Sharka: The Past, The Present and The Future



Abstract
:
1. Introduction


2. Plum pox virus

{kind=link}
{kind=link}
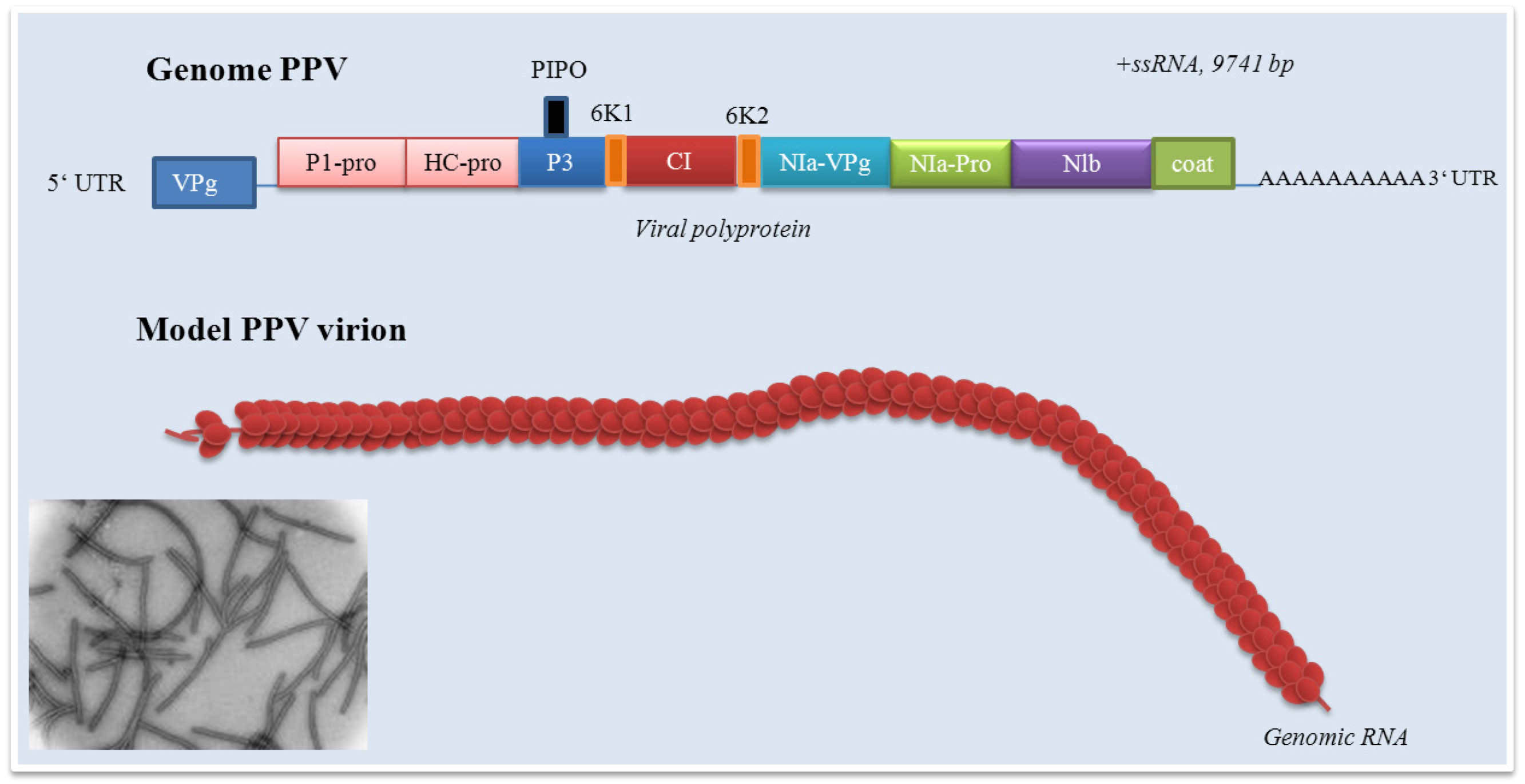{kind=link}
| Protein (size) | Function | Cellular localization | Reference(s) |
|---|---|---|---|
| P1 (32 – 64 K) | serine proteinase with cis-cleavage activity, P1/HCpro cleavage site. Responsible for symptomatolology. Usable for investigation of potyviruses phylogeny and relationship | Crystalline inclusions in cytoplasm and nuclei. | [56,61,63,168,169,170] |
| HCpro (56 – 58 K) | cysteine proteinase with cis-cleavage activity, HCpro/P3 cleavage site. Aphid transmission, systemic movement, suppression of gene silencing, symptoms development. | Cytoplasm, aggregates along the endoplasmic reticulum. | [82,171,172,173,174,175] |
| P3 (37 K) | plant pathogenicity – virulence, viral infection cycle. | Crystalline inclusions in cytoplasm and nuclei. Endoplasmic reticulum (punctuate inclusions in association with Golgi) | [86,91,92,169,176,177] |
| 6K1 (6 K?) | cell-to-cell movement?, role in virulence? | The cell periphery. | [94,178,179] |
| CI (70 K) | RNA helicase required for genome replication. Cell-to-cell movement. Long-distance movement. Development of symptoms. Breaking the host resistance. | Cytoplasm, conical structures attached to plasmodesmata. | [95,97,98,99,180] |
| 6K2 | anchoring the viral replication complex to membranes. Long-distance movement. Development of systemic infection. | Crystalline inclusions in cytoplasm and nuclei. | [103,167,169] |
| VPg | covalently links the 5´ end of the viral RNA via tyrosine residue. Viral cycle. Systemic infection. Overcoming the eIF4E-based recessive resistance. | Endoplasmic reticulum, nucleus, nucleolus. | [100,102,104,114,117] |
| NIa-Pro | NIa-pro cysteine proteinase. Host specificity. Host DNA cleavage activity. | Nuclei of infected cells in the form of inclusion bodies. | [71,75] |
| NIb | RNA-dependent RNA polymerase. Genome/viral replication. | Nuclei of infected cells in the form of inclusion bodies. Cytoplasm. | [116,127,131] |
| CP | Aphid transmission. Cell-to-cell and long-distance movement. Virion assembly. | Plasmodesmatal connections between infected leaf mesophyll cells. Cytoplasm of infected cells. | [96,144,145,146,181,182,183] |
| Disease status | Country | Reference(s) |
|---|---|---|
| Widespread (high level) | Croatia, Greece, Bulgaria, Germany, Hungary, Poland, Romania, Russia, Serbia, Slovakia | [197,198,199,200,201,202,203,204,205,206,207,208,209,210] |
| Restricted distribution(heterogeneous levels) | Albania, Argentina, Austria, Canada, Cyprus, Czech Republic, Egypt, France, Italy, Iran , Kazakhstan, Luxembourg, Moldova, Norway, Pakistan, Portugal, Southern Russia, Slovenia, Spain, Syria, Turkey, Ukraine, United Kingdom, United States | [192,195,197,208,211,212,213,214,215,216,217,218,219,220,221,222,223,224,225,226,227,228,229] |
| Introduced, established | Azores, Bosnia-Herzegovina, Chile, Some states of former USSR including Central Asia, India, Jordan, Lithuania, The Netherlands, Switzerland, Tunisia | [197,208,218,230,231,232,233,234,235] |
| Introduced, eradicated | Belgium, Denmark | [197,208] |
| Not present | Estonia, Finland, Ireland, Israel, Lebanon, Malta, Morocco, Palestine, Sweden | [197,208,236,237] |
| Unknown status | China*, Libya | [208,238,239] |
| Strain/Isolate* | First identification | Characterization | Distribution | References |
|---|---|---|---|---|
| PPV-M(Marcus) | Peach in northern Greece | Common strain in southern, eastern and central Europe. Spread rapidly by aphids, epidemic strain of PPV. Apricot, peach, plum. | Austria, Bosnia and Herzegovina, Bulgaria, Croatia, Czech Republic, France, Germany, Hungary, Iran, Italy, Jordan, Kosovo, Serbia, Slovakia, Slovenia, Spain, Romania, Turkey | [10,14,18,199,250,254,255,256,257,258,259,260,261,262,263,264,265,266,267,268,269,270,271] |
| PPV-D (Dideron) | Apricot in Southern France | The most common in western Europe, epidemic strain of PPV. Apricot, peach (only poorly), plum. | Bosnia and Herzegovina, Bulgaria, Canada, Chile, Croatia, Czech Republic, France, Hungary, Japan, Kosovo, Lithuania, Romania, Slovakia, Spain, Turkey, Ukraine | [18,19,196,218,254,258,265,268,271,272,273,274,275,276,277,278,279] |
| PPV-rec(recombinant) | Recombination between PPV-M and PPV-D | Widespread in several central and eastern Europe countries. Efficiently transmitted by aphids. Plums. | Bosnia and Herzegovina, Bulgaria, Croatia, Czech republic, Germany, Hungary, Italy, Kosovo, Serbia, Slovakia, Slovenia | [14,265,267,271,277,278,279,280,281] |
| PPV-EA(El Amar) | Apricot from Egypt | Only Egypt. Limited data about distribution. Apricot, peach, plum trees. | Egypt | [282,283,284,285,286] |
| PPV-C**(Cherry) | Cherry from Moldova | Eastern and central European countries, Italy (probably eradicated). Transmitted efficiently by aphids.Sweet (PPV-SwC) and sour cherry (PVP-SoC). | Hungary, Italy, Moldova, Romania | [287,288,289,290] |
| PPV-T* (Turkey) | Plum from Ancara region, Turkey | Turkey, data still limited. | Turkey | [250] |
| PPV-Penn* (Pennsylvania) | Three different growers. Peach, nectarine, Plum. | USA, firstly in Adams County, Pensylvania in 1999. | USA | [291,292] |
| PPV-W(Winona) | Plum from Canada | Canada, found in Latvia (The LV-141pl and LV-145bt isolates appear to be representatives of the "ancestral" PPV-W strain) | Canada, Latvia | [253,293] |
3. Plum pox virus genome organization
4. Plum pox virus transmission and cytological, histological and biochemical changes in infected plants
5. Detection of PPV
| Plant host species | Symptoms | References |
|---|---|---|
| Chenopodium foetidum Lam.(Amaranthaceae) | yellow spots, some with necrotic centres or necrotic spots; not systemic | [327,346,347,348] |
| Nicandra physalodes (L.) P. Gaertn. (Solanaceae) | black-brown necrotic local lesions | [344,345] |
| Prunus domestica L. cv. Italian Pruneandcv. Pozegaca (Rosaceae) | chlorotic rings, bands and spots | [242,349,350] |
| Prunus japonica Thunb. (Rosaceae) | diffused chlorotic spots | [18,351] |
| Prunus maritima Marsh.(Rosaceae) | chlorotic spots, vein necrosis | [351] |
| Prunus persica L. (Batsch)cv. GF305 | vein necrosis, malformation of leaves | [352,353] |
| Prunus sibirica L. (Rosaceae) | green spots and faint line pattern | [351,354] |
| Prunus tomentosa Thunb. (Rosaceae) | epinasty and malformation of young leaves, chlorotic spots, necrotic spots of older leaves | [23,355,356] |
| Sorbus domestica L. (Rosaceae) | yellow spots, leaf chlorosis | [208,357] |
Acknowledgments
Conflict of Interest
References and Notes
- Albrechtova, L. Investigations on the distribution of sharka virus (plum pox virus) in tissue of prunus-domestica. J. Plant Dis. Prot. 1986, 93, 190–201. [Google Scholar]
- Neumuller, M.; Hartmann, W. The hypersensitivity of european plum (prunus domestica l.) against the plum pox virus. In Proceedings of the twentieth international symposium on virus and virus–like diseases of temperate fruit crops – fruit tree diseases, 2008; pp. 273–279.
- Badenes, M.L.; Asins, M.J.; Carbonell, E.A.; Glacer, G. Genetic diversity in apricot, prunus armeniaca, aimed at improving resistance to plum pox virus. Plant Breed. 1996, 115, 133–139. [Google Scholar]
- Balan, V.; Ivascu, A.; Toma, S. Susceptibility of apricot, nectarine and peach cultivars and hybrids to plum-pox virus. In Xvith international symposium on fruit tree virus diseases; 1995; pp. 299–305. [Google Scholar]
- Martinez-Gomez, P.; Rubio, M.; Dicenta, F.; Gradziel, T.M. Utilization of almond as source of plum pox virus resistance in peach breeding. In Proceedings of the xixth international symposium on virus and virus-like diseases of temperate fruit crops: Fruit tree diseases, 2004; pp. 289–293.
- Pribek, D.; Gaborjanyi, R.; Palkovics, L. Molecular characterization of plum pox virus almond isolate. In Proceedings of the xixth international symposium on virus and virus–like diseases of temperate fruit crops: Fruit tree diseases, 2004; pp. 289–293.
- Rubio, M.; Martinez-Gomez, P.; Dicenta, F. Resistance of almond cultivars to plum pox virus (sharka). Plant Breed. 2003, 122, 462–464. [Google Scholar] [CrossRef]
- Crescenzi, A.; Nuzzaci, M.; Levy, L.; Piazzolla, P.; Hadidi, A. Plum pox virus (ppv) in sweet cherry. In Xvith international symposium on fruit tree virus diseases; 1995; pp. 219–225. [Google Scholar]
- Fanigliulo, A.; Comes, S.; Crescenzi, A. Evaluation of cherry cultivars for their response to infection by plum pox virus sweet cherry strain. In Proceedings of the xixth international symposium on virus and virus–like diseases of temperate fruit crops: Fruit tree diseases, 2004; pp. 309–316.
- Boeglin, M.; Quiot, J.B.; Labonne, G. Risk assessment of contamination of cherry trees by plum pox virus in france. In Proceedings of the xixth international symposium on virus and virus–like diseases of temperate fruit crops: Fruit tree diseases, 2004; pp. 221–224.
- Damsteegt, V.D.; Scorza, R.; Stone, A.L.; Schneider, W.L.; Webb, K.; Demuth, M.; Gildow, F.E. Prunus host range of plum pox virus (ppv) in the united states by aphid and graft inoculation. Plant Dis. 2007, 91, 18–23. [Google Scholar] [CrossRef]
- Nemchinov, L.; Hadidi, A. Characterization of the sour cherry strain of plum pox virus. Phytopathology 1996, 86, 575–580. [Google Scholar] [CrossRef]
- Elibuyuk, I.O. Detection of plum pox virus in ornamental prunus cerasifera. Phytoparasitica 2006, 34, 347–352. [Google Scholar] [CrossRef]
- Kamenova, I. Prunus cerasifera as a host of plum pox virus in bulgaria. J. Plant Pathol. 2008, 90, 15–18. [Google Scholar]
- Rubio, M.; Garcia-Ibarra, A.; Dicenta, F.; Martinez-Gomez, P. Plum pox virus (sharka) sensitivity in prunus salicina and prunus cerasifera cultivars against a dideron-type isolate. Plant Breed. 2011, 130, 283–286. [Google Scholar]
- Kalinina, A.; Brown, D.C.W.; Ravelonandro, M. Susceptibility of ornamental prunus to plum pox potyvirus infection. In Proceedings of the international symposium on biotechnology of temperate fruit crops and tropical species, 2007; pp. 601–605.
- Kegler, H.; Fuchs, E.; Gruntzig, M.; Krczal, G.; Wegener, B. Susceptibility of genotypes of the genus prunus to plum pox potyvirus. J. Plant Dis. Prot. 1996, 103, 255–261. [Google Scholar]
- Sebestyen, D.; Nemeth, M.; Hangyal, R.; Krizbai, L.; Ember, I.; Nyerges, K.; Kolber, M.; Kiss, E.; Bese, G. Ornamental prunus species as new natural hosts of plum pox virus and their importance in the spread of the virus in hungary. J. Plant Pathol. 2008, 90, 57–61. [Google Scholar]
- Stobbs, L.W.; Van Driel, L.; Whybourne, K.; Carlson, C.; Tulloch, M.; Van Lier, J. Distribution of plum pox virus in residential sites, commercial nurseries, and native plant species in the niagara region, ontario, canada. Plant Dis. 2005, 89, 822–827. [Google Scholar] [CrossRef]
- Ilbagi, H.; Citir, A.; Bostan, H. Prunus spinosa l. - a natural wild host of some important fruit viruses in tekirdag,turkey. In Proceedings of the twentieth international symposium on virus and virus–like diseases of temperate fruit crops – fruit tree diseases, 2008; pp. 33–36.
- Polak, J. The role of prunus spinosa l in epidemiology of plum pox virus in the czech republic. In Diagnosis and identification of plant pathogens; 1997; pp. 527–530. [Google Scholar]
- Polak, J.; Oukropec, I. The determination of sources of resistance to plum pox virus suitable for peach. In Proceedings of the twentieth international symposium on virus and virus–like diseases of temperate fruit crops – fruit tree diseases, 2008; pp. 269–272.
- Cambra, M.; Capote, N.; Olmos, A.; Bertolini, E.; Gorris, M.T.; Africander, N.L.; Levy, L.; Lenardon, S.L.; Clover, G.; Wright, D. Proposal for a new international protocol for detection and identification of plum pox virus: Validation of the techniques. In Proceedings of the twentieth international symposium on virus and virus–like diseases of temperate fruit crops – fruit tree diseases, 2008; pp. 181–191.
- Damsteegt, V.D.; Scorza, R.; Gildow, F.E.; Schneider, W.L.; Stone, A.L.; Luster, D.G. Comparative host range of us isolates of plum pox virus among prunus and other woody plant species following graft inoculation or aphid transmission. Phytopathology 2004, 94, S24–S24. [Google Scholar]
- Monsion, M.; Briard, P.; Glasa, M.; Ravelonandro, M. Comparative techniques to perform koch's postulates with plum pox virus. In Proceedings of the twentieth international symposium on virus and virus–like diseases of temperate fruit crops – fruit tree diseases, 2008; pp. 221–225.
