Balibalosides, an Original Family of Glucosylated Sesterterpenes Produced by the Mediterranean Sponge Oscarella balibaloi

 ,
,  ,
, 
Abstract
:
1. Introduction

2. Results and Discussion
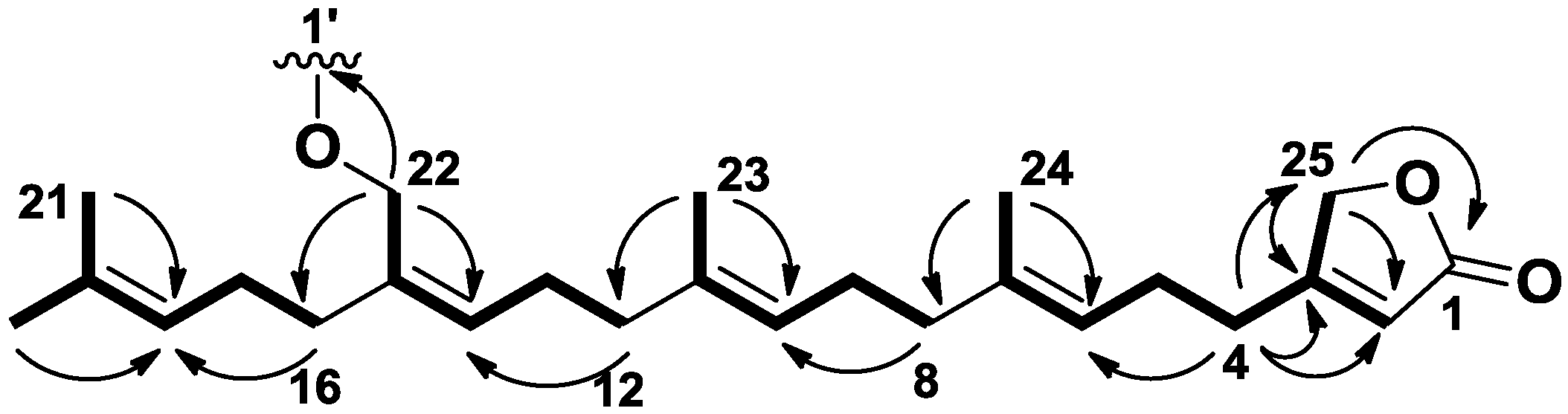
2.1. Structure Identification

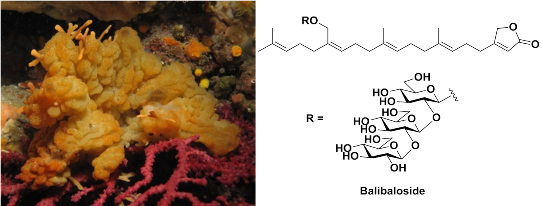{kind=link}
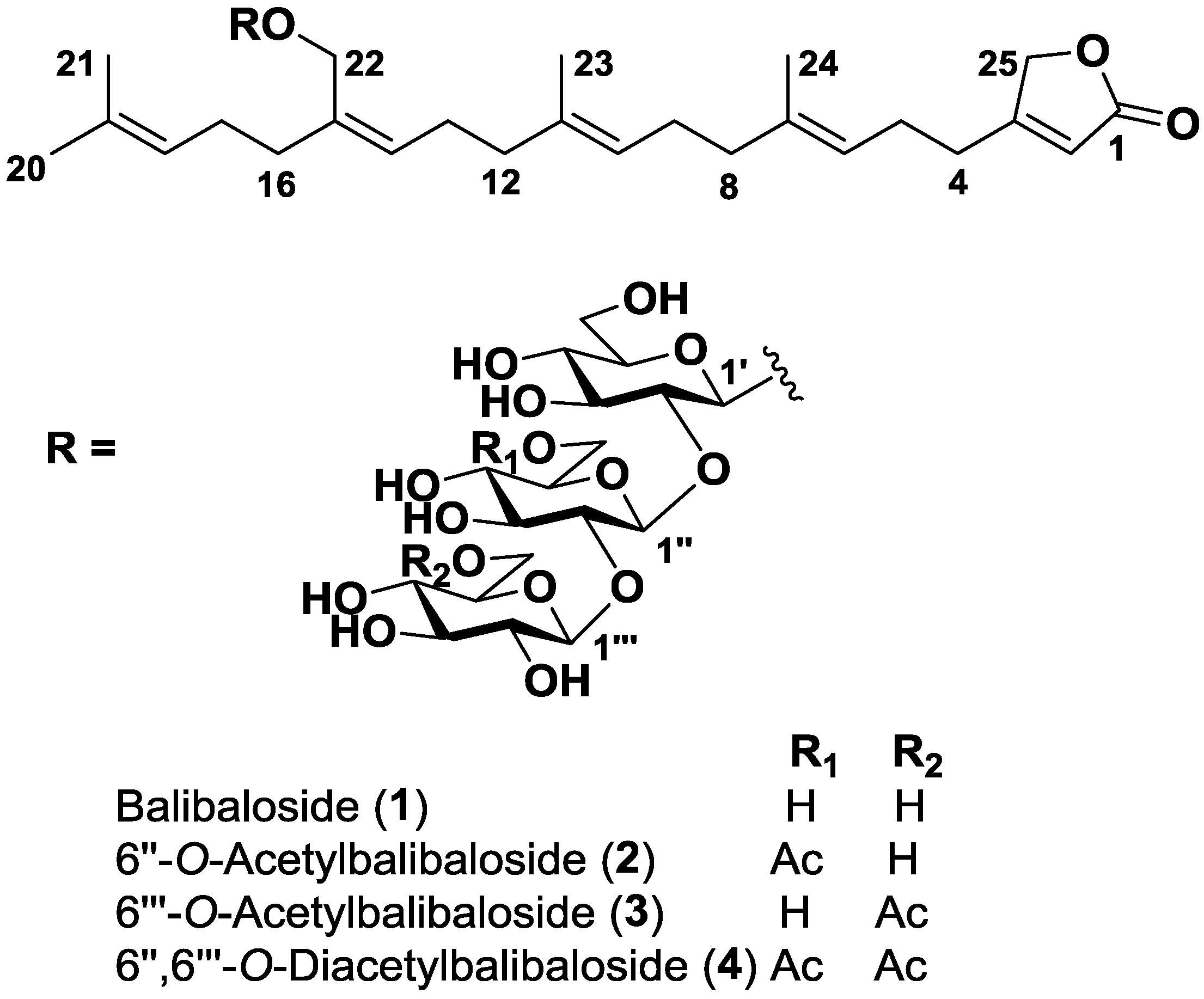{kind=link}
{kind=link}
{kind=link}
| Residue | Position | 1 | 2 | 3 | 4 | ||||
|---|---|---|---|---|---|---|---|---|---|
| δC | δH, mult. (J in Hz) | δC | δH, mult. (J in Hz) | δC | δH, mult. (J in Hz) | δC | δH, mult. (J in Hz) | ||
| Glu-1 | 1′ | 101.3 | 4.36, d (7.3) | 101.3 | 4.35, d (7.6) | 101.3 | 4.36, d (8.0) | 101.3 | 4.34, d (8.0) |
| 2′ | 83.8 | 3.37, dd (9.1, 7.3) | 84.9 | 3.34, m | 83.9 | 3.36, m | 85.0 | 3.30, m | |
| 3′ | 77.9 | 3.64, t (9.1) | 77.7 | 3.64, t (9.1) | 77.8 | 3.56, t (9.0) | 77.5 | 3.57, t (9.0) | |
| 4′ | 70.8 | 3.35, m | 70.9 | 3.36, m | 70.9 | 3.36, m | 70.9 | 3.35, m | |
| 5′ | 77.9 | 3.24, m | 77.9 | 3.24, ddd (9.3, 5.4, 2.3) | 78.1 | 3.24, m | 77.8 | 3.24, m | |
| 6′ | 62.7 | 3.86, dd (11.8, 2.1) | 62.7 | 3.86, dd (11.9, 2.3) | 62.7 | 3.86, dd (11.9, 2.3) | 62.5 | 3.85, dd (11.9, 2.3) | |
| 3.63, dd (11.8, 5.6) | 3.69, dd (11.9, 5.6) | 3.69, dd (11.9, 5.6) | 3.68, dd (11.9, 5.6) | ||||||
| Glu-2 | 1″ | 103.8 | 4.71, d (7.9) | 104.2 | 4.65, d (7.8) | 103.6 | 4.69, d (7.7) | 103.8 | 4.62, d (8.0) |
| 2″ | 85.2 | 3.40, dd (9.1, 7.9) | 84.8 | 3.44, dd (9.1, 7.8) | 84.8 | 3.39, dd (9.0, 7.7) | 84.6 | 3.41, dd (9.1, 7.8) | |
