Biodegradable and Biocompatible Biomaterial, Polyhydroxybutyrate, Produced by an Indigenous Vibrio sp. BM-1 Isolated from Marine Environment


Abstract
:1. Introduction
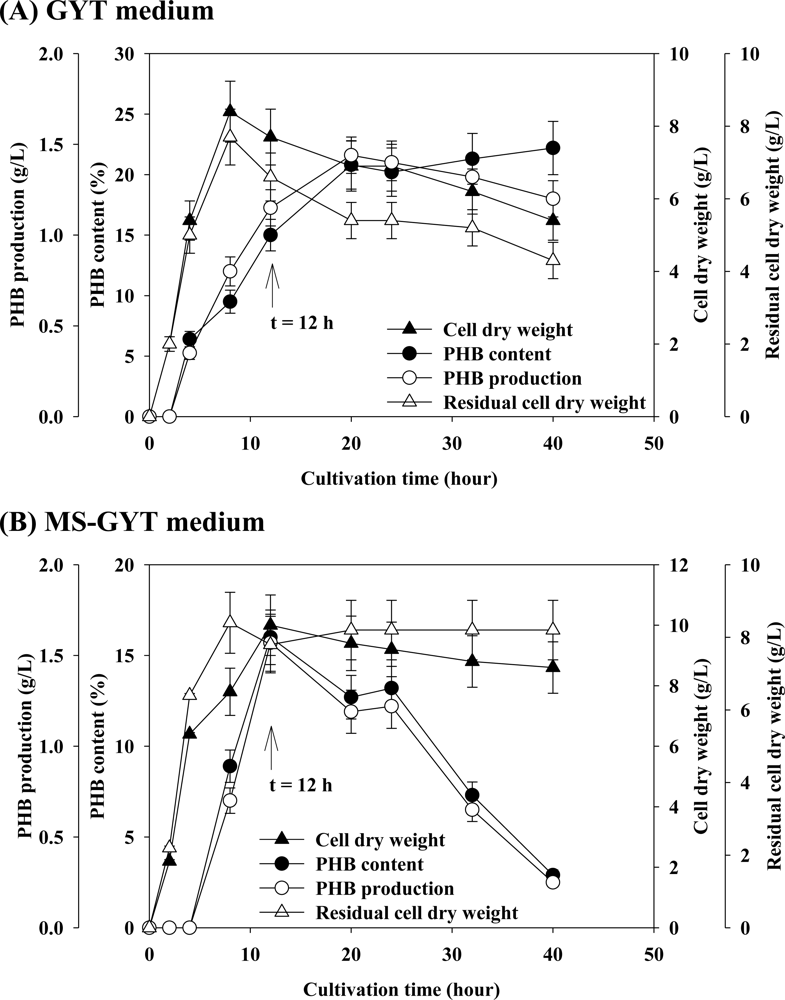
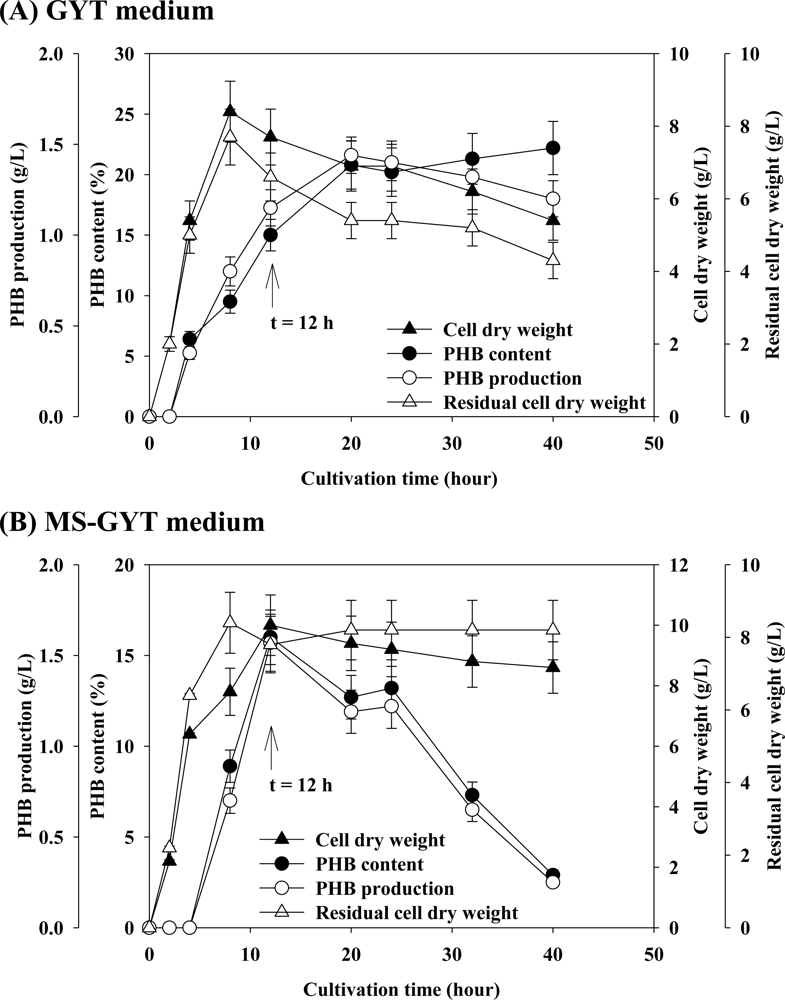
2. Results and Discussions
2.1. Effect of Carbon Source on PHB Production
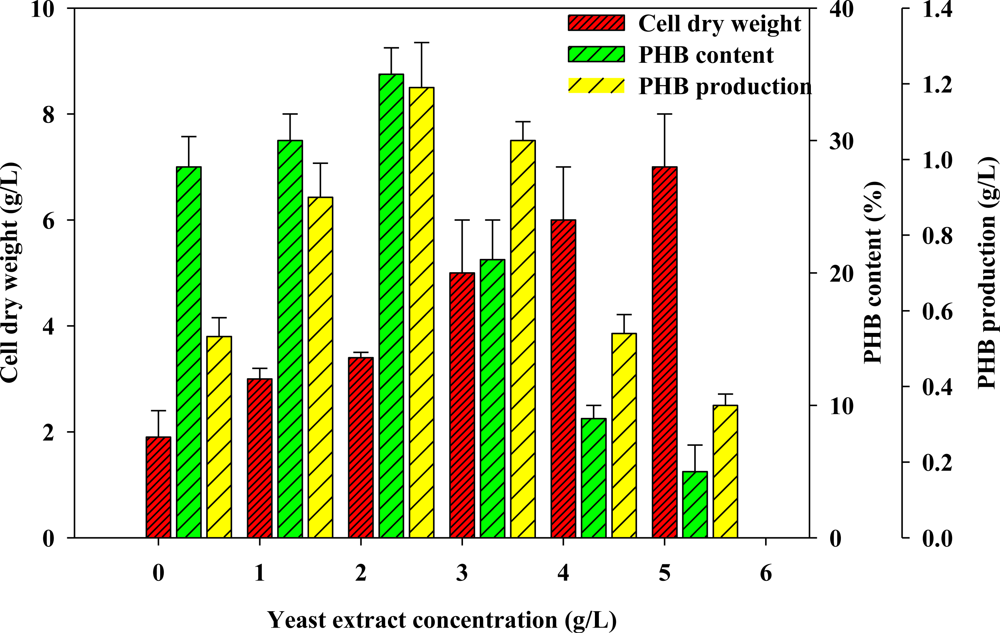
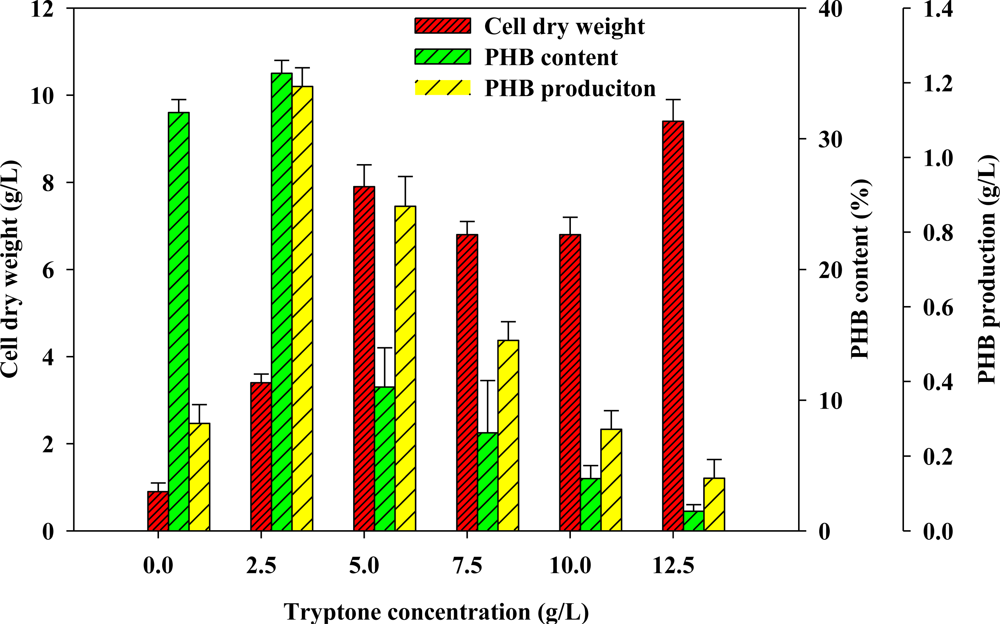
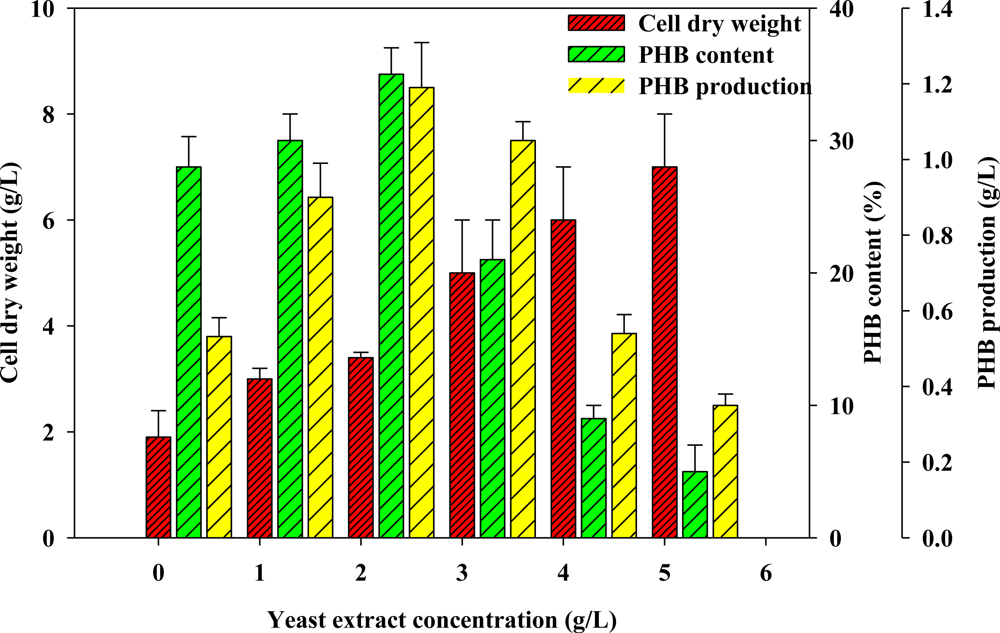
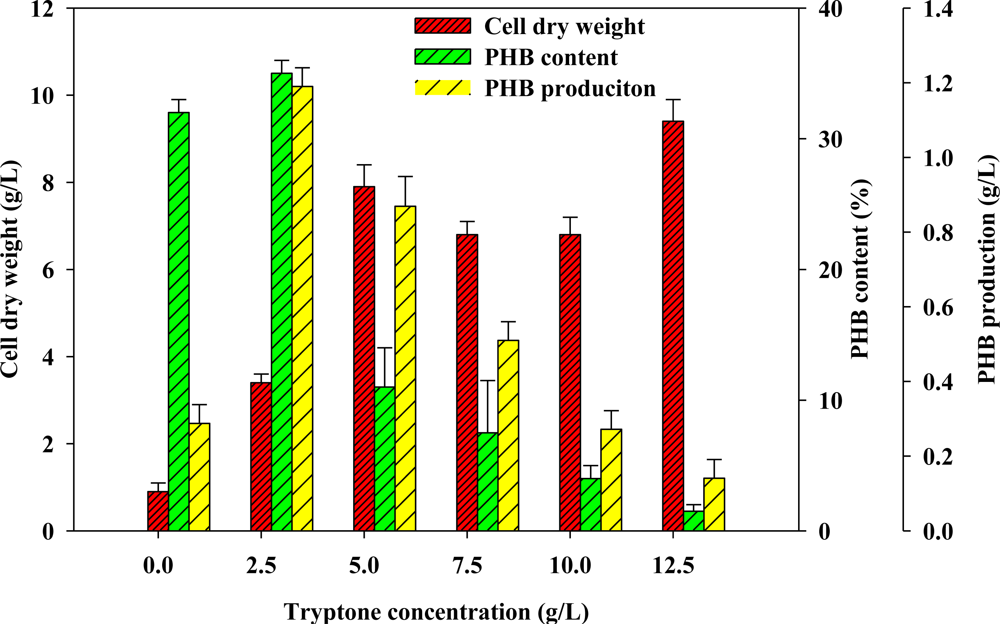
2.2. Effect of Various Concentrations of Complex Nitrogen Sources on PHB Production
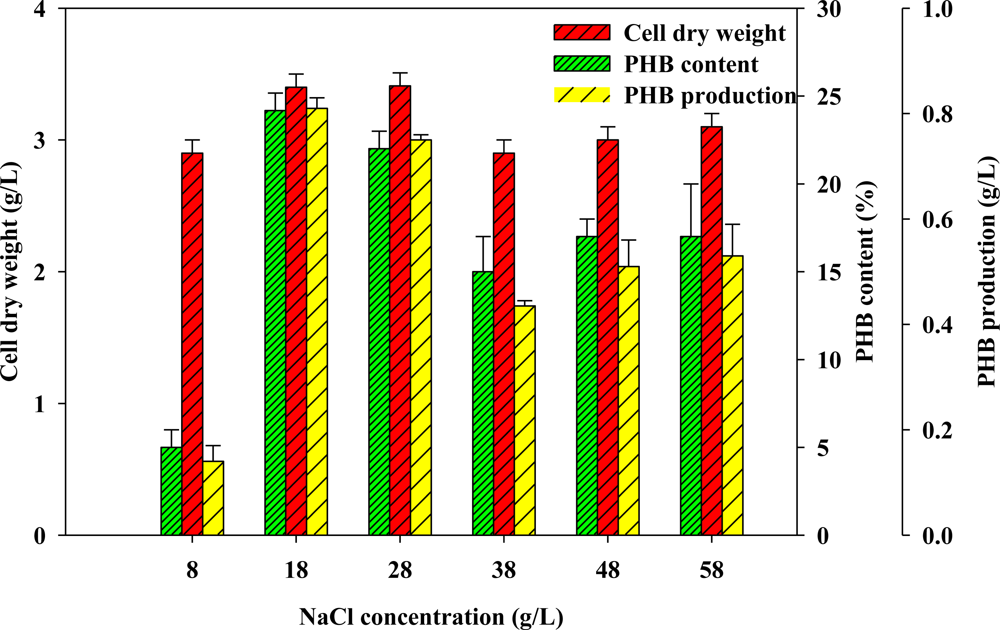
2.3. Effect of Concentration of Sodium Chloride on PHB Production
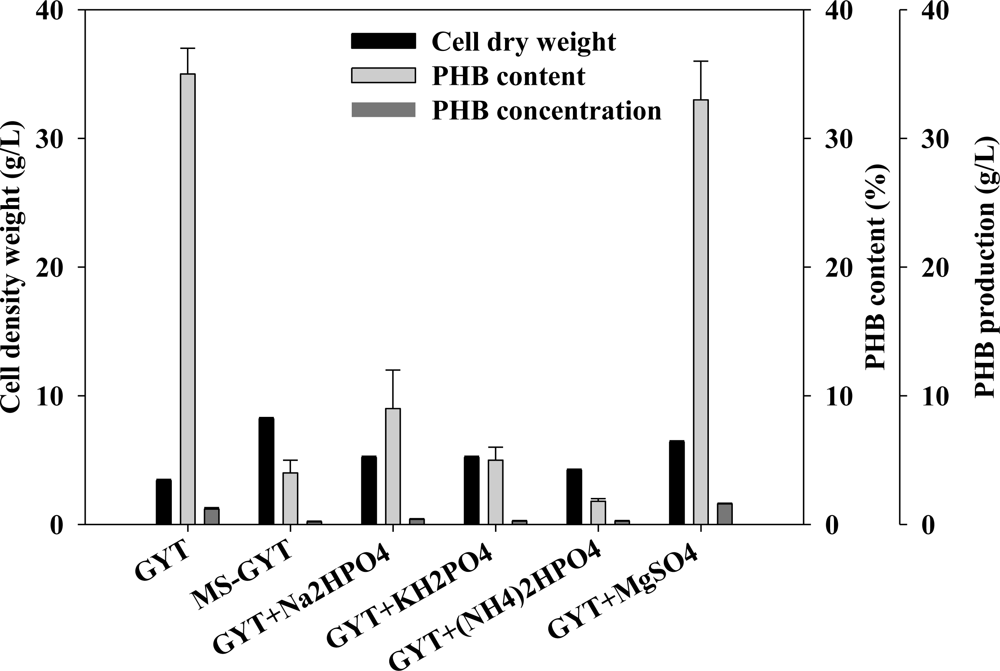
2.4. Effect of Mineral Salts on PHB Production
3. Experimental Section
3.1. Microorganism and Culture Medium
3.2. Culture Condition
3.3. Quantifications of Cell Growth and PHB
3.4. Analytical Methods
4. Conclusions
Acknowledgments
- Samples Availability: Available from the authors.
References
- Keshavarz, T; Roy, I. Polyhydroxyalkanoates: Bioplastics with a green agenda. Curr Opin Microbiol 2010, 13, 321–326. [Google Scholar]
