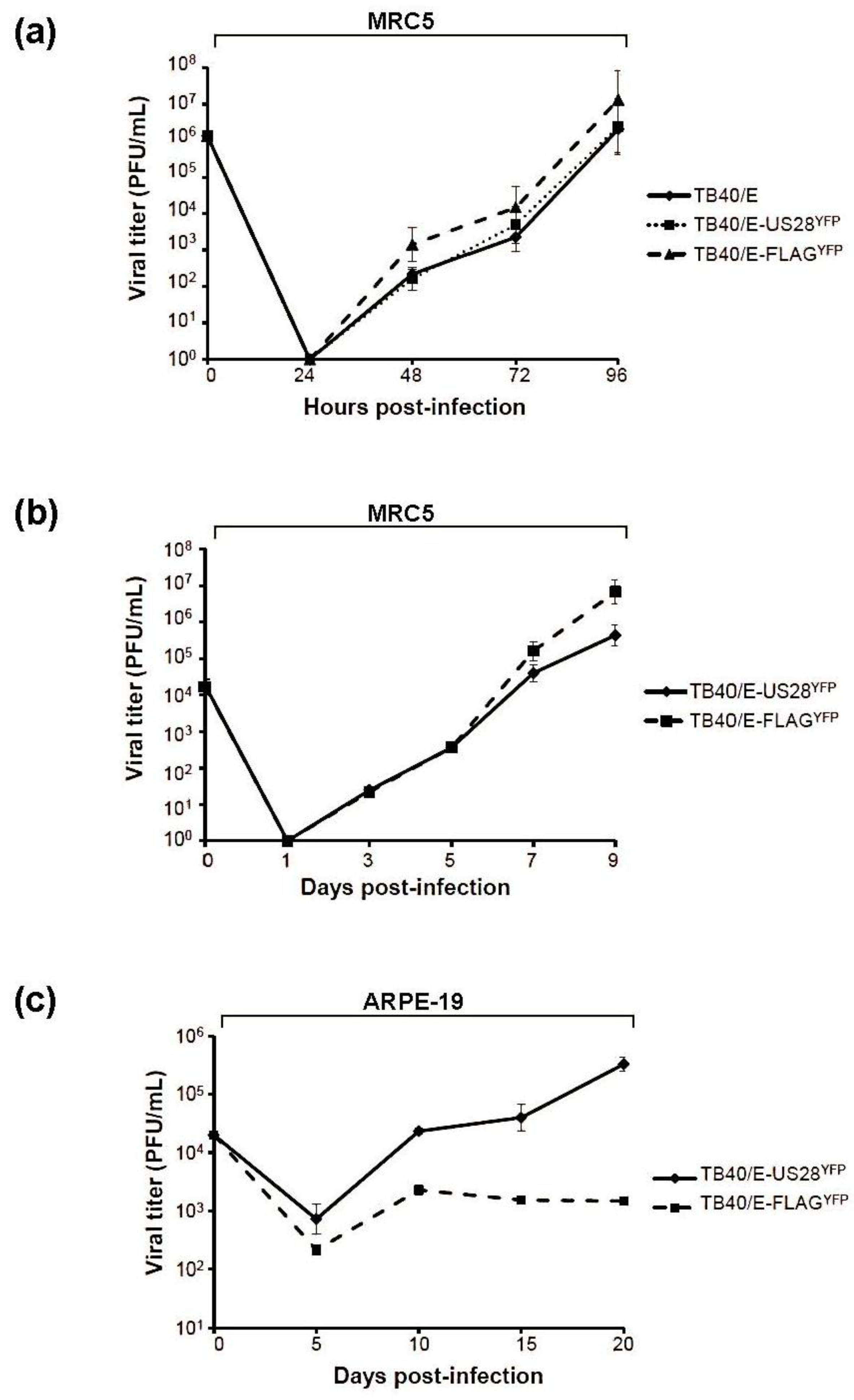
2.4.1. TB40/E ΔUS28 Accumulates Increased Amounts of Extracellular Virus in Fibroblasts
2.4.2. US28 Modulates HCMV Cell-to-Cell Dissemination

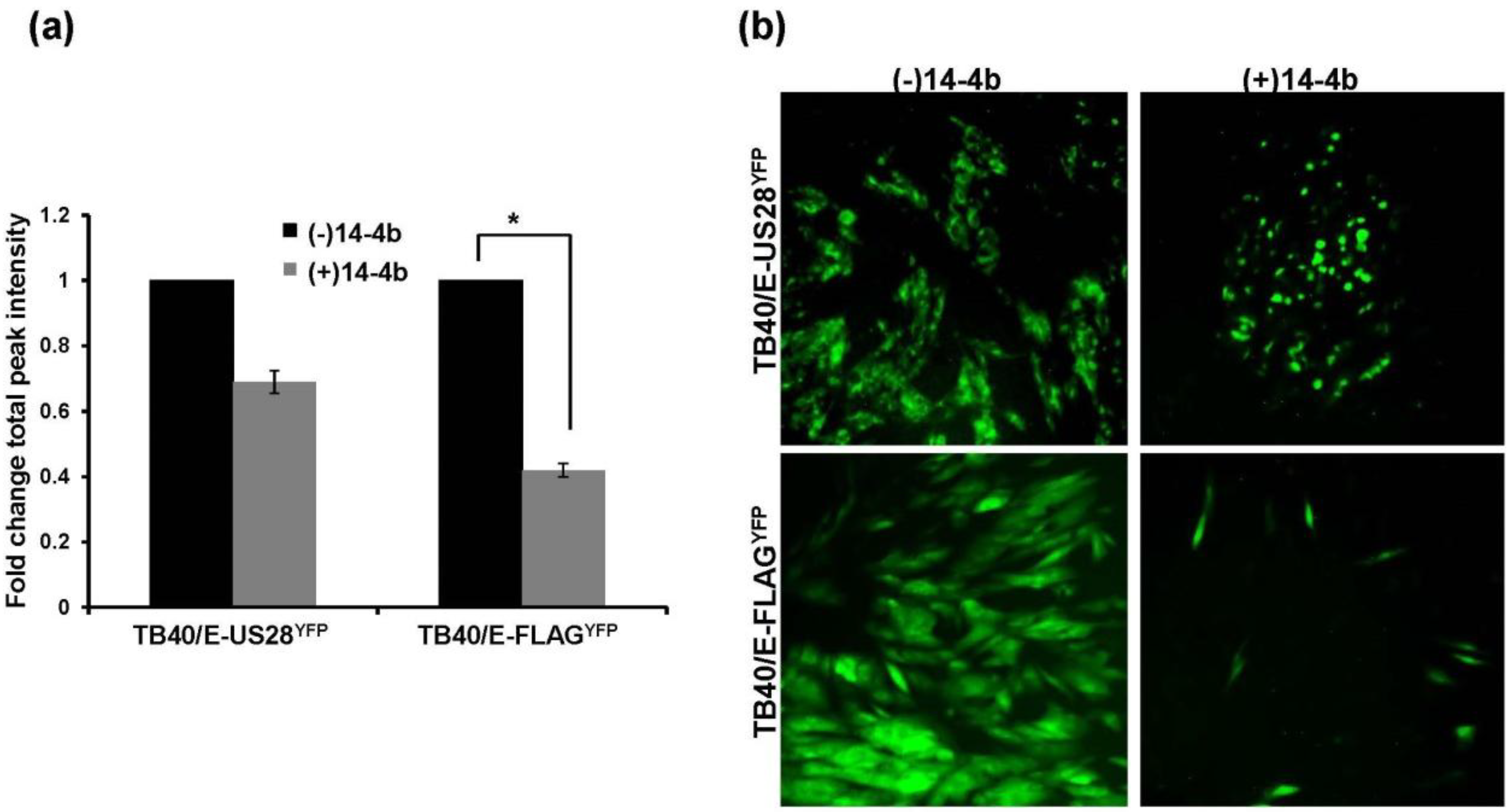
2.5. Inhibition of TB40/E ΔUS28 Dissemination by Anti-HCMV Neutralizing Antibody
3. Experimental Section
3.1. Cells and Viruses
3.2. Fluorescence Microscopy
3.3. Cell Fractionation and Immunoblot Analysis
3.4. N-Linked Protein Glycosylation Analysis
3.5. Analysis of Virus Growth and Spread
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References and Notes
- Britt, W. Manifestations of human cytomegalovirus infection: Proposed mechanisms of acute and chronic disease. Curr. Top. Microbiol. Immunol. 2008, 325, 417–470. [Google Scholar]
- Arvin, A.M. Human herpesviruses: Biology, therapy, and immunoprophylaxis; Cambridge University Press: Cambridge, New York, NY, USA, 2007; p. xx, 1388. [Google Scholar]
- Soderberg-Naucler, C. Does cytomegalovirus play a causative role in the development of various inflammatory diseases and cancer? J. Intern. Med. 2006, 259, 219–246. [Google Scholar] [CrossRef]
- Sissons, J.G.; Carmichael, A.J.; McKinney, N.; Sinclair, J.H.; Wills, M.R. Human cytomegalovirus and immunopathology. Springer Semin. Immunopathol. 2002, 24, 169–185. [Google Scholar] [CrossRef]
- Razonable, R.R. Epidemiology of cytomegalovirus disease in solid organ and hematopoietic stem cell transplant recipients. Am. J. Health Syst. Pharm. 2005, 62, S7–S13. [Google Scholar]
- Murphy, E.; Yu, D.; Grimwood, J.; Schmutz, J.; Dickson, M.; Jarvis, M.A.; Hahn, G.; Nelson, J.A.; Myers, R.M.; Shenk, T.E. Coding potential of laboratory and clinical strains of human cytomegalovirus. Proc. Natl. Acad. Sci. USA 2003, 100, 14976–14981. [Google Scholar] [CrossRef]