| 3″ | 77.8 | 3.56, t (9.1) | 77.7 | 3.55, t (9.1) | 77.8 | 3.56, t (9.0) | 77.5 | 3.55, t (9.1) | |
| 4″ | 71.4 | 3.29, dd (9.1, 7.4) | 71.4 | 3.30, m | 71.6 | 3.30, m | 71.2 | 3.30, m | |
| 5″ | 77.9 | 3.24, m | 75.1 | 3.41, m | 78.0 | 3.24, m | 75.0 | 3.41, m | |
| 6″ | 63.0 | 3.80, dd (12.0, 2.3) | 64.7 | 4.31, br d (12.0) | 63.0 | 3.80, dd (12.0, 2.3) | 64.7 | 4.31, br d (12.0) | |
| 3.63, dd (12.0, 6.0) | 4.20, dd (12.0, 5.0) | 3.63, dd (12.0, 6.0) | 4.19, dd (12.0, 5.0) | ||||||
| 6″-O-Ac | 20.7 | 2.04, s | 20.7 | 2.04, s | |||||
| 170.5 | 170.5 | ||||||||
| Glu-3 | 1″′ | 106.2 | 4.60, d (7.7) | 106.0 | 4.63, d (8.0) | 105.8 | 4.63,d (8.0) | 105.8 | 4.66, d (8.0) |
| 2″′ | 76.3 | 3.27, dd (9.1, 7.7) | 76.2 | 3.27, m | 76.0 | 3.28, m | 76.0 | 3.28, m | |
| 3″′ | 77.6 | 3.38, dd (9.0, 7.8) | 77.5 | 3.38, m | 77.7 | 3.38, t (9.0) | 77.6 | 3.38, t (9.0) | |
| 4″′ | 71.0 | 3.33, m | 70.9 | 3.35, m | 71.0 | 3.35, m | 70.9 | 3.35, m | |
| 5″′ | 78.9 | 3.34, m | 78.9 | 3.35, m | 76.1 | 3.52, ddd (9.0, 5.0, 2.0) | 76.0 | 3.52, ddd (9.0, 5.0, 2.0) | |
| 6″′ | 62.4 | 3.91, dd (12.0, 1.6) | 62.4 | 3.91, dd (12.0, 1.8) | 64.3 | 4.47, dd (12.0, 2.0) | 64.3 | 4.47, dd (12.0, 2.0) | |
| 3.73, dd (12.0, 6.0) | 3.73, dd (12.0, 5.0) | 4.22, dd (12.0, 5.0) | 4.22, dd (12.0, 5.0) | ||||||
| 2″′-O-Ac | |||||||||
| 6″′-O-Ac | 21.0 | 2.10, s | 21.0 | 2.10, s | |||||
| 170.5 | 170.5 | ||||||||
| Position | δC, mult. | δH, mult. (J in Hz) |
|---|---|---|
