Higher Mosquito Production in Low-Income Neighborhoods of Baltimore and Washington, DC: Understanding Ecological Drivers and Mosquito-Borne Disease Risk in Temperate Cities


Abstract
:1. Introduction
2. Mosquitoes and Coupled Natural-Human Systems
3. Experimental Section
3.1. Case Study in Washington, DC and Baltimore, MD
3.2. Mosquito Species in Northeastern United States
3.3. Sampling Protocol
3.4. Analyses
4. Results and Discussion
4.1. Species Composition

{kind=link}
{kind=link}
| Neighborhood | City | Relative Median Income | Culex pipiens | Culex restuans | Aedes albopictus | Aedes triseriatus |
|---|---|---|---|---|---|---|
| N1 | Baltimore | L | 12.72% | 0.29% | 83.09% | 3.51% |
| N2 | Baltimore | M | 53.66% | 14.81% | 30.64% | 0.48% |
| N3 | Washington | L | 1.76% | 0.08% | 93.57% | 4.21% |
| N4 | Washington | M | 0.16% | 0.00% | 99.84% | 0.00% |
| N5 | Washington | H | 0.00% | 0.00% | 90.27% | 9.73% |
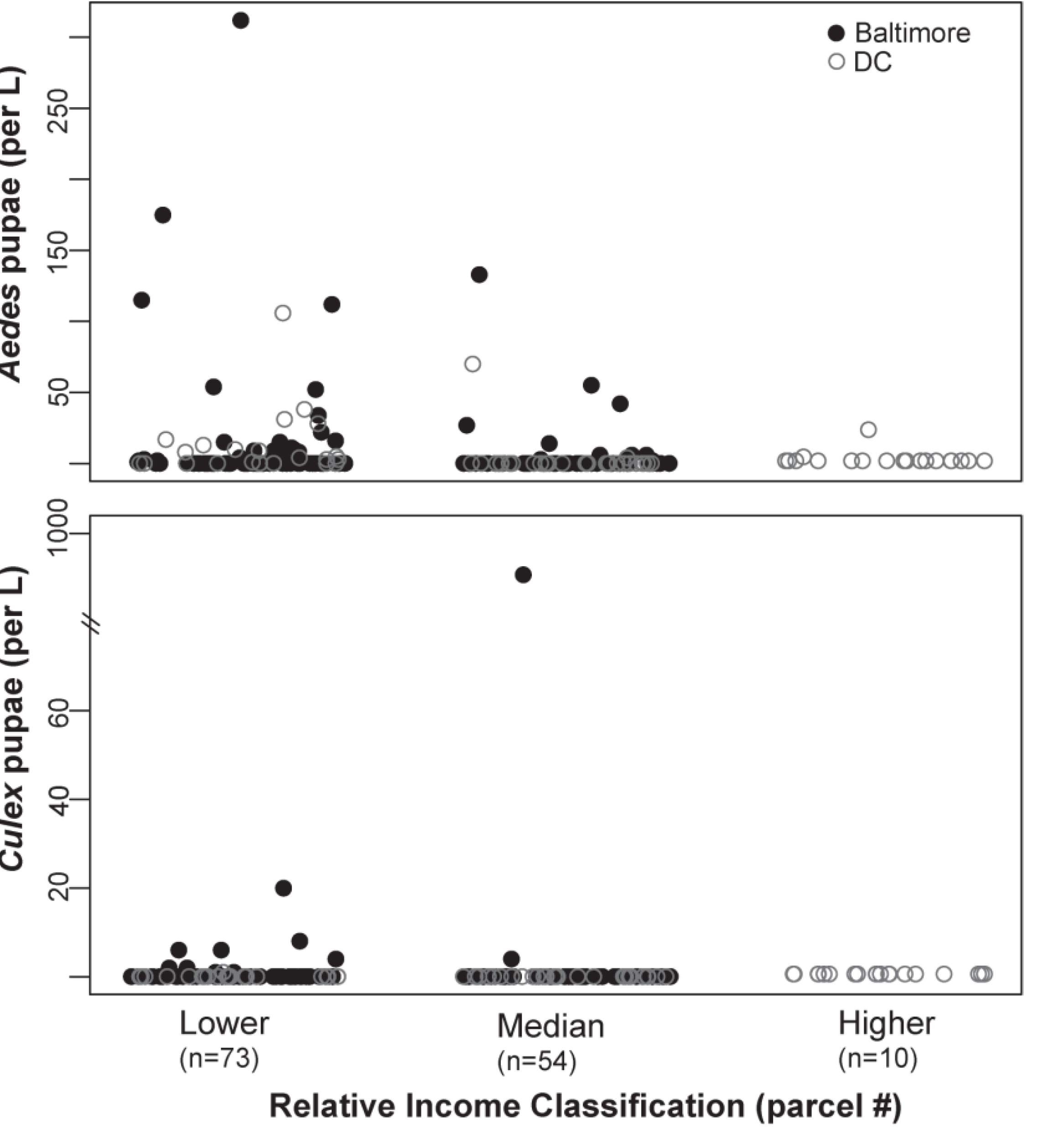
4.2. Potential Vector Production across Relative Income Categories
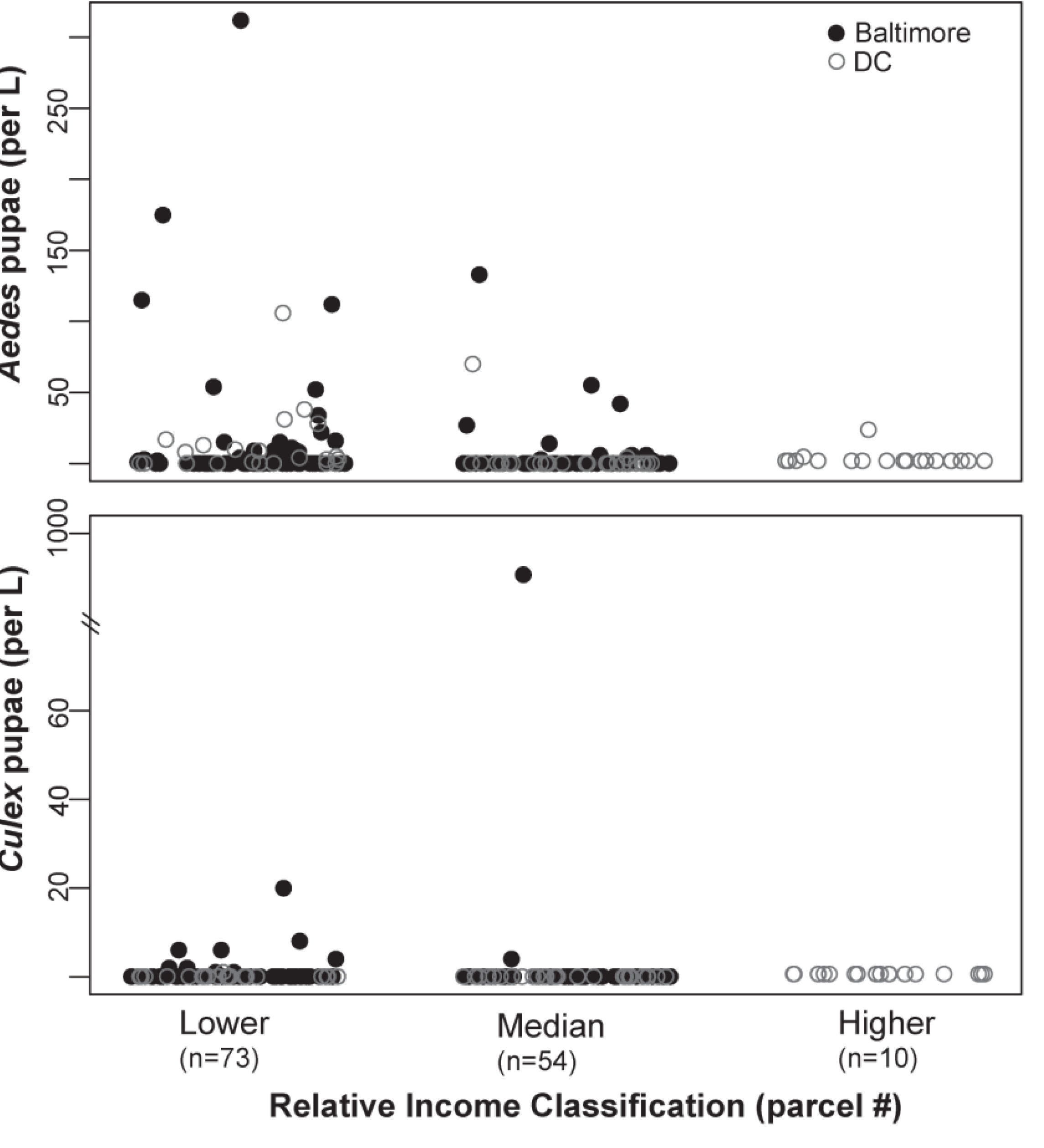
4.3. The Composition of Container Habitats
| Container Function | Container # per parcel | Density Coefficient (pupae) | |||
|---|---|---|---|---|---|
| Lower | Median | Higher | Culex | Aedes | |
| Storage | 0.133 | 0.087 | 0.222 | ns | 2.19 (0.17) p < 0.001 |
| Recreation | 0.067 | 0.065 | 0.111 | ns | ns |
| Structural | 0.100 | 0.087 | 0.444 | ns | ns |
| Yard care | 0.666 | 1.152 | 1.222 | ns | 1.53 (0.13) p < 0.001 |
| Trash | 0.567 | 0.500 | 0.000 | ns | ns |
| Tires | 0.375 | 0.057 | 0.000 | 5.83 (1.95) p = 0.003 | 3.14 (0.53) p < 0.001 |
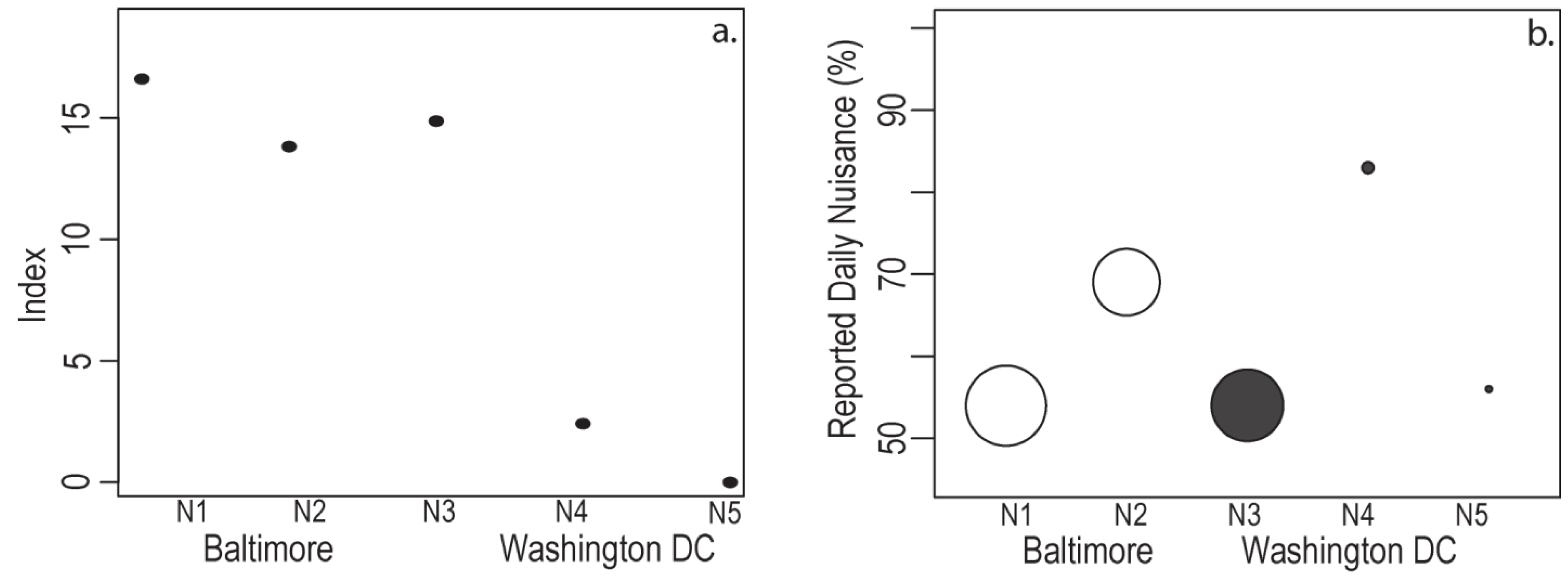
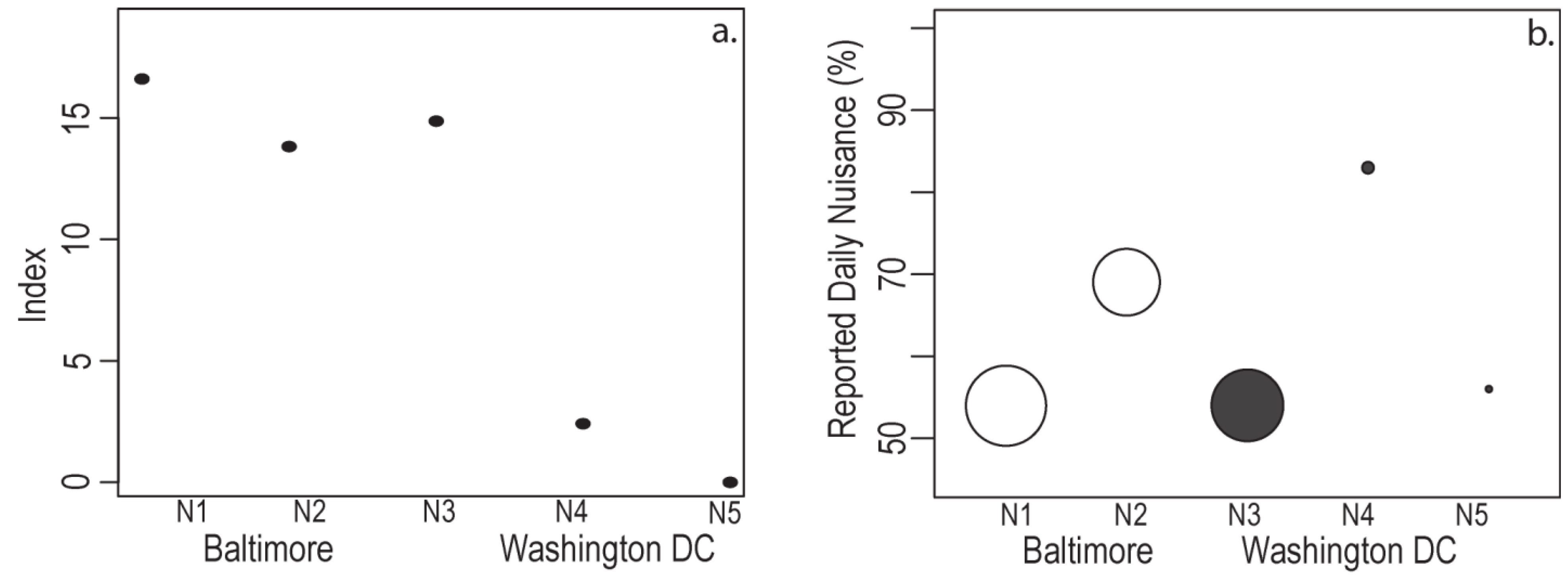
4.4. Resident Response and Relative Exposure
| NBHD | Median Income | Container # | Mosquito + | Pupae # | Every Day | Never |
|---|---|---|---|---|---|---|
| Per Parcel | Per Container | Resident Reported Nuisance | ||||
| N1 | L | 1.49 | 77.0% | 14.96 | 54.0% | 8.0% |
| N2 | M | 1.71 | 40.0% | 21.00 | 69.0% | 6.0% |
| N3 | L | 1.69 | 73.0% | 12.55 | 54.0% | 8.0% |
| N4 | M | 2.45 | 44.0% | 2.74 | 83.0% | 17.0% |
| N5 | H | 2.11 | 21.0% | 1.32 | 56.0% | 8.0% |
5. Conclusions