- Vischer, H.F.; Leurs, R.; Smit, M.J. Hcmv-encoded g-protein-coupled receptors as constitutively active modulators of cellular signaling networks. Trends Pharmacol. Sci. 2006, 27, 56–63. [Google Scholar] [CrossRef]
- Sodhi, A.; Montaner, S.; Gutkind, J.S. Viral hijacking of g-protein-coupled-receptor signalling networks. Nat. Rev. Mol. Cell Biol. 2004, 5, 998–1012. [Google Scholar] [CrossRef]
- Evers, D.L.; Wang, X.; Huang, E.S. Cellular stress and signal transduction responses to human cytomegalovirus infection. Microbes Infect. 2004, 6, 1084–1093. [Google Scholar] [CrossRef]
- Margulies, B.J.; Gibson, W. The chemokine receptor homologue encoded by us27 of human cytomegalovirus is heavily glycosylated and is present in infected human foreskin fibroblasts and enveloped virus particles. Virus Res. 2007, 123, 57–71. [Google Scholar] [CrossRef]
- Margulies, B.J.; Browne, H.; Gibson, W. Identification of the human cytomegalovirus g protein-coupled receptor homologue encoded by ul33 in infected cells and enveloped virus particles. Virology 1996, 225, 111–125. [Google Scholar] [CrossRef]
- O'Connor, C.M.; Shenk, T. Human cytomegalovirus pul78 g protein-coupled receptor homologue is required for timely cell entry in epithelial cells but not fibroblasts. J. Virol. 2012, 86, 11425–11433. [Google Scholar] [CrossRef]
- Casarosa, P.; Gruijthuijsen, Y.K.; Michel, D.; Beisser, P.S.; Holl, J.; Fitzsimons, C.P.; Verzijl, D.; Bruggeman, C.A.; Mertens, T.; Leurs, R.; et al. Constitutive signaling of the human cytomegalovirus-encoded receptor ul33 differs from that of its rat cytomegalovirus homolog r33 by promiscuous activation of g proteins of the gq, gi, and gs classes. J. Biol. Chem. 2003, 278, 50010–50023. [Google Scholar] [CrossRef]
