Neonatal Calf Infection with Respiratory Syncytial Virus: Drawing Parallels to the Disease in Human Infants


Abstract
:1. Importance of Human and Bovine Respiratory Syncytial Viruses
2. Historical Perspectives on Respiratory Syncytial Viruses
3. Age and Seasonal Affects
4. RSV Viral Proteins
5. Experimental Bovine RSV in Calves and Similarities to Lesions seen in Human Infants
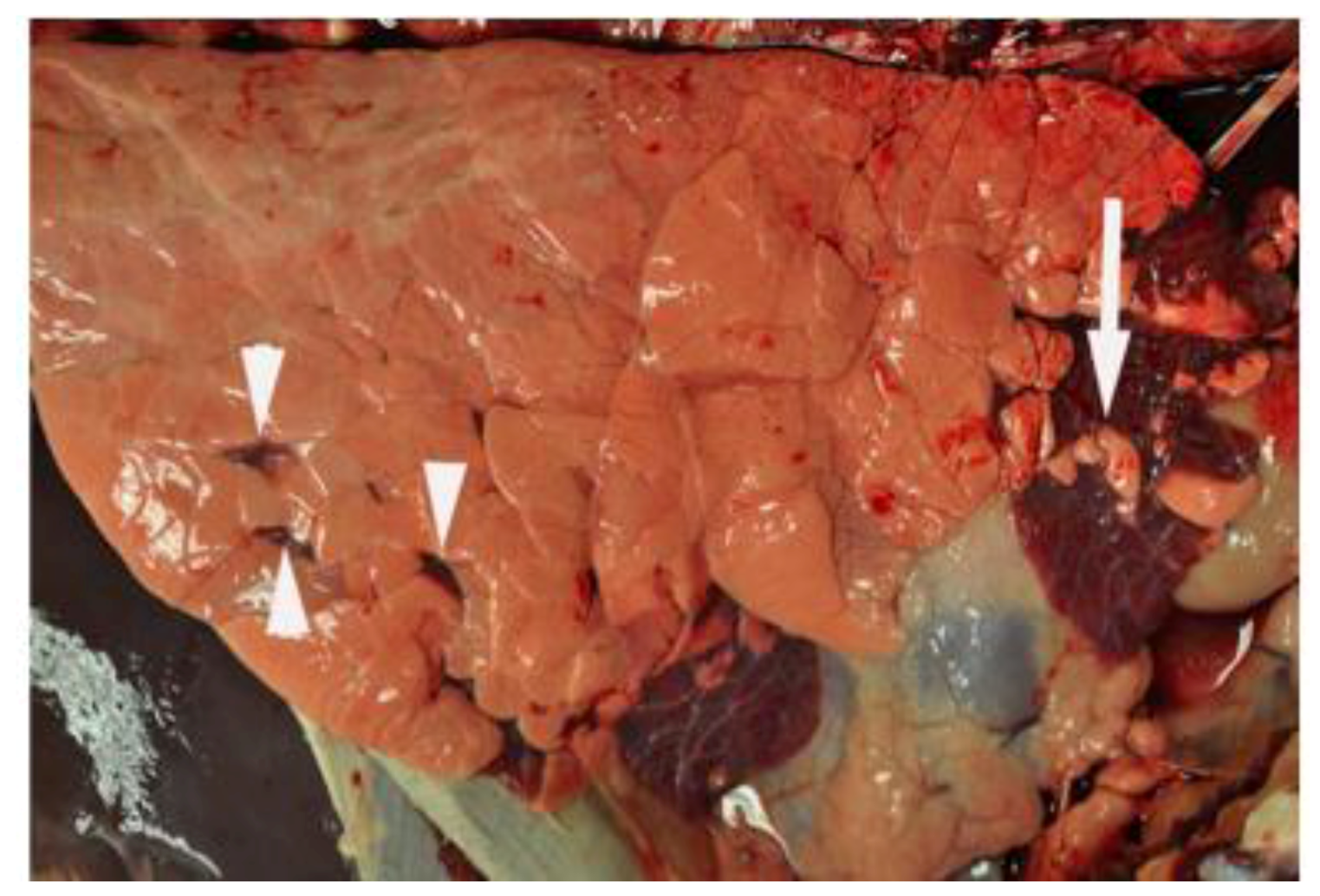
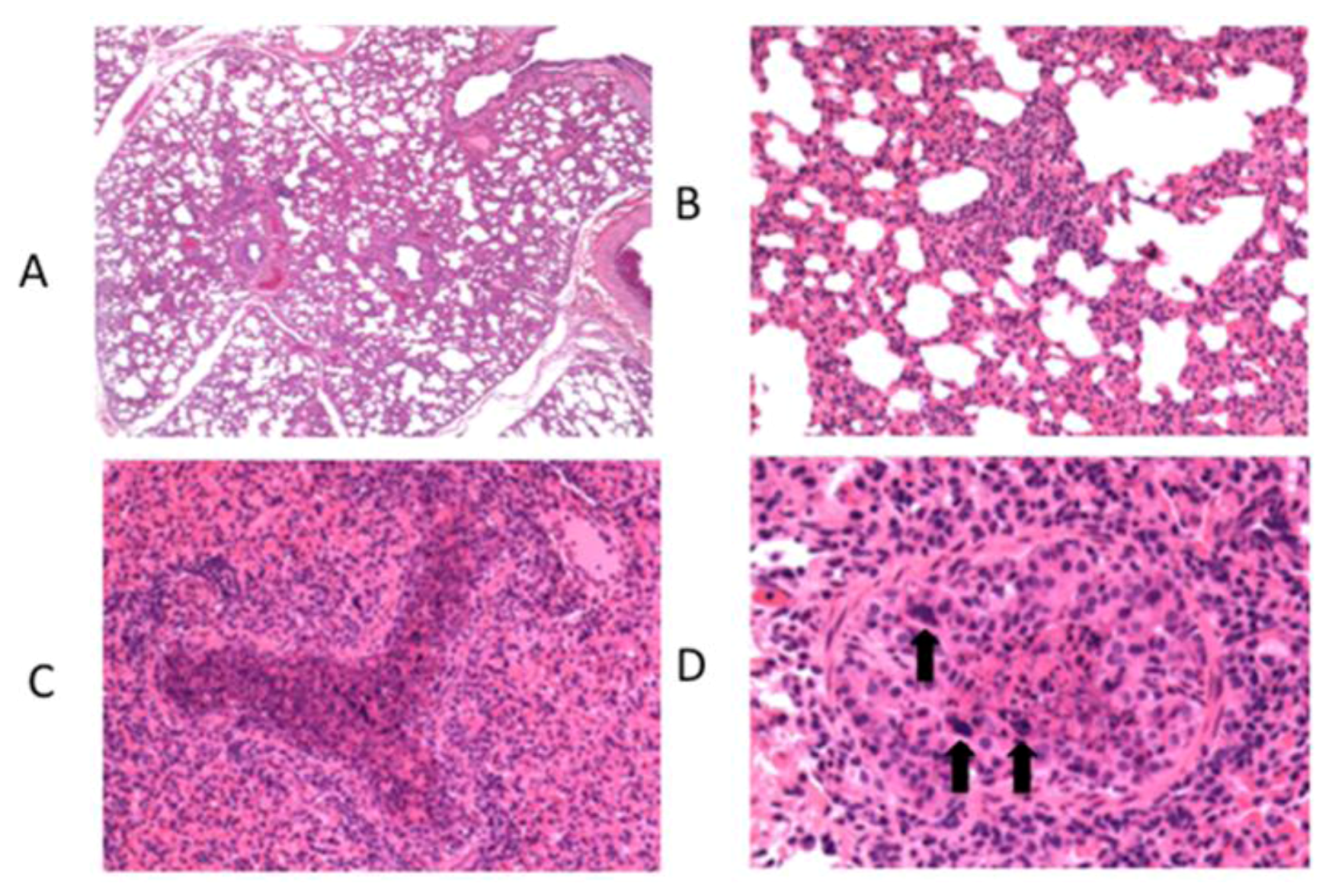
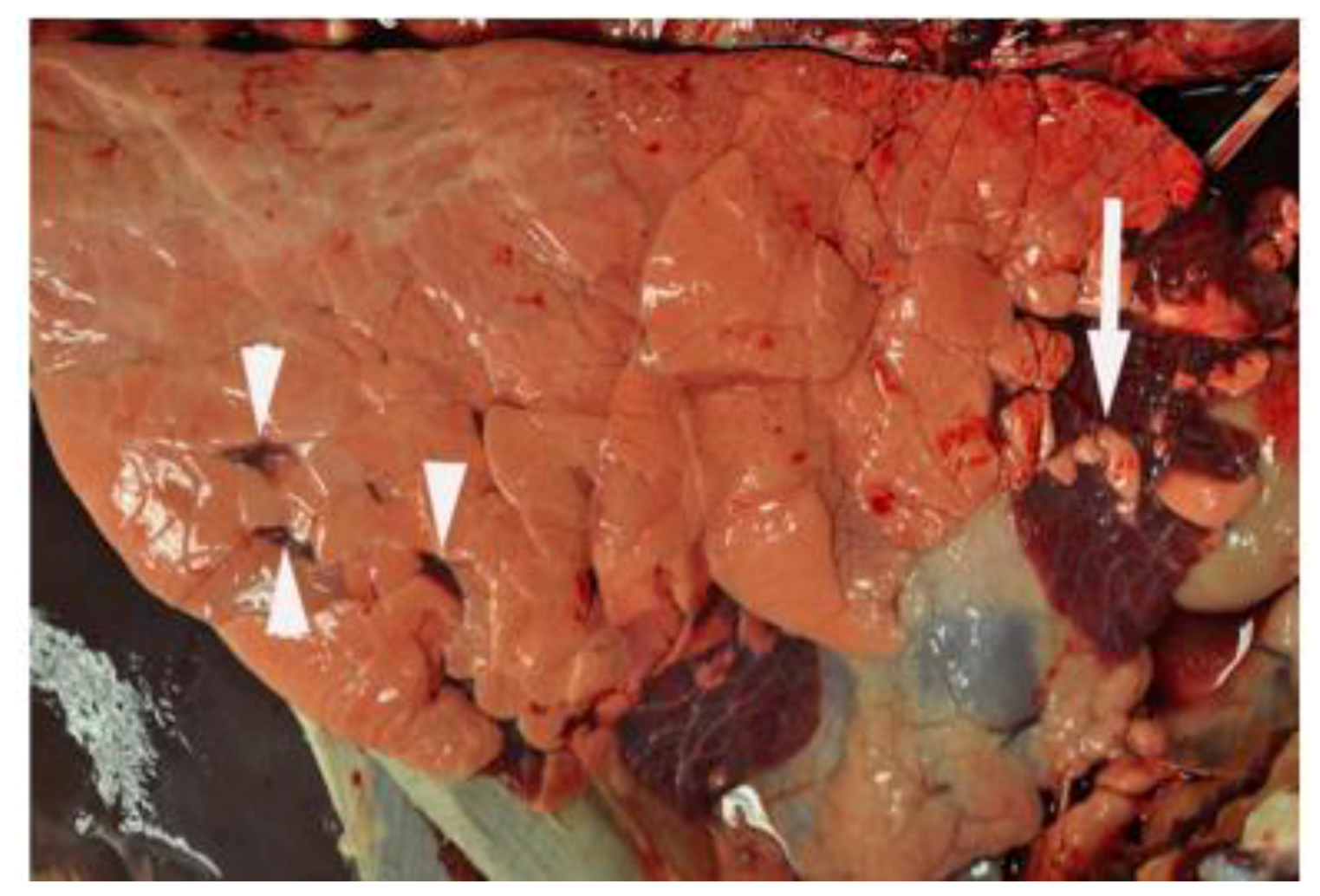
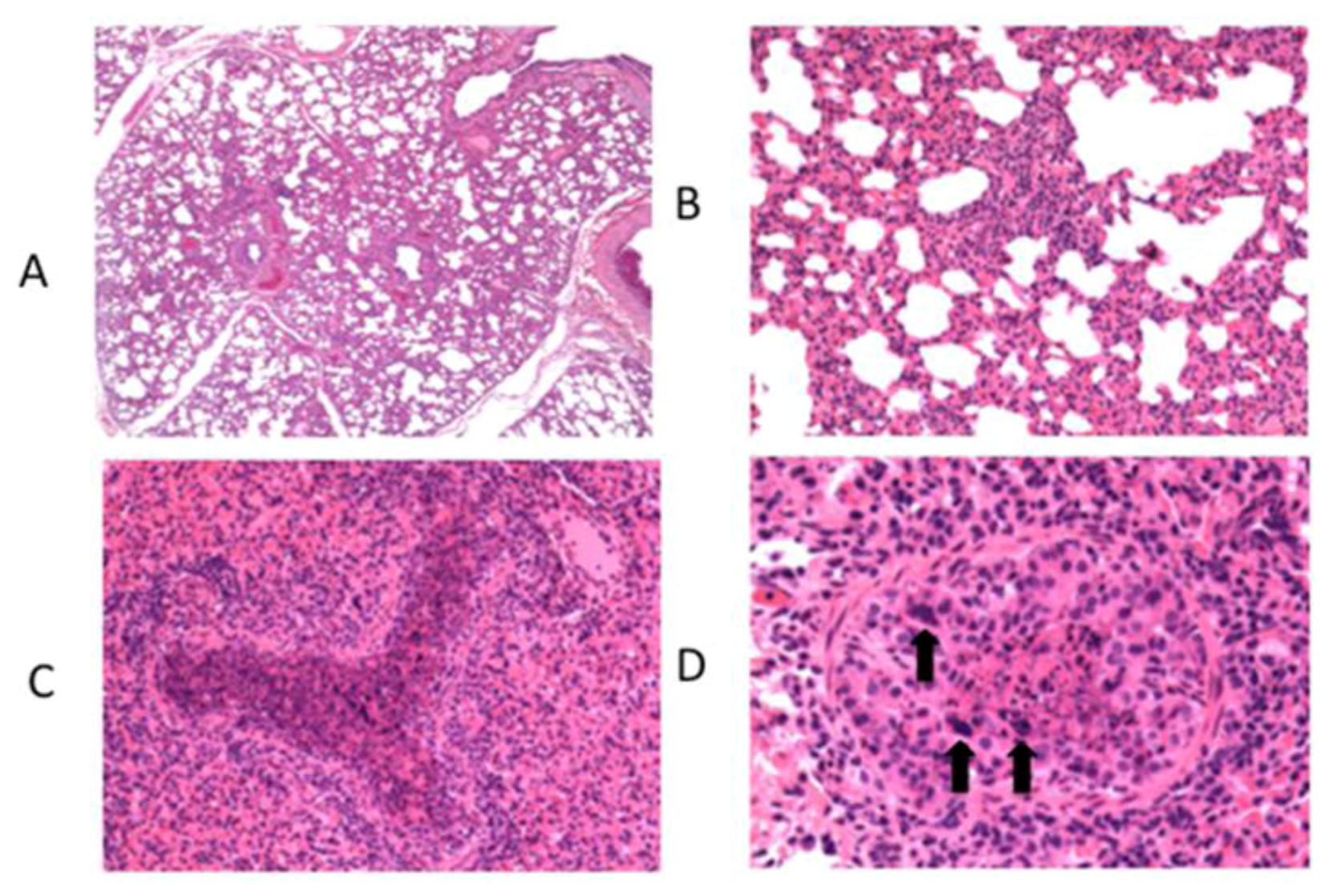
6. Innate Immunity to RSV
7. Adaptive Immunity to RSV
8. RSV Vaccines
9. Potential Role of Vitamin D as an Immunomodulator During RSV Infection
10. Summary

{kind=link}
{kind=link}
| Feature | Human RSV | Bovine RSV | References |
|---|---|---|---|
| Age-dependency | More prevalent in children <2 yrs old | More prevalent in calves <6 mo old | [16,17] |
| Seasonal periodicity | More common in fall and winter | More common in fall and winter | [18] |
| Histopathology | Bronchiolitis; interstitial pneumonia; | Bronchiolitis; interstitial pneumonia; | [31,33,34,35] |
| Prominent neutrophil and macrophage infiltration | Prominent neutrophil and macrophage infiltration | ||
| CXC chemokines | CXCL8 (IL-8) upregulated | CXCL8 (IL-8) upregulated | [31,139] |
| Adaptive immunity | Th2 cytokine bias | Th2 cytokine bias | [67,68,70,71,72] |
| Vaccine-enhanced disease | Observed with formalin-inactivated vaccine | Observed with formalin-inactivated vaccine | [109,110,111,112,113] |
Acknowledgments
Conflict of Interest
References
- Nair, H.; Nokes, D.J.; Gessner, B.D.; Dherani, M.; Madhi, S.A.; Singleton, R.J.; O'Brien, K.L.; Roca, A.; Wright, P.F.; Bruce, N.; et al. Global burden of acute lower respiratory infections due to respiratory syncytial virus in young children: a systematic review and meta-analysis. Lancet 2010, 375, 1545–1555. [Google Scholar] [CrossRef]
- Silvestri, M.; Sabatini, F.; Defilippi, A.C.; Rossi, G.A. The wheezy infant -- immunological and molecular considerations. Paediatr. Respir. Rev. 2004, 5 Suppl A, S81–S87. [Google Scholar] [CrossRef]
- Stein, R.T.; Sherrill, D.; Morgan, W.J.; Holberg, C.J.; Halonen, M.; Taussig, L.M.; Wright, A.L.; Martinez, F.D. Respiratory syncytial virus in early life and risk of wheeze and allergy by age 13 years. Lancet 1999, 354, 541–545. [Google Scholar]
- Falsey, A.R.; Hennessey, P.A.; Formica, M.A.; Cox, C.; Walsh, E.E. Respiratory syncytial virus infection in elderly and high-risk adults. N. Engl. J. Med. 2005, 352, 1749–1759. [Google Scholar] [CrossRef]
- Falsey, A.R.; Walsh, E.E. Respiratory syncytial virus infection in adults. Clin. Microbiol. Rev. 2000, 13, 371–384. [Google Scholar] [CrossRef]
- Raboni, S.M.; Nogueira, M.B.; Tsuchiya, L.R.; Takahashi, G.A.; Pereira, L.A.; Pasquini, R.; Siqueira, M.M. Respiratory tract viral infections in bone marrow transplant patients. Transplantation 2003, 76, 142–146. [Google Scholar] [CrossRef]
- Gershwin, L.J. Bovine respiratory syncytial virus infection: immunopathogenic mechanisms. Anim. Health Res. Rev. 2007, 8, 207–213. [Google Scholar] [CrossRef]
- Blount, R.E., Jr.; Morris, J.A.; Savage, R.E. Recovery of cytopathogenic agent from chimpanzees with coryza. Proc. Soc. Exp. Biol. Med. 1956, 92, 544–549. [Google Scholar]
- Chanock, R.; Finberg, L. Recovery from infants with respiratory illness of a virus related to chimpanzee coryza agent (CCA). II. Epidemiologic aspects of infection in infants and young children. Am. J. Hyg. 1957, 66, 291–300. [Google Scholar]