Acknowledgments
Conflict of Interest
References
- Gubler, D.J. Dengue and dengue hemorrhagic fever. Clin. Microbiol. Rev. 1998, 11, 480–496. [Google Scholar]
- Wilcox, B.A.; Gubler, D.J. Disease ecology and the global emergence of zoonotic pathogens. Environ. Health Prev. Med. 2005, 10, 263–272. [Google Scholar] [CrossRef]
- Gubler, D.J. The global emergence/resurgence of arboviral diseases as public health problems. Arch. Med. Res. 2002, 33, 330–342. [Google Scholar] [CrossRef]
- Weaver, S.C.; Reisen, W.K. Present and future arboviral threats. Antivir. Res. 2010, 85, 328–345. [Google Scholar] [CrossRef]
- Patterson, G. The Mosquito Wars; University Presses of Florida: Gainesville, FL, USA, 2004. [Google Scholar]
- Gubler, D.J. Vector-borne diseases: Understanding the environmental, human health, and ecological connections. In Vector-Borne Disease Emergence and Resurgence; National Academies Press: Washington, DC, USA, 2008; Volume 1. [Google Scholar]
- Woolhouse, M.E.J.; Gowtage-Sequeria, S. Host range and emerging and reemerging pathogens. Emerg. Infect. Dis. 2005, 11, 1842–1847. [Google Scholar] [CrossRef]
- Leisnham, P.T.; Slaney, D.P. Urbanization and the increasing risk from mosquito-borne diseases: Linking human well-being with ecosystem health. In Focus on Urbanization Trends; De Smet, L.M., Ed.; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2009; pp. 47–82. [Google Scholar]
- Gratz, N.G. Emerging and resurging vector-borne diseases. Ann. Rev. Entomol. 1999, 44, 51–75. [Google Scholar] [CrossRef]
- Morens, D.M.; Folkers, G.K.; Fauci, A.S. The challenge of emerging and re-emerging infectious diseases. Nature 2004, 430, 242–249. [Google Scholar] [CrossRef]
- Utzinger, J.; Keiser, J. Urbanization and tropical health—Then and now. Ann. Trop. Med. Parasitol. 2006, 100, 517–533. [Google Scholar] [CrossRef]
- Toussaint, J.F.; Kerkhofs, P.; De Clercq, K. Influence of global climate changes on arboviruses spread. Ann. Med. Vet. 2006, 150, 56–63. [Google Scholar]
- Kumari, R.; Kumar, K.; Chauhan, L.S. First dengue virus detection in Aedes albopictus from Delhi, India: Its breeding ecology and role in dengue transmission. Trop. Med. Int. Health 2011, 16, 949–954. [Google Scholar] [CrossRef]