| 1 | 177.0, C | |
| 2 | 115.7, CH | 5.87, quint (1.6) |
| 3 | 174.2, C | |
| 4 | 29.5, CH2 | 2.52, br t (7.2) |
| 5 | 26.7, CH2 | 2.34, q (7.2) |
| 6 | 123.9, CH | 5.17, br t (7.2) |
| 7 | 138.1, C | |
| 8 | 40.7, CH2 | 2.04, br t (7.4) |
| 9 | 27.5, CH2 | 2.12, m |
| 10 | 125.7, CH | 5.12, t (7.0) |
| 11 | 135.8, C | |
| 12 | 40.9, CH2 | 2.02, m |
| 13 | 27.2, CH2 | 2.19, br q (6.8) |
| 14 | 130.8, CH | 5.39, t (7.2) |
| 15 | 136.5, C | |
| 16 | 35.9, CH2 | 2.13, m |
| 17 | 27.8, CH2 | 2.12, m |
| 18 | 125.7, CH | 5.12, t (7.0) |
| 19 | 132.2, C | |
| 20 | 26.0, CH3 | 1.69, s |
| 21 | 18.6, CH3 | 1.63, s |
| 22 | 67.0, CH2 | 4.32, s a |
| 23 | 16.2, CH3 | 1.62, s |
| 24 | 16.2, CH3 | 1.65, s |
| 25 | 75.0, CH2 | 4.85 b |
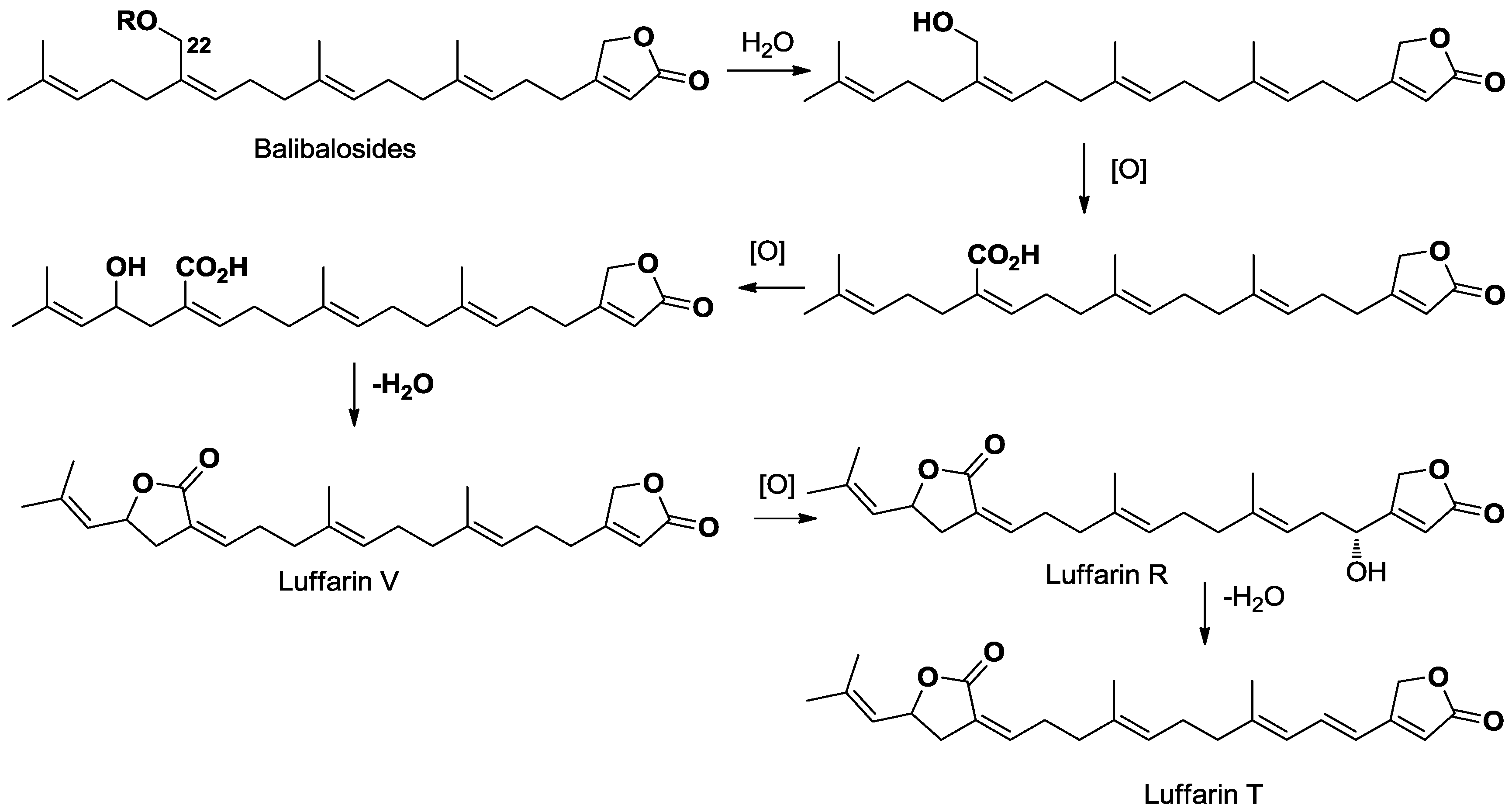
2.2. Biosynthetic Considerations
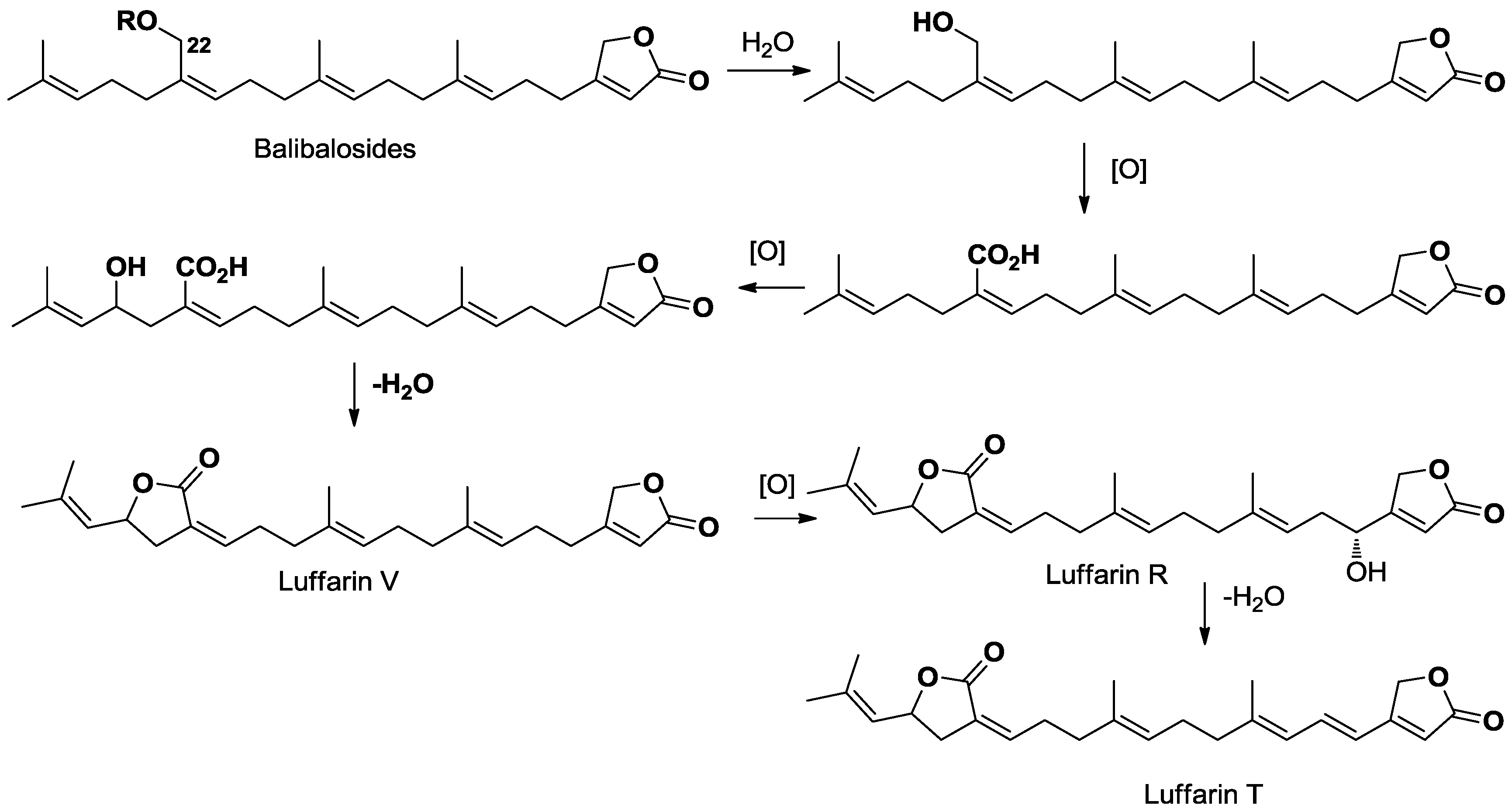
2.3. Bioassays
3. Experimental Section
3.1. General Experimental Procedures
3.2. Animal Material
3.3. Extraction and Isolation
3.4. Balibaloside (1)
3.5. 6″-O-Acetylbalibaloside (2)
3.6. 6″′-O-Acetylbalibaloside (3)
3.7. 6″,6″′-O-Diacetylbalibaloside (4)
3.8. Absolute Configuration of the Sugar Residues
3.9. Cytotoxicity Bioassays
3.10. Antimicrobial Bioassays

4. Conclusions
Acknowledgments
References
- Gerovasileiou, V.; Voultsiadou, E. Marine caves of the Mediterranean Sea: A sponge biodiversity reservoir within a biodiversity hotspot. PLoS One 2012, 7, e39873. [Google Scholar] [CrossRef]