- Ravelonandro, M.; Briard, P.; Monsion, M. Strategy to identify and to characterize new isolates of plum pox virus. In Proceedings of the xixth international symposium on virus and virus–like diseases of temperate fruit crops: Fruit tree diseases, 2004; pp. 171–175.
- Salvador, B.; Delgadillo, M.O.; Saenz, P.; Garcia, J.A.; Simon-Mateo, C. Identification of plum pox virus pathogenicity determinants in herbaceous and woody hosts. Mol. Plant-Microbe Interact. 2008, 21, 20–29. [Google Scholar] [CrossRef]
- Baumgartnerova, H. First findings of plum pox virus in walnut trees (juglans regia l). Acta Virol. 1996, 40, 59–60. [Google Scholar]
- Polak, J. Hosts and symptoms of plum pox virus: Woody species other than fruit and ornamental species of prunus. EPPO Bull. 2006, 36, 225–226. [Google Scholar] [CrossRef]
- Gibbs, A.; Ohshima, K. Potyviruses and the digital revolution. In Annu. Rev. Phytopathol. ; 2010; Volume 48, pp. 205–223. [Google Scholar]
- Gibbs, A.J.; Mackenzie, A.M.; Wei, K.J.; Gibbs, M.J. The potyviruses of australia. Arch. Virol. 2008, 153, 1411–1420. [Google Scholar] [CrossRef]
- Revers, F.; Le Gall, O.; Candresse, T.; Maule, A.J. New advances in understanding the molecular biology of plant/potyvirus interactions. Mol. Plant-Microbe Interact. 12, 367–376.
- Riechmann, J.L.; Lain, S.; Garcia, J.A. Highlights and prospects of potyvirus molecular-biology. Journal of General Virology 1992, 73, 1–16. [Google Scholar] [CrossRef]
- Ward, C.W.; Shukla, D.D. Taxonomy of potyviruses - current problems and some solutions. Intervirology 1991, 32, 269–296. [Google Scholar]
- Fereres, A.; Kampmeier, G.E.; Irwin, M.E. Aphid attraction and preference for soybean and pepper plants infected with potyviridae. Ann. Entomol. Soc. Am. 1999, 92, 542–548. [Google Scholar]
- Manachini, B.; Casati, P.; Cinanni, L.; Bianco, P. Role of myzus persicae (hemiptera : Aphididae) and its secondary hosts in plum pox virus propagation. J. Econ. Entomol. 2007, 100, 1047–1052. [Google Scholar] [CrossRef]
- Srinivasan, R.; Alvarez, J.M. Effect of mixed viral infections (potato virus y-potato leafroll virus) on biology and preference of vectors myzus persicae and macrosiphum euphorbiae (hemiptera : Aphididae). J. Econ. Entomol. 2007, 100, 646–655. [Google Scholar] [CrossRef]
- Symmes, E.J.; Perring, T.M. Intraspecific variation in zucchini yellow mosaic virus transmission by myzus persicae and the impact of aphid host plant. J. Econ. Entomol. 2007, 100, 1764–1772. [Google Scholar] [CrossRef]
- Choi, I.R.; French, R.; Hein, G.L.; Stenger, D.C. Fully biologically active in vitro transcripts of the eriophyid mite-transmitted wheat streak mosaic tritimovirus. Phytopathology 1999, 89, 1182–1185. [Google Scholar] [CrossRef]
- Stephan, D.; Moeller, I.; Skoracka, A.; Ehrig, F.; Maiss, E. Eriophyid mite transmission and host range of a brome streak mosaic virus isolate derived from a full-length cdna clone. Arch. Virol. 2008, 153, 181–185. [Google Scholar] [CrossRef]
- Sanchez-Sanchez, H.; Henry, M.; Cardenas-Soriano, E.; Alvizo-Villasana, H.F. Identification of wheat streak mosaic virus and its vector aceria tosichella in mexico. Plant Dis. 2001, 85, 13–17. [Google Scholar] [CrossRef]
- Adkins, S.; Webb, S.E.; Achor, D.; Roberts, P.D.; Baker, C.A. Identification and characterization of a novel whitefly-transmitted member of the family potyviridae isolated from cucurbits in florida. Phytopathology 2007, 97, 145–154. [Google Scholar] [CrossRef]
- Adkins, S.; Webster, C.G.; Kousik, C.S.; Webb, S.E.; Roberts, P.D.; Stansly, P.A.; Turechek, W.W. Ecology and management of whitefly-transmitted viruses of vegetable crops in florida. Virus Res. 2011, 159, 110–114. [Google Scholar] [CrossRef]
- Valverde, R.A.; Sim, J.; Lotrakul, P. Whitefly transmission of sweet potato viruses. Virus Res. 2004, 100, 123–128. [Google Scholar] [CrossRef]
- Dessens, J.T.; Meyer, M. Identification of structural similarities between putative transmission proteins of polymyxa and spongospora transmitted bymoviruses and furoviruses. Virus Genes 1996, 12, 95–99. [Google Scholar] [CrossRef]
- Chen, J.P. Progress and prospects of studies on polymyxa graminis and its transmitted cereal viruses in china. Progress Nat. Sci. 2005, 15, 481–490. [Google Scholar] [CrossRef]
- McGrann, G.R.D.; Adams, M.J. Investigating resistance to barley mild mosaic virus. Plant Pathol. 2004, 53, 161–169. [Google Scholar] [CrossRef]
- Thompson, J.P.; Clewett, T.G.; Jennings, R.E.; Sheedy, J.G.; Owen, K.J.; Persley, D.M. Detection of polymyxa graminis in a barley crop in australia. Austral. Plant Pathol. 2011, 40, 66–75. [Google Scholar] [CrossRef]
- Campbell, R.N. Fungal transmission of plant viruses. Annu. Rev. Phytopathol. 1996, 34, 87–108. [Google Scholar] [CrossRef]
- Kuroda, T.; Nabata, K.; Hori, T.; Ishikawa, K.; Natsuaki, T. Soybean leaf rugose mosaic virus, a new soilborne virus in the family potyviridae, isolated from soybean in japan. Journal of General Plant Pathology 2010, 76, 382–388. [Google Scholar] [CrossRef]
- Chung, B.Y.W.; Miller, W.A.; Atkins, J.F.; Firth, A.E. An overlapping essential gene in the potyviridae. Proc. Natl. Acad. Sci. U. S. A. 2008, 105, 5897–5902. [Google Scholar]
- Wen, R.H.; Hajimorad, M.R. Mutational analysis of the putative pipo of soybean mosaic virus suggests disruption of pipo protein impedes movement. Virology 2010, 400, 1–7. [Google Scholar] [CrossRef]
- Wen, R.H.; Maroof, M.A.S.; Hajimorad, M.R. Amino acid changes in p3, and not the overlapping pipo-encoded protein, determine virulence of soybean mosaic virus on functionally immune rsv1-genotype soybean. Molecular Plant Pathology 2011, 12, 799–807. [Google Scholar] [CrossRef]
- Kneller, E.L.P.; Rakotondrafara, A.M.; Miller, W.A. Cap-independent translation of plant viral rnas. Virus Research 2006, 119, 63–75. [Google Scholar] [CrossRef]
- Vijayapalani, P.; Maeshima, M.; Nagasaki-Takekuchi, N.; Miller, W.A. Interaction of the trans-frame potyvirus protein p3n-pipo with host protein pcap1 facilitates potyvirus movement. PLoS Pathog. 2012, 8, 1–15. [Google Scholar]
- Adams, M.J.; Antoniw, J.F.; Beaudoin, F. Overview and analysis of the polyprotein cleavage sites in the family potyviridae. Mol. Plant Pathol. 2005, 6, 471–487. [Google Scholar] [CrossRef]
- Verchot, J.; Koonin, E.V.; Carrington, J.C. The 35-kda protein from the n-terminus of the potyviral polyprotein functions as a 3rd virus-encoded proteinase. Virology 1991, 185, 527–535. [Google Scholar] [CrossRef]
- Yoshida, N.; Shimura, H.; Yamashita, K.; Suzuki, M.; Masuta, C. Variability in the p1 gene helps to refine phylogenetic relationships among leek yellow stripe virus isolates from garlic. Arch. Virol. 2012, 157, 147–153. [Google Scholar] [CrossRef]
- Salvador, B.; Saenz, P.; Yanguez, E.; Quiot, J.B.; Quiot, L.; Delgadillo, M.O.; Garcia, J.A.; Simon-Mateo, C. Host-specific effect of p1 exchange between two potyviruses. Mol. Plant Pathol. 2008, 9, 147–155. [Google Scholar] [CrossRef]
- Shi, Y.; Chen, J.; Hong, X.; Adams, M.J. A potyvirus p1 protein interacts with the rieske fe/s protein of its host. Mol. Plant Pathol. 2007, 8, 785–790. [Google Scholar] [CrossRef]
- Valli, A.; Lopez-Moya, J.J.; Garcia, J.A. Recombination and gene duplication in the evolutionary diversification of p1 proteins in the family potyviridae. J. Gen. Virol. 2007, 88, 1016–1028. [Google Scholar] [CrossRef]
- Young, B.A.; Stenger, D.C.; Qu, F.; Morris, T.J.; Tatineni, S.; French, R. Tritimovirus p1 functions as a suppressor of rna silencing and an enhancer of disease symptoms. Virus Res. 2012, 163, 672–677. [Google Scholar] [CrossRef]
- Rohozkova, J.; Navratil, M. P1 peptidase - a mysterious protein of family potyviridae. J. Biosci. 2011, 36, 189–200. [Google Scholar] [CrossRef]
- Stenger, D.C.; Hein, G.L.; Gildow, F.E.; Horken, K.M.; French, R. Plant virus hc-pro is a determinant of eriophyid mite transmission. J. Virol. 2005, 79, 9054–9061. [Google Scholar] [CrossRef]
- Endres, M.; Mlotshwa, S.; Schauer, S.; Bowman, L.; Vance, V. Hc-pro suppression of rna silencing: Towards a mechanism. Phytopathology 2005, 95, S123–S123. [Google Scholar]
- Torres-Barcelo, C.; Daros, J.A.; Elena, S.F. Compensatory molecular evolution of hc-pro, an rna-silencing suppressor from a plant rna virus. Mol. Biol. Evol. 2010, 27, 543–551. [Google Scholar] [CrossRef]
- Shen, W.T.; Yan, P.; Gao, L.; Pan, X.Y.; Wu, J.Y.; Zhou, P. Helper component-proteinase (hc-pro) protein of papaya ringspot virus interacts with papaya calreticulin. Mol. Plant Pathol. 2010, 11, 335–346. [Google Scholar] [CrossRef]
- Guo, B.H.; Lin, J.Z.; Ye, K.Q. Structure of the autocatalytic cysteine protease domain of potyvirus helper-component proteinase. J. Biol. Chem. 2011, 286, 21937–21943. [Google Scholar] [CrossRef]
- Matthews, D.A.; Smith, W.W.; Ferre, R.A.; Condon, B.; Budahazi, G.; Sisson, W.; Villafranca, J.E.; Janson, C.A.; McElroy, H.E.; Gribskov, C.L.; et al. Structure of human rhinovirus 3c protease reveals a trypsin-like polypeptide fold, rna-binding site, and means for cleaving precursor polyprotein. Cell 1994, 77, 761–771. [Google Scholar] [CrossRef]
- Riechmann, J.L.; Cervera, M.T.; Garcia, J.A. Processing of the plum pox virus polyprotein at the p3-6k(1) junction is not required for virus viability. J. Gen. Virol. 1995, 76, 951–956. [Google Scholar] [CrossRef]
- Anindya, R.; Savithri, H.S. Potyviral nia proteinase, a proteinase with novel deoxyribonuclease activity. J. Biol. Chem. 2004, 279, 32159–32169. [Google Scholar] [CrossRef]
- Han, H.E.; Sellamuthu, S.; Shin, B.H.; Lee, Y.J.; Song, S.; Seo, J.S.; Baek, I.S.; Bae, J.; Kim, H.; Yoo, Y.J.; et al. The nuclear inclusion a (nia) protease of turnip mosaic virus (tumv) cleaves amyloid-beta. Plos One 2010, 5. [Google Scholar]
- Han, J.S.; Kang, H.J.; Song, B.D.; Choi, K.Y. Novel mechanism of activity control of potyvirus nia protease. Faseb J. 2002, 16, A905–A905. [Google Scholar]
- Puhl, A.C.; Giacomini, C.; Irazoqui, G.; Batista-Viera, F.; Villarino, A.; Terenzi, H. Covalent immobilization of tobacco-etch-virus nia protease: A useful tool for cleavage: Of the histidine tag of recombinant proteins. Biotechnol. Appl. Biochem. 2009, 53, 165–174. [Google Scholar] [CrossRef]
- Chen, K.C.; Chiang, C.H.; Raja, J.A.J.; Liu, F.L.; Tai, C.H.; Yeh, S.D. A single amino acid of niapro of papaya ringspot virus determines host specificity for infection of papaya. Mol. Plant-Microbe Interact. 2008, 21, 1046–1057. [Google Scholar] [CrossRef]
- Mathur, C.; Jimsheena, V.K.; Banerjee, S.; Makinen, K.; Gowda, L.R.; Savithri, H.S. Functional regulation of pvbv nuclear inclusion protein-a protease activity upon interaction with viral protein genome-linked and phosphorylation. Virology 2012, 422, 254–264. [Google Scholar] [CrossRef]
- Restrepo, M.A.; Freed, D.D.; Carrington, J.C. Nuclear transport of plant potyviral proteins. Plant Cell 1990, 2, 987–998. [Google Scholar] [Green Version]
- Knuhtsen, H.; Hiebert, E.; Purciful, D. Partial-purification and some properties of tobacco etch virus-induced intranuclear inclusions. Virology 1974, 61, 200–209. [Google Scholar] [CrossRef]
- Shepard, J.F. Electron microscopy of subtilisin-treated tobacco etch virus nuclear and cytoplasmic inclusions. Virology 1968, 36, 20. [Google Scholar] [CrossRef]
- Sheffield, F.M.L. The cytoplasmic and nuclear inclusions associated with severe etch virus. J. Royal Microscop. Soc. 1941, 61, 30–45. [Google Scholar] [CrossRef]
- Fellers, J.P.; Collins, G.B.; Hunt, A.G. The nia-proteinase of different plant potyviruses provides specific resistance to viral infection. Crop Sci. 1998, 38, 1309–1319. [Google Scholar] [CrossRef]
- Fukuzawa, N.; Itchoda, N.; Ishihara, T.; Goto, K.; Masuta, C.; Matsumura, T. Hc-pro, a potyvirus rna silencing suppressor, cancels cycling of cucumber mosaic virus in nicotiana benthamiana plants. Virus Genes 2010, 40, 440–446. [Google Scholar] [CrossRef]
- Giner, A.; Lopez-Moya, J.J.; Lakatos, L. Rna silencing in plants and the role of viral suppressors. In Rna interference and viruses: Current innovations and future trends; CAISTER ACADEMIC PRESS: WYMONDHAM, 2010; pp. 25–46. [Google Scholar]
- Reyes, C.A.; De Francesco, A.; Pena, E.J.; Costa, N.; Plata, M.I.; Sendin, L.; Castagnaro, A.P.; Garcia, M.L. Resistance to citrus psorosis virus in transgenic sweet orange plants is triggered by coat protein-rna silencing. J. Biotechnol. 2011, 151, 151–158. [Google Scholar]
- Jenner, C.E.; Wang, X.W.; Tomimura, K.; Ohshima, K.; Ponz, F.; Walsh, J.A. The dual role of the potyvirus p3 protein of turnip mosaic virus as a symptom and avirulence determinant in brassicas. Mol. Plant-Microbe Interact. 2003, 16, 777–784. [Google Scholar] [CrossRef]
- Lin, L.; Luo, Z.P.; Yan, F.; Lu, Y.W.; Zheng, H.Y.; Chen, J.P. Interaction between potyvirus p3 and ribulose-1,5-bisphosphate carboxylase/oxygenase (rubisco) of host plants. Virus Genes 2011, 43, 90–92. [Google Scholar] [CrossRef]
- Chowda-Reddy, R.V.; Sun, H.Y.; Chen, H.Y.; Poysa, V.; Ling, H.; Gijzen, M.; Wang, A.M. Mutations in the p3 protein of soybean mosaic virus g2 isolates determine virulence on rsv4-genotype soybean. Mol. Plant-Microbe Interact. 2011, 24, 37–43. [Google Scholar] [CrossRef]
- Ryslava, H.; Doubnerova, V.; Janoskova, M.; Subr, Z.; Novakova, S.; Cerovska, N. Influence of viral infection on transgenic plants containing genes for nonstructural potyviral proteins hc-pro and p3. Febs J. 2005, 272, 464–464. [Google Scholar] [CrossRef]