- Chanock, R.; Roizman, B.; Myers, R. Recovery from infants with respiratory illness of a virus related to chimpanzee coryza agent (CCA). I. Isolation, properties and characterization. Am. J. Hyg. 1957, 66, 281–290. [Google Scholar]
- Doggett, J.E.; Taylor-Robinson, D.; Gallop, R.G. A study of an inhibitor in bovine serum active against respiratory syncytial virus. Arch. Gesamte Virusforsch 1968, 23, 126–137. [Google Scholar]
- Paccaud, M.F.; Jacquier, C. A respiratory syncytial virus of bovine origin. Arch. Gesamte Virusforsch 1970, 30, 327–342. [Google Scholar] [CrossRef]
- Jacobs, J.W.; Edington, N. Isolation of respiratory syncytial virus from cattle in Britain. Vet. Rec. 1971, 88, 694. [Google Scholar]
- Inaba, Y.; Tanaka, Y.; Sato, K.; Ito, H.; Omori, T. Nomi virus, a virus isolated from an apparently new epizootic respiratory disease of cattle. Jpn. J. Microbiol. 1970, 14, 246–248. [Google Scholar]
- Inaba, Y.; Tanaka, Y.; Omori, T.; Matumoto, M. Isolation of bovine respiratory syncytial virus. Jpn J. Exp. Med. 1970, 40, 473–474. [Google Scholar]
- Glezen, W.P.; Taber, L.H.; Frank, A.L.; Kasel, J.A. Risk of primary infection and reinfection with respiratory syncytial virus. Am. J. Dis. Child. 1986, 140, 543–546. [Google Scholar]
- Valarcher, J.F.; Schelcher, F.; Bourhy, H. Evolution of bovine respiratory syncytial virus. J. Virol. 2000, 74, 10714–10728. [Google Scholar] [CrossRef]
- Van der Poel, W.H.; Brand, A.; Kramps, J.A.; Van Oirschot, J.T. Respiratory syncytial virus infections in human beings and in cattle. J. Infect. 1994, 29, 215–228. [Google Scholar] [CrossRef]
- Anderson, L.J.; Hierholzer, J.C.; Tsou, C.; Hendry, R.M.; Fernie, B.F.; Stone, Y.; McIntosh, K. Antigenic characterization of respiratory syncytial virus strains with monoclonal antibodies. J. Infect. Dis. 1985, 151, 626–633. [Google Scholar]
- Cristina, J.; Lopez, J.A.; Albo, C.; Garcia-Barreno, B.; Garcia, J.; Melero, J.A.; Portela, A. Analysis of genetic variability in human respiratory syncytial virus by the RNase A mismatch cleavage method: subtype divergence and heterogeneity. Virology 1990, 174, 126–134. [Google Scholar] [CrossRef]
- Furze, J.M.; Roberts, S.R.; Wertz, G.W.; Taylor, G. Antigenically distinct G glycoproteins of BRSV strains share a high degree of genetic homogeneity. Virology 1997, 231, 48–58. [Google Scholar] [CrossRef]
- Deplanche, M.; Lemaire, M.; Mirandette, C.; Bonnet, M.; Schelcher, F.; Meyer, G. In vivo evidence for quasispecies distributions in the bovine respiratory syncytial virus genome. J. Gen. Virol. 2007, 88, 1260–1265. [Google Scholar] [CrossRef]
- McLellan, J.S.; Yang, Y.; Graham, B.S.; Kwong, P.D. Structure of respiratory syncytial virus fusion glycoprotein in the postfusion conformation reveals preservation of neutralizing epitopes. J. Virol. 2011, 85, 7788–7796. [Google Scholar]
- Johnson, P.R.; Spriggs, M.K.; Olmsted, R.A.; Collins, P.L. The G glycoprotein of human respiratory syncytial viruses of subgroups A and B: extensive sequence divergence between antigenically related proteins. Proc. Natl. Acad. Sci. U. S. A. 1987, 84, 5625–5629. [Google Scholar] [CrossRef]
- Valarcher, J.F.; Schelcher, F.; Bourhy, H. Evolution of bovine respiratory syncytial virus. J. Virol. 2000, 74, 10714–10728. [Google Scholar]