- Gazave, E.; Lapébie, P.; Ereskovsky, A.; Vacelet, J.; Renard, E.; Cárdenas, P.; Borchiellini, C. No longer Demospongiae: Homoscleromorpha formal nomination as a fourth class of Porifera. Hydrobiologia 2012, 687, 3–10. [Google Scholar] [CrossRef]
- Ozbek, S.; Balasubramanian, P.G.; Holstein, T.W. Cnidocyst structure and the biomechanics of discharge. Toxicon 2009, 54, 1038–1045. [Google Scholar] [CrossRef]
- Maçek, P.; Belmonte, G.; Pederzolli, C.; Menestrina, G. Mechanism of action of equinatoxin II, a cytolysin from the sea anemone Actinia equina L. belonging to the family of actinoporins. Toxicology 1994, 87, 205–227. [Google Scholar] [CrossRef]
- Wang, X.; Lavrov, D.V. Mitochondrial genome of the Homoscleromorph Oscarella carmela (Porifera, Demospongiae) reveals unexpected complexity in the common ancestor of sponges and other animals. Mol. Biol. Evol. 2007, 24, 363–373. [Google Scholar] [CrossRef]
- Ivanišević, J.; Thomas, O.P.; Lejeusne, C.; Chevaldonné, P.; Pérez, T. Metabolic fingerprinting as an indicator of biodiversity: towards understanding inter-specific relationships among Homoscleromorpha sponges. Metabolomics 2011, 7, 289–304. [Google Scholar] [CrossRef]
- Martins, R.D.; Alves, R.S.; Martins, A.M.C.; Barbosa, P.S.F.; Evangelista, J.S.A.M.; Evangelista, J.J.F.; Ximenes, R.M.; Toyama, M.H.; Toyama, D.O.; Souza, A.J.F.; et al. Purification and characterization of the biological effects of phospholipase A2 from sea anemone Bunodosoma caissarum. Toxicon 2009, 54, 413–420. [Google Scholar] [CrossRef]