- Saenz, P.; Cervera, M.T.; Dallot, S.; Quiot, L.; Quiot, J.B.; Riechmann, J.L.; Garcia, J.A. Identification of a pathogenicity determinant of plum pox virus in the sequence encoding the c-terminal region of protein p3+6k(1). J. Gen. Virol. 2000, 81, 557–566. [Google Scholar]
- Suehiro, N.; Natsuaki, T.; Watanabe, T.; Okuda, S. An important determinant of the ability of turnip mosaic virus to infect brassica spp. And/or raphanus sativus is in its p3 protein. J. Gen. Virol. 2004, 85, 2087–2098. [Google Scholar] [CrossRef]
- Wen, R.H.; Maroof, M.A.S.; Hajimorad, M.R. Amino acid changes in p3, and not the overlapping pipo-encoded protein, determine virulence of soybean mosaic virus on functionally immune rsv1-genotype soybean. Mol. Plant Pathol. 2011, 12, 799–807. [Google Scholar] [CrossRef]
- Cui, X.Y.; Wei, T.Y.; Chowda-Reddy, R.V.; Sun, G.Y.; Wang, A.M. The tobacco etch virus p3 protein forms mobile inclusions via the early secretory pathway and traffics along actin microfilaments. Virology 2010, 397, 56–63. [Google Scholar] [CrossRef]
- Eiamtanasate, S.; Juricek, M.; Yap, Y.K. C-terminal hydrophobic region leads prsv p3 protein to endoplasmic reticulum. Virus Genes 2007, 35, 611–617. [Google Scholar] [CrossRef]
- Hong, X.Y.; Chen, J.; Shi, Y.H.; Chen, J.P. The '6k1' protein of a strain of soybean mosaic virus localizes to the cell periphery. Arch. Virol. 2007, 152, 1547–1551. [Google Scholar] [CrossRef]
- Carrington, J.C.; Jensen, P.E.; Schaad, M.C. Genetic evidence for an essential role for potyvirus ci protein in cell-to-cell movement. Plant J. 1998, 14, 393–400. [Google Scholar] [CrossRef]
- RodriguezCerezo, E.; Findlay, K.; Shaw, J.G.; Lomonossoff, G.P.; Qiu, S.G.; Linstead, P.; Shanks, M.; Risco, C. The coat and cylindrical inclusion proteins of a potyvirus are associated with connections between plant cells. Virology 1997, 236, 296–306. [Google Scholar] [CrossRef]
- Roberts, I.M.; Wang, D.; Findlay, K.; Maule, A.J. Ultrastructural and temporal observations of the potyvirus cylindrical inclusions (cis) show that the ci protein acts transiently in aiding virus movement. Virology 1998, 245, 173–181. [Google Scholar] [CrossRef]
- Jimenez, I.; Lopez, L.; Alamillo, J.M.; Valli, A.; Garcia, J.A. Identification of a plum pox virus ci-interacting protein from chloroplast that has a negative effect in virus infection. Mol. Plant-Microbe Interact. 2006, 19, 350–358. [Google Scholar] [CrossRef]
- Abdul-Razzak, A.; Guiraud, T.; Peypelut, M.; Walter, J.; Houvenaghel, M.C.; Candresse, T.; Le Gall, O.; German-Retana, S. Involvement of the cylindrical inclusion (ci) protein in the overcoming of an eif4e-mediated resistance against lettuce mosaic potyvirus. Mol. Plant Pathol. 2009, 10, 109–113. [Google Scholar] [CrossRef]
- Tavert-Roudet, G.; Abdul-Razzak, A.; Doublet, B.; Walter, J.; Delaunay, T.; German-Retana, S.; Michon, T.; Le Gall, O.; Candresse, T. The c terminus of lettuce mosaic potyvirus cylindrical inclusion helicase interacts with the viral vpg and with lettuce translation eukaryotic initiation factor 4e. J. Gen. Virol. 2012, 93, 184–193. [Google Scholar] [CrossRef]
- Ayme, V.; Souche, S.; Caranta, C.; Jacquemond, M.; Chadoeuf, J.; Palloix, A.; Moury, B. Different mutations in the genome-linked protein vpg of potato virus y confer virulence on the pvr2(3) resistance in pepper. Mol. Plant-Microbe Interact. 2006, 19, 557–563. [Google Scholar] [CrossRef]
- Charron, C.; Nicolai, M.; Gallois, J.L.; Robaglia, C.; Moury, B.T.; Palloix, A.; Caranta, C. Natural variation and functional analyses provide evidence for co-evolution between plant eif4e and potyviral vpg. Plant J. 2008, 54, 56–68. [Google Scholar] [CrossRef]
- Rajamaki, M.L.; Valkonen, J.P.T. The 6k2 protein and the vpg of potato virus a are determinants of systemic infection in nicandra physaloides. Mol. Plant-Microbe Interact. 1999, 12, 1074–1081. [Google Scholar] [CrossRef]
- Valkonen, J.P.T.; Rajamaki, M.L.; Hamalainen, J.; Kekarainen, T.; Oruetxebarria, I. Incompatible potyvirus-host interactions in resistant plants: The viral genome-linked protein (vpg) of potato virus a as an avirulence determinant. In Proceedings of the 7th aschersleben symposium new aspects of resistance research on culitivated plants: Virus diseases; BUNDESANSTALT FUR ZUCHTUNGS FORSCHUNG KULTURPFLANZEN: QUEDLINBURG, 2000; pp. 92–95. [Google Scholar]
- Puurand, U.; Makinen, K.; Paulin, L.; Saarma, M. The nucleotide-sequence of potato-virus-a genomic rna and its sequence similarities with other potyviruses. J. Gen. Virol. 1994, 75, 457–461. [Google Scholar] [CrossRef]
- Rantalainen, K.I.; Eskelin, K.; Tompa, P.; Makinen, K. Structural flexibility allows the functional diversity of potyvirus genome-linked protein vpg. J. Virol. 2011, 85, 2449–2457. [Google Scholar] [CrossRef]
- Rajamaki, M.L.; Valkonen, J.P.T. Localization of a potyvirus and the viral genome-linked protein in wild potato leaves at an early stage of systemic infection. Mol. Plant-Microbe Interact. 2003, 16, 25–34. [Google Scholar] [CrossRef]
- Vuorinen, A.L.; Kelloniemi, J.; Valkonen, J.P.T. Why do viruses need phloem for systemic invasion of plants? Plant Sci. 2011, 181, 355–363. [Google Scholar] [CrossRef]
- Rajamaki, M.L.; Valkonen, J.P.T. Viral genome-linked protein (vpg) controls accumulation and phloem-loading of a potyvirus in inoculated potato leaves. Mol. Plant-Microbe Interact. 2002, 15, 138–149. [Google Scholar] [CrossRef]
- Puustinen, P.; Rajamaki, M.L.; Ivanov, K.I.; Valkonen, J.P.T.; Makinen, K. Detection of the potyviral genome-linked protein vpg in virions and its phosphorylation by host kinases. J. Virol. 2002, 76, 12703–12711. [Google Scholar]
- Roudet-Tavert, G.; Michon, T.; Walter, J.; Delaunay, T.; Redondo, E.; Le Gall, O. Central domain of a potyvirus vpg is involved in the interaction with the host translation initiation factor eif4e and the viral protein hcpro. J. Gen. Virol. 2007, 88, 1029–1033. [Google Scholar] [CrossRef]
- Yambao, M.L.M.; Masuta, C.; Nakahara, K.; Uyeda, I. The central and c-terminal domains of vpg of clover yellow vein virus are important for vpg-hcpro and vpg-vpg interactions. J. Gen. Virol. 2003, 84, 2861–2869. [Google Scholar] [CrossRef]
- Dunoyer, P.; Thomas, C.; Harrison, S.; Revers, F.; Maule, A. A cysteine-rich plant protein potentiates potyvirus movement through an interaction with the virus genome-linked protein vpg. J. Virol. 2004, 78, 2301–2309. [Google Scholar] [CrossRef]
- Hong, Y.L.; Levay, K.; Murphy, J.F.; Klein, P.G.; Shaw, J.G.; Hunt, A.G. A potyvirus polymerase interacts with the viral coat protein and vpg in yeast-cells. Virology 1995, 214, 159–166. [Google Scholar] [CrossRef]
- Carrington, J.C.; Haldeman, R.; Dolja, V.V.; Restrepohartwig, M.A. Internal cleavage and trans-proteolytic activities of the vpg-proteinase (nia) of tobacco etch potyvirus in-vivo. J. Virol. 1993, 67, 6995–7000. [Google Scholar]
- Hajimorad, M.R.; Ding, X.S.; Flasinski, S.; Mahajan, S.; Graff, E.; HaldemanCahill, R.; Carrington, J.C.; Cassidy, B.G. Nla and nlb of peanut stripe potyvirus are present in the nucleus of infected cells, but do not form inclusions. Virology 1996, 224, 368–379. [Google Scholar] [CrossRef]
- Beauchemin, C.; Boutet, N.; Laliberte, J.F. Visualization of the interaction between the precursors of vpg, the viral protein linked to the genome of turnip mosaic virus, and the translation eukaryotic initiation factor iso 4e in planta. J. Virol. 2007, 81, 775–782. [Google Scholar] [CrossRef]
- Borgstrom, B.; Johansen, I.E. Mutations in pea seedborne mosaic virus genome-linked protein vpg alter pathotype-specific virulence in pisum sativum. Mol. Plant-Microbe Interact. 2001, 14, 707–714. [Google Scholar] [CrossRef]
- Daughenbaugh, K.F.; Fraser, C.S.; Hershey, J.W.B.; Hardy, M.E. The genome-linked protein vpg of the norwalk virus binds eif3, suggesting its role in translation initiation complex recruitment. Embo J. 2003, 22, 2852–2859. [Google Scholar] [CrossRef]
- Leonard, S.; Plante, D.; Wittmann, S.; Daigneault, N.; Fortin, M.G.; Laliberte, J.F. Complex formation between potyvirus vpg and translation eukaryotic initiation factor 4e correlates with virus infectivity. J. Virol. 2000, 74, 7730–7737. [Google Scholar] [CrossRef]
- Leonard, S.; Viel, C.; Beauchemin, C.; Daigneault, N.; Fortin, M.G.; Laliberte, J.F. Interaction of vpg-pro of turnip mosaic virus with the translation initiation factor 4e and the poly(a)-binding protein in planta. J. Gen. Virol. 2004, 85, 1055–1063. [Google Scholar] [CrossRef]
- Plante, D.; Viel, C.; Leonard, S.; Tampo, H.; Laliberte, J.F.; Fortin, M.G. Tumip mosaic virus vpg does not disrupt the translation initiation complex but interferes with cap binding. Physiol. Mol. Plant Pathol. 2004, 64, 219–226. [Google Scholar] [CrossRef]
- Grzela, R.; Strokovska, L.; Andrieu, J.P.; Dublet, B.; Zagorski, W.; Chroboczek, J. Potyvirus terminal protein vpg, effector of host eukaryotic initiation factor eif4e. Biochimie 2006, 88, 887–896. [Google Scholar] [CrossRef]
- Michon, T.; Estevez, Y.; Walter, J.; German-Retana, S.; Le Gall, O. The potyviral virus genome-linked protein vpg forms a ternary complex with the eukaryotic initiation factors eif4e and eif4g and reduces eif4e affinity for a mrna cap analogue. Febs J. 2006, 273, 1312–1322. [Google Scholar] [CrossRef]
- Eskelin, K.; Hafren, A.; Rantalainen, K.I.; Makinen, K. Potyviral vpg enhances viral rna translation and inhibits reporter mrna translation in planta. J. Virol. 2011, 85, 9210–9221. [Google Scholar] [CrossRef]
- Rantalainen, K.I.; Christensen, P.A.; Hafren, A.; Otzen, D.E.; Kalkkinen, N.; Makinen, K. Interaction of a potyviral vpg with anionic phospholipid vesicles. Virology 2009, 395, 114–120. [Google Scholar]
- Li, X.H.; Valdez, P.; Olvera, R.E.; Carrington, J.C. Functions of the tobacco etch virus rna polymerase (nib): Subcellular transport and protein-protein interaction with vpg/proteinase (nia). J. Virol. 1997, 71, 1598–1607. [Google Scholar]
- Daros, J.A.; Schaad, M.C.; Carrington, J.C. Functional analysis of the interaction between vpg-proteinase (nia) and rna polymerase (nib) of tobacco etch potyvirus, using conditional and suppressor mutants. J. Virol. 1999, 73, 8732–8740. [Google Scholar]
- Guo, D.Y.; Rajamaki, M.L.; Saarma, M.; Valkonen, J.P.T. Towards a protein interaction map of potyviruses: Protein interaction matrixes of two potyviruses based on the yeast two-hybrid system. J. Gen. Virol. 2001, 82, 935–939. [Google Scholar]
- Lin, L.; Shi, Y.H.; Luo, Z.P.; Lu, Y.W.; Zheng, H.Y.; Yan, F.; Chen, J.; Chen, J.P.; Adams, M.J.; Wu, Y.F. Protein-protein interactions in two potyviruses using the yeast two-hybrid system. Virus Res. 2009, 142, 36–40. [Google Scholar] [CrossRef]
- Puustinen, P.; Makinen, K. Uridylylation of the potyvirus vpg by viral replicase nib correlates with the nucleotide binding capacity of vpg. J. Biol. Chem. 2004, 279, 38103–38110. [Google Scholar] [CrossRef]
- Jones, A.L.; Johansen, I.E.; Bean, S.J.; Bach, I.; Maule, A.J. Specificity of resistance to pea seed-borne mosaic potyvirus in transgenic peas expressing the viral replicase (nib) gene. J. Gen. Virol. 1998, 79, 3129–3137. [Google Scholar]
- Sivamani, E.; Brey, C.W.; Dyer, W.E.; Talbert, L.E.; Qu, R.D. Resistance to wheat streak mosaic virus in transgenic wheat expressing the viral replicase (nib) gene. Mol. Breed. 2000, 6, 469–477. [Google Scholar] [CrossRef]
- Cassidy, B.G.; Nelson, R.S. Differences in protection phenotypes in tobacco plants expressing coat protein genes from peanut stripe potyvirus with or without an engineered atg. Mol. Plant-Microbe Interact. 1995, 8, 357–365. [Google Scholar] [CrossRef]
- Crescenzi, A.; d'Aquino, L.; Nuzzaci, M.; Ostuni, A.; Bavoso, A.; Comes, S.; De Stradis, A.; Piazzolla, P. Production of strain specific antibodies against a synthetic polypeptide corresponding to the n-terminal region of the plum pox potyvirus coat protein. J. Virol. Methods 1997, 69, 181–189. [Google Scholar] [CrossRef]
- Gotz, R.; Huth, W.; Maiss, E. Molecular analyses of the coat protein region of different viruses on poaceae belonging to the potyviridae. Agronomie 1995, 15, 491–494. [Google Scholar] [CrossRef]
- Handley, J.A.; Smith, G.R.; Dale, J.L.; Harding, R.M. Sequence diversity in the coat protein coding region of twelve sugarcane mosaic potyvirus isolates from australia, USA and south africa. Arch. Virol. 1998, 143, 1145–1153. [Google Scholar] [CrossRef]
- Kobayashi, K.; Rabinowicz, P.; BravoAlmonacid, F.; Helguera, M.; Conci, V.; Lot, H.; Mentaberry, A. Coat protein gene sequences of garlic and onion isolates of the onion yellow dwarf potyvirus (oydv). Arch. Virol. 1996, 141, 2277–2287. [Google Scholar] [CrossRef]
- Ryu, K.H.; Kim, S.J.; Park, W.M. Nucleotide sequence analysis of the coat protein genes of two korean isolates of sweet potato feathery mottle potyvirus. Arch. Virol. 1998, 143, 557–562. [Google Scholar] [CrossRef]
- Valkonen, J.P.T.; Puurand, U.; Slack, S.A.; Makinen, K.; Saarma, M. 3 strain groups of potato-a-potyvirus based on hypersensitive responses in potato, serological properties, and coat protein sequences. Plant Dis. 1995, 79, 748–753. [Google Scholar] [CrossRef]
- Massey, B.; Cui, X.; Hiebert, E.; Elliott, M.S.; Waipara, N.; Hayes, L.; Charudattan, R. Partial sequencing of the genomic rna of araujia mosaic virus and comparison of the coat protein sequence with those of other potyviruses. Arch. Virol. 2007, 152, 2125–2129. [Google Scholar] [CrossRef]