- Garcia, O.; Martin, M.; Dopazo, J.; Arbiza, J.; Frabasile, S.; Russi, J.; Hortal, M.; Perez-Brena, P.; Martinez, I.; Garcia-Barreno, B.; et al. Evolutionary pattern of human respiratory syncytial virus (subgroup A): cocirculating lineages and correlation of genetic and antigenic changes in the G glycoprotein. J. Virol. 1994, 68, 5448–5459. [Google Scholar]
- Woelk, C.H.; Holmes, E.C. Variable immune-driven natural selection in the attachment (G) glycoprotein of respiratory syncytial virus (RSV). J. Mol. Evol. 2001, 52, 182–192. [Google Scholar]
- Prozzi, D.; Walravens, K.; Langedijk, J.P.; Daus, F.; Kramps, J.A.; Letesson, J.J. Antigenic and molecular analyses of the variability of bovine respiratory syncytial virus G glycoprotein. J. Gen. Virol. 1997, 78 ( Pt 2), 359–366. [Google Scholar]
- Fach, S.J.; Meyerholz, D.K.; Gallup, J.M.; Ackermann, M.R.; Lehmkuhl, H.D.; Sacco, R.E. Neonatal ovine pulmonary dendritic cells support bovine respiratory syncytial virus replication with enhanced interleukin (IL)-4 And IL-10 gene transcripts. Viral. Immunol. 2007, 20, 119–130. [Google Scholar] [CrossRef]
- Meyerholz, D.K.; Grubor, B.; Fach, S.J.; Sacco, R.E.; Lehmkuhl, H.D.; Gallup, J.M.; Ackermann, M.R. Reduced clearance of respiratory syncytial virus infection in a preterm lamb model. Microbes. Infect. 2004, 6, 1312–1319. [Google Scholar]
- Sacco, R.E.; Nonnecke, B.J.; Palmer, M.V.; Waters, W.R.; Lippolis, J.D.; Reinhardt, T.A. Differential expression of cytokines in response to respiratory syncytial virus infection of calves with high or low circulating 25-hydroxyvitamin D3. PLoS One 2012, 7, e33074. [Google Scholar]
- Woolums, A.R.; Anderson, M.L.; Gunther, R.A.; Schelegle, E.S.; LaRochelle, D.R.; Singer, R.S.; Boyle, G.A.; Friebertshauser, K.E.; Gershwin, L.J. Evaluation of severe disease induced by aerosol inoculation of calves with bovine respiratory syncytial virus. Am J. Vet. Res. 1999, 60, 473–480. [Google Scholar]
- Aherne, W.; Bird, T.; Court, S.D.; Gardner, P.S.; McQuillin, J. Pathological changes in virus infections of the lower respiratory tract in children. J. Clin. Pathol. 1970, 23, 7–18. [Google Scholar] [CrossRef]
- Bem, R.A.; Domachowske, J.B.; Rosenberg, H.F. Animal models of human respiratory syncytial virus disease. Am. J. Physiol. Lung Cell. Mol. Physiol. 2011, 301, L148–L156. [Google Scholar] [CrossRef]
- Johnson, J.E.; Gonzales, R.A.; Olson, S.J.; Wright, P.F.; Graham, B.S. The histopathology of fatal untreated human respiratory syncytial virus infection. Mod. Pathol. 2007, 20, 108–119. [Google Scholar]
- Bendelja, K.; Vojvoda, V.; Aberle, N.; Cepin-Bogovic, J.; Gagro, A.; Mlinaric-Galinovic, G.; Rabatic, S. Decreased Toll-like receptor 8 expression and lower TNF-alpha synthesis in infants with acute RSV infection. Respir. Res. 2010, 11, 143. [Google Scholar] [CrossRef]
- Zeng, R.; Cui, Y.; Hai, Y.; Liu, Y. Pattern recognition receptors for respiratory syncytial virus infection and design of vaccines. Virus Res. 2012, 167, 138–145. [Google Scholar]
- Ning, S.; Pagano, J.S.; Barber, G.N. IRF7: activation, regulation, modification and function. Genes Immun. 2011, 12, 399–414. [Google Scholar] [CrossRef]
- Yang, H.; Lin, C.H.; Ma, G.; Baffi, M.O.; Wathelet, M.G. Interferon regulatory factor-7 synergizes with other transcription factors through multiple interactions with p300/CBP coactivators. J. Biol. Chem. 2003, 278, 15495–15504. [Google Scholar]