- Minisini, R.; Tulone, C.; Luske, A.; Michel, D.; Mertens, T.; Gierschik, P.; Moepps, B. Constitutive inositol phosphate formation in cytomegalovirus-infected human fibroblasts is due to expression of the chemokine receptor homologue pus28. J. Virol. 2003, 77, 4489–4501. [Google Scholar] [CrossRef]
- Billstrom, M.A.; Johnson, G.L.; Avdi, N.J.; Worthen, G.S. Intracellular signaling by the chemokine receptor us28 during human cytomegalovirus infection. J. Virol. 1998, 72, 5535–5544. [Google Scholar]
- Streblow, D.N.; Soderberg-Naucler, C.; Vieira, J.; Smith, P.; Wakabayashi, E.; Ruchti, F.; Mattison, K.; Altschuler, Y.; Nelson, J.A. The human cytomegalovirus chemokine receptor us28 mediates vascular smooth muscle cell migration. Cell 1999, 99, 511–520. [Google Scholar] [CrossRef]
- Waldhoer, M.; Kledal, T.N.; Farrell, H.; Schwartz, T.W. Murine cytomegalovirus (cmv) m33 and human cmv us28 receptors exhibit similar constitutive signaling activities. J. Virol. 2002, 76, 8161–8168. [Google Scholar] [CrossRef]
- Arnolds, K.L.; Lares, A.P.; Spencer, J.V. The us27 gene product of human cytomegalovirus enhances signaling of host chemokine receptor cxcr4. Virology 2013, 439, 122–131. [Google Scholar] [CrossRef]
- O'Connor, C.M.; Shenk, T. Human cytomegalovirus pus27 g protein-coupled receptor homologue is required for efficient spread by the extracellular route but not for direct cell-to-cell spread. J. Virol. 2011, 85, 3700–3707. [Google Scholar] [CrossRef]
- Davis-Poynter, N.J.; Lynch, D.M.; Vally, H.; Shellam, G.R.; Rawlinson, W.D.; Barrell, B.G.; Farrell, H.E. Identification and characterization of a g protein-coupled receptor homolog encoded by murine cytomegalovirus. J. Virol. 1997, 71, 1521–1529. [Google Scholar]
- Oliveira, S.A.; Shenk, T.E. Murine cytomegalovirus m78 protein, a g protein-coupled receptor homologue, is a constituent of the virion and facilitates accumulation of immediate-early viral mrna. Proc. Natl. Acad. Sci. USA 2001, 98, 3237–3242. [Google Scholar] [CrossRef]