- Grandadam, M. Arbovirus diseases extension: Who is guilty? M S-Med. Sci. 2011, 27, 339–340. [Google Scholar]
- Leisnham, P.; Juliano, S. Impacts of climate, land use, and biological invasion on the ecology of immature Aedes mosquitoes: Implications for La Crosse emergence. EcoHealth 2012, 9, 217–228. [Google Scholar] [CrossRef]
- LaDeau, S.L.; Calder, C.A.; Doran, P.J.; Marra, P.P. West Nile virus impacts in American crow populations are associated with human land use and climate. Ecol. Res. 2011, 26, 909–916. [Google Scholar] [CrossRef]
- Liu, A.; Lee, V.; Galusha, D.; Slade, M.; Diuk-Wasser, M.; Andreadis, T.; Scotch, M.; Rabinowitz, P. Risk factors for human infection with West Nile virus in Connecticut: A multi-year analysis. Int. J. Health Geogr. 2009, 8. [Google Scholar] [CrossRef]
- Bradley, C.A.; Altizer, S. Urbanization and the ecology of wildlife diseases. Trends Ecol. Evol. 2007, 22, 95–102. [Google Scholar] [CrossRef]
- Gibbs, S.E.J.; Wimberly, M.C.; Madden, M.; Masour, J.; Yabsley, M.J.; Stallknecht, D.E. Factors affecting the geographic distribution of West Nile virus in Georgia, USA: 2002–2004. Vector-Borne Zoonotic. Dis. 2006, 6, 73–82. [Google Scholar] [CrossRef]
- Andreadis, T.G.; Anderson, J.F.; Vossbrinck, C.R.; Main, A.J. Epidemiology of West Nile virus in Connecticut: A five-year analysis of mosquito data 1999–2003. Vector-Borne Zoonotic. Dis. 2004, 4, 360–378. [Google Scholar] [CrossRef]
- Weinberg, M.; Waterman, S.; Lucas, C.A.; Falcon, V.C.; Morales, P.K.; Lopez, L.A.; Peter, C.; Gutierrez, A.E.; Gonzalez, E.R.; Flisser, A.; Bryar, R.; Valle, E.N.; Rodriguez, A.; Hernandez, G.A.; Rosales, C.; Ortiz, J.A.; Lander, M.; Vilchis, H.; Rawlings, J.; Leal, F.L.; Ortega, L.; Flagg, E.; Conyer, R.T.; Cetron, M.; Border Infectious Disease Surveillance Project Working Group. The US-Mexico border infectious disease surveillance project: Establishing binational border surveillance. Emerg. Infect. Dis. 2003, 9, 97–102. [Google Scholar] [CrossRef]
- CDC. Dengue Hemorrhagic Fever—U.S.-Mexico Border, 2005. Available online: http://www.cdc.gov/mmwr/preview/mmwrhtml/mm5631a1.htm (accessed on 13 January 2013).