- Pfosser, M.F.; Baumann, H. Phylogeny and geographical differentiation of zucchini yellow mosaic virus isolates (potyviridae) based on molecular analysis of the coat protein and part of the cytoplasmic inclusion protein genes. Arch. Virol. 2002, 147, 1599–1609. [Google Scholar] [CrossRef]
- Sanchez, F.; Wang, X.; Jenner, C.E.; Walsh, J.A.; Ponz, F. Strains of turnip mosaic potyvirus as defined by the molecular analysis of the coat protein gene of the virus. Virus Res. 2003, 94, 33–43. [Google Scholar] [CrossRef]
- Andrejeva, J.; Puurand, U.; Merits, A.; Rabenstein, F.; Jarvekulg, L.; Valkonen, J.P.T. Potyvirus helper component-proteinase and coat protein (cp) have coordinated functions in virus-host interactions and the same cp motif affects virus transmission and accumulation. J. Gen. Virol. 1999, 80, 1133–1139. [Google Scholar]
- Atreya, P.L.; Lopezmoya, J.J.; Chu, M.H.; Atreya, C.D.; Pirone, T.P. Mutational analysis of the coat protein n-terminal amino-acids involved in potyvirus transmission by aphids. J. Gen. Virol. 1995, 76, 265–270. [Google Scholar]
- Blanc, S.; LopezMoya, J.J.; Wang, R.Y.; GarciaLampasona, S.; Thornbury, D.W.; Pirone, T.P. A specific interaction between coat protein and helper component correlates with aphid transmission of a potyvirus. Virology 1997, 231, 141–147. [Google Scholar] [CrossRef]
- Flasinski, S.; Cassidy, B.G. Potyvirus aphid transmission requires helper component and homologous coat protein for maximal efficiency. Arch. Virol. 1998, 143, 2159–2172. [Google Scholar] [CrossRef]
- Lopez-Moya, J.J.; Pirone, T.P. Charge changes near the n terminus of the coat protein of two potyviruses affect virus movement. J. Gen. Virol. 1998, 79, 161–165. [Google Scholar]
- Lopez-Moya, J.J.; Wang, R.Y.; Pirone, T.P. Context of the coat protein dag motif affects potyvirus transmissibility by aphids. J. Gen. Virol. 1999, 80, 3281–3288. [Google Scholar]
- Dombrovsky, A.; Huet, H.; Chejanovsky, N.; Raccah, B. Aphid transmission of a potyvirus depends on suitability of the helper component and the n terminus of the coat protein. Arch. Virol. 2005, 150, 287–298. [Google Scholar] [CrossRef]
- Andersen, K.; Johansen, I.E. A single conserved amino acid in the coat protein gene of pea seed-borne mosaic potyvirus modulates the ability of the virus to move systemically in chenopodium quinoa. Virology 1998, 241, 304–311. [Google Scholar] [CrossRef]
- Aparicio, F.; Thomas, C.L.; Lederer, C.; Niu, Y.; Wang, D.W.; Maule, A.J. Virus induction of heat shock protein 70 reflects a general response to protein accumulation in the plant cytosol. Plant Physiol. 2005, 138, 529–536. [Google Scholar] [CrossRef]
- Sugio, A.; Dreos, R.; Aparicio, F.; Maule, A.J. The cytosolic protein response as a subcomponent of the wider heat shock response in arabidopsis. Plant Cell 2009, 21, 642–654. [Google Scholar] [CrossRef]
- Hafren, A.; Hofius, D.; Ronnholm, G.; Sonnewald, U.; Makinen, K. Hsp70 and its cochaperone cpip promote potyvirus infection in nicotiana benthamiana by regulating viral coat protein functions. Plant Cell 2010, 22, 523–535. [Google Scholar] [CrossRef]
- Wang, Y.Y.; Gardner, R.C.; Pearson, M.N. Resistance to vanilla necrosis potyvirus in transgenic nicotiana benthamiana plants containing the virus coat protein gene. J. Phytopathol. 1997, 145, 7–15. [Google Scholar] [CrossRef]
- Dinant, S.; Maisonneuve, B.; Albouy, J.; Chupeau, Y.; Chupeau, M.C.; Bellec, Y.; Gaudefroy, F.; Kusiak, C.; Souche, S.; Robaglia, C.; et al. Coat protein gene-mediated protection in lactuca sativa against lettuce mosaic potyvirus strains. Mol. Breed. 1997, 3, 75–86. [Google Scholar] [CrossRef]
- Xu, D.; Liu, H.; Li, F.; Howell, B.; Tian, T.; Li, R. Biological characterization and complete genomic sequence of carrot thin leaf virus. Phytopathology 2011, 101, S195–S195. [Google Scholar]
- Fuchs, M.; Gal-On, A.; Raccah, B.; Gonsalves, D. Epidemiology of an aphid nontransmissible potyvirus in fields of nontransgenic and coat protein transgenic squash. Transgenic Res. 1999, 8, 429–439. [Google Scholar] [CrossRef]
- Higgins, C.M.; Hall, R.M.; Mitter, N.; Cruickshank, A.; Dietzgen, R.G. Peanut stripe potyvirus resistance in peanut (arachis hypogaea l.) plants carrying viral coat protein gene sequences. Transgenic Res. 2004, 13, 59–67. [Google Scholar] [CrossRef]
- Ravelonandro, M.; Dunez, J.; Scorza, R.; Labonne, G. Challenging transgenic plums expressing potyvirus coat protein genes with viruliferous aphids. In 17th international symposium on virus and virus-like diseases of temperate fruit crops: Fruit tree diseases, vols 1 and 2; INTERNATIONAL SOCIETY HORTICULTURAL SCIENCE: LEUVEN, 1998; pp. 413–420. [Google Scholar]
- Scorza, R.; Callahan, A.M.; Levy, L.; Damsteegt, V.; Ravelonandro, M. Transferring potyvirus coat protein genes through hybridization of transgenic plants to produce plum pox virus resistant plums (prunus domestica l.). In 17th international symposium on virus and virus-like diseases of temperate fruit crops: Fruit tree diseases, vols 1 and 2; INTERNATIONAL SOCIETY HORTICULTURAL SCIENCE: LEUVEN 1, 1998; pp. 421–427. [Google Scholar]
- Yadav, N.; Khan, J.A. Over expression of narcissus potyvirus coat protein in e. Coli. Indian J. Virol. 2009, 20, 44–44. [Google Scholar]
- Savenkov, E.I.; Valkonen, J.P.T. Coat protein gene-mediated resistance to potato virus a in transgenic plants is suppressed following infection with another potyvirus. J. Gen. Virol. 2001, 82, 2275–2278. [Google Scholar]
- Shand, K.; Theodoropoulos, C.; Stenzel, D.; Dale, J.L.; Harrison, M.D. Expression of potato virus y cytoplasmic inclusion protein in tobacco results in disorganization of parenchyma cells, distortion of epidermal cells, and induces mitochondrial and chloroplast abnormalities, formation of membrane whorls and atypical lipid accumulation. Micron 2009, 40, 730–736. [Google Scholar] [CrossRef]
- Restrepohartwig, M.A.; Carrington, J.C. Regulation of nuclear transport of a plant potyvirus protein by autoproteolysis. J. Virol. 1992, 66, 5662–5666. [Google Scholar]
- Restrepohartwig, M.A.; Carrington, J.C. The tobacco etch potyvirus 6-kilodalton protein is membrane-associated and involved in viral replication. J. Virol. 1994, 68, 2388–2397. [Google Scholar]
- Spetz, C.; Valkonen, J.P.T. Potyviral 6k2 protein long-distance movement and symptom-induction functions are independent and host-specific. Mol. Plant-Microbe Interact. 2004, 17, 502–510. [Google Scholar] [CrossRef]
- Yoshida, N.; Shimura, H.; Yamashita, K.; Suzuki, M.; Masuta, C. Variability in the p1 gene helps to refine phylogenetic relationships among leek yellow stripe virus isolates from garlic. Arch. Virol. 2011, 157, 147–153. [Google Scholar]
- Martin, M.T.; Gelie, B. Non-structural plum pox potyvirus proteins detected by immunogold labelling. Eur. J. Plant Pathol. 1997, 103, 427–431. [Google Scholar] [CrossRef]
- Arbatova, J.; Lehto, K.; Pehu, E.; Pehu, T. Localization of the p1 protein of potato y potyvirus in association with cytoplasmic inclusion bodies and in the cytoplasm of infected cells. J. Gen. Virol. 1998, 79, 2319–2323. [Google Scholar]
- Mangrauthia, S.K.; Jain, R.K.; Praveen, S. Sequence motifs comparisons establish a functional portrait of a multifunctional protein hc-pro from papaya ringspot potyvirus. J. Plant Biochem. Biotechnol. 2008, 17, 201–204. [Google Scholar]
- Plisson, C.; Drucker, M.; Blanc, S.; German-Retana, S.; Le Gall, O.; Thomas, D.; Bron, P. Structural characterization of hc-pro, a plant virus multifunctional protein. J. Biol. Chem. 2003, 278, 23753–23761. [Google Scholar]
- Soitamo, A.J.; Jada, B.; Lehto, K. Hc-pro silencing suppressor significantly alters the gene expression profile in tobacco leaves and flowers. BMC Plant Biol. 2011, 11. [Google Scholar]
- Wu, H.W.; Lin, S.S.; Chen, K.C.; Yeh, S.D.; Chua, N.H. Discriminating mutations of hc-pro of zucchini yellow mosaic virus with differential effects on small rna pathways involved in viral pathogenicity and symptom development. Mol. Plant-Microbe Interact. 2010, 23, 17–28. [Google Scholar] [CrossRef]
- Zheng, H.Y.; Yan, F.; Lu, Y.W.; Sun, L.Y.; Lin, L.; Cai, L.; Hou, M.S.; Chen, J.P. Mapping the self-interacting domains of tumv hc-pro and the subcellular localization of the protein. Virus Genes 2011, 42, 110–116. [Google Scholar] [CrossRef]
- Hjulsager, C.K.; Olsen, B.S.; Jensen, D.M.K.; Cordea, M.I.; Krath, B.N.; Johansen, I.E.; Lund, O.S. Multiple determinants in the coding region of pea seed-borne mosaic virus p3 are involved in virulence against sbm-2 resistance. Virology 2006, 355, 52–61. [Google Scholar] [CrossRef]
- Langenberg, W.G.; Zhang, L.Y. Immunocytology shows the presence of tobacco etch virus p3 protein in nuclear inclusions. J. Struct. Biol. 1997, 118, 243–247. [Google Scholar] [CrossRef]
- Hjulsager, C.K.; Lund, O.S.; Johansen, I.E. A new pathotype of pea seedborne mosaic virus explained by properties of the p3-6k1-and viral genome-linked protein (vpg)-coding regions. Mol. Plant-Microbe Interact. 2002, 15, 169–171. [Google Scholar] [CrossRef]
- Waltermann, A.; Maiss, E. Detection of 6k1 as a mature protein of 6 kda in plum pox virus-infected nicotiana benthamiana. J. Gen. Virol. 2006, 87, 2381–2386. [Google Scholar] [CrossRef]
- Wei, T.Y.; Zhang, C.W.; Hong, J.A.; Xiong, R.Y.; Kasschau, K.D.; Zhou, X.P.; Carrington, J.C.; Wang, A.M. Formation of complexes at plasmodesmata for potyvirus intercellular movement is mediated by the viral protein p3n-pipo. PLoS Pathog. 2010, 6. [Google Scholar]
- Riedel, D.; Lesemann, D.E.; Maiss, E. Ultrastructural localization of nonstructural and coat proteins of 19 potyviruses using antisera to bacterially expressed proteins of plum pox potyvirus. Arch. Virol. 1998, 143, 2133–2158. [Google Scholar] [CrossRef]
- Voloudakis, A.E.; Malpica, C.A.; Aleman-Verdaguer, M.E.; Stark, D.M.; Fauquet, C.M.; Beachy, R.N. Structural characterization of tobacco etch virus coat protein mutants. Arch. Virol. 2004, 149, 699–712. [Google Scholar] [CrossRef]
- Hema, M.; Reddy, C.V.S.; Savithri, H.S.; Sreenivasulu, P. Assembly of recombinant coat protein of sugarcane streak mosaic virus into potyvirus-like particles. Indian J. Exp. Biol. 2008, 46, 793–796. [Google Scholar]
- Atanasoff, D. Plum pox. A new virus disease. In Yearbook university of sofia; University of Sofia, 1932; Volume 11, pp. 49–69. [Google Scholar]
- Atanasoff, D. Mosaic of stone fruits. Phytopathology 1935, 8. [Google Scholar]
- Candresse, T.; Svanella-Dumas, L.; Gentit, P.; Caglayan, K.; Cevik, B. First report of the presence of plum pox virus rec strain in turkey. Plant Dis. 2007, 91, 331–331. [Google Scholar]
- Myrta, A.; Rwahnih, M.A.; Savino, V. Presence of a recombinant isolate of plum pox virus in apulia. J. Plant Pathol. 2005, 87, 127–130. [Google Scholar]
- Snover-Clift, K.L.; Clement, P.A.; Jablonski, R.; Mungari, R.J.; Mavrodieva, V.A.; Negi, S.; Levy, L. First report of plum pox virus on plum in new york state. Plant Dis. 2007, 91, 1512–1512. [Google Scholar]
- Zagrai, I.; Gaboreanu, I.; Ferencz, B.; Zagrai, L.; Pamfil, D.; Popescu, O.; Ravelonandro, M.; Capote, N.; Kovacs, K. First detection and molecular caracterization of plum pox virus recombinant strain in romania. In Bulletin of the university of agricultural sciences and veterinary medicine, vol 62, 2006: Animal husbandry and biotechnologies; 2006; pp. 291–298. [Google Scholar]
- Thompson, D.; Varga, A.; De Costa, H.; Birch, C.; Glasa, M.; James, D. First report of plum pox virus recombinant strain on prunus spp. In canada. Plant Dis. 2009, 93, 674–674. [Google Scholar]
- Kamenova, I.; Dallot, S.; Bozkova, V.; Milusheva, S. First report of the plum pox virus recombinant strain on peach in bulgaria. Plant Dis. 2011, 95, 1320–1321. [Google Scholar]
- Cambra, M.; Capote, N. Plum pox virus (ppv) in kazakhstan. EPPO Bull. 2006, 36, 210–211. [Google Scholar] [CrossRef]
- Navratil, M.; Safarova, D.; Karesova, R.; Petrzik, K. First incidence of plum pox virus on apricot trees in china. Plant Dis. 2005, 89, 338–338. [Google Scholar]
- Dal Zotto, A.; Ortego, J.M.; Raigon, J.M.; Caloggero, S.; Rossini, M.; Ducasse, D.A. First report in argentina of plum pox virus causing sharka disease in prunus. Plant Dis. 2006, 90, 523–523. [Google Scholar]
- Capote, N.; Cambra, M. Plum pox virus (ppv) in pakistan. EPPO Bull. 2006, 36, 212–212. [Google Scholar] [CrossRef]
- Maejima, K.; Hoshi, H.; Hashimoto, M.; Himeno, M.; Kawanishi, T.; Komatsu, K.; Yamaji, Y.; Hamamoto, H.; Namba, S. First report of plum pox virus infecting japanese apricot (prunus mume sieb. Et zucc.) in japan. J. Gen. Plant Pathol. 2010, 76, 229–231. [Google Scholar] [CrossRef]
- Kolber, M.; Nemeth, M.; Chernets, A.; Kalashian, Y.; Dulic-Markovic, I.; Glasa, M.; Isac, M.; Kriska, B.; Malinowski, T.; Zawadzka, B.; et al. Current situation of plum pox disease on stone fruit species in middle and eastern europe. In Proceedings of the 18th international symposium on virus & virus–like diseases of temperate fruit crops – top fruit diseases, vols 1 and 2; INTERNATIONAL SOCIETY HORTICULTURAL SCIENCE: LEUVEN 1, 2001; pp. 73–78. [Google Scholar]
- Mikec, I.; Kajić, V.; Križanac, I. Plum pox virus (ppv) in croatia. EPPO Bull. 2006, 36, 207–207. [Google Scholar]
- Mikec, I.; Kajic, V.; Krajacic, M.; Skoric, D. Occurrence and distribution of plum pox virus in croatia. In Proceedings of the twentieth international symposium on virus and virus–like diseases of temperate fruit crops – fruit tree diseases, 2008; pp. 193–196.