- Fraile-Ramos, A.; Kledal, T.N.; Pelchen-Matthews, A.; Bowers, K.; Schwartz, T.W.; Marsh, M. The human cytomegalovirus us28 protein is located in endocytic vesicles and undergoes constitutive endocytosis and recycling. Mol. Biol. Cell 2001, 12, 1737–1749. [Google Scholar] [CrossRef]
- Fraile-Ramos, A.; Pelchen-Matthews, A.; Kledal, T.N.; Browne, H.; Schwartz, T.W.; Marsh, M. Localization of hcmv ul33 and us27 in endocytic compartments and viral membranes. Traffic 2002, 3, 218–232. [Google Scholar] [CrossRef]
- Stropes, M.P.; Miller, W.E. Functional analysis of human cytomegalovirus pus28 mutants in infected cells. J. Gen. Virol. 2008, 89, 97–105. [Google Scholar] [CrossRef]
- Maley, F.; Trimble, R.B.; Tarentino, A.L.; Plummer, T.H., Jr. Characterization of glycoproteins and their associated oligosaccharides through the use of endoglycosidases. Anal. Biochem. 1989, 180, 195–204. [Google Scholar] [CrossRef]
- Noriega, V.M.; Tortorella, D. Human cytomegalovirus-encoded immune modulators partner to downregulate major histocompatibility complex class i molecules. J. Virol. 2009, 83, 1359–1367. [Google Scholar] [CrossRef]
- Silva, M.C.; Schroer, J.; Shenk, T. Human cytomegalovirus cell-to-cell spread in the absence of an essential assembly protein. Proc. Natl. Acad. Sci. USA 2005, 102, 2081–2086. [Google Scholar] [CrossRef]
- Li, L.; Coelingh, K.L.; Britt, W.J. Human cytomegalovirus neutralizing antibody-resistant phenotype is associated with reduced expression of glycoprotein h. J. Virol. 1995, 69, 6047–6053. [Google Scholar]
- Warming, S.; Costantino, N.; Court, D.L.; Jenkins, N.A.; Copeland, N.G. Simple and highly efficient bac recombineering using galk selection. Nucleic Acids Res. 2005, 33, e36. [Google Scholar] [CrossRef]
- Adobe Photoshop, CS5.1; Adobe Systems: New York, NY, USA, 2011.