- Bossert, B.; Conzelmann, K.K. Respiratory syncytial virus (RSV) nonstructural (NS) proteins as host range determinants: a chimeric bovine RSV with NS genes from human RSV is attenuated in interferon-competent bovine cells. J. Virol. 2002, 76, 4287–4293. [Google Scholar] [CrossRef]
- Schlender, J.; Bossert, B.; Buchholz, U.; Conzelmann, K.K. Bovine respiratory syncytial virus nonstructural proteins NS1 and NS2 cooperatively antagonize alpha/beta interferon-induced antiviral response. J. Virol. 2000, 74, 8234–8242. [Google Scholar]
- Bossert, B.; Marozin, S.; Conzelmann, K.K. Nonstructural proteins NS1 and NS2 of bovine respiratory syncytial virus block activation of interferon regulatory factor 3. J. Virol. 2003, 77, 8661–8668. [Google Scholar] [CrossRef]
- Lo, M.S.; Brazas, R.M.; Holtzman, M.J. Respiratory syncytial virus nonstructural proteins NS1 and NS2 mediate inhibition of Stat2 expression and alpha/beta interferon responsiveness. J. Virol. 2005, 79, 9315–9319. [Google Scholar] [CrossRef]
- Liu, P.; Jamaluddin, M.; Li, K.; Garofalo, R.P.; Casola, A.; Brasier, A.R. Retinoic acid-inducible gene I mediates early antiviral response and Toll-like receptor 3 expression in respiratory syncytial virus-infected airway epithelial cells. J. Virol. 2007, 81, 1401–1411. [Google Scholar]
- Ferrick, D.A.; King, D.P.; Jackson, K.A.; Braun, R.K.; Tam, S.; Hyde, D.M.; Beaman, B.L. Intraepithelial gamma delta T lymphocytes: sentinel cells at mucosal barriers. Springer Semin. Immunopathol. 2000, 22, 283–296. [Google Scholar]
- Hayday, A.C. gamma][delta] cells: a right time and a right place for a conserved third way of protection. Annu. Rev. Immunol. 2000, 18, 975–1026. [Google Scholar] [CrossRef]
- Hein, W.R.; Mackay, C.R. Prominence of gamma delta T cells in the ruminant immune system. Immunol. Today 1991, 12, 30–34. [Google Scholar]
- Jutila, M.A.; Holderness, J.; Graff, J.C.; Hedges, J.F. Antigen-independent priming: a transitional response of bovine gammadelta T-cells to infection. Anim. Health Res. Rev. 2008, 9, 47–57. [Google Scholar] [CrossRef]
- De Weerd, W.; Twilhaar, W.N.; Kimpen, J.L. T cell subset analysis in peripheral blood of children with RSV bronchiolitis. Scand. J. Infect. Dis. 1998, 30, 77–80. [Google Scholar]
- Aoyagi, M.; Shimojo, N.; Sekine, K.; Nishimuta, T.; Kohno, Y. Respiratory syncytial virus infection suppresses IFN-gamma production of gammadelta T cells. Clin. Exp. Immunol. 2003, 131, 312–317. [Google Scholar] [CrossRef]
- Dodd, J.; Riffault, S.; Kodituwakku, J.S.; Hayday, A.C.; Openshaw, P.J. Pulmonary V gamma 4+ gamma delta T cells have proinflammatory and antiviral effects in viral lung disease. J. Immunol. 2009, 182, 1174–1181. [Google Scholar]
- Taylor, G.; Thomas, L.H.; Wyld, S.G.; Furze, J.; Sopp, P.; Howard, C.J. Role of T-lymphocyte subsets in recovery from respiratory syncytial virus infection in calves. J. Virol. 1995, 69, 6658–6664. [Google Scholar]
- Thomas, L.H.; Cook, R.S.; Howard, C.J.; Gaddum, R.M.; Taylor, G. Influence of selective T-lymphocyte depletion on the lung pathology of gnotobiotic calves and the distribution of different T-lymphocyte subsets following challenge with bovine respiratory syncytial virus. Res. Vet. Sci. 1996, 61, 38–44. [Google Scholar] [CrossRef]
- McGill, J.L.; Nonnecke, B.J.; Lippolis, J.D.; Reinhardt, T.A.; Sacco, R.E. Differential chemokine and cytokine production by neonatal bovine γδ T cell subsets in response to viral toll-like receptor agonists and in vivo RSV infection. Immunology. Manuscript under review.