- Isac, M.; Zagrai, I. Plum pox virus (ppv) in romania. EPPO Bull. 2006, 36, 213–213. [Google Scholar] [CrossRef]
- Varveri, C. Plum pox virus (ppv) in greece. EPPO Bull. 2006, 36, 209–210. [Google Scholar] [CrossRef]
- Navrátil, M. Plum pox virus (ppv) in the czech republic. EPPO Bull. 2006, 36, 208–208. [Google Scholar] [CrossRef]
- Jarausch, W. Plum pox virus (ppv) in germany. EPPO Bull. 2006, 36, 209–209. [Google Scholar] [CrossRef]
- Milusheva, S.; Kamenova, I.; Stoev, A. Plum pox virus (ppv) in bulgaria. EPPO Bull. 2006, 36, 206–206. [Google Scholar]
- Kölber, M. Plum pox virus (ppv) in hungary. EPPO Bull. 2006, 36, 210–210. [Google Scholar] [CrossRef]
- Prichodko, Y. Plum pox virus (ppv) in russia. EPPO Bull. 2006, 36, 213–213. [Google Scholar] [CrossRef]
- Malinowski, T. Plum pox virus (ppv) in poland. EPPO Bull. 2006, 36, 212–213. [Google Scholar] [CrossRef]
- EPPO. Plum pox potyvirus. EPPO Bull. 2004, 34, 247–256. [CrossRef]
- Dulic-Markovic, I.; Jevremovic, D. Plum pox virus (ppv) in serbia. EPPO Bull. 2006, 36, 213–214. [Google Scholar] [CrossRef]
- Glasa, M. Plum pox virus (ppv) in slovakia. EPPO Bull. 2006, 36, 214–214. [Google Scholar] [CrossRef]
- Ortego, J.; Dal Zotto, A.; Caloggero, S.; Raigón, J.M.; Gasparini, M.L.; Ojeda, M.E.; Ducasse, D.A. Current status of ppv worldwide - plum pox virus (ppv) in argentina. EPPO Bull. 2006, 36, 205–205. [Google Scholar]
- Stamo, B.; Myrta, A. Current status of ppv worldwide - plum pox virus (ppv) in albania. EPPO Bull. 2006, 36, 205–205. [Google Scholar] [CrossRef]
- Akbas, B.; Degirmenci, K.; Ciftci, O.; Kaya, A.; Yurtmen, M.; Uzunogullari, N.; Celik, N.; Torkolmez, S. Update on plum pox virus distribution in turkey. Phytopathologia Mediterranea 2011, 50, 75–83. [Google Scholar]
- Di Terlizzi, B.; Boscia, D. Plum pox virus (ppv) in italy. EPPO Bull. 2006, 36, 210–210. [Google Scholar] [CrossRef]
- Ortego, J.; Zotto, A.D.; Caloggero, S.; Raigón, J.M.; Gasparini, M.L.; Ojeda, M.E.; Ducasse, D.A. Plum pox virus (ppv) in argentina. EPPO Bull. 2006, 36, 205–205. [Google Scholar]
- Kalashian, Y.; Chernets, A. Plum pox virus (ppv) in moldova. EPPO Bull. 2006, 36, 211–211. [Google Scholar] [CrossRef]
- Youssef, S.A.; Shalaby, A. Plum pox virus (ppv) in egypt. EPPO Bull. 2006, 36, 208–208. [Google Scholar] [CrossRef]
- Norkus, T.; Staniulis, J.; Zizyte, M.; Melnyk, M.; Yusko, L.; Snihur, H.; Budzanivska, I.; Polischuk, V. Molecular identification of plum pox virus isolates from lithuania and ukraine. Zemdirbyste-Agriculture 2008, 95, 277–285. [Google Scholar]
- Wang, A.; Sanfacon, H.; Stobbs, L.W.; James, D.; Thompson, D.; Svircev, A.M.; Brown, D.C.W. Plum pox virus in canada: Progress in research and future prospects for disease control. Can. J. Plant Pathol.-Rev. Can. Phytopathol. 2006, 28, 182–196. [Google Scholar]
- Blystad, D.R.; Munthe, T. Plum pox virus (ppv) in norway. EPPO Bull. 2006, 36, 212–212. [Google Scholar] [CrossRef]
- Zamharir, M.G.; Bashir, N.S.; Khakvar, R. Plum pox virus (ppv) in iran. EPPO Bull. 2006, 36, 210–210. [Google Scholar]
- Speich, P. Plum pox virus (ppv) in france. EPPO Bull. 2006, 36, 208–209. [Google Scholar] [CrossRef]
- Marn, M.V.; Mavric, I. Plum pox virus (ppv) in slovenia. EPPO Bull. 2006, 36, 214–214. [Google Scholar]
- Kondratenko, P.; Udovychenko, V. Plum pox virus (ppv) in ukraine. EPPO Bull. 2006, 36, 217–217. [Google Scholar] [CrossRef]
- Mumford, R.A. Plum pox virus (ppv) in the united kingdom. EPPO Bull. 2006, 36, 217–217. [Google Scholar] [CrossRef]
- Ismaeil, F. Plum pox virus (ppv) in syria. EPPO Bull. 2006, 36, 216–216. [Google Scholar] [CrossRef]
- Levy, L. Plum pox virus (ppv) in the united states of america. EPPO Bull. 2006, 36, 217–218. [Google Scholar] [CrossRef]
- Caglayan, K. Plum pox virus (ppv) in turkey. EPPO Bull. 2006, 36, 216–217. [Google Scholar] [CrossRef]
- Cambra, M.A.; Serra, J.; Cano, A.; Cambra, M. Plum pox virus (ppv) in spain. EPPO Bull. 2006, 36, 215–215. [Google Scholar]
- Muñoz, M.; Collao, M. Plum pox virus (ppv) in chile. EPPO Bull. 2006, 36, 207–207. [Google Scholar]
- Matić, S.; Durić, G.; Myrta, A. Current status of ppv worldwide - plum pox virus (ppv) in bosnia and herzegovina. EPPO Bull. 2006, 36, 205–206. [Google Scholar]
- Staniulis, J. Plum pox virus (ppv) in lithuania. EPPO Bull. 2006, 36, 211–211. [Google Scholar] [CrossRef]
- Verhoeven, J.T.J.; Roenhorst, J.W.; Jongedijk, G.P. Plum pox virus (ppv) in the netherlands. EPPO Bull. 2006, 36, 211–212. [Google Scholar]
- Boulila, M.; Ravelonandro, M. Plum pox virus (ppv) in tunisia. EPPO Bull. 2006, 36, 216–216. [Google Scholar]
- Ramel, M.E.; Gugerli, P.; Bünter, M. Plum pox virus (ppv) in switzerland. EPPO Bull. 2006, 36, 215–216. [Google Scholar]
- Lemmetty, A. Plum pox virus (ppv) in finland. EPPO Bull. 2006, 36, 208–208. [Google Scholar] [CrossRef]
- Choueiri, E. Plum pox virus (ppv) in lebanon. EPPO Bull. 2006, 36, 211–211. [Google Scholar] [CrossRef]
- Navratil, M.; Safarova, D.; Karesova, R.; Petrzik, K. Plum pox virus (ppv) in china. EPPO Bull. 2006, 36, 207–207. [Google Scholar]
- Reyes, F.; Fiore, N.; Reyes, M.A.; Sepulveda, P.; Paredes, V.; Prieto, H. Biological behavior and partial molecular characterization of six chilean isolates of plum pox virus. Plant Dis. 2003, 87, 15–20. [Google Scholar] [CrossRef]
- Manachini, B.; Casati, P.; Aliverti, I.; Cinanni, L. Transmission of ppv-m to prunus persica by brachycaudus schwartzi and phorodon humuli (hem. aphididae). J. Appl. Entomol. 2004, 128, 677–680. [Google Scholar] [CrossRef]
- Zawadzka, B.; Rozpara, E.; Grzyb, Z. The response of some new plum cultivars to plum pox virus (ppv). In Vi international symposium on plum and prune genetics, breeding and pomology; INTERNATIONAL SOCIETY HORTICULTURAL SCIENCE: LEUVEN 1, 1998; pp. 81–85. [Google Scholar]
- Paprstein, F.; Karesova, R.; Navratil, M. Evaluation of ppv symptoms on plum fruits. In Proceedings of the eighth international symposium on plum and prune genetics, breeding and pomology, 2007; pp. 255–257.
- Varveri, C.; Zintzaras, E.; Dimou, D.; Di Terlizzi, B. Monitoring and spatiotemporal analysis of ppv-m spread in two apricot orchards in southern greece. In Proceedings of the 18th international symposium on virus & virus–like diseases of temperate fruit crops – top fruit diseases, vols 1 and 2; INTERNATIONAL SOCIETY HORTICULTURAL SCIENCE: LEUVEN 1, 2001; pp. 129–133. [Google Scholar]
- Clemente-Moreno, M.J.; Piqueras, A.; Hernandez, J.A. Implication of peroxidase activity in development of healthy and ppv-infected micropropagated gf305 peach plants. Plant Growth Regul. 2011, 65, 359–367. [Google Scholar] [CrossRef]
- Pacheco, R.; Garcia-Marcos, A.; Manzano, A.; de Lacoba, M.G.; Camanes, G.; Garcia-Agustin, P.; Diaz-Ruiz, J.R.; Tenllado, F. Comparative analysis of transcriptomic and hormonal responses to compatible and incompatible plant-virus interactions that lead to cell death. Mol. Plant-Microbe Interact. 2012, 25, 709–723. [Google Scholar] [CrossRef]
- Nagyova, A.; Kamencayova, M.; Glasa, M.; Subr, Z.W. The 3'-proximal part of the plum pox virus p1 gene determinates the symptom expression in two herbaceous host plants. Virus Genes 2012, 44, 505–512. [Google Scholar]
- Varveri, C.; Vassilakos, N. Preliminary biological and molecular characterization of mild ppv isolates naturally induced in n-benthamiana plants. In Proceedings of the twentieth international symposium on virus and virus–like diseases of temperate fruit crops – fruit tree diseases; INTERNATIONAL SOCIETY HORTICULTURAL SCIENCE: LEUVEN 1, 2008; pp. 249–253. [Google Scholar]
- Sutic, D.; Jordovic, M.; Rankovic, M.; Festi, H. Comparative studies of some sharka virus isolates. Proceedings of the VIII Symposium sur les Maladies a Virus des Arbres Frutiers. Annales de Phytopathologie, 1971; 1971; pp. 185–194.
- James, D.; Varga, A.; Thompson, D.; Hayes, S. Detection of a new and unusual isolate of plum pox virus in plum (prunus domestica). Plant Dis. 2003, 87, 1119–1124. [Google Scholar] [CrossRef]
- Serce, C.U.; Candresse, T.; Svanella-Dumas, L.; Krizbai, L.; Gazel, M.; Caglayan, K. Further characterization of a new recombinant group of plum pox virus isolates, ppv-t, found in orchards in the ankara province of turkey. Virus Res. 2009, 142, 121–126. [Google Scholar] [CrossRef]
- Matic, S.; Elmaghraby, I.; Law, V.; Varga, A.; Reed, C.; Myrta, A.; James, D. Serological and molecular characterization of isolates of plum pox virus strain el amar to better understand its diversity, evolution, and unique geographical distribution. Journal of Plant Pathology 2011, 93, 303–310. [Google Scholar]
- Candresse, T.; Macquaire, G.; Lanneau, M.; Bousalem, M.; Wetze, T.; Quiot-Douine, L.; Quiot, J.B.; Dunez, J. Detection of plum pox potyvirus and analysis of its molecular variability using immunocapture-pcr1. EPPO Bull. 1994, 24, 585–594. [Google Scholar] [CrossRef]
- Glasa, M.; Malinowski, T.; Predajna, L.; Pupola, N.; Dekena, D.; Michalczuk, L.; Candresse, T. Sequence variability, recombination analysis, and specific detection of the w strain of plum pox virus. Phytopathology 2011, 101, 980–985. [Google Scholar] [CrossRef]
- Glasa, M.; Matisova, J.; Kudela, O. Characterization of plum pox virus isolates from slovakia. Acta Virol. 1998, 42, 226–229. [Google Scholar]
- Navratil, M.; Simonova, V.; Fialova, R.; Valova, P. Molecular variability of czech plum pox virus isolates. Acta Virol. 1998, 42, 254–256. [Google Scholar]
- Navratil, M.; Simonova, V.; Paprstein, F.; Karesova, R. Detection and serological identification of plum pox virus isolates in the czech republic. In 17th international symposium on virus and virus-like diseases of temperate fruit crops: Fruit tree diseases, vols 1 and 2; 1998; pp. 373–378. [Google Scholar]
- Al Rwahnih, M.; Boscia, D.; Myrta, A.; Di Terlizzi, B. First record of plum pox virus in jordan. In Proceedings of the 18th international symposium on virus & virus–like diseases of temperate fruit crops – top fruit diseases, vols 1 and 2, 2001; pp. 141–144.
- Kamenova, I.; Milusheva, S.; Borisova, A.; Stoev, A.; Myrta, A. Typing of plum pox virus isolates in bulgaria: Preliminary results. Biotechnol. Biotechnol. Equip. 2002, 16, 10–13. [Google Scholar]
- Bianco, P.A.; Aliverti, I.; Casati, P.; Belli, G.; Comes, S.; Fanigliulo, A.; Crescenzi, A. Detection and characterization of plum pox virus isolates in lombardia and veneto regions (italy). In Proceedings of the xixth international symposium on virus and virus–like diseases of temperate fruit crops: Fruit tree diseases, 2004; pp. 165–169.
- Cambra, M.; Gorris, M.T.; Capote, N.; Asensio, M.; Martinez, M.C.; Bertolini, E.; Collado, C.; de Mendoza, A.H.; Mataix, E.; Lopez, A. Epidemiology of plum pox virus in japanese plums in spain. In Proceedings of the xixth international symposium on virus and virus–like diseases of temperate fruit crops: Fruit tree diseases, 2004; pp. 195–200.
- Jarausch, W.; Bassler, A.; Molla, N.; Krezal, G. First detection and molecular characterisation of ppv-m strains in plum orchards in south-western germany. In Proceedings of the xixth international symposium on virus and virus–like diseases of temperate fruit crops: Fruit tree diseases, 2004; pp. 159–164.
- Marn, M.V.; Mavric, I. Current status of sharka disease in slovenia. In Proceedings of the xixth international symposium on virus and virus–like diseases of temperate fruit crops: Fruit tree diseases, 2004; pp. 245–249.
- Dallot, S.; Boeglin, M.; Labonne, G. Spatial pattern and genetic structure of ppv-m in a delimited area of stone fruit orchards in southern france. In Proceedings of the twentieth international symposium on virus and virus–like diseases of temperate fruit crops – fruit tree diseases, 2008; pp. 235–242.
- Dallot, S.; Kamenova, I.; Glasa, M.; Pittnerova, S.; Kominek, P.; Paunovic, S.; Jevremovic, D.; Virscek-Marn, M.; Plesko, I.M.; Milusheva, S. Prevalence and genetic structure of ppv-m in six european countries. In Proceedings of the twentieth international symposium on virus and virus–like diseases of temperate fruit crops – fruit tree diseases, 2008; pp. 227–234.
- Kajic, V.; Cerni, S.; Krajacic, M.; Mikec, I.; Skoric, D. Molecular typing of plum pox virus isolates in croatia. J. Plant Pathol. 2008, 90, 9–13. [Google Scholar]
- Zagrai, L.; Zagrai, I.; Ferencz, B.; Gaboreanu, I.; Kovacs, K.; Petricele, I.; Popescu, O.; Pamfil, D.; Capote, N. Serological and molecular typing of plum pox virus isolates in the north of romania. J. Plant Pathol. 2008, 90, 41–46. [Google Scholar]
- Zagrai, I.; Zagrai, L.; Ferencz, B.; Petricele, I.; Pamfil, D.; Popescu, O.; Briciu, A.; Festila, A. Serological and molecular typing of plum pox virus isolates in the transylvania, romania. Not. Bot. Horti Agrobot. Cluj-Na. 2009, 37, 265–272. [Google Scholar]
- Capote, N.; Cambra, M.A.; Botella, P.; Gorris, M.T.; Martinez, M.C.; Lopez-Quilez, A.; Cambra, M. Detection, characterization, epidemiology and eradication of plum pox virus marcus type in spain. J. Plant Pathol. 2010, 92, 619–628. [Google Scholar]
- Zagrai, I.; Zagrai, L.; Preda, S.; Kelemen, B.; Petricele, I.; Popescu, O.; Pamfil, D.; Isac, M. Genetic diversity of plum pox virus isolates in muntenia, romania. Rom. Biotech. Lett. 2010, 15, 5303–5309. [Google Scholar]
- Dallot, S.; Glasa, M.; Jevremovic, D.; Kamenova, I.; Paunovic, S.; Labonne, G. Mediterranean and central-eastern european countries host viruses of two different clades of plum pox virus strain m. Arch. Virol. 2011, 156, 539–542. [Google Scholar] [CrossRef]
- Myrta, A.; Palmisano, F.; Pulaj, B.; Susuri, L.R.; Boscia, D. Incidence of plum pox virus and its strains in kosovo. J. Plant Pathol. 2011, 93, 725–728. [Google Scholar]
- Rosales, M.; Hinrichsen, P.; Herrera, G. Molecular characterization of plum pox virus isolated from apricots, plums and peaches in chile. In 17th international symposium on virus and virus-like diseases of temperate fruit crops: Fruit tree diseases, vols 1 and 2; INTERNATIONAL SOCIETY HORTICULTURAL SCIENCE: LEUVEN 1, 1998; pp. 401–405. [Google Scholar]
- Rochon, D.; Theilmann, J.; James, D.; Reade, R.; Yang, L.; Upton, C. Partial molecular characterization of plum pox virus isolates occurring in canada. Can. J. Plant Pathol.-Rev. Can. Phytopathol. 2003, 25, 198–208. [Google Scholar]
- Marie-Jeanne, V.; Thebaud, G.; Peyre, J.; Labonne, G. Spatially arranged diversity of plum pox virus (ppv) isolates in a cluster of orchards in southern france. In Proceedings of the xixth international symposium on virus and virus-like diseases of temperate fruit crops: Fruit tree diseases,INTERNATIONAL SOCIETY HORTICULTURAL SCIENCE: LEUVEN 1, 2004; INTERNATIONAL SOCIETY HORTICULTURAL SCIENCE: LEUVEN 1; pp. 213–215.