- Ivanisevic, J.; Thomas, O.P.; Pedel, L.; Penez, N.; Ereskovsky, A.V.; Culioli, G.; Perez, T. Biochemical trade-offs: Evidence for ecologically linked secondary metabolism of the sponge Oscarella balibaloi. PLoS One 2011, 6, e28059. [Google Scholar]
- Turk, T.; Kem, W.R. The phylum Cnidaria and investigations of its toxins and venoms until 1990. Toxicon 2009, 54, 1031–1037. [Google Scholar] [CrossRef]
- Duus, J.Ø.; Gotfredsen, C.H.; Bock, K. Carbohydrate structural determination by NMR spectroscopy: Modern methods and limitations. Chem. Rev. 2000, 100, 4589–4614. [Google Scholar] [CrossRef]
- Costantino, V.; Fattorusso, E.; Menna, M.; Taglialatela-Scafati, O. Chemical diversity of bioactive marine natural products: An illustrative case study. Curr. Med. Chem. 2004, 11, 1671–1692. [Google Scholar] [CrossRef]
- Rahm, F.; Hayes, P.Y.; Kitching, W. Metabolites from marine sponges of the genus Plakortis. Heterocycles 2004, 64, 523–575. [Google Scholar] [CrossRef]
- Aiello, A.; Fattorusso, E.; Magno, S.; Mayol, L.; Menna, M. Isolation of two novel 5α,6α-epoxy-7-ketosterols from the encrusting demospongia Oscarella lobularis. J. Nat. Prod. 1990, 53, 487–491. [Google Scholar] [CrossRef]
- Aiello, A.; Fattorusso, E.; Magno, S.; Menna, M. Isolation of five new 5α-hydroxy-6-keto-Δ7 sterols from the marine sponge Oscarella lobularis. Steroids 1991, 56, 337–340. [Google Scholar] [CrossRef]