- Meyer, G.; Deplanche, M.; Schelcher, F. Human and bovine respiratory syncytial virus vaccine research and development. Comp. Immunol. Microbiol. Infect. Dis. 2008, 31, 191–225. [Google Scholar]
- Bueno, S.M.; Gonzalez, P.A.; Pacheco, R.; Leiva, E.D.; Cautivo, K.M.; Tobar, H.E.; Mora, J.E.; Prado, C.E.; Zuniga, J.P.; Jimenez, J.; et al. Host immunity during RSV pathogenesis. Int. Immunopharmacol. 2008, 8, 1320–1329. [Google Scholar] [CrossRef]
- van Bleek, G.M.; Poelen, M.C.; van der Most, R.; Brugghe, H.F.; Timmermans, H.A.; Boog, C.J.; Hoogerhout, P.; Otten, H.G.; van Els, C.A. Identification of immunodominant epitopes derived from the respiratory syncytial virus fusion protein that are recognized by human CD4 T cells. J. Virol. 2003, 77, 980–988. [Google Scholar]
- Cannon, M.J.; Openshaw, P.J.; Askonas, B.A. Cytotoxic T cells clear virus but augment lung pathology in mice infected with respiratory syncytial virus. J. Exp. Med. 1988, 168, 1163–1168. [Google Scholar] [CrossRef]
- Graham, B.S.; Bunton, L.A.; Wright, P.F.; Karzon, D.T. Role of T lymphocyte subsets in the pathogenesis of primary infection and rechallenge with respiratory syncytial virus in mice. J. Clin. Invest. 1991, 88, 1026–1033. [Google Scholar]
- Graham, B.S.; Bunton, L.A.; Rowland, J.; Wright, P.F.; Karzon, D.T. Respiratory syncytial virus infection in anti-mu-treated mice. J. Virol. 1991, 65, 4936–4942. [Google Scholar]
- Openshaw, P.J.; Tregoning, J.S. Immune responses and disease enhancement during respiratory syncytial virus infection. Clin. Microbiol. Rev. 2005, 18, 541–555. [Google Scholar] [CrossRef]
- Guerrero-Plata, A.; Casola, A.; Suarez, G.; Yu, X.; Spetch, L.; Peeples, M.E.; Garofalo, R.P. Differential response of dendritic cells to human metapneumovirus and respiratory syncytial virus. Am. J. Respir. Cell. Mol. Biol. 2006, 34, 320–329. [Google Scholar]
- Bartz, H.; Turkel, O.; Hoffjan, S.; Rothoeft, T.; Gonschorek, A.; Schauer, U. Respiratory syncytial virus decreases the capacity of myeloid dendritic cells to induce interferon-gamma in naive T cells. Immunology 2003, 109, 49–57. [Google Scholar] [CrossRef]
- Boelen, A.; Kwakkel, J.; Barends, M.; de Rond, L.; Dormans, J.; Kimman, T. Effect of lack of Interleukin-4, Interleukin-12, Interleukin-18, or the Interferon-gamma receptor on virus replication, cytokine response, and lung pathology during respiratory syncytial virus infection in mice. J Med Virol 2002, 66, 552–560. [Google Scholar] [CrossRef]
- Hussell, T.; Baldwin, C.J.; O'Garra, A.; Openshaw, P.J. CD8+ T cells control Th2-driven pathology during pulmonary respiratory syncytial virus infection. Eur. J. Immunol. 1997, 27, 3341–3349. [Google Scholar]
- Durbin, J.E.; Johnson, T.R.; Durbin, R.K.; Mertz, S.E.; Morotti, R.A.; Peebles, R.S.; Graham, B.S. The role of IFN in respiratory syncytial virus pathogenesis. J. Immunol. 2002, 168, 2944–2952. [Google Scholar]
- Lindemans, C.A.; Kimpen, J.L.; Luijk, B.; Heidema, J.; Kanters, D.; van der Ent, C.K.; Koenderman, L. Systemic eosinophil response induced by respiratory syncytial virus. Clin. Exp. Immunol. 2006, 144, 409–417. [Google Scholar] [CrossRef]
- Kristjansson, S.; Bjarnarson, S.P.; Wennergren, G.; Palsdottir, A.H.; Arnadottir, T.; Haraldsson, A.; Jonsdottir, I. Respiratory syncytial virus and other respiratory viruses during the first 3 months of life promote a local TH2-like response. J. Allergy Clin. Immunol. 2005, 116, 805–811. [Google Scholar]
- van Benten, I.J.; van Drunen, C.M.; Koevoet, J.L.; Koopman, L.P.; Hop, W.C.; Osterhaus, A.D.; Neijens, H.J.; Fokkens, W.J. Reduced nasal IL-10 and enhanced TNFalpha responses during rhinovirus and RSV-induced upper respiratory tract infection in atopic and non-atopic infants. J. Med. Virol. 2005, 75, 348–357. [Google Scholar] [CrossRef]
- Miao, C.; Woolums, A.R.; Zarlenga, D.S.; Brown, C.C.; Brown, J.C., Jr.; Williams, S.M.; Scott, M.A. Effects of a single intranasal dose of modified-live bovine respiratory syncytial virus vaccine on cytokine messenger RNA expression following viral challenge in calves. Am. J. Vet. Res. 2004, 65, 725–733. [Google Scholar] [CrossRef]
- Stewart, R.S.; Gershwin, L.J. Role of IgE in the pathogenesis of bovine respiratory syncytial virus in sequential infections in vaccinated and nonvaccinated calves. Am. J. Vet. Res. 1989, 50, 349–355. [Google Scholar]
- Stewart, R.S.; Gershwin, L.J. Detection of IgE antibodies to bovine respiratory syncytial virus. Vet. Immunol. Immunopathol. 1989, 20, 313–323. [Google Scholar] [CrossRef]
- Gershwin, L.J.; Gunther, R.A.; Anderson, M.L.; Woolums, A.R.; McArthur-Vaughan, K.; Randel, K.E.; Boyle, G.A.; Friebertshauser, K.E.; McInturff, P.S. Bovine respiratory syncytial virus-specific IgE is associated with interleukin-2 and -4, and interferon-gamma expression in pulmonary lymph of experimentally infected calves. Am. J. Vet. Res. 2000, 61, 291–298. [Google Scholar] [CrossRef]
- Sigurs, N.; Gustafsson, P.M.; Bjarnason, R.; Lundberg, F.; Schmidt, S.; Sigurbergsson, F.; Kjellman, B. Severe respiratory syncytial virus bronchiolitis in infancy and asthma and allergy at age 13. Am. J. Respir. Crit. Care Med. 2005, 171, 137–141. [Google Scholar]
- Sigurs, N.; Bjarnason, R.; Sigurbergsson, F.; Kjellman, B.; Bjorksten, B. Asthma and immunoglobulin E antibodies after respiratory syncytial virus bronchiolitis: a prospective cohort study with matched controls. Pediatrics 1995, 95, 500–505. [Google Scholar]
- Gershwin, L.J.; Gunther, R.A.; Hornof, W.J.; Larson, R.F. Effect of infection with bovine respiratory syncytial virus on pulmonary clearance of an inhaled antigen in calves. Am. J. Vet. Res. 2008, 69, 416–422. [Google Scholar] [CrossRef]