- Elibuyuk, I.O. Current situation of sharka disease in ankara, turkey. Phytoparasitica 2004, 32, 417–420. [Google Scholar] [CrossRef]
- Theilmann, J.; Yang, L.; Rochon, D. Sequence analysis of isolates of the canadian plum pox virus, and comparisons to isolates from europe and the united states. Can. J. Plant Pathol.-Rev. Can. Phytopathol. 2006, 28, 144–151. [Google Scholar]
- Matic, S.; Al-Rwahnih, M.; Myrta, A. Diversity of plum pox virus isolates in bosnia and herzegovina. Plant Pathol. 2006, 55, 11–17. [Google Scholar] [CrossRef]
- Gadiou, S.; Safarova, D.; Navratil, M. Genetic variability of plum pox virus isolates in the czech republic. Eur. J. Plant Pathol. 2008, 121, 513–517. [Google Scholar] [CrossRef]
- Zagrai, I.; Zagrai, L.; Preda, S.; Kelemen, B.; Petricele, I.; Popescu, O.; Pamfil, D.; Isac, M. Genetic diversity of plum pox virus isolates in muntenia, romania. Rom. Biotech. Lett. 2011, 15, 5303–5309. [Google Scholar]
- Glasa, M.; Paunovic, S.; Jevremovic, D.; Myrta, A.; Pittnerova, S.; Candresse, T. Analysis of recombinant plum pox virus (ppv) isolates from serbia confirms genetic homogeneity and supports a regional origin for the ppv-rec subgroup. Arch. Virol. 2005, 150, 2051–2060. [Google Scholar] [CrossRef]
- Navratil, M.; Safarova, D.; Gadiou, S.; Franova, J.; Kucerova, J.; Talacko, L. The partial molecular characterization of plum pox virus infecting sweet cherry trees in the czech republic. In Proceedings of the twentieth international symposium on virus and virus–like diseases of temperate fruit crops – fruit tree diseases; INTERNATIONAL SOCIETY HORTICULTURAL SCIENCE: LEUVEN 1, 2008; pp. 203–208. [Google Scholar]
- Wetzel, T.; Candresse, T.; Ravelonandro, M.; Delbos, R.P.; Mazyad, H.; Aboulata, A.E.; Dunez, J. Nucleotide-sequence of the 3'-terminal region of the rna of the elamar strain of plum pox potyvirus. J. Gen. Virol. 1991, 72, 1741–1746. [Google Scholar] [CrossRef]
- Abdel-Ghaffar, M.H.; El-Nasr, M.A.A.; Hari, V. Studies on an apricot strain of plum pox potyvirus isolated from el amar, egypt. In 17th international symposium on virus and virus-like diseases of temperate fruit crops: Fruit tree diseases, vols 1 and 2; 1998; pp. 385–391. [Google Scholar]
- Aboul-Ela, A.; Aboul-Ata, A.E.; Mazyad, H.M. Plum pox potyvirus situation in egyptian stone fruit trees. In International symposium on apricot culture, vols 1 and 2; INTERNATIONAL SOCIETY HORTICULTURAL SCIENCE: LEUVEN 1, 1999; pp. 745–751. [Google Scholar]
- Myrta, A.; Varga, A.; James, D. The complete genome sequence of an el amar isolate of plum pox virus (ppv) and its phylogenetic relationship to other ppv strains. Arch. Virol. 2006, 151, 1189–1198. [Google Scholar] [CrossRef]
- Matic, S.; Elmaghraby, I.; Law, V.; Varga, A.; Reed, C.; Myrta, A.; James, D. Serological and molecular characterization of isolates of plum pox virus strain el amar to better understand its diversity, evolution, and unique geographical distribution. J. Plant Pathol. 2011, 93, 303–310. [Google Scholar]
- Nemchinov, L.; Hadidi, A.; Kolber, M.; Nemeth, M. Molecular evidence for the occurrence of plum pox virus - cherry subgroup in hungary. In 17th international symposium on virus and virus-like diseases of temperate fruit crops: Fruit tree diseases, vols 1 and 2; INTERNATIONAL SOCIETY HORTICULTURAL SCIENCE: LEUVEN 1, 1998; pp. 503–508. [Google Scholar]
- Nemchinov, L.; Hadidi, A.; Maiss, E.; Cambra, M.; Candresse, T.; Damsteegt, V. Sour cherry strain of plum pox potyvirus (ppv): Molecular and serological evidence for a new subgroup of ppv strains. Phytopathology 1996, 86, 1215–1221. [Google Scholar] [CrossRef]
- Crescenzi, A.; dAquino, L.; Comes, S.; Nuzzaci, M.; Piazzolla, P.; Boscia, D.; Hadidi, A. Characterization of the sweet cherry isolate of plum pox potyvirus. Plant Dis. 1997, 81, 711–714. [Google Scholar] [CrossRef]
- Isac, M.; Plopa, C.; Calinescu, M.; Myrta, A. Detection of the viral diseases presently with the stone fruit species in romania. In Proceedings of the twentieth international symposium on virus and virus–like diseases of temperate fruit crops – fruit tree diseases, 2008; pp. 59–63.
- Schneider, W.L.; Damsteegt, V.D.; Gildow, F.E.; Stone, A.L.; Sherman, D.J.; Levy, L.E.; Mavrodieva, V.; Richwine, N.; Welliver, R.; Luster, D.G. Molecular, ultrastructural, and biological characterization of pennsylvania isolates of plum pox virus. Phytopathology 2011, 101, 627–636. [Google Scholar] [CrossRef]
- Damsteegt, V.D.; Stone, A.L.; Schneider, W.L.; Luster, D.G.; Gildow, F.E. Potential prunus host range of ppv-penn isolates by aphid transmission. In Proceedings of the xixth international symposium on virus and virus–like diseases of temperate fruit crops: Fruit tree diseases; INTERNATIONAL SOCIETY HORTICULTURAL SCIENCE: LEUVEN 1, 2004; pp. 201–205. [Google Scholar]
- James, D.; Varga, A. Nucleotide sequence analysis of plum pox virus isolate w3174: Evidence of a new strain. Virus Res. 2005, 110, 143–150. [Google Scholar] [CrossRef]
- Riechmann, J.L.; Lain, S.; Garcia, J.A. Highlights and prospects of potyvirus molecular-biology. J. Gen. Virol. 1992, 73, 1–16. [Google Scholar] [CrossRef]
- Salvador, B.; García, J.A.; Simón-Mateo, C. Causal agent of sharka disease: Plum pox virus genome and function of gene products. EPPO Bull. 2006, 36, 229–238. [Google Scholar] [CrossRef]
- Teycheney, P.Y.; Tavert, G.; Delbos, R.; Ravelonandro, M.; Dunez, J. The complete nucleotide-sequence of plum pox virus-rna (strain-d). Nucleic Acids Res. 1989, 17, 10115–10116. [Google Scholar] [CrossRef]
- Riechmann, J.L.; Lain, S.; Garcia, J.A. Identification of the initiation codon of plum pox potyvirus genomic rna. Virology 1991, 185, 544–552. [Google Scholar] [CrossRef]
- Khan, M.A.; Goss, D.J. Poly(a)-binding protein increases the binding affinity and kinetic rates of interaction of viral protein linked to genome with translation initiation factors eifiso4f and eifiso4f. 4b complex. Biochemistry 2012, 51, 1388–1395. [Google Scholar]
- Alvarez, V.; Ducasse, D.A.; Biderbost, E.; Nome, S.F. Sequencing and characterization of the coat protein and 3' non-coding region of a new sweet potato potyvirus. Arch. Virol. 1997, 142, 1635–1644. [Google Scholar] [CrossRef]
- Colinet, D.; Kummert, J.; Lepoivre, P. Evidence for the assignment of two strains of splv to the genus potyvirus based on coat protein and 3' non-coding region sequence data. Virus Res. 1997, 49, 91–100. [Google Scholar] [CrossRef]
- Carrington, J.C.; Freed, D.D. Cap-independent enhancement of translation by a plant potyvirus 5' nontranslated region. J. Virol. 1990, 64, 1590–1597. [Google Scholar]
- Kneller, E.L.P.; Rakotondrafara, A.M.; Miller, W.A. Cap-independent translation of plant viral rnas. Virus Res. 2006, 119, 63–75. [Google Scholar] [CrossRef]
- SimonBuela, L.; Guo, H.S.; Garcia, J.A. Cap-independent leaky scanning as the mechanism of translation initiation of a plant viral genomic rna. J. Gen. Virol. 1997, 78, 2691–2699. [Google Scholar]
- Fanigliulo, A.; Comes, S.; Maiss, E.; Piazzolla, P.; Crescenzi, A. The complete nucleotide sequence of plum pox virus isolates from sweet (ppv-swc) and sour (ppv-soc) cherry and their taxonomic relationships within the species. Arch. Virol. 2003, 148, 2137–2153. [Google Scholar] [CrossRef]
- Glasa, M.; Marie-Jeanne, V.; Moury, B.; Kudela, O.; Quiot, J.B. Molecular variability of the p3-6k(1) genomic region among geographically and biologically distinct isolates of plum pox virus. Arch. Virol. 2002, 147, 563–575. [Google Scholar] [CrossRef]
- Poncarova, Z.; Kominek, P. Restriction fragment length polymorphism differentiation of plum pox virus isolates. Acta Virol. 1998, 42, 268–269. [Google Scholar]
- SimonBuela, L.; Guo, H.S.; Garcia, J.A. Long sequences in the 5' noncoding region of plum pox virus are not necessary for viral infectivity but contribute to viral competitiveness and pathogenesis. Virology 1997, 233, 157–162. [Google Scholar] [CrossRef]
- Simon-Buela, L.; Osaba, L.; Garcia, J.A.; Lopez-Moya, J.J. Preservation of 5 '-end integrity of a potyvirus genomic rna is not dependent on template specificity. Virology 2000, 269, 377–382. [Google Scholar] [CrossRef]
- Fernandez-Calvino, L.; Lopez-Abella, D.; Lopez-Moya, J.J.; Fereres, A. Comparison of potato virus y and plum pox virus transmission by two aphid species in relation to their probing behavior. Phytoparasitica 2006, 34, 315–324. [Google Scholar] [CrossRef]
- Fos, A.; Massonie, G. Experimental transmission of plum pox virus by brachycaudus-persicae passerini. Agronomie 1993, 13, 515–518. [Google Scholar] [CrossRef]
- Gildow, F.E.; Damsteegt, V.D.; Stone, A.L.; Schneider, W.L.; Luster, D.G.; Levy, L. Transmission of three north american isolates of plum pox virus: Identification of aphid vectors and species-specific transmission from infected stone fruits. In Proceedings of the xixth international symposium on virus and virus–like diseases of temperate fruit crops: Fruit tree diseases, 2004; pp. 207–211.
- Glasa, M.; Boeglin, M.; Labonne, G. Aphid transmission of natural recombinant plum pox virus isolates to different prunus ssp - a contribution for understanding the epidemiology of an atypical ppv. In Proceedings of the xixth international symposium on virus and virus–like diseases of temperate fruit crops: Fruit tree diseases; INTERNATIONAL SOCIETY HORTICULTURAL SCIENCE: LEUVEN 1, 2004; pp. 217–220. [Google Scholar]
- Lopezmoya, J.J.; Canto, T.; Diazruiz, J.R.; Lopezabella, D. Transmission by aphids of a naturally non-transmissible plum pox virus isolate with the aid of potato-virus-y helper component. J. Gen. Virol. 1995, 76, 2293–2297. [Google Scholar] [CrossRef]
- Paprstein, F.; Karesova, R.; Navratil, M. Results of plum pox virus transmission into germplasm of plums. In Vi international symposium on plum and prune genetics, breeding and pomology; INTERNATIONAL SOCIETY HORTICULTURAL SCIENCE: LEUVEN 1, 1998; pp. 279–282. [Google Scholar]
- Milusheva, S.; Gercheva, P.; Bozhkova, V.; Kamenova, I. Experiments on transmission of plum pox virus through prunus seeds. J. Plant Pathol. 2008, 90, 23–26. [Google Scholar]
- Glasa, M.; Hriovsky, I.; Kudela, O. Evidence for non-transmission of plum pox virus by seed in infected plum and myrobalan. Biologia 1999, 54, 481–484. [Google Scholar]
- Schimanski, H.H.; Gruntzig, M.; Fuchs, E. Non-transmission of the plum pox virus in plum and apricot seed source clones. Zentralblatt Fur Mikrobiologie 1988, 143, 121–123. [Google Scholar]
- Dicenta, F.; Audergon, J.M. Localization of plum pox virus in apricot stem and petiole tissues by tissue printing onto nitrocellulose membrane. Ann. Appl. Biol. 1995, 126, 105–110. [Google Scholar] [CrossRef]
- Hoffmann, A.; Stockert, T.; Hartmann, W.; Stosser, R. Localization of plum pox virus (ppv, sharka) in plum (prunus domestica l.) using tissue print immunoblotting (tpib). J. Plant Dis. Prot. 1997, 104, 411–414. [Google Scholar]
- Rubio, M.; Gosalvez, B.; Hernandez, J.A.; Sanchez-Navarro, J.A.; Pallas, V. Localization of coat protein and nucleic acid of plum pox virus in prunus petiole and stem tissues. In Proceedings of the xixth international symposium on virus and virus–like diseases of temperate fruit crops: Fruit tree diseases; INTERNATIONAL SOCIETY HORTICULTURAL SCIENCE: LEUVEN 1, 2004; pp. 295–300. [Google Scholar]
- Rubio, M.; Martinez-Gomez, P.; Dicenta, F. Study of long-distance movement of plum pox virus (sharka) as an alternative resistance-evaluation method in prunus. Sci. Hortic. 2008, 118, 223–227. [Google Scholar]
- Dicenta, F.; Martinez-Gomez, P.; Rubio, M.; Audergon, J.M. Localisation and movement of plum pox virus in apricot stem tissues. Ann. Appl. Biol. 2003, 142, 99–105. [Google Scholar] [CrossRef]
- Quiot, J.B.; Tab, F.; Labonne, G.; Adamolle, C.; Boeglin, M. Location of plum pox virus in peach and apricot trees. In Proceedings of the xiith ishs symposium on apricot culture and decline, vols 1 and 2; INTERNATIONAL SOCIETY HORTICULTURAL SCIENCE: LEUVEN 1, 2006; pp. 489–492. [Google Scholar]
- Elouaghlidi, F.; Sautter, C. Immunofluorescence localization of plum pox virus and prunus necrotic ringspot virus in tissue-sections of prunus-cerasus and prunus-juxa. J. Plant Dis. Prot. 1988, 95, 70–76. [Google Scholar]
- Hernandez, J.A.; Talavera, J.M.; Martinez-Gomez, P.; Dicenta, F.; Sevilla, F. Response of antioxidative enzymes to plum pox virus in two apricot cultivars. Physiol. Plant. 2001, 111, 313–321. [Google Scholar] [CrossRef]
- Visedo, G.; Fernandezpiqueras, J.; Garcia, J.A. Peroxidase isozyme analysis of factors involved in development of symptoms in nicotiana-clevelandii infected by plum pox virus. Physiol. Plant. 1991, 83, 165–169. [Google Scholar] [CrossRef]
- Visedo, G.; Fernandezpiqueras, J.; Garcia, J.A. Comparison among the isozyme profiles associated with ethrel treatments of leaves, and with senescence and plum pox virus-infection in chenopodium-foetidum. Physiol. Plant. 1991, 83, 159–164. [Google Scholar] [CrossRef]
- Hernandez, J.A.; Olmos, J.E.; Portillo, B.; Rubio, M.; Dicenta, F.; Martinez-Gomez, P. Effect of plum pox virus infection on photosynthesis and antioxidant enzyme activity in peach 'gf305'. In Proceedings of the xixth international symposium on virus and virus–like diseases of temperate fruit crops: Fruit tree diseases; INTERNATIONAL SOCIETY HORTICULTURAL SCIENCE: LEUVEN 1, 2004; pp. 349–353. [Google Scholar]
- Diaz-Vivancos, P.; Clemente-Moreno, M.J.; Rubio, M.; Olmos, E.; Garcia, J.A.; Martinez-Gomez, P.; Hernandez, J.A. Alteration in the chloroplastic metabolism leads to ros accumulation in pea plants in response to plum pox virus. J. Exp. Bot. 2008, 59, 2147–2160. [Google Scholar] [CrossRef]
- Hernandez, J.A.; Diaz-Vivancos, P.; Rubio, M.; Olmos, E.; Ros-Barcelo, A.; Martinez-Gomez, P. Long-term plum pox virus infection produces an oxidative stress in a susceptible apricot, prunus armeniaca, cultivar but not in a resistant cultivar. Physiol. Plant. 2006, 126, 140–152. [Google Scholar] [CrossRef]
- Diaz-Vivancos, P.; Rubio, M.; Mesonero, V.; Martinez-Gomez, P.; Hernandez, J.A.; Periago, P.M. Effect of plum pox virus infection on the antioxidant capacity of leaf apoplast from apricot plants. In Proceedings of the xiiith international symposium on apricot breeding and culture; INTERNATIONAL SOCIETY HORTICULTURAL SCIENCE: LEUVEN 1, 2006; pp. 119–122. [Google Scholar]
- Diaz-Vivancos, P.; Rubio, M.; Mesonero, V.; Periago, P.M.; Barcelo, A.R.; Martinez-Gomez, P.; Hernandez, J.A. The apoplastic antioxidant system in prunus: Response to long-term plum pox virus infection. J. Exp. Bot. 2006, 57, 3813–3824. [Google Scholar] [CrossRef]
- Escalettes, V.S.L.; Hullot, C.; Wawrzy'nczak, D.; Mathieu, E.; Eyquard, J.P.; Le Gall, O.; Decroocq, V. Plum pox virus induces differential gene expression in the partially resistant stone fruit tree prunus armeniaca cv. Goldrich. Gene 2006, 374, 96–103. [Google Scholar] [CrossRef]
- Babu, M.; Griffiths, J.S.; Huang, T.S.; Wang, A. Altered gene expression changes in arabidopsis leaf tissues and protoplasts in response to plum pox virus infection. Bmc Genomics 2008, 9. [Google Scholar]
- Wang, A.; Chapman, P.; Chen, L.; Stobbs, L.W.; Brown, D.C.W.; Brandle, J.E. A comparative survey, by expressed sequence tag analysis, of genes expressed in peach leaves infected with plum pox virus (ppv) and free from ppv. Can. J. Plant Pathol.-Rev. Can. Phytopathol. 2005, 27, 410–419. [Google Scholar]
- Milosevic, T.; Milosevic, N. Plum pox virus as a stress factor in the one-year-old shoot and fruit growth and yield of plum cv. Stanley. Cereal Res. Commun. 2009, 37, 241–244. [Google Scholar]
- Milosevic, T.M.; Glisic, I.P.; Milosevic, N.T.; Glisic, I.S. Plum pox virus as a stress factor in the vegetative growth, fruit growth and yield of plum (prunus domestica) cv. 'Cacanska rodna'. Eur. J. Plant Pathol. 2010, 126, 73–79. [Google Scholar] [CrossRef]
- Garcia-Ibarra, A.; Clemente-Moreno, M.J.; Barba-Espin, G.; Diaz-Vivancos, P.; Rubio, M.; Dicenta, F.; Martinez-Gomez, P.; Hernandez, J.A. Changes in the antioxidative metabolism induced by apple chlorotic leaf spot virus infection in peach [prunus persica (l.) batsch]. Environ. Exp. Bot. 2011, 70, 277–282. [Google Scholar] [CrossRef]
- Amari, K.; Diaz-Vivancos, P.; Pallas, V.; Amelia Sanchez-Pina, M.; Antonio Hernandez, J. Oxidative stress induction by prunus necrotic ringspot virus infection in apricot seeds. Physiol. Plant. 2007, 131, 302–310. [Google Scholar]
- Clarke, S.F.; Guy, P.L.; Burritt, D.J.; Jameson, P.E. Changes in the activities of antioxidant enzymes in response to virus infection and hormone treatment. Physiol. Plant. 2002, 114, 157–164. [Google Scholar] [CrossRef]
- Farkas, G.L.; Kiraly, Z.; Solymosy, F. Role of oxidative metabolism in the localization of plant viruses. Virology 1960, 12, 408–421. [Google Scholar] [CrossRef]
- Fodor, J.; Gullner, G.; Adam, A.L.; Barna, B.; Komives, T.; Kiraly, Z. Local and systemic responses of antioxidants to tobacco mosaic virus infection and to salicylic acid in tobacco - role in systemic acquired resistance. Plant Physiol. 1997, 114, 1443–1451. [Google Scholar]
- Kiraly, L.; Barnaz, B.; Kiralyz, Z. Plant resistance to pathogen infection: Forms and mechanisms of innate and acquired resistance. J. Phytopathol. 2007, 155, 385–396. [Google Scholar] [CrossRef]
- Kröll, J. Experiments to verify the plum pox virus (ppv) by communicating it on chenopodium foetidum schrad. Zentralbl. Bakteriol. Parasitenkd. Infektionskr. Hyg. 1975, 130, 226–231. [Google Scholar]
- Kröll, J. Verification of the plum pox virus (ppv) using the tray test. Zentralbl. Bakteriol. Naturwiss. 1978, 133, 59–64. [Google Scholar]
- Glasa, M.; Matisova, J.; Hricovsky, I.; Kudela, O. Susceptibility of peach gf 305 seedlings and selected herbaceous plants to plum pox virus isolates from western slovakia. Acta Virol. 1997, 41, 341–344. [Google Scholar]
- Polak, J. European spindle tree and common privet a new natural hosts of plum pox virus. In Proceedings of the 18th international symposium on virus & virus–like diseases of temperate fruit crops – top fruit diseases, vols 1 and 2, 2001; pp. 125–128.