- Cimino, G.; de Stefano, S.; Minale, L. Long alkyl chains, 3-substituted pyrrole-2-aldehyde (-2-carboxylic acid and methyl ester) from the marine sponge Oscarella lobularis. Experientia 1975, 31, 1387–1389. [Google Scholar] [CrossRef]
- De Rosa, S.; Carbonelli, S. Two new luffarin derivatives from the Adriatic Sea sponge Fasciospongia cavernosa. Tetrahedron 2006, 62, 2845–2849. [Google Scholar] [CrossRef]
- Gerwig, G.J.; Kamerling, J.P.; Vliegenthart, J.F.G. Determination of the absolute configuration of monosaccharides in complex carbohydrates by capillary g.l.c. Carbohydr. Res. 1979, 77, 1–7. [Google Scholar] [CrossRef]
- Kalinin, V.I.; Ivanchina, N.V.; Krasokhin, V.B.; Makarieva, T.N.; Stonik, V.A. Glycosides from marine sponges (Porifera, Demospongiae): Structures, taxonomical distribution, biological activities and biological roles. Mar. Drugs 2012, 10, 1671–1710. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, L.; Jung, J.H.; Zhang, S. Sesterterpenoids. Nat. Prod. Rep. 2007, 24, 1401–1429. [Google Scholar] [CrossRef]
- Ebada, S.S.; Lin, W.; Proksch, P. Bioactive sesterterpenes and triterpenes from marine sponges: Occurrence and pharmacological significance. Mar. Drugs 2010, 8, 313–346. [Google Scholar] [CrossRef]
- Costantino, V.; Fattorusso, E.; Imperatore, C.; Mangoni, A. Glycolipids from sponges, 10. Plaxyloside from the marine sponge Plakortis simplex: An improved strategy for NMR structural studies of carbohydrate chains. Eur. J. Org. Chem. 2001, 4457–4462. [Google Scholar]