- Gershwin, L.J.; Anderson, M.L.; Wang, C.; Berghaus, L.J.; Kenny, T.P.; Gunther, R.A. Assessment of IgE response and cytokine gene expression in pulmonary efferent lymph collected after ovalbumin inhalation during experimental infection of calves with bovine respiratory syncytial virus. Am. J. Vet. Res. 2011, 72, 134–145. [Google Scholar]
- Heidema, J.; Lukens, M.V.; van Maren, W.W.; van Dijk, M.E.; Otten, H.G.; van Vught, A.J.; van der Werff, D.B.; van Gestel, S.J.; Semple, M.G.; Smyth, R.L.; et al. CD8+ T cell responses in bronchoalveolar lavage fluid and peripheral blood mononuclear cells of infants with severe primary respiratory syncytial virus infections. J. Immunol. 2007, 179, 8410–8417. [Google Scholar]
- McInnes, E.; Sopp, P.; Howard, C.J.; Taylor, G. Phenotypic analysis of local cellular responses in calves infected with bovine respiratory syncytial virus. Immunology 1999, 96, 396–403. [Google Scholar]
- Openshaw, P.J.; Anderson, K.; Wertz, G.W.; Askonas, B.A. The 22,000-kilodalton protein of respiratory syncytial virus is a major target for Kd-restricted cytotoxic T lymphocytes from mice primed by infection. J. Virol. 1990, 64, 1683–1689. [Google Scholar]
- Cherrie, A.H.; Anderson, K.; Wertz, G.W.; Openshaw, P.J. Human cytotoxic T cells stimulated by antigen on dendritic cells recognize the N, SH, F, M, 22K, and 1b proteins of respiratory syncytial virus. J. Virol. 1992, 66, 2102–2110. [Google Scholar]
- Bangham, C.R.; McMichael, A.J. Specific human cytotoxic T cells recognize B-cell lines persistently infected with respiratory syncytial virus. Proc. Natl. Acad. Sci. U. S. A. 1986, 83, 9183–9187. [Google Scholar] [CrossRef]
- Gaddum, R.M.; Cook, R.S.; Furze, J.M.; Ellis, S.A.; Taylor, G. Recognition of bovine respiratory syncytial virus proteins by bovine CD8+ T lymphocytes. Immunology 2003, 108, 220–229. [Google Scholar]
- Taylor, G.; Thomas, L.H.; Furze, J.M.; Cook, R.S.; Wyld, S.G.; Lerch, R.; Hardy, R.; Wertz, G.W. Recombinant vaccinia viruses expressing the F, G or N, but not the M2, protein of bovine respiratory syncytial virus (BRSV) induce resistance to BRSV challenge in the calf and protect against the development of pneumonic lesions. J. Gen. Virol. 1997, 78 ( Pt 12), 3195–3206. [Google Scholar]
- Antonis, A.F.; Claassen, E.A.; Hensen, E.J.; de Groot, R.J.; de Groot-Mijnes, J.D.; Schrijver, R.S.; van der Most, R.G. Kinetics of antiviral CD8 T cell responses during primary and post-vaccination secondary bovine respiratory syncytial virus infection. Vaccine 2006, 24, 1551–1561. [Google Scholar] [CrossRef]
- Chang, J.; Braciale, T.J. Respiratory syncytial virus infection suppresses lung CD8+ T-cell effector activity and peripheral CD8+ T-cell memory in the respiratory tract. Nat. Med. 2002, 8, 54–60. [Google Scholar]
- Chang, J.; Srikiatkhachorn, A.; Braciale, T.J. Visualization and characterization of respiratory syncytial virus F-specific CD8(+) T cells during experimental virus infection. J. Immunol. 2001, 167, 4254–4260. [Google Scholar]
- Gaddum, R.M.; Cook, R.S.; Thomas, L.H.; Taylor, G. Primary cytotoxic T-cell responses to bovine respiratory syncytial virus in calves. Immunology 1996, 88, 421–427. [Google Scholar]
- Woolums, A.R.; Gunther, R.A.; McArthur-Vaughan, K.; Anderson, M.L.; Omlor, A.; Boyle, G.A.; Friebertshauser, K.E.; McInturff, P.S.; Gershwin, L.J. Cytotoxic T lymphocyte activity and cytokine expression in calves vaccinated with formalin-inactivated bovine respiratory syncytial virus prior to challenge. Comp. Immunol. Microbiol. Infect. Dis. 2004, 27, 57–74. [Google Scholar]
- Piedra, P.A.; Jewell, A.M.; Cron, S.G.; Atmar, R.L.; Glezen, W.P. Correlates of immunity to respiratory syncytial virus (RSV) associated-hospitalization: establishment of minimum protective threshold levels of serum neutralizing antibodies. Vaccine 2003, 21, 3479–3482. [Google Scholar] [CrossRef]
- Stensballe, L.G.; Ravn, H.; Kristensen, K.; Meakins, T.; Aaby, P.; Simoes, E.A. Seasonal variation of maternally derived respiratory syncytial virus antibodies and association with infant hospitalizations for respiratory syncytial virus. J. Pediatr. 2009, 154, 296–298. [Google Scholar] [CrossRef]
- Stensballe, L.G.; Ravn, H.; Kristensen, K.; Agerskov, K.; Meakins, T.; Aaby, P.; Simoes, E.A. Respiratory syncytial virus neutralizing antibodies in cord blood, respiratory syncytial virus hospitalization, and recurrent wheeze. J. Allergy. Clin. Immunol. 2009, 123, 398–403. [Google Scholar] [CrossRef]
- Kimman, T.G.; Zimmer, G.M.; Westenbrink, F.; Mars, J.; van Leeuwen, E. Epidemiological study of bovine respiratory syncytial virus infections in calves: influence of maternal antibodies on the outcome of disease. Vet. Rec. 1988, 123, 104–109. [Google Scholar]
- Crowe, J.E., Jr.; Williams, J.V. Immunology of viral respiratory tract infection in infancy. Paediatr Respir. Rev. 2003, 4, 112–119. [Google Scholar]
- Kimman, T.G.; Westenbrink, F.; Schreuder, B.E.; Straver, P.J. Local and systemic antibody response to bovine respiratory syncytial virus infection and reinfection in calves with and without maternal antibodies. J. Clin. Microbiol. 1987, 25, 1097–1106. [Google Scholar]
- Groothuis, J.R. The role of RSV neutralizing antibodies in the treatment and prevention of respiratory syncytial virus infection in high-risk children. Antiviral Res. 1994, 23, 1–10. [Google Scholar]
- Connors, M.; Collins, P.L.; Firestone, C.Y.; Murphy, B.R. Respiratory syncytial virus (RSV) F, G, M2 (22K), and N proteins each induce resistance to RSV challenge, but resistance induced by M2 and N proteins is relatively short-lived. J. Virol. 1991, 65, 1634–1637. [Google Scholar]
- Oshansky, C.M.; Zhang, W.; Moore, E.; Tripp, R.A. The host response and molecular pathogenesis associated with respiratory syncytial virus infection. Future Microbiol. 2009, 4, 279–297. [Google Scholar]