- Visedo, G.; Fernandezpiqueras, J.; Garcia, J.A. Isozyme profiles associated with the hypersensitive response of chenopodium-foetidum to plum pox virus-infection. Physiol. Plant. 1990, 78, 218–224. [Google Scholar] [CrossRef]
- Di Terlizzi, B.; Savino, V. Sanitary status and sanitation of stone fruit trees in south east italy. In Iobc/wprs and ishs international conference on integrated fruit production, proceedings of the meeting, 1996; pp. 326–327.
- Paprstein, F.; Karesova, R. Long term evaluation of plum cultivars artificially infected by ppv. In Proceedings of the 18th international symposium on virus & virus–like diseases of temperate fruit crops – top fruit diseases, vols 1 and 2, 2001; pp. 121–123.
- James, D.; Thompson, D. Hosts and symptoms of plum pox virus: Ornamental and wild prunus species. EPPO Bull. 2006, 36, 222–224. [Google Scholar] [CrossRef]
- Rubio, M.; Martinez-Gomez, P.; Pinochet, J.; Dicenta, F. Evaluation of resistance to sharka (plum pox virus) of several prunus rootstocks. Plant Breed. 2005, 124, 67–70. [Google Scholar]
- Martinez-Gomez, P.; Rubio, M.; Gambin, M.; Dicenta, F. Behaviour of six prunus rootstocks in presence of sharka (plum pox potyvirus). In Proceedings of the xiith ishs symposium on apricot culture and decline, vols 1 and 2; INTERNATIONAL SOCIETY HORTICULTURAL SCIENCE: LEUVEN 1, 2006; pp. 227–231. [Google Scholar]
- Zhebentyayeva, T.N.; Reighard, G.L.; Lalli, D.; Gorina, V.M.; Krska, B.; Abbott, A.G. Origin of resistance to plum pox virus in apricot: What new aflp and targeted ssr data analyses tell. Tree Genet. Genomes 2008, 4, 403–417. [Google Scholar] [CrossRef]
- Damsteegt, V.D. The versatility of prunus tomentosa as a bioindicator of viruses. In 17th international symposium on virus and virus-like diseases of temperate fruit crops: Fruit tree diseases, vols 1 and 2; 1998; pp. 143–146. [Google Scholar]
- Damsteegt, V.D.; Waterworth, H.E.; Mink, G.I.; Howell, W.E.; Levy, L. Prunus tomentosa as a diagnostic host for detection of plum pox virus and other prunus viruses. Plant Dis. 1997, 81, 329–332. [Google Scholar] [CrossRef]
- Dosba, F. Progress and prospects in stone fruit breeding. In Genetics and breeding of tree fruits and nuts; INTERNATIONAL SOCIETY HORTICULTURAL SCIENCE: LEUVEN 1, 2003; pp. 35–43. [Google Scholar]
- Barnavetro, I.; Gyorgy, B.; Schuster, V.; Varro, R. Use of peroxidase labeled antibodies for detection of plum pox virus. Acta Phyto. Acad. Sci. Hung. 1980, 15, 333–338. [Google Scholar]
- Adams, A.N.; Guise, C.M.; Crossley, S.J. Plum pox virus detection in dormant plum trees by pcr and elisa. Plant Pathol. 1999, 48, 240–244. [Google Scholar] [CrossRef]
- Caglayan, K.; Sertkaya, G.; Ulubas, C.; Kolber, M.; Krizbai, L. Characterization of plum pox potyvirus (ppv) by das-elisa and rt-pcr/rflp analysis in turkey. In Proceedings of the xixth international symposium on virus and virus–like diseases of temperate fruit crops: Fruit tree diseases, 2004; pp. 183–188.
- Dovas, C.I.; Mamolos, A.P.; Katis, N.I. Fluctuations in concentration of two potyviruses in garlic during the growing period and sampling conditions for reliable detection by elisa. Ann. Appl. Biol. 2002, 140, 21–28. [Google Scholar] [CrossRef]
- Thomidis, T.; Karajiannis, I. Using elisa and pcr to test the potential for spread of plum pox virus by seeds of different stone fruit cultivars. N. Z. J. Crop Hortic. Sci. 2003, 31, 69–72. [Google Scholar] [CrossRef]
- Asensio, M.; Gorris, M.T.; Sanz, A.; Camarasa, E.; Perez, E.; Carbonell, E.A.; Cambra, M. Characterization and detection of plum pox virus using monoclonal antibodies. In Xvith international symposium on fruit tree virus diseases, 1995; pp. 354–356.
- Lopezmoya, J.J.; Sanz, A.; Cambra, M.; Gorris, M.T.; Anaya, C.; Miguet, J.G.; Cortes, E.; Lopezabella, D. Production and characterization of monoclonal-antibodies to plum pox virus and their use in differentiation of mediterranean isolates. Arch. Virol. 1994, 135, 293–304. [Google Scholar] [CrossRef]
- Orban, S.; Theilmann, J.; Rochon, D. Production of polyclonal antibodies for the detection of plum pox virus in leaf tissue by a modified das-elisa assay. Can. J. Plant Pathol.-Rev. Can. Phytopathol. 2007, 29, 86–86. [Google Scholar]
- Richter, J.; Proll, E.; Rabenstein, F.; Stanarius, A. Serological detection of members of the potyviridae with polyclonal antisera. J. Phytopathol. 1994, 142, 11–18. [Google Scholar]
- Hilgert, I.; Cikanek, D.; Kristofova, H.; Karesova, R.; Navratil, M. Monoclonal-antibodies suitable for plum pox virus determination. Hybridoma 1993, 12, 215–220. [Google Scholar] [CrossRef]
- Myrta, A.; Potere, O.; Boscia, D.; Candresse, T.; Cambra, M.; Savino, V. Production of a monoclonal antibody specific to the el amar strain of plum pox virus. Acta Virol. 1998, 42, 248–250. [Google Scholar]
- Cambra, M.; Asensio, M.; Gorris, M.T.; Pérez, E.; Camarasa, E.; Garcia, J.A.; Moya, J.J.; Lopez-Abella, D.; Vela, C.; Sanz, A. Detection of plum pox potyvirus using monoclonal antibodies to structural and non-structural proteins. EPPO Bull. 1994, 24, 569–577. [Google Scholar] [CrossRef]
- Martin, M.T.; Garcia, J.A.; Cervera, M.T.; Goldbach, R.W.; Vanlent, J.W.M. Intracellular-localization of 3 nonstructural plum pox potyvirus proteins by immunogold labeling. Virus Res. 1992, 25, 201–211. [Google Scholar] [CrossRef]
- Purcifull, D.E.; Hiebert, E. Serological relationships involving potyviral nonstructural proteins. Arch. Virol. 1992, 97–122. [Google Scholar] [CrossRef]
- Ravelonandro, M.; Peyruchaud, O.; Garrigue, L.; Demarcillac, G.; Dunez, J. Immunodetection of the plum pox virus helper component in infected plants and expression of its gene in transgenic plants. Arch. Virol. 1993, 130, 251–268. [Google Scholar] [CrossRef]
- Gil, M.; Esteban, O.; Garcia, J.A.; Pena, L.; Cambra, M. Resistance to plum pox virus in plants expressing cytosolic and nuclear single-chain antibodies against the viral rna nib replicase. Plant Pathol. 2011, 60, 967–976. [Google Scholar] [CrossRef]
- Olmos, A.; Cambra, M.; Dasi, M.A.; Candresse, T.; Esteban, O.; Gorris, M.T.; Asensio, M. Simultaneous detection and typing of plum pox potyvirus (ppv) isolates by heminested-pcr and pcr-elisa. J. Virol. Methods 1997, 68, 127–137. [Google Scholar] [CrossRef]
- Kamenova, I.; Peters, D. The differential reactivity of two polyclonal antisera to plum pox virus isolates collected at different stages after immunization. Biotechnol. Biotechnol. Equip. 1999, 13, 33–39. [Google Scholar]
- Subr, Z.; Matisova, J. Preparation of diagnostic monoclonal antibodies against two potyviruses. Acta Virol. 1999, 43, 255–257. [Google Scholar]
- Himmler, G.; Brix, U.; Steinkellner, H.; Laimer, M.; Mattanovich, D.; Katinger, H.W.D. Early screening for anti-plum pox virus monoclonal-antibodies with different epitope specificities by means of gold-labeled immunosorbent electron-microscopy. J. Virol. Methods 1988, 22, 351–358. [Google Scholar] [CrossRef]
- Jarocka, U.; Wasowicz, M.; Radecka, H.; Malinowski, T.; Michalczuk, L.; Radecki, J. Impedimetric immunosensor for detection of plum pox virus in plant extracts. Electroanalysis 2011, 23, 2197–2204. [Google Scholar] [CrossRef]
- Pollini, C.P.; Giunchedi, L.; Bissani, R. Specific detection of d- and m-isolates of plum pox virus by immunoenzymatic determination of pcr products. J. Virol. Methods 1997, 67, 127–133. [Google Scholar] [CrossRef]
- Youssef, S.A.; Shalaby, A.A.; Mazyad, H.M.; Hadidi, A. Detection and identification of prune dwarf virus and plum pox virus by standard and multiplex rt-pcr probe capture hybridization (rt-pcr-elisa). J. Plant Pathol. 2002, 84, 113–119. [Google Scholar]
- Wetzel, T.; Candresse, T.; Ravelonandro, M.; Delbos, R.P.; Mazyad, H.; Aboulata, A.E.; Dunez, J. Nucleotide-sequence of the 3'-terminal region of the rna of the elamar strain of plum pox potyvirus. Journal of General Virology 1991, 72, 1741–1746. [Google Scholar] [CrossRef]
- Wetzel, T.; Candresse, T.; Ravelonandro, M.; Dunez, J. A polymerase chain-reaction assay adapted to plum pox potyvirus detection. J. Virol. Methods 1991, 33, 355–365. [Google Scholar] [CrossRef]
- Subr, Z.; Glasa, M. Plum pox virus variability detected by the advanced analytical methods. Acta Virol. 2008, 52, 75–89. [Google Scholar]
- Wetzel, T.; Tavert, G.; Teycheney, P.Y.; Ravelonandro, M.; Candresse, T.; Dunez, J. Dot hybridization detection of plum pox virus using p-32 labeled rna probes representing nonstructural viral protein genes. J. Virol. Methods 1990, 30, 161–171. [Google Scholar] [CrossRef]
- Varveri, C.; Candresse, T.; Cugusi, M.; Ravelonandro, M.; Dunez, J. Use of a p-32-labeled transcribed rna probe for dot hybridization detection of plum pox virus. Phytopathology 1988, 78, 1280–1283. [Google Scholar] [CrossRef]
- Hammond, J.; Puhringer, H.; Machado, A.D.; Machado, M.L.D. A broad-spectrum pcr assay combined with rflp analysis for detection and differentiation of plum pox virus isolates. In 17th international symposium on virus and virus-like diseases of temperate fruit crops: Fruit tree diseases, vols 1 and 2; INTERNATIONAL SOCIETY HORTICULTURAL SCIENCE: LEUVEN 1, 1998; pp. 483–490. [Google Scholar]
- Gadiou, S.; Safarova, D.; Navratil, M. Differentiation of plum pox virus isolates by single-strand conformation polymorphism and low-stringency single specific primer pcr analysis of hc-pro genome region. Acta Virol. 2009, 53, 53–56. [Google Scholar]
- Subr, Z.; Pittnerova, S.; Glasa, M. A simplified rt-pcr-based detection of recombinant plum pox virus isolates. Acta Virol. 2004, 48, 173–176. [Google Scholar]
- Spiegel, S.; Kovalenko, E.M.; Varga, A.; James, D. Detection and partial molecular characterization of two plum pox virus isolates from plum and wild apricot in southeast kazakhstan. Plant Dis. 2004, 88, 973–979. [Google Scholar] [CrossRef]
- Szemes, M.; Kalman, M.; Myrta, A.; Boscia, D.; Nemeth, M.; Kolber, M.; Dorgai, L. Integrated rt-pcr/nested pcr diagnosis for differentiating between subgroups of plum pox virus. J. Virol. Methods 2001, 92, 165–175. [Google Scholar] [CrossRef]
- Pasquini, G.; Barba, M.; Hadidi, A.; Faggioli, F.; Negri, R.; Sobol, I.; Tiberini, A.; Caglayan, K.; Mazyad, H.; Anfoka, G.; et al. Oligonucleotide microarray-based detection and genotyping of plum pox virus. J. Virol. Methods 2008, 147, 118–126. [Google Scholar] [CrossRef]
- Olmos, A.; Bertolini, E.; Gil, M.; Cambra, M. Real-time assay for quantitative detection of non-persistently transmitted plum pox virus rna targets in single aphids. J. Virol. Methods 2005, 128, 151–155. [Google Scholar] [CrossRef]
- Capote, N.; Gorris, M.T.; Martinez, M.C.; Asensio, M.; Olmos, A.; Cambra, M. Interference between d and m types of plum pox virus in japanese plum assessed by specific monoclonal antibodies and quantitative real-time reverse transcription-polymerase chain reaction. Phytopathology 2006, 96, 320–325. [Google Scholar] [CrossRef]
- Kim, W.S.; Stobbs, L.W.; Lehman, S.M.; James, D.; Svircev, A.M. Direct real-time pcr detection of plum pox virus in field surveys in ontario. Can. J. Plant Pathol.-Rev. Can. Phytopathol. 2008, 30, 308–317. [Google Scholar]
- Capote, N.; Bertolini, E.; Olmos, A.; Vidal, E.; Martinez, M.C.; Cambra, M. Direct sample preparation methods for the detection of plum pox virus by real-time rt-pcr. Int. Microbiol. 2009, 12, 1–6. [Google Scholar]
- Levy, L.; Kolber, M.; Tokes, G.; Nemeth, M.; Hadidi, A. 3' non-coding region rt-pcr detection and molecular hybridization of plum pox virus in anthers of infected stone fruit. In Xvith international symposium on fruit tree virus diseases; INTERNATIONAL SOCIETY HORTICULTURAL SCIENCE: LOUVAIN, 1995; pp. 331–339. [Google Scholar]
- Schneider, W.L.; Sherman, D.J.; Stone, A.L.; Damsteegt, V.D.; Frederick, R.D. Specific detection and quantification of plum pox virus by real-time fluorescent reverse transcription-pcr. J. Virol. Methods 2004, 120, 97–105. [Google Scholar] [CrossRef]
- Varga, A.; James, D. Real-time rt-pcr and sybr green i melting curve analysis for the identification of plum pox virus strains c, ea, and w: Effect of amplicon size, melt rate, and dye translocation. J. Virol. Methods 2006, 132, 146–153. [Google Scholar] [CrossRef]
- Varga, A.; James, D. Detection and differentiation of plum pox virus using real-time multiplex pcr with sybr green and melting curve analysis: A rapid method for strain typing. J. Virol. Methods 2005, 123, 213–220. [Google Scholar] [CrossRef]
- Korschineck, I.; Himmler, G.; Sagl, R.; Steinkellner, H.; Katinger, H.W.D. A pcr membrane spot assay for the detection of plum pox virus-rna in bark of infected trees. J. Virol. Methods 1991, 31, 139–146. [Google Scholar] [CrossRef]
- Olmos, A.; Bertolini, E.; Cambra, M. Simultaneous and co-operational amplification (co-pcr): A new concept for detection of plant viruses. J. Virol. Methods 2002, 106, 51–59. [Google Scholar] [CrossRef]
- Bertolini, E.; Torres, E.; Olmos, A.; Martin, M.P.; Bertaccini, A.; Cambra, M. Co-operational pcr coupled with dot blot hybridization for detection and 16srx grouping of phytoplasmas. Plant Pathol. 2007, 56, 677–682. [Google Scholar] [CrossRef]
- Martos, S.; Torres, E.; El Bakali, M.A.; Raposo, R.; Gramaje, D.; Armengol, J.; Luque, J. Co-operational pcr coupled with dot blot hybridization for the detection of phaeomoniella chlamydospora on infected grapevine wood. J. Phytopathol. 2011, 159, 247–254. [Google Scholar] [CrossRef]
- Wetzel, T.; Candresse, T.; Macquaire, G.; Ravelonandro, M.; Dunez, J. A highly sensitive immunocapture polymerase chain-reaction method for plum pox potyvirus detection. J. Virol. Methods 1992, 39, 27–37. [Google Scholar] [CrossRef]