- CDC. Locally Acquired Dengue—Key West, Florida, 2009–2010. Available online: http://www.cdc.gov/MMWr/preview/mmwrhtml/mm5919a1.htm (accessed on 13 January 2013).
- Morens, D.M.; Fauci, A.S. The 1918 influenza pandemic: Insights for the 21st century. Int. J. Infect. Dis. 2007, 195, 1018–1028. [Google Scholar] [CrossRef]
- Hamer, G.L.; Chaves, L.F.; Anderson, T.K.; Kitron, U.D.; Brawn, J.D.; Ruiz, M.O.; Loss, S.R.; Walker, E.D.; Goldberg, T.L. Fine-Scale variation in vector host use and force of infection drive localized patterns of West Nile virus transmission. PLoS One 2011, 6. [Google Scholar] [CrossRef]
- Lounibos, L.P. Invasions by insect vectors of human disease. Ann. Rev. Entomol. 2002, 47, 233–266. [Google Scholar] [CrossRef]
- Blaustein, L.; Chase, J.M. Interactions between mosquito larvae and species that share the same trophic level. Ann. Rev. Entomol. 2007, 52, 489–507. [Google Scholar] [CrossRef]
- Juliano, S.A. Species interactions among Larval mosquitoes: Context dependence across habitat gradients. Ann. Rev. Entomol. 2009, 54, 37–56. [Google Scholar] [CrossRef]
- Juliano, S.A.; Lounibos, L.P. Ecology of invasive mosquitoes: Effects on resident species and on human health. Ecol. Lett. 2005, 8, 558–574. [Google Scholar] [CrossRef]
- LoGiudice, K.; Ostfeld, R.S.; Schmidt, K.A.; Keesing, F. The ecology of infectious disease: Effects of host diversity and community composition on Lyme disease risk. PNAS 2003, 100, 567–571. [Google Scholar] [CrossRef]
- Apperson, C.S.; Hassan, H.K.; Harrison, B.A.; Savage, H.M.; Aspen, S.E.; Farajollahi, A.; Crans, W.; Daniels, T.J.; Falco, R.C.; Benedict, M.; Anderson, M.; McMillen, L.; Unnasch, T.R. Host feeding patterns of established and potential mosquito vectors of West Nile virus in the eastern United States. Vector-Borne Zoonotic. Dis. 2004, 4, 71–82. [Google Scholar] [CrossRef]
- Reisen, W.K.; Fang, Y.; Martinez, V.M. Avian host and mosquito (Diptera: Culicidae) vector competence determine the efficiency of west nile and St. Louis encephalitis virus transmission. J. Med. Entomol. 2005, 42, 367–375. [Google Scholar] [CrossRef]
- Kilpatrick, A.M.; Daszak, P.; Jones, M.J.; Marra, P.P.; Kramer, L.D. Host heterogeneity dominates West Nile virus transmission. Proc. R. Soc. B Biol. Sci. 2006, 273, 2327–2333. [Google Scholar] [CrossRef]
- Kilpatrick, A.M.; Kramer, L.D.; Jones, M.J.; Marra, P.P.; Daszak, P. West Nile virus epidemics in North America are driven by shifts in mosquito feeding behavior. PLoS Biol. 2006, 4, 606–610. [Google Scholar]
- Chase, J.M.; Knight, T.M. Drought-induced mosquito outbreaks in wetlands. Ecol. Lett. 2003, 6, 1017–1024. [Google Scholar] [CrossRef]
- Blaustein, L.; Kiflawi, M.; Eitam, A.; Mangel, M.; Cohen, J.E. Oviposition habitat selection in response to risk of predation in temporary pools: Mode of detection and consistency across experimental venue. Oecologia 2004, 138, 300–305. [Google Scholar] [CrossRef]
- Juliano, S.A.; Lounibos, L.P.; Nishimura, N.; Greene, K. Your worst enemy could be your best friend: Predator contributions to invasion resistance and persistence of natives. Oecologia 2010, 162, 709–718. [Google Scholar] [CrossRef]
- Juliano, S.A. Population dynamics. J. Amer. Mosquito Contr. Assn. 2007, 23, 265–275. [Google Scholar] [CrossRef]
- Yee, D.A.; Kaufman, M.G.; Juliano, S.A. The significance of ratios of detritus types and micro-organism productivity to competitive interactions between aquatic insect detritivores. J. Anim. Ecol. 2007, 76, 1105–1115. [Google Scholar] [CrossRef]
- Kaufman, M.G.; Pelz-Stelinski, K.S.; Yee, D.A.; Juliano, S.A.; Ostrom, P.H.; Walker, E.D. Stable isotope analysis reveals detrital resource base sources of the tree hole mosquito, Aedes triseriatus. Ecol. Entomol. 2010, 35, 586–593. [Google Scholar] [CrossRef]
- Kraus, J.M.; Vonesh, J.R. Fluxes of terrestrial and aquatic carbon by emergent mosquitoes: A test of controls and implications for cross-ecosystem linkages. Oecologia 2012, 170, 1111–1122. [Google Scholar] [CrossRef]
- Leisnham, P.T.; Sala, L.M.; Juliano, S.A. Geographic variation in adult survival and reproductive tactics of the mosquito Aedes albopictus. J. Med. Entomol. 2008, 45, 210–221. [Google Scholar] [CrossRef]
- AMCA. American Mosquito Control Association. Available online: http://www.mosquito.org/faq#few (accessed on 13 January 2013).