- Costantino, V.; Fattorusso, E.; Mangoni, A.; Di Rosa, M.; Ianaro, A. Glycolipids from sponges. Part 8: Plakopolyprenoside from the marine sponge Plakortis simplex. An improved procedure for isolation of glycolipids as peracetyl derivatives. Tetrahedron 2000, 56, 1393–1395. [Google Scholar] [CrossRef]
- Butler, M.S.; Capon, R.J. The luffarins (A–Z), novel terpenes from an Australian marine sponge, Luffariella geometrica. Aust. J. Chem. 1992, 45, 1705–1743. [Google Scholar] [CrossRef]
- Alley, M.C.; Scudiero, D.A.; Monks, A.; Hursey, M.L.; Czerwinski, M.J.; Fine, D.L.; Abbott, B.J.; Mayo, J.G.; Shoemaker, R.H.; Boyd, M.R. Feasibility of drug screening with panels of human tumor cell lines using a microculture tetrazolium assay. Cancer Res. 1988, 48, 589–601. [Google Scholar]
- Van de Loosdrecht, A.A.; Beelen, R.H.J.; Ossenkoppele, G.J.; Broekhoven, M.G.; Langenhuijsen, M.M.A.C. A tetrazolium-based colorimetric MTT assay to quantitate human monocyte mediated cytotoxicity against leukemic cells from cell lines and patients with acute myeloid leukemia. J. Immunol. Methods 1994, 174, 311–320. [Google Scholar] [CrossRef]
- Slater, T.F.; Sawyer, B.; Sträuli, U. Studies on succinate-tetrazolium reductase systems: III. Points of coupling of four different tetrazolium salts III. Points of coupling of four different tetrazolium salts. Biochim. Biophys. Acta 1963, 77, 383–393. [Google Scholar] [CrossRef]
- Ecimar. Available online: http://www.ecimar.org (accessed on 1 April 2013).
- BAMMBO. Available online: http://www.bammbo.eu (accessed on 1 April 2013).