- Sreenivasulu, M.; Gopal, D. Development of recombinant coat protein antibody based ic-rt-pcr and comparison of its sensitivity with other immunoassays for the detection of papaya ringspot virus isolates from india, (vol 26, pg 25, 2010). Plant Pathol. J. 2010, 26, 207. [Google Scholar] [CrossRef]
- Sreenivasulu, M.; Gopal, D. Development of recombinant coat protein antibody based ic-rt-pcr and comparison of its sensitivity with other immunoassays for the detection of papaya ringspot virus isolates from india. Plant Pathol. J. 2010, 26, 25–31. [Google Scholar] [CrossRef]
- Huang, C.H.; Hu, W.C.; Yang, T.C.; Chang, Y.C. Zantedeschia mild mosaic virus, a new widespread virus in calla lily, detected by elisa, dot-blot hybridization and ic-rt-pcr. Plant Pathol. 2007, 56, 183–189. [Google Scholar]
- Hema, M.; Kirthi, N.; Sreeivasulu, P.; Savithri, H.S. Development of recombinant coat protein antibody based ic-rt-pcr for detection and discrimination of sugarcane streak mosaic virus isolates from southern india. Arch. Virol. 2003, 148, 1185–1193. [Google Scholar] [CrossRef]
- Olmos, A.; Bertolini, E.; Cambra, M. Isothermal amplification coupled with rapid flow-through hybridisation for sensitive diagnosis of plum pox virus. J. Virol. Methods 2007, 139, 111–115. [Google Scholar] [CrossRef]
- Olmos, A.; Bertolini, E.; Cambra, M. Isothermal amplification for detection of plum pox virus. In Proceedings of the twentieth international symposium on virus and virus–like diseases of temperate fruit crops – fruit tree diseases; INTERNATIONAL SOCIETY HORTICULTURAL SCIENCE: LEUVEN 1, 2008; pp. 209–213. [Google Scholar]
- Hadersdorfer, J.; Neumuller, M.; Treutter, D.; Fischer, T.C. Fast and reliable detection of plum pox virus in woody host plants using the blue lamp protocol. Ann. Appl. Biol. 2011, 159, 456–466. [Google Scholar] [CrossRef]
- Capote, N.; Bertolini, E.; Martinez, M.C.; Olmos, A.; Gorris, M.T.; Cambra, M. Spot real-time rt-pcr: A method for direct detection of plum pox virus avoiding rna extraction. In Proceedings of the twentieth international symposium on virus and virus–like diseases of temperate fruit crops – fruit tree diseases; INTERNATIONAL SOCIETY HORTICULTURAL SCIENCE: LEUVEN 1, 2008; pp. 215–219. [Google Scholar]
- Sanchez, F.; Sobolev, K. Nanotechnology in concrete - a review. Constr. Build. Mater. 2010, 24, 2060–2071. [Google Scholar] [CrossRef]
- Shea, C.M.; Grinde, R.; Elmslie, B. Nanotechnology as general-purpose technology: Empirical evidence and implications. Tech. Anal. Strat. Manag. 2011, 23, 175–192. [Google Scholar] [CrossRef]
- Banerjee, R.; Katsenovich, Y.; Lagos, L.; McIintosh, M.; Zhang, X.; Li, C.Z. Nanomedicine: Magnetic nanoparticles and their biomedical applications. Curr. Med. Chem. 2010, 17, 3120–3141. [Google Scholar] [CrossRef]
- Cai, S.; Yang, Q.H.; Bagby, T.R.; Forrest, M.L. Lymphatic drug delivery using engineered liposomes and solid lipid nanoparticles. Adv. Drug Deliv. Rev. 2011, 63, 901–908. [Google Scholar] [CrossRef]
- Feng, B.; Matsui, H.; Tomizawa, K. Nanoparticle-based drug delivery systems for solid brain tumors. Curr. Nanosci. 2011, 7, 47–54. [Google Scholar] [CrossRef]
- Ghaderi, S.; Ramesh, B.; Seifalian, A.M. Fluorescence nanoparticles "quantum dots" as drug delivery system and their toxicity: A review. J. Drug Target. 2011, 19, 475–486. [Google Scholar] [CrossRef]
- Chen, G.J.; Wang, L.F. Design of magnetic nanoparticles-assisted drug delivery system. Curr. Pharm. Design 2011, 17, 2331–2351. [Google Scholar] [CrossRef]
- Malam, Y.; Lim, E.J.; Seifalian, A.M. Current trends in the application of nanoparticles in drug delivery. Curr. Med. Chem. 2011, 18, 1067–1078. [Google Scholar] [CrossRef]
- Parveen, S.; Misra, R.; Sahoo, S.K. Nanoparticles: A boon to drug delivery, therapeutics, diagnostics and imaging. Nanomed.-Nanotechnol. Biol. Med. 2012, 8, 147–166. [Google Scholar] [CrossRef]
- Paulo, C.S.O.; das Neves, R.P.; Ferreira, L.S. Nanoparticles for intracellular-targeted drug delivery. Nanotechnology 2011, 22. [Google Scholar]
- Prow, T.W.; Grice, J.E.; Lin, L.L.; Faye, R.; Butler, M.; Becker, W.; Wurm, E.M.T.; Yoong, C.; Robertson, T.A.; Soyer, H.P.; et al. Nanoparticles and microparticles for skin drug delivery. Adv. Drug Deliv. Rev. 2011, 63, 470–491. [Google Scholar] [CrossRef]
- Su, X.; Zhan, X.; Tang, F.; Yao, J.Y.; Wu, J. Magnetic nanoparticles in brain disease diagnosis and targeting drug delivery. Curr. Nanosci. 2011, 7, 37–46. [Google Scholar] [CrossRef]
- Talekar, M.; Kendall, J.; Denny, W.; Garg, S. Targeting of nanoparticles in cancer: Drug delivery and diagnostics. Anti-Cancer Drugs 2011, 22, 949–962. [Google Scholar] [CrossRef]
- Yoo, J.W.; Doshi, N.; Mitragotri, S. Adaptive micro and nanoparticles: Temporal control over carrier properties to facilitate drug delivery. Adv. Drug Deliv. Rev. 2011, 63, 1247–1256. [Google Scholar] [CrossRef]
- Eggenberger, K.; Frey, N.; Zienicke, B.; Siebenbrock, J.; Schunck, T.; Fischer, R.; Brase, S.; Birtalan, E.; Nann, T.; Nick, P. Use of nanoparticles to study and manipulate plant cells. Adv. Eng. Mater. 2010, 12, B406–B412. [Google Scholar] [CrossRef]
- Baruah, S.; Dutta, J. Nanotechnology applications in pollution sensing and degradation in agriculture: A review. Environ. Chem. Lett. 2009, 7, 191–204. [Google Scholar] [CrossRef]
- Knauer, K.; Bucheli, T. Nano-materials - the need for research in agriculture. Agrarforschung 2009, 16, 390–395. [Google Scholar]
- Knauer, K.; Bucheli, T.D. Nano-materials: Research needs in agriculture. Rev. Suisse Agric. 2009, 41, 341–345. [Google Scholar]
- Shrivastava, S.; Dash, D. Agrifood nanotechnology: A tiny revolution in food and agriculture. J. Nano Res. 2009, 6, 1–14. [Google Scholar] [CrossRef]
- Lokko, Y.; Mba, C.; Spencer, M.; Till, B.; Lagoda, P. Nanotechnology and synthetic biology - potential in crop improvement. J. Food Agric. Environ. 2011, 9, 599–604. [Google Scholar]
- Young, M.; Willits, D.; Uchida, M.; Douglas, T. Plant viruses as biotemplates for materials and their use in nanotechnology. In Annu. Rev. Phytopathol. 2008, 361–384. [Google Scholar]
- Aniagyei, S.E.; DuFort, C.; Kao, C.C.; Dragnea, B. Self-assembly approaches to nanomaterial encapsulation in viral protein cages. J. Mater. Chem. 2008, 18, 3763–3774. [Google Scholar] [CrossRef]
- Srivastava, S.K. Use of plant viruses and their genes in the areas of biotechnology and nanotechnology. Indian J. Virol. 2008, 19, 101–102. [Google Scholar]
- Behrens, S.S. Synthesis of inorganic nanomaterials mediated by protein assemblies. J. Mater. Chem. 2008, 18, 3788–3798. [Google Scholar] [CrossRef]
- Lee, S.Y.; Lim, J.S.; Harris, M.T. Synthesis and application of virus-based hybrid nanomaterials. Biotechnol. Bioeng. 2012, 109, 16–30. [Google Scholar] [CrossRef]
- Royston, E.; Ghosh, A.; Kofinas, P.; Harris, M.T.; Culver, J.N. Self-assembly of virus-structured high surface area nanomaterials and their application as battery electrodes. Langmuir 2008, 24, 906–912. [Google Scholar] [CrossRef]
- Soto, C.M.; Ratna, B.R. Virus hybrids as nanomaterials for biotechnology. Curr. Opin. Biotechnol. 2010, 21, 426–438. [Google Scholar] [CrossRef]
- Zhou, Y. Recent progress in biomolecule-templated nanomaterials. Curr. Nanosci. 2006, 2, 123–134. [Google Scholar] [CrossRef]
- Flenniken, M.L.; Uchida, M.; Liepold, L.O.; Kang, S.; Young, M.J.; Douglas, T. A library of protein cage architectures as nanomaterials. In Viruses and nanotechnology; 2009; pp. 71–93. [Google Scholar]
- Franzen, S.; Lockney, D.M.; Wang, R.Q.; Lommel, S.; Hauck, M. In vivo imaging and tumor targeting using a plant virus nanoparticle. Abstr. Pap. Am. Chem. Soc. 2011, 241. [Google Scholar]
- Lockney, D.M.; Guenther, R.N.; Loo, L.; Overton, W.; Antonelli, R.; Clark, J.; Hu, M.; Luft, C.; Lommel, S.A.; Franzen, S. The red clover necrotic mosaic virus capsid as a multifunctional cell targeting plant viral nanoparticle. Bioconjugate Chem. 2011, 22, 67–73. [Google Scholar] [CrossRef]
- Liang, M.M.; Guo, L.H. Application of nanomaterials in environmental analysis and monitoring. J. Nanosci. Nanotechnol. 2009, 9, 2283–2289. [Google Scholar] [CrossRef]
- Martin, C.R. Nanomaterials: Evolving applications in membranes, energy production, and analytical chemistry. Abstr. Pap. Am. Chem. Soc. 2000, 219, U727–U727. [Google Scholar]
- Martin, C.R.; Mitchell, D.T. Nanomaterials in analytical chemistry. Anal. Chem. 1998, 70, 322–327. [Google Scholar] [CrossRef]
- Scida, K.; Stege, P.W.; Haby, G.; Messina, G.A.; Garcia, C.D. Recent applications of carbon-based nanomaterials in analytical chemistry: Critical review. Anal. Chim. Acta 2011, 691, 6–17. [Google Scholar] [CrossRef]
- Valentini, F.; Palleschi, G. Nanomaterials and analytical chemistry. Anal. Lett. 2008, 41, 479–520. [Google Scholar] [CrossRef]
- Bandyopadhyay, A.; Chatterjee, S.; Sarkar, K. Rapid isolation of genomic DNA from e. Coli xl1 blue strain approaching bare magnetic nanoparticles. Curr. Sci. 2011, 101, 210–214. [Google Scholar]
- Chiang, C.L.; Sung, C.S.; Chen, C.Y. Application of silica-magnetite nanocomposites to the isolation of ultrapure plasmid DNA from bacterial cells. J. Magn. Magn. Mater. 2006, 305, 483–490. [Google Scholar] [CrossRef]
- Sarkar, T.R.; Irudayaraj, J. Carboxyl-coated magnetic nanoparticles for mrna isolation and extraction of supercoiled plasmid DNA. Anal. Biochem. 2008, 379, 130–132. [Google Scholar]
- Zhang, H.P.; Bai, S.; Xu, L.; Sun, Y. Fabrication of mono-sized magnetic anion exchange beads for plasmid DNA purification. J. Chromatogr. B 2009, 877, 127–133. [Google Scholar] [CrossRef]
- Masarik, M.; Gumulec, J.; Sztalmachova, M.; Hlavna, M.; Babula, P.; Krizkova, S.; Ryvolova, M.; Jurajda, M.; Sochor, J.; Adam, V.; et al. Isolation of metallothionein from cells derived from aggressive form of high-grade prostate carcinoma using paramagnetic antibody-modified microbeads off-line coupled with electrochemical and electrophoretic analysis. Electrophoresis 2011, 32, 3576–3588. [Google Scholar]
- Adam, V.; Huska, D.; Hubalek, J.; Kizek, R. Easy to use and rapid isolation and detection of a viral nucleic acid by using paramagnetic microparticles and carbon nanotubes-based screen-printed electrodes. Microfluid. Nanofluid. 2010, 8, 329–339. [Google Scholar] [CrossRef]
- Huska, D.; Hubalek, J.; Adam, V.; Vajtr, D.; Horna, A.; Trnkova, L.; Havel, L.; Kizek, R. Automated nucleic acids isolation using paramagnetic microparticles coupled with electrochemical detection. Talanta 2009, 79, 402–411. [Google Scholar] [CrossRef]
- Masarik, M.; Huska, D.; Adam, V.; Hubalek, J.; Provaznik, I.; Trnkova, L.; Kizek, R. Automated pipetting system coupled with micro- and nanopartieles as a new tool for study of nucleic acids. Int. J. Mol. Med. 2010, 26, S46–S46. [Google Scholar]
- Wang, J.; Hui, F.; Fang, X.W.; Vinciguerra, J.; Hunicke-Smith, S.; WalkerPeach, C.R. Magnetic bead-based viral nucleic acid isolation incorporating a quantitative internal control. J. Mol. Diagn. 2004, 6, 427–427. [Google Scholar]
- Zhang, Z.C.; Cui, Y.; Wan, Q.H. Surface modification of magnetic silica microspheres and its application to the isolation of plant genomic nucleic acids. Chin. J. Anal. Chem. 2007, 35, 31–36. [Google Scholar] [CrossRef]
- Miszczak, F.; Shuck, K.M.; Lu, Z.C.; Go, Y.Y.; Zhang, J.Q.; Sells, S.; Vabret, A.; Pronost, S.; Fortier, G.; Timoney, P.J.; et al. Evaluation of two magnetic-bead-based viral nucleic acid purification kits and three real-time reverse transcription-pcr reagent systems in two taqman assays for equine arteritis virus detection in semen. J. Clin. Microbiol. 2011, 49, 3694–3696. [Google Scholar] [CrossRef]
- Sun, H.X.; Zeng, X.; Liu, M.; Elingarami, S.; Li, G.P.; Shen, B.; He, N.Y. Synthesis of size-controlled fe3o4@sio2 magnetic nanoparticles for nucleic acid analysis. J. Nanosci. Nanotechnol. 2012, 12, 267–273. [Google Scholar] [CrossRef]
- Yang, S.Y.; Chieh, J.J.; Wang, W.C.; Yu, C.Y.; Hing, N.S.; Horng, H.E.; Hong, C.Y.; Yang, H.C.; Chang, C.F.; Lin, H.Y. Magnetic nanoparticles for high-sensitivity detection on nucleic acids via superconducting-quantum-interference-device-based immunomagnetic reduction assay. J. Magn. Magn. Mater. 2011, 323, 681–685. [Google Scholar] [CrossRef]
- Byzova, N.A.; Safenkova, I.V.; Chirkov, S.N.; Avdienko, V.G.; Guseva, A.N.; Mitrofanova, I.V.; Zherdev, A.V.; Dzantiev, B.B.; Atabekov, J.G. Interaction of plum pox virus with specific colloidal gold-labeled antibodies and development of immunochromatographic assay of the virus. Biochem.-Moscow 2010, 75, 1393–1403. [Google Scholar] [CrossRef]
- Safenkova, I.V.; Zherdev, A.V.; Dzantiev, B.B. Correlation between the composition of multivalent antibody conjugates with colloidal gold nanoparticles and their affinity. J. Immunol. Methods 2010, 357, 17–25. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Sochor, J.; Babula, P.; Adam, V.; Krska, B.; Kizek, R. Sharka: The Past, The Present and The Future. Viruses 2012, 4, 2853-2901. https://doi.org/10.3390/v4112853
Sochor J, Babula P, Adam V, Krska B, Kizek R. Sharka: The Past, The Present and The Future. Viruses. 2012; 4(11):2853-2901. https://doi.org/10.3390/v4112853
Chicago/Turabian StyleSochor, Jiri, Petr Babula, Vojtech Adam, Boris Krska, and Rene Kizek. 2012. "Sharka: The Past, The Present and The Future" Viruses 4, no. 11: 2853-2901. https://doi.org/10.3390/v4112853
APA StyleSochor, J., Babula, P., Adam, V., Krska, B., & Kizek, R. (2012). Sharka: The Past, The Present and The Future. Viruses, 4(11), 2853-2901. https://doi.org/10.3390/v4112853