- Farajollahi, A.; Fonseca, D.M.; Kramer, L.D.; Kilpatrick, A.M. “Bird biting” mosquitoes and human disease: A review of the role of Culex pipiens complex mosquitoes in epidemiology. Infect. Genet. Evol. 2011, 11, 1577–1585. [Google Scholar] [CrossRef]
- Kilpatrick, A.M.; Kramer, L.D.; Jones, M.J.; Marra, P.P.; Daszak, P.; Fonseca, D.M. Genetic influences on mosquito feeding behavior and the emergence of zoonotic pathogens. Amer. J. Trop. Med. Hyg. 2007, 77, 667–671. [Google Scholar]
- Hamer, G.L.; Kitron, U.D.; Brawn, J.D.; Loss, S.R.; Ruiz, M.O.; Goldberg, T.L.; Walker, E.D. Culex pipiens (Diptera: Culicidae): A bridge vector of West Nile virus to humans. J. Med. Entomol. 2008, 45, 125–128. [Google Scholar] [CrossRef]
- Molaei, G.; Andreadis, T.G.; Armstrong, P.M.; Bueno, R.; Dennett, J.A.; Real, S.V.; Sargent, C.; Bala, A.; Randle, Y.; Guzman, H.; da Rosa, A.T.; Wuithiranyagool, T.; Tesh, R.B. Host feeding pattern of Culex quinquefasciatus (Diptera: Culicidae) and its role in transmission of West Nile virus in Harris County, Texas. Amer. J. Trop. Med. Hyg. 2007, 77, 73–81. [Google Scholar]
- Molaei, G.; Andreadis, T.A.; Armstrong, P.M.; Anderson, J.F.; Vossbrinck, C.R. Host feeding patterns of Culex mosquitoes and West Nile virus transmission, northeastern United States. Emerg. Infect. Dis. 2006, 12, 468–474. [Google Scholar] [CrossRef]
- Apperson, C.S.; Harrison, B.A.; Unnasch, T.R.; Hassan, H.K.; Irby, W.S.; Savage, H.M.; Aspen, S.E.; Watson, D.W.; Rueda, L.M.; Engber, B.R.; Nasci, R.S. Host-feeding habits of Culex and other mosquitoes (Diptera: Culicidae) in the Borough of Queens in New York City, with characters and techniques for identification of Culex mosquitoes. J. Med. Entomol. 2002, 39, 777–785. [Google Scholar] [CrossRef]
- Su, T.Y.; Mulla, M.S. Effects of temperature on development, mortality, mating and blood feeding behavior of Culiseta incidens (Diptera : Culicidae). J. Vector Ecol. 2001, 26, 83–92. [Google Scholar]
- Reisen, W.K.; Cayan, D.; Tyree, M.; Barker, C.A.; Eldridge, B.; Dettinger, M. Impact of climate variation on mosquito abundance in California. J. Vector Ecol. 2008, 33, 89–98. [Google Scholar] [CrossRef]
- Paz, S.; Albersheim, I. Influence of warming tendency on Culex pipiens population abundance and on the probability of West Nile Fever outbreaks (Israeli case study: 2001–2005). EcoHealth 2008, 5, 40–48. [Google Scholar] [CrossRef]
- Delatte, H.; Gimonneau, G.; Triboire, A.; Fontenille, D. Influence of temperature on immature development, survival, longevity, fecundity, and gonotrophic cycles of Aedes albopictus, vector of Chikungunya and Dengue in the Indian ocean. J. Med. Entomol. 2009, 46, 33–41. [Google Scholar] [CrossRef]
- Leisnham, P.T.; Towler, L.; Juliano, S.A. Geographic variation of photoperiodic diapause but not adult survival or reproduction of the invasive mosquito Aedes albopictus (Diptera: Culicidae) in North America. Ann. Entomol. Soc. Amer. 2011, 104, 1309–1318. [Google Scholar] [CrossRef]
- Patz, J.A.; Daszak, P.; Tabor, G.M.; Aguirre, A.A.; Pearl, M.; Epstein, J.; Wolfe, N.D.; Kilpatrick, A.M.; Foufopoulos, J.; Molyneux, D.; Bradley, D.J. Working Group on Land Use Change and Disease Emergence. Unhealthy landscapes: Policy recommendations on land use change and infectious disease emergence. Environ. Health Perspect. 2004, 112, 1092–1098. [Google Scholar] [CrossRef]
- Patz, J.A.; Olson, S.H.; Uejio, C.K.; Gibbs, H.K. Disease emergence from global climate and land use change. Med. Clin. N Amer. 2008, 92, 1473–1491. [Google Scholar] [CrossRef]
- Yasuoka, J.; Levins, R. Impact of deforestation and agricultural development on anopheline ecology and malaria epidemiology. Amer. J. Trop. Med. Hyg. 2007, 76, 450–460. [Google Scholar]
- Myers, S.S.; Patz, J.A. Emerging threats to human health from global environmental change. Annu. Rev. Env. Resour. 2009, 34, 223–252. [Google Scholar] [CrossRef]
- Anderson, R.M.; May, R.M. Infectious Diseases of Humans: Dynamics and Control; Oxford University Press: Oxford, UK, 1992. [Google Scholar]
- Ruiz, M.O.; Walker, E.D.; Foster, E.S.; Haramis, L.D.; Kitron, U.D. Association of West Nile virus illness and urban landscapes in Chicago and Detroit. Int. J. Health Geogr. 2007, 6. [Google Scholar] [CrossRef]
- Harrigan, R.J.; Thomassen, H.A.; Buermann, W.; Cummings, R.F.; Kahn, M.E.; Smith, T.B. Economic conditions predict prevalence of West Nile Virus. PLoS One 2010, 5. [Google Scholar] [CrossRef]
- Dowling, Z.; LaDeau, S.; Armbruster, P.; Biehler, D.; Leisnham, P. Socioeconomic status affects mosquito (Diptera: Culicidae) larval habitat types and infestation. J. Med. Entomol. 2013. in review. [Google Scholar]
- Ruiz, M.O.; Chaves, L.F.; Hamer, G.L.; Sun, T.; Brown, W.M.; Walker, E.D.; Haramis, L.; Goldberg, T.L.; Kitron, U.D. Local impact of temperature and precipitation on West Nile virus infection in Culex species mosquitoes in northeast Illinois, USA. Parasit. Vectors 2010, 3. [Google Scholar] [CrossRef]
- Tedesco, C.; Ruiz, M.; McLafferty, S. Mosquito politics: Local vector control policies and the spread of West Nile Virus in the Chicago region. Health Place 2010, 16, 1188–1195. [Google Scholar] [CrossRef]
- Mayer, J.D. The political ecology of disease as one new focus for medical geography. Prog. Hum. Geogr. 1996, 20, 441–456. [Google Scholar] [CrossRef]