- Melero, J.A.; Garcia-Barreno, B.; Martinez, I.; Pringle, C.R.; Cane, P.A. Antigenic structure, evolution and immunobiology of human respiratory syncytial virus attachment (G) protein. J. Gen. Virol. 1997, 78 ( Pt 10), 2411–2418. [Google Scholar]
- Bukreyev, A.; Yang, L.; Fricke, J.; Cheng, L.; Ward, J.M.; Murphy, B.R.; Collins, P.L. The secreted form of respiratory syncytial virus G glycoprotein helps the virus evade antibody-mediated restriction of replication by acting as an antigen decoy and through effects on Fc receptor-bearing leukocytes. J. Virol. 2008, 82, 12191–12204. [Google Scholar] [CrossRef]
- Graham, B.S. Biological challenges and technological opportunities for respiratory syncytial virus vaccine development. Immunol. Rev. 2011, 239, 149–166. [Google Scholar]
- Mills, J.T.; Van Kirk, J.E.; Wright, P.F.; Chanock, R.M. Experimental respiratory syncytial virus infection of adults. Possible mechanisms of resistance to infection and illness. J. Immunol. 1971, 107, 123–130. [Google Scholar]
- Prince, G.A.; Hemming, V.G.; Horswood, R.L.; Chanock, R.M. Immunoprophylaxis and immunotherapy of respiratory syncytial virus infection in the cotton rat. Virus Res. 1985, 3, 193–206. [Google Scholar] [CrossRef]
- Prince, G.A.; Horswood, R.L.; Chanock, R.M. Quantitative aspects of passive immunity to respiratory syncytial virus infection in infant cotton rats. J. Virol. 1985, 55, 517–520. [Google Scholar]
- Singleton, R.; Etchart, N.; Hou, S.; Hyland, L. Inability to evoke a long-lasting protective immune response to respiratory syncytial virus infection in mice correlates with ineffective nasal antibody responses. J. Virol. 2003, 77, 11303–11311. [Google Scholar]
- Singleton, R.; Dooley, L.; Bruden, D.; Raelson, S.; Butler, J.C. Impact of palivizumab prophylaxis on respiratory syncytial virus hospitalizations in high risk Alaska Native infants. Pediatr. Infect. Dis. J. 2003, 22, 540–545. [Google Scholar]
- Malley, R.; DeVincenzo, J.; Ramilo, O.; Dennehy, P.H.; Meissner, H.C.; Gruber, W.C.; Sanchez, P.J.; Jafri, H.; Balsley, J.; Carlin, D.; et al. Reduction of respiratory syncytial virus (RSV) in tracheal aspirates in intubated infants by use of humanized monoclonal antibody to RSV F protein. J. Infect. Dis. 1998, 178, 1555–1561. [Google Scholar] [CrossRef]
- Simoes, E.A.; Groothuis, J.R.; Carbonell-Estrany, X.; Rieger, C.H.; Mitchell, I.; Fredrick, L.M.; Kimpen, J.L. Palivizumab prophylaxis, respiratory syncytial virus, and subsequent recurrent wheezing. J. Pediatr. 2007, 151, 34–42, 42 e31.. [Google Scholar] [CrossRef]
- Chin, J.; Magoffin, R.L.; Shearer, L.A.; Schieble, J.H.; Lennette, E.H. Field evaluation of a respiratory syncytial virus vaccine and a trivalent parainfluenza virus vaccine in a pediatric population. Am. J. Epidemiol. 1969, 89, 449–463. [Google Scholar]
- Kapikian, A.Z.; Mitchell, R.H.; Chanock, R.M.; Shvedoff, R.A.; Stewart, C.E. An epidemiologic study of altered clinical reactivity to respiratory syncytial (RS) virus infection in children previously vaccinated with an inactivated RS virus vaccine. Am. J. Epidemiol. 1969, 89, 405–421. [Google Scholar]
- Kim, H.W.; Canchola, J.G.; Brandt, C.D.; Pyles, G.; Chanock, R.M.; Jensen, K.; Parrott, R.H. Respiratory syncytial virus disease in infants despite prior administration of antigenic inactivated vaccine. Am. J. Epidemiol. 1969, 89, 422–434. [Google Scholar]
- Kimman, T.G.; Sol, J.; Westenbrink, F.; Straver, P.J. A severe outbreak of respiratory tract disease associated with bovine respiratory syncytial virus probably enhanced by vaccination with modified live vaccine. Vet. Q. 1989, 11, 250–253. [Google Scholar] [CrossRef]
- Schreiber, P.; Matheise, J.P.; Dessy, F.; Heimann, M.; Letesson, J.J.; Coppe, P.; Collard, A. High mortality rate associated with bovine respiratory syncytial virus (BRSV) infection in Belgian white blue calves previously vaccinated with an inactivated BRSV vaccine. J. Vet. Med. B. Infect. Dis. Vet. Public Health 2000, 47, 535–550. [Google Scholar]
- Antonis, A.F.; Schrijver, R.S.; Daus, F.; Steverink, P.J.; Stockhofe, N.; Hensen, E.J.; Langedijk, J.P.; van der Most, R.G. Vaccine-induced immunopathology during bovine respiratory syncytial virus infection: exploring the parameters of pathogenesis. J. Virol. 2003, 77, 12067–12073. [Google Scholar]
- Gershwin, L.J.; Schelegle, E.S.; Gunther, R.A.; Anderson, M.L.; Woolums, A.R.; Larochelle, D.R.; Boyle, G.A.; Friebertshauser, K.E.; Singer, R.S. A bovine model of vaccine enhanced respiratory syncytial virus pathophysiology. Vaccine 1998, 16, 1225–1236. [Google Scholar] [CrossRef]
- West, K.; Petrie, L.; Haines, D.M.; Konoby, C.; Clark, E.G.; Martin, K.; Ellis, J.A. The effect of formalin-inactivated vaccine on respiratory disease associated with bovine respiratory syncytial virus infection in calves. Vaccine 1999, 17, 809–820. [Google Scholar] [CrossRef]
- Mohanty, S.B.; Rockemann, D.D.; Davidson, J.P.; Sharabrin, O.I.; Forst, S.M. Effect of vaccinal serum antibodies on bovine respiratory syncytial viral infection in calves. Am. J. Vet. Res. 1981, 42, 881–883. [Google Scholar]
- Kalina, W.V.; Woolums, A.R.; Berghaus, R.D.; Gershwin, L.J. Formalin-inactivated bovine RSV vaccine enhances a Th2 mediated immune response in infected cattle. Vaccine 2004, 22, 1465–1474. [Google Scholar] [CrossRef]
- Hurwitz, J.L. Respiratory syncytial virus vaccine development. Expert Rev. Vaccines 2011, 10, 1415–1433. [Google Scholar]
- Karron, R.A.; Wright, P.F.; Belshe, R.B.; Thumar, B.; Casey, R.; Newman, F.; Polack, F.P.; Randolph, V.B.; Deatly, A.; Hackell, J.; et al. Identification of a recombinant live attenuated respiratory syncytial virus vaccine candidate that is highly attenuated in infants. J. Infect. Dis. 2005, 191, 1093–1104. [Google Scholar] [CrossRef]
- Luongo, C.; Winter, C.C.; Collins, P.L.; Buchholz, U.J. Increased genetic and phenotypic stability of a promising live-attenuated respiratory syncytial virus vaccine candidate by reverse genetics. J. Virol. 2012, 86, 10792–10804. [Google Scholar]