- Reddy, CS; Ghai, R; Rashmi Kalia, VC. Polyhydroxyalkanoates: An overview. Bioresour Technol 2003, 87, 137–146. [Google Scholar]
- Brandl, H; Gross, RA; Lenz, RW; Fuller, RC. Plastics from bacteria and for bacteria: poly(beta-hydroxyalkanoates) as natural, biocompatible, and biodegradable polyesters. Adv Biochem Eng Biotechnol 1990, 41, 77–93. [Google Scholar]
- Khanna, S; Srivastava, AK. Recent advances in microbial polyhydroxyalkanoates. Proc Biochem 2005, 40, 607–619. [Google Scholar]
- Sun, W; Cao, JG; Teng, K; Meighen, EA. Biosynthesis of poly-3-hydroxybutyrate in the luminescent bacterium, Vibrio harveyi, and regulation by the lux autoinducer, N-(3-hydroxybutanoyl)homoserine lactone. J Biol Chem 1994, 269, 20785–20790. [Google Scholar]
- Weiner, RM. Biopolymers from marine prokaryotes. Trends Biotechnol 1997, 15, 390–394. [Google Scholar]
- Higgins, DG; Sharp, PM. Fast and sensitive multiple sequence alignments on a microcomputer. Comput Appl Biosci 1989, 5, 151–153. [Google Scholar]