- Robbins, P.; Farnsworth, R.; Jones, J.P. Insects and institutions: Managing emergent hazards in the US southwest. J. Environ. Policy Plann. 2008, 10, 95–112. [Google Scholar] [CrossRef]
- Grimm, N.B.; Foster, D.; Groffman, P.; Grove, J.M.; Hopkinson, C.S.; Nadelhoffer, K.J.; Pataki, D.E.; Peters, D.P.C. The changing landscape: Ecosystem responses to urbanization and pollution across climatic and societal gradients. Front. Ecol. Environ. 2008, 6, 264–272. [Google Scholar] [CrossRef]
- Pickett, S.T.A.; Buckley, G.L.; Kaushal, S.S.; Williams, Y. Social-ecological science in the humane metropolis. Urban Ecosyst. 2011, 14, 319–339. [Google Scholar] [CrossRef]
- Pataki, D.E.; Carreiro, M.M.; Cherrier, J.; Grulke, N.E.; Jennings, V.; Pincetl, S.; Pouyat, R.V.; Whitlow, T.H.; Zipperer, W.C. Coupling biogeochemical cycles in urban environments: ecosystem services, green solutions, and misconceptions. Front. Ecol. Environ. 2011, 9, 27–36. [Google Scholar] [CrossRef]
- Khormi, H.M.; Kumar, L. Assessing the risk for dengue fever based on socioeconomic and environmental variables in a geographical information system environment. Geospat. Health 2012, 6, 171–176. [Google Scholar]
- Ruhago, G.M.; Mujinja, P.G.M.; Norheim, O.F. Equity implications of coverage and use of insecticide treated nets distributed for free or with co-payment in two districts in Tanzania: A cross-sectional comparative household survey. Int. J. Equity Health 2011, 10. [Google Scholar] [CrossRef]
- Furst, T.; Raso, G.; Acka, C.A.; Tschannen, A.B.; N’Goran, E.K.; Utzinger, J. Dynamics of socioeconomic risk factors for neglected tropical diseases and Malaria in an armed conflict. PLoS Negl. Trop. Dis. 2009, 3. [Google Scholar] [CrossRef]
- Honorio, N.A.; Nogueira, R.M.R.; Codeco, C.T.; Carvalho, M.S.; Cruz, O.G.; Magalhaes, M.; de Araujo, J.M.G.; de Araujo, E.S.M.; Gomes, M.Q.; Pinheiro, L.S.; Pinel, C.D.; Lourenco-de-Oliveira, R. Spatial evaluation and modeling of Dengue seroprevalence and vector density in Rio de Janeiro, Brazil. PLoS Negl. Trop. Dis. 2009, 3. [Google Scholar] [CrossRef]
- Nguyen, L.A.P.; Clements, A.C.A.; Jeffery, J.A.L.; Yen, N.T.; Nam, V.S.; Vaughan, G.; Shinkfield, R.; Kutcher, S.C.; Gatton, M.L.; Kay, B.H.; Ryan, P.A. Abundance and prevalence of Aedes aegypti immatures and relationships with household water storage in rural areas in southern Viet Nam. Int. Health 2011, 3, 115–125. [Google Scholar] [CrossRef]
- Reisen, W.K.; Meyer, R.P.; Tempelis, C.H.; Spoehel, J.J. Mosquito abundance and bionomics in residential communities in Orange and Los-Angeles counties, California. J. Med. Entomol. 1990, 27, 356–367. [Google Scholar]
- Rochlin, I.; Turbow, D.; Gomez, F.; Ninivaggi, D.V.; Campbell, S.R. Predictive mapping of human risk for West Nile Virus (WNV) based on environmental and socioeconomic factors. PLoS One 2011, 6. [Google Scholar] [CrossRef]
- Reisen, W.K.; Takahashi, R.M.; Carroll, B.D.; Quiring, R. Delinquent mortgages, neglected swimming pools, and West Nile Virus, California. Emerg. Infect. Dis. 2008, 14, 1747–1749. [Google Scholar] [CrossRef]
- Dowling, Z.; Armbruster, P.; LaDeau, S.; DeCotis, M.; Motley, J.; Leisnham, P. Linking mosquito infestation to resident socioeconomic status, knowledge and source reduction practices in suburban Washington, D.C. EcoHealth 2013. [Google Scholar] [CrossRef]
- Bartlett-Healy, K.; Unlu, I.; Obenauer, P.; Hughes, T.; Healy, S.; Crepeau, T.; Farajollahi, A.; Kesavaraju, B.; Fonseca, D.; Schoeler, G.; Gaugler, R.; Strickman, D. Larval mosquito habitat utilization and community dynamics of Aedes albopictus and Aedes japonicus (Diptera: Culicidae). J. Med. Entomol. 2012, 49, 813–824. [Google Scholar] [CrossRef]
- Bartlett-Healy, K.; Hamilton, G.; Healy, S.; Crepeau, T.; Unlu, I.; Farajollahi, A.; Fonseca, D.; Gaugler, R.; Clark, G.G.; Strickman, D. Source reduction behavior as an independent measurement of the impact of a public health education campaign in an integrated vector management program for the Asian tiger mosquito. Int. J. Environ. Res. Public Health 2011, 8, 1358–1367. [Google Scholar] [CrossRef]
- Alto, B.W.; Juliano, S.A. Precipitation and temperature effects on populations of Aedes albopictus (Diptera : Culicidae): Implications for range expansion. J. Med. Entomol. 2001, 38, 646–656. [Google Scholar] [CrossRef]
- Strickman, D.; Kittayapong, P. Dengue and its vectors in Thailand: Calculated transmission risk from total pupal counts of Aedes aegypti and association of wing-length measurements with aspects of the larval habitat. Amer. J. Trop. Med. Hyg. 2003, 68, 209–217. [Google Scholar]
- Harlan, N.P.; Paradise, C.J. Do habitat size and shape modify abiotic factors and communities in artificial treeholes? Community Ecol. 2006, 7, 211–222. [Google Scholar] [CrossRef]
- Lounibos, L.P.; Nishimura, N.; Escher, R.L. Fitness of a treehole mosquito—Influences of food type and predation. Oikos 1993, 66, 114–118. [Google Scholar] [CrossRef]
- Vezzani, D.; Albicocco, A.P. The effect of shade on the container index and pupal productivity of the mosquitoes Aedes aegypti and Culex pipiens breeding in artificial containers. Med. Vet. Entomol. 2009, 23, 78–84. [Google Scholar] [CrossRef]
- Bartlett-Healy, K.; Healy, S.P.; Hamilton, G.C. A model to predict evaporation rates in habitats used by container-dwelling mosquitoes. J. Med. Entomol. 2011, 48, 712–716. [Google Scholar] [CrossRef]
