Glycobiology of Reproductive Processes in Marine Animals: The State of the Art



{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
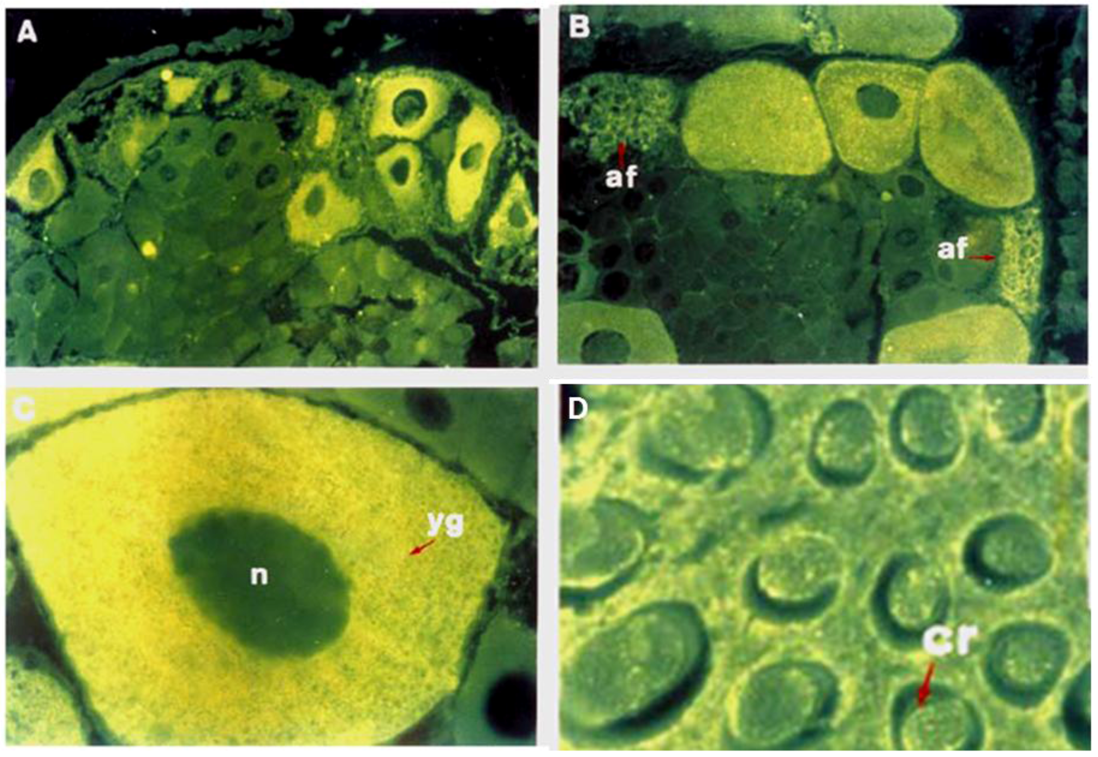
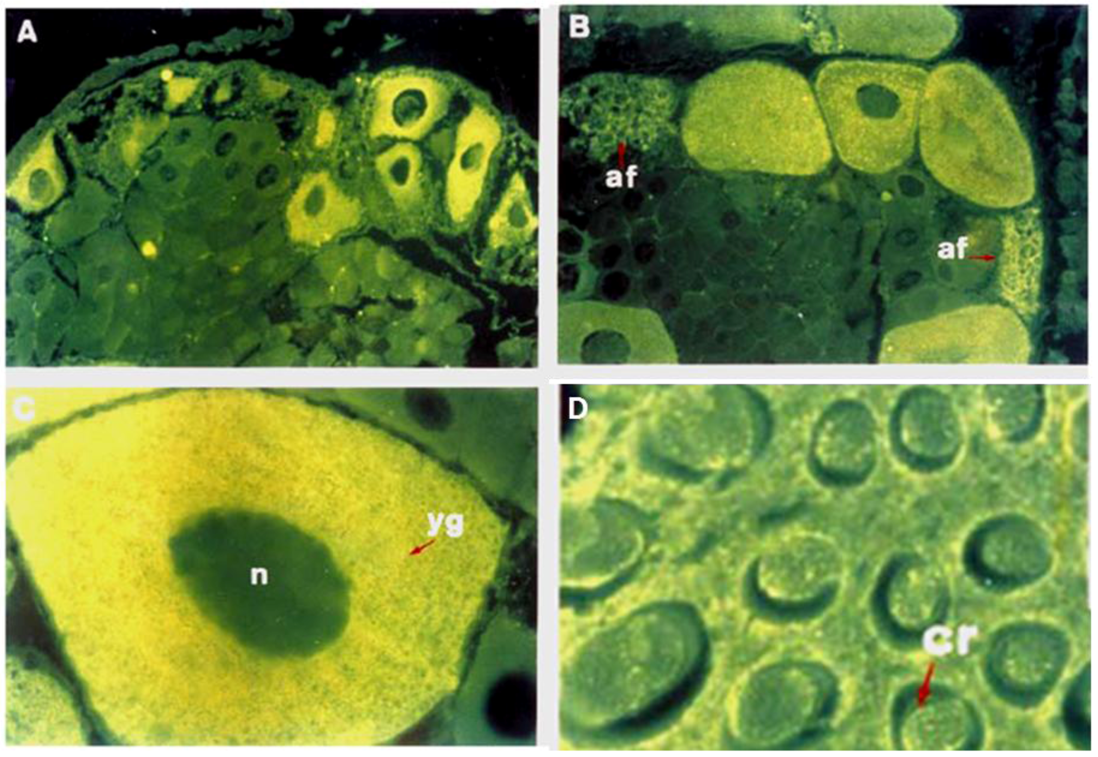
2. The Glycobiology in Oogenesis
3. The Glycobiology in Spermatogenesis
4. The Glycobiology in Fertilization
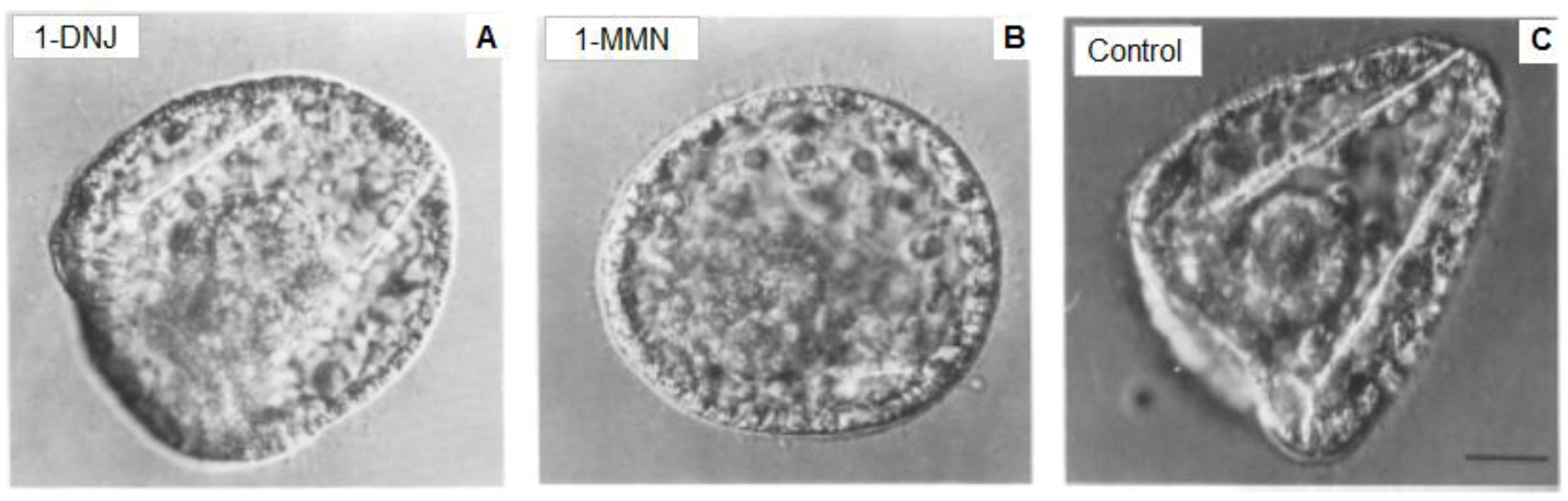
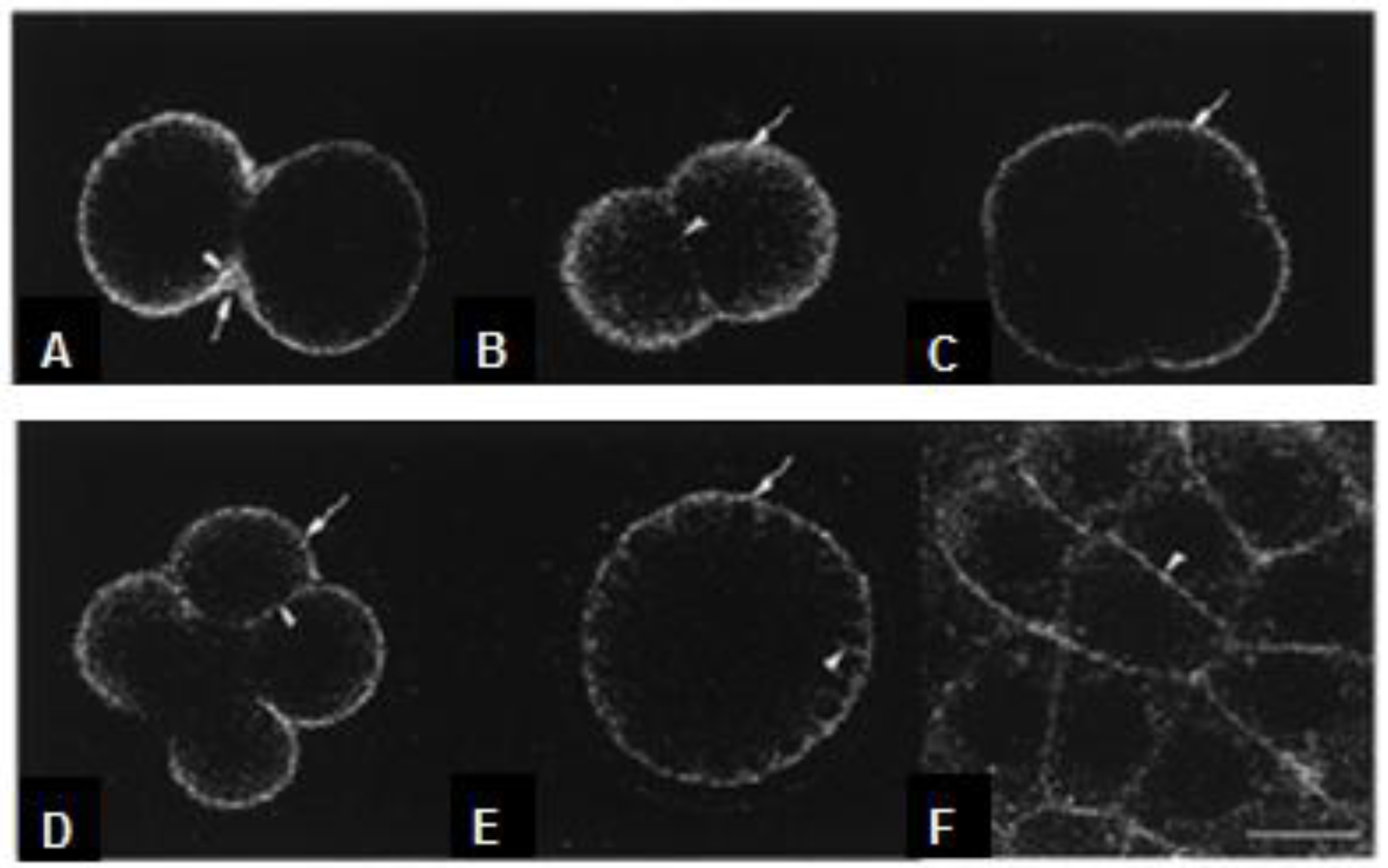
5. The Glycobiology in Embryo Development