- Chen, CW; Don, TM; Yen, HF. Enzymatic extruded starch as a carbon source for the production of poly(3-hydroxybutyrate-co-3-hydroxyvalerate) by Haloferax mediterranei. Process Biochem 2006, 41, 2289–2296. [Google Scholar]
- Don, TM; Chen, CW; Chan, TH. Preparation and characterizationof poly(hydroxyalkanoate) from the fermentation of Haloferax mediterranei. J Biomater Sci Polym Ed 2006, 17, 1425–1438. [Google Scholar]
- Huang, TY; Duan, KJ; Huang, SY; Chen, CW. Production of polyhydroxyalkanoates from inexpensive extruded rice bran and starch by Haloferax mediterranei. J Ind Microbiol Biotechnol 2006, 33, 701–706. [Google Scholar]
- Mokhtari-Hosseini, ZB; Vasheghani-Farahani, E; Heidarzadeh-Vazifekhoran, A; Shojaosadati, SA; Karimzadeh, R; Khosravi Darani, K. Statistical media optimization for growth and PHB production from methanol by a methylotrophic bacterium. Bioresour Technol 2009, 100, 2436–2443. [Google Scholar]
- Ghanem, NB; Mabrouk, ME; Sabry, SA; El-Badan, DE. Degradation of polyesters by a novel marine Nocardiopsis aegyptia sp. nov.: Application of Plackett-Burman experimental design for the improvement of PHB depolymerase activity. J Gen Appl Microbiol 2005, 51, 151–158. [Google Scholar]