- Metzger, M.E.; Myers, C.M.; Kluh, S.; Wekesa, J.W.; Hu, R.J.; Kramer, V.L. An assessment of mosquito production and nonchemical control measures in structural stormwater best management practices in Southern California. J. Amer. Mosquito Contr. Assn. 2008, 24, 70–81. [Google Scholar] [CrossRef]
- Kwan, J.A.; Riggs-Nagy, J.M.; Fritz, C.L.; Shindelbower, M.; Castro, P.A.; Kramer, V.L.; Metzger, M.E. Mosquito production in stormwater treatment devices in the Lake Tahoe Basin, California. J. Amer. Mosquito Contr. Assn 2008, 24, 82–89. [Google Scholar] [CrossRef]
- Gardner, A.M.; Hamer, G.L.; Hines, A.M.; Newman, C.M.; Walker, E.D.; Ruiz, M.O. Weather variability affects abundance of Larval Culex (Diptera: Culicidae) in storm water catch basins in suburban Chicago. J. Med. Entomol. 2012, 49, 270–276. [Google Scholar] [CrossRef]
- Chaves, L.F.; Keogh, C.L.; Nguyen, A.M.; Decker, G.M.; Vazquez-Prokopec, G.M.; Kitron, U.D. Combined sewage overflow accelerates immature development and increases body size in the urban mosquito Culex quinquefasciatus. J. Appl. Entomol. 2011, 135, 611–620. [Google Scholar] [CrossRef]
- Couzin-Frankel, J. Fears of lax surveillance if CDC program cut. Science 2010, 328, 1088. [Google Scholar] [CrossRef]
- LaBeaud, A.D.; Aksoy, S. Neglected funding for vector-borne diseases: A near miss this time, a possible disaster next time. PLoS Negl. Trop. Dis. 2010, 4. [Google Scholar] [CrossRef]
- World Health Organization, Dengue Haemorrhagic Fever: Diagnosis, Treatment, Prevention and Control; World Health Organization: Geneva, Switzerland, 1997.
- Tuiten, W.; Koenraadt, C.J.M.; McComas, K.; Harrington, L.C. The effect of West Nile Virus perceptions and knowledge on protective behavior and mosquito breeding in residential yards in upstate New York. EcoHealth 2009, 6, 42–51. [Google Scholar] [CrossRef]
- Biehler, D.D. Flies, Bedbugs, Roaches, and Rats: A History of Pests in the City; University of Washington Press: Seattle, WA, USA, 2013. [Google Scholar]
- US Census Bureau, American Community Survey. Available online: https://www.census.gov/acs/www/ (accessed on 15 January 2013).
- Baltimore City Office of Sustainability. Cleanliness: Vacant Lots. Available online: http://www.baltimoresustainability.org/cleanliness/vacant-lots (accessed on 19 January 2013).
- Hille, M. Animal disease control. DC Department of Health; Personal comment to Leisnham, P. 2010. [Google Scholar]
- Westbrook, C.J.; Reiskind, M.H.; Pesko, K.N.; Greene, K.E.; Lounibos, L.P. Larval environmental temperature and the susceptibility of Aedes albopictus Skuse (Diptera: Culicidae) to Chikungunya virus. Vector-Borne Zoonotic. Dis. 2010, 10, 241–247. [Google Scholar] [CrossRef]
- Turell, M.J.; Dohm, D.J.; Sardelis, M.R.; Oquinn, M.L.; Andreadis, T.G.; Blow, J.A. An update on the potential of North American mosquitoes (Diptera: Culicidae) to transmit West Nile virus. J. Med. Entomol. 2005, 42, 57–62. [Google Scholar] [CrossRef]
- Sardelis, M.R.; Turell, M.J.; O’Guinn, M.L.; Andre, R.G.; Roberts, D.R. Vector competence of three North American strains of Aedes albopictus for West Nile virus. J. Amer. Mosquito Contr. Assn. 2002, 18, 284–289. [Google Scholar]
- Sardelis, M.R.; Turell, M.J.; Dohm, D.J.; O’Guinn, M.L. Vector competence of selected North American Culex and Coquillettidia mosquitoes for West Nile virus. Emerg. Infect. Dis. 2001, 7, 1018–1022. [Google Scholar] [CrossRef]
- McTighe, S.P.; Vaidyanathan, R. Vector competence of Aedes albopictus from Virginia and Georgia for Chikungunya virus isolated in the Comoros Islands, 2005. Vector-Borne Zoonotic. Dis. 2012, 12, 867–871. [Google Scholar] [CrossRef]
- Lambrechts, L.; Scott, T.W.; Gubler, D.J. Consequences of the expanding global distribution of Aedes albopictus for Dengue Virus Transmission. PLoS Negl. Trop. Dis. 2010, 4. [Google Scholar] [CrossRef]
- Gratz, N.G. Critical review of the vector status of Aedes albopictus. Med. Vet. Entomol. 2004, 18, 215–227. [Google Scholar] [CrossRef]
- Crans, W.J.; Chomsky, M.S.; Guthrie, D.; Acquaviva, A. First record of Aedes albopictus from New Jersey. J. Amer. Mosquito Contr. Assn. 1996, 12, 307–309. [Google Scholar]
- Hawley, W.A. The biology of Aedes albopictus. J. Amer. Mosquito Contr. Assn. 1988, 434, 1–40. [Google Scholar]
- Leisnham, P.T.; Lounibos, L.P.; O'Meara, G.F.; Juliano, S.A. Interpopulation divergence in competitive interactions of the mosquito Aedes albopictus. Ecology 2009, 90, 2405–2413. [Google Scholar] [CrossRef]
- Kilpatrick, A.M.; Kramer, L.D.; Campbell, S.R.; Alleyne, E.O.; Dobson, A.P.; Daszak, P. West Nile virus risk assessment and the bridge vector paradigm. Emerg. Infect. Dis. 2005, 11, 425–429. [Google Scholar] [CrossRef]
- Peyton, E.L.; Campbell, S.R.; Candeletti, T.M.; Romanowski, M.; Crans, W.J. Aedes (Finlaya) japonicus japonicus (Theobald), a new introduction into the United States. J. Amer. Mosquito Contr. Assn. 1999, 15, 238–241. [Google Scholar]
- Andreadis, T.G.; Wolfe, B.J. Evidence for reduction of native mosquitoes with increased expansion of invasive Ochlerotatus japonicus japonicus (Diptera: Culicidae) in the Northeastern United States. J. Med. Entomol. 2010, 47, 43–52. [Google Scholar]
- Bevins, S.N. Establishment and abundance of a recently introduced mosquito species Ochlerotatus japonicus (Diptera: Culicidae) in the Southern Appalachians, USA. J. Med. Entomol. 2007, 44, 945–952. [Google Scholar] [CrossRef]