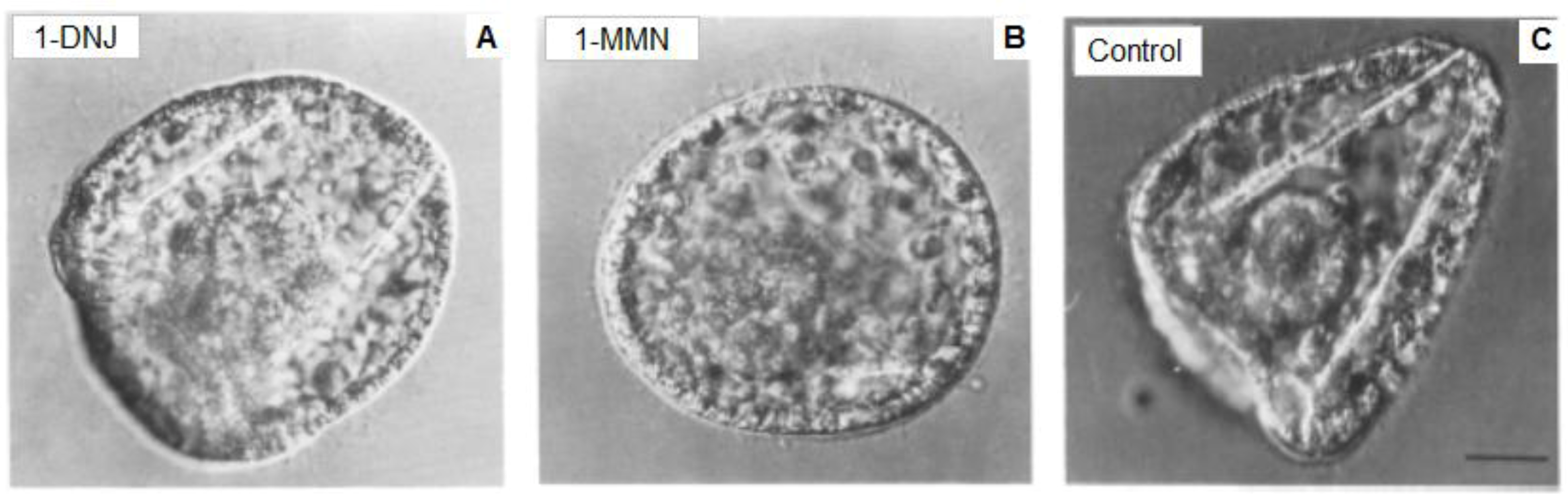

6. Conclusions
Acknowledgments
Conflict of Interest
References
- Caldwell, G.S.; Pagett, H.E. Marine glycobiology: Current status and future perspectives. Mar. Biotechnol. 2010, 12, 241–252. [Google Scholar] [CrossRef]
- Shriver, Z.; Raguram, S.; Sasisekharan, R. Glycomics: A pathway to a class of new and improved therapeutics. Nat. Rev. Drug Discov. 2004, 3, 863–873. [Google Scholar]