- Taran, M; Amirkhani, H. Strategies of poly(3-hydroxybutyrate) synthesis by Haloarcula sp. IRU1 utilizing glucose as carbon source: Optimization of culture conditions by Taguchi methodology. Int J Biol Macromol 2010, 47, 632–634. [Google Scholar]
- Quillaguamán, J; Muñoz, M; Mattiasson, B; Hatti-Kaul, R. Optimizing conditions for poly(beta-hydroxybutyrate) production by Halomonas boliviensis LC1 in batch culture with sucrose as carbon source. Appl Microbiol Biotechnol 2007, 74, 981–986. [Google Scholar]
- Chien, CC; Chen, CC; Choi, MH; Kung, SS; Wei, YH. Production of poly-beta-hydroxybutyrate (PHB) by Vibrio spp. isolated from marine environment. J Biotechnol 2007, 132, 259–263. [Google Scholar]
- Merrick, JM; Steger, R; Dombroski, D. Hydrolysis of native poly(hydroxybutyrate) granules (PHB), crystalline PHB, and artificial amorphous PHB granules by intracellular and extracellular depolymerases. Int J Biol Macromol 1999, 25, 129–134. [Google Scholar]
- Jendrossek, D; Handrick, R. Microbial degradation of polyhydroxyalkanoates. Annu Rev Microbiol 2002, 56, 403–432. [Google Scholar]
- Jendrossek, D; Selchow, O; Hoppert, M. Poly(3-hydroxybutyrate) granules at the early stages of formation are localized close to the cytoplasmic membrane in Caryophanon latum. Appl Environ Microbiol 2007, 73, 586–593. [Google Scholar]
- Wei, YH; Chen, WC; Huang, CK; Wu, HS; Sun, YM; Lo, CW; Janarthanan, OM. Screening and Evaluation of Polyhydroxybutyrate-Producing Strains from Indigenous Isolate Cupriavidus taiwanensis Strains. Int J Mol Sci 2011, 12, 252–265. [Google Scholar]
- Braunegg, G; Sonnleitner, B; Lafferty, RM. A rapid gas chromatographic method for the determination of poly-β-hydroxybutyric acid in microbial biomass. Appl Microbiol Biotechnol 1978, 6, 29–37. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Carbon Sources | CDW (g/L) | PHB Content (%) | PHB Produciton (g/L) |
|---|---|---|---|
| Glucose | 2.73 ±0.03 | 17.49 ±0.18 | 0.35 ±0.03 |
| Fructose | 2.78 ±0.03 | 17.92 ±2.11 | 0.52 ±0.05 |
| Sucrose | 0.76 ±0.01 | *ND. | *ND. |
| Molasses | 0.69 ±0.01 | 2.39 ±0.23 | 0.02 ±0.01 |