- Tang, R.S.; MacPhail, M.; Schickli, J.H.; Kaur, J.; Robinson, C.L.; Lawlor, H.A.; Guzzetta, J.M.; Spaete, R.R.; Haller, A.A. Parainfluenza virus type 3 expressing the native or soluble fusion (F) Protein of Respiratory Syncytial Virus (RSV) confers protection from RSV infection in African green monkeys. J. Virol. 2004, 78, 11198–11207. [Google Scholar]
- Bernstein, D.I.; Malkin, E.; Abughali, N.; Falloon, J.; Yi, T.; Dubovsky, F. Phase 1 study of the safety and immunogenicity of a live, attenuated respiratory syncytial virus and parainfluenza virus type 3 vaccine in seronegative children. Pediatr. Infect. Dis. J. 2012, 31, 109–114. [Google Scholar] [CrossRef]
- Tang, R.S.; Spaete, R.R.; Thompson, M.W.; MacPhail, M.; Guzzetta, J.M.; Ryan, P.C.; Reisinger, K.; Chandler, P.; Hilty, M.; Walker, R.E.; et al. Development of a PIV-vectored RSV vaccine: preclinical evaluation of safety, toxicity, and enhanced disease and initial clinical testing in healthy adults. Vaccine 2008, 26, 6373–6382. [Google Scholar] [CrossRef]
- Valarcher, J.F.; Furze, J.; Wyld, S.; Cook, R.; Conzelmann, K.K.; Taylor, G. Role of alpha/beta interferons in the attenuation and immunogenicity of recombinant bovine respiratory syncytial viruses lacking NS proteins. J. Virol. 2003, 77, 8426–8439. [Google Scholar]
- Mapletoft, J.W.; Oumouna, M.; Townsend, H.G.; Gomis, S.; Babiuk, L.A.; van Drunen Littel-van den Hurk, S. Formulation with CpG oligodeoxynucleotides increases cellular immunity and protection induced by vaccination of calves with formalin-inactivated bovine respiratory syncytial virus. Virology 2006, 353, 316–323. [Google Scholar] [CrossRef]
- Hagglund, S.; Hu, K.F.; Larsen, L.E.; Hakhverdyan, M.; Valarcher, J.F.; Taylor, G.; Morein, B.; Belak, S.; Alenius, S. Bovine respiratory syncytial virus ISCOMs--protection in the presence of maternal antibodies. Vaccine 2004, 23, 646–655. [Google Scholar] [CrossRef]
- Hagglund, S.; Hu, K.; Vargmar, K.; Pore, L.; Olofson, A.S.; Blodorn, K.; Anderson, J.; Ahooghalandari, P.; Pringle, J.; Taylor, G.; et al. Bovine respiratory syncytial virus ISCOMs-Immunity, protection and safety in young conventional calves. Vaccine 2011, 29, 8719–8730. [Google Scholar] [CrossRef]
- Hewison, M. Vitamin D and innate and adaptive immunity. Vitam. Horm. 2011, 86, 23–62. [Google Scholar]
- Mora, J.R.; Iwata, M.; von Andrian, U.H. Vitamin effects on the immune system: vitamins A and D take centre stage. Nat. Rev. Immunol. 2008, 8, 685–698. [Google Scholar] [CrossRef]
- Adams, J.S.; Liu, P.T.; Chun, R.; Modlin, R.L.; Hewison, M. Vitamin D in defense of the human immune response. Ann. N. Y. Acad. Sci. 2007, 1117, 94–105. [Google Scholar] [CrossRef]
- Bikle, D.D. Vitamin D regulation of immune function. Vitam Horm 2011, 86, 1–21. [Google Scholar]
- Nelson, C.D.; Reinhardt, T.A.; Beitz, D.C.; Lippolis, J.D. In vivo activation of the intracrine vitamin D pathway in innate immune cells and mammary tissue during a bacterial infection. PLoS One 2010, 5, e15469. [Google Scholar]
- Nelson, C.D.; Reinhardt, T.A.; Thacker, T.C.; Beitz, D.C.; Lippolis, J.D. Modulation of the bovine innate immune response by production of 1alpha,25-dihydroxyvitamin D(3) in bovine monocytes. J. Dairy Sci. 2010, 93, 1041–1049. [Google Scholar] [CrossRef]
- Grant, C.C.; Wall, C.R.; Gibbons, M.J.; Morton, S.M.; Santosham, M.; Black, R.E. Child nutrition and lower respiratory tract disease burden in New Zealand: a global context for a national perspective. J. Paediatr. Child. Health 2011, 47, 497–504. [Google Scholar] [CrossRef]
- Karatekin, G.; Kaya, A.; Salihoglu, O.; Balci, H.; Nuhoglu, A. Association of subclinical vitamin D deficiency in newborns with acute lower respiratory infection and their mothers. Eur. J. Clin. Nutr. 2009, 63, 473–477. [Google Scholar] [CrossRef]
- Wayse, V.; Yousafzai, A.; Mogale, K.; Filteau, S. Association of subclinical vitamin D deficiency with severe acute lower respiratory infection in Indian children under 5 y. Eur. J. Clin. Nutr. 2004, 58, 563–567. [Google Scholar]
- Hansdottir, S.; Monick, M.M.; Lovan, N.; Powers, L.; Gerke, A.; Hunninghake, G.W. Vitamin D decreases respiratory syncytial virus induction of NF-kappaB-linked chemokines and cytokines in airway epithelium while maintaining the antiviral state. J. Immunol. 2010, 184, 965–974. [Google Scholar] [CrossRef]
- Abu-Harb, M.; Bell, F.; Finn, A.; Rao, W.H.; Nixon, L.; Shale, D.; Everard, M.L. IL-8 and neutrophil elastase levels in the respiratory tract of infants with RSV bronchiolitis. Eur. Respir. J. 1999, 14, 139–143. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Sacco, R.E.; McGill, J.L.; Palmer, M.V.; Lippolis, J.D.; Reinhardt, T.A.; Nonnecke, B.J. Neonatal Calf Infection with Respiratory Syncytial Virus: Drawing Parallels to the Disease in Human Infants. Viruses 2012, 4, 3731-3753. https://doi.org/10.3390/v4123731
Sacco RE, McGill JL, Palmer MV, Lippolis JD, Reinhardt TA, Nonnecke BJ. Neonatal Calf Infection with Respiratory Syncytial Virus: Drawing Parallels to the Disease in Human Infants. Viruses. 2012; 4(12):3731-3753. https://doi.org/10.3390/v4123731
Chicago/Turabian StyleSacco, Randy E., Jodi L. McGill, Mitchell V. Palmer, John D. Lippolis, Timothy A. Reinhardt, and Brian J. Nonnecke. 2012. "Neonatal Calf Infection with Respiratory Syncytial Virus: Drawing Parallels to the Disease in Human Infants" Viruses 4, no. 12: 3731-3753. https://doi.org/10.3390/v4123731
APA StyleSacco, R. E., McGill, J. L., Palmer, M. V., Lippolis, J. D., Reinhardt, T. A., & Nonnecke, B. J. (2012). Neonatal Calf Infection with Respiratory Syncytial Virus: Drawing Parallels to the Disease in Human Infants. Viruses, 4(12), 3731-3753. https://doi.org/10.3390/v4123731