- MetaXpress, version 5.3; Molecular Devices: Sunnyvale, CA, USA, 2013.
- Baker, B.M.; Tortorella, D. Dislocation of an endoplasmic reticulum membrane glycoprotein involves the formation of partially dislocated ubiquitinated polypeptides. J. Biol. Chem. 2007, 282, 26845–26856. [Google Scholar] [CrossRef]
- Noriega, V.M.; Tortorella, D. A bipartite trigger for dislocation directs the proteasomal degradation of an endoplasmic reticulum membrane glycoprotein. J. Biol. Chem. 2008, 283, 4031–4043. [Google Scholar] [CrossRef]
- Gardner, T.J.; Bolovan-Fritts, C.; Teng, M.W.; Redmann, V.; Kraus, T.A.; Sperling, R.; Moran, T.; Britt, W.; Weinberger, L.S.; Tortorella, D. Development of a high-throughput assay to measure the neutralization capability of anti-cytomegalovirus antibodies. Clin. Vaccine Immunol. 2013, 20, 540–550. [Google Scholar] [CrossRef]
- Kledal, T.N.; Rosenkilde, M.M.; Schwartz, T.W. Selective recognition of the membrane-bound cx3c chemokine, fractalkine, by the human cytomegalovirus-encoded broad-spectrum receptor us28. FEBS Lett. 1998, 441, 209–214. [Google Scholar] [CrossRef]
- Pleskoff, O.; Treboute, C.; Alizon, M. The cytomegalovirus-encoded chemokine receptor us28 can enhance cell-cell fusion mediated by different viral proteins. J. Virol. 1998, 72, 6389–6397. [Google Scholar]
- Tschische, P.; Tadagaki, K.; Kamal, M.; Jockers, R.; Waldhoer, M. Heteromerization of human cytomegalovirus encoded chemokine receptors. Biochem. Pharmacol. 2011, 82, 610–619. [Google Scholar] [CrossRef] [Green Version]
- Beisser, P.S.; Laurent, L.; Virelizier, J.L.; Michelson, S. Human cytomegalovirus chemokine receptor gene us28 is transcribed in latently infected thp-1 monocytes. J. Virol. 2001, 75, 5949–5957. [Google Scholar] [CrossRef]
- Goda, S.; Imai, T.; Yoshie, O.; Yoneda, O.; Inoue, H.; Nagano, Y.; Okazaki, T.; Imai, H.; Bloom, E.T.; Domae, N.; Umehara, H. Cx3c-chemokine, fractalkine-enhanced adhesion of thp-1 cells to endothelial cells through integrin-dependent and -independent mechanisms. J. Immunol. 2000, 164, 4313–4320. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Noriega, V.M.; Gardner, T.J.; Redmann, V.; Bongers, G.; Lira, S.A.; Tortorella, D. Human Cytomegalovirus US28 Facilitates Cell-to-Cell Viral Dissemination. Viruses 2014, 6, 1202-1218. https://doi.org/10.3390/v6031202
Noriega VM, Gardner TJ, Redmann V, Bongers G, Lira SA, Tortorella D. Human Cytomegalovirus US28 Facilitates Cell-to-Cell Viral Dissemination. Viruses. 2014; 6(3):1202-1218. https://doi.org/10.3390/v6031202
Chicago/Turabian StyleNoriega, Vanessa M., Thomas J. Gardner, Veronika Redmann, Gerold Bongers, Sergio A. Lira, and Domenico Tortorella. 2014. "Human Cytomegalovirus US28 Facilitates Cell-to-Cell Viral Dissemination" Viruses 6, no. 3: 1202-1218. https://doi.org/10.3390/v6031202