| Starch | 0.54 ±0.05 | *ND. | *ND. |
| Acetic acid | 5.04 ±0.05 | 4.99 ±0.67 | 0.25 ±0.04 |
| Mannitol | 3.06 ±0.03 | 10.77 ±1.12 | 0.33 ±0.02 |
| Glycerol | 3.69 ±0.04 | 15.68 ±1.73 | 0.58 ±0.05 |
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wei, Y.-H.; Chen, W.-C.; Wu, H.-S.; Janarthanan, O.-M. Biodegradable and Biocompatible Biomaterial, Polyhydroxybutyrate, Produced by an Indigenous Vibrio sp. BM-1 Isolated from Marine Environment. Mar. Drugs 2011, 9, 615-624. https://doi.org/10.3390/md9040615
Wei Y-H, Chen W-C, Wu H-S, Janarthanan O-M. Biodegradable and Biocompatible Biomaterial, Polyhydroxybutyrate, Produced by an Indigenous Vibrio sp. BM-1 Isolated from Marine Environment. Marine Drugs. 2011; 9(4):615-624. https://doi.org/10.3390/md9040615
Chicago/Turabian StyleWei, Yu-Hong, Wei-Chuan Chen, Ho-Shing Wu, and Om-Murugan Janarthanan. 2011. "Biodegradable and Biocompatible Biomaterial, Polyhydroxybutyrate, Produced by an Indigenous Vibrio sp. BM-1 Isolated from Marine Environment" Marine Drugs 9, no. 4: 615-624. https://doi.org/10.3390/md9040615