- Fonseca, D.M.; Widdel, A.K.; Hutchinson, M.; Spichiger, S.E.; Kramer, L.D. Fine-scale spatial and temporal population genetics of Aedes japonicus, a new US mosquito, reveal multiple introductions. Mol. Ecol. 2010, 19, 1559–1572. [Google Scholar] [CrossRef]
- Sardelis, M.R.; Turell, M.J.; Andre, A.R.G. Laboratory transmission of La Crosse virus by Ochlerotatus j. japonicus (Diptera: Culicidae). J. Med. Entomol. 2002, 39, 635–639. [Google Scholar] [CrossRef]
- Sardelis, M.R.; Turell, M.J. Ochlerotatus j. japonicus in Frederick County, Maryland: Discovery, distribution, and vector competence for West Nile virus. J. Amer. Mosquito Contr. Assn. 2001, 17, 137–141. [Google Scholar]
- Sardelis, M.R.; Dohm, D.J.; Pagac, B.; Andre, R.G.; Turell, M.J. Experimental transmission of eastern equine encephalitis virus by Ochlerotatus j. japonicus (Diptera: Culicidae). J. Med. Entomol. 2002, 39, 480–484. [Google Scholar] [CrossRef]
- Sardelis, M.R.; Turell, M.J.; Andre, R.G. Experimental transmission of St. Louis encephalitis virus by Ochlerotatus j. japonicus. J. Amer. Mosquito Contr. Assn. 2003, 19, 159–162. [Google Scholar]
- Darsie, R.; Ward, R. Identification and Geographical Distribution of the Mosquitoes of North America, North of Mexico; University of Florida Press: Gainesville, FL, USA, 2005. [Google Scholar]
- Gelman, A.; Hill, J. Data Analysis Using Regression and Multilevel/Hierarchical Models; Cambridge University Press: New York, NY, USA, 2007; pp. 325–342. [Google Scholar]
- Yee, D.A. Tires as habitats for mosquitoes: A review of studies within the eastern United States. J. Med. Entomol. 2008, 45, 581–593. [Google Scholar] [CrossRef]
- Kaufman, P.E.; Harrington, L.C.; Waldron, J.K.; Rutz, D.A. The importance of agricultural tire habitats for mosquitoes of public health importance in New York state. J. Amer. Mosquito Contr. Assn. 2005, 21, 171–176. [Google Scholar] [CrossRef]
- Focks, D.A.; Chadee, D.D. Pupal survey: An epidemiologically significant surveillance method for Aedes aegypti: An example using data from Trinidad. Amer. J. Trop. Med. Hyg. 1997, 56, 159–167. [Google Scholar]
- Focks, D.A.; Sackett, S.R.; Bailey, D.L.; Dame, D.A. Observations on container-breeding mosquitoes in New Orleans, Louisiana, with an estimate of the population-density of Aedes aegypti (L). Amer. J. Trop. Med. Hyg. 1981, 30, 1329–1335. [Google Scholar]
- Kitron, U.; Clennon, J.A.; Cecere, M.C.; Gurtler, R.E.; King, C.H.; Vazquez-Prokopec, G. Upscale or downscale: Applications of fine scale remotely sensed data to Chagas disease in Argentina and schistosomiasis in Kenya. Geospatial Health 2006, 1, 49–58. [Google Scholar]
- Reiter, P.; Lathrop, S.; Bunning, M.; Biggerstaff, B.; Singer, D.; Tiwari, T.; Baber, L.; Amador, M.; Thirion, J.; Hayes, J.; Seca, C.; Mendez, J.; Ramirez, B.; Robinson, J.; Rawlings, J.; Vorndam, V.; Waterman, S.; Gubler, D.; Clark, G.; Hayes, E. Texas lifestyle limits transmission of dengue virus. Emerg. Infect. Dis. 2003, 9, 86–89. [Google Scholar] [CrossRef]
- Ruiz, M.O.; Brown, W.M.; Brawn, J.D.; Hamer, G.L.; Kunkel, K.E.; Loss, S.R.; Walker, E.D.; Kitron, U.D. Spatiotemporal patterns of precipitation and West Nile virus in Chicago, Illinois, 2002–2005 and implications for surveillance. Amer. J. Trop. Med. Hyg. 2006, 75, 269–270. [Google Scholar]
- Hammond, S.N.; Gordon, A.L.; Lugo, E.D.C.; Moreno, G.; Kuan, G.M.; Lopez, M.M.; Lopez, J.D.; Delgado, M.A.; Valle, S.I.; Espinoza, P.M.; Harris, E. Characterization of Aedes aegypti (Diptera: Culcidae) production sites in urban Nicaragua. J. Med. Entomol. 2007, 44, 851–860. [Google Scholar] [CrossRef]
- Luz, P.M.; Vanni, T.; Medlock, J.; Paltiel, A.D.; Galvani, A.P. Dengue vector control strategies in an urban setting: An economic modelling assessment. Lancet 2011, 377, 1673–1680. [Google Scholar] [CrossRef]
- Miller, J.R. The control of mosquito-borne diseases in New York City. J. Urban Health 2001, 78, 359–366. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
LaDeau, S.L.; Leisnham, P.T.; Biehler, D.; Bodner, D. Higher Mosquito Production in Low-Income Neighborhoods of Baltimore and Washington, DC: Understanding Ecological Drivers and Mosquito-Borne Disease Risk in Temperate Cities. Int. J. Environ. Res. Public Health 2013, 10, 1505-1526. https://doi.org/10.3390/ijerph10041505
LaDeau SL, Leisnham PT, Biehler D, Bodner D. Higher Mosquito Production in Low-Income Neighborhoods of Baltimore and Washington, DC: Understanding Ecological Drivers and Mosquito-Borne Disease Risk in Temperate Cities. International Journal of Environmental Research and Public Health. 2013; 10(4):1505-1526. https://doi.org/10.3390/ijerph10041505
Chicago/Turabian StyleLaDeau, Shannon L., Paul T. Leisnham, Dawn Biehler, and Danielle Bodner. 2013. "Higher Mosquito Production in Low-Income Neighborhoods of Baltimore and Washington, DC: Understanding Ecological Drivers and Mosquito-Borne Disease Risk in Temperate Cities" International Journal of Environmental Research and Public Health 10, no. 4: 1505-1526. https://doi.org/10.3390/ijerph10041505
APA StyleLaDeau, S. L., Leisnham, P. T., Biehler, D., & Bodner, D. (2013). Higher Mosquito Production in Low-Income Neighborhoods of Baltimore and Washington, DC: Understanding Ecological Drivers and Mosquito-Borne Disease Risk in Temperate Cities. International Journal of Environmental Research and Public Health, 10(4), 1505-1526. https://doi.org/10.3390/ijerph10041505