- Samples Availability: Available from the authors.
Supplementary Files
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Audoin, C.; Bonhomme, D.; Ivanisevic, J.; Cruz, M.D.l.; Cautain, B.; Monteiro, M.C.; Reyes, F.; Rios, L.; Perez, T.; Thomas, O.P. Balibalosides, an Original Family of Glucosylated Sesterterpenes Produced by the Mediterranean Sponge Oscarella balibaloi. Mar. Drugs 2013, 11, 1477-1489. https://doi.org/10.3390/md11051477
Audoin C, Bonhomme D, Ivanisevic J, Cruz MDl, Cautain B, Monteiro MC, Reyes F, Rios L, Perez T, Thomas OP. Balibalosides, an Original Family of Glucosylated Sesterterpenes Produced by the Mediterranean Sponge Oscarella balibaloi. Marine Drugs. 2013; 11(5):1477-1489. https://doi.org/10.3390/md11051477
Chicago/Turabian StyleAudoin, Coralie, Dominique Bonhomme, Julijana Ivanisevic, Mercedes De la Cruz, Bastien Cautain, Maria Cândida Monteiro, Fernando Reyes, Laurent Rios, Thierry Perez, and Olivier P. Thomas. 2013. "Balibalosides, an Original Family of Glucosylated Sesterterpenes Produced by the Mediterranean Sponge Oscarella balibaloi" Marine Drugs 11, no. 5: 1477-1489. https://doi.org/10.3390/md11051477
APA StyleAudoin, C., Bonhomme, D., Ivanisevic, J., Cruz, M. D. l., Cautain, B., Monteiro, M. C., Reyes, F., Rios, L., Perez, T., & Thomas, O. P. (2013). Balibalosides, an Original Family of Glucosylated Sesterterpenes Produced by the Mediterranean Sponge Oscarella balibaloi. Marine Drugs, 11(5), 1477-1489. https://doi.org/10.3390/md11051477