- Shur, B.D. Glycobiology: The beginning of a sweet tale. Curr. Biol. 1994, 4, 996–999. [Google Scholar] [CrossRef]
- O’Donnell, N. Intracellular glycosylation and development. Biochim. Biophys. Acta 2002, 1573, 336–345. [Google Scholar] [CrossRef]
- Morris, H.R.; Dell, A.; Easton, R.L.; Panico, M.; Koistinen, H.; Koistinen, R.; Oehninger, S.; Patankar, M.S.; Seppala, M.; Clark, G.F. Gender-specific glycosylation of human glycodelin affects its contraceptive activity. J. Biol. Chem. 1996, 271, 32159–32167. [Google Scholar]
- Dell, A.; Morris, H.R.; Easton, R.L.; Patankar, M.; Clark, G.F. The glycobiology of gametes and fertilization. Biochim. Biophys. Acta 1999, 1473, 196–205. [Google Scholar] [CrossRef]
- Sarasquete, C.; Cardenas, S.; de Gonzalez, C.M.; Pascual, E. Oogenesis in the bluefin tuna, Thunnus thynnus L.: A histological and histochemical study. Histol. Histopathol. 2002, 17, 775–788. [Google Scholar]
- Ortiz-Delgado, J.B.; Porcelloni, S.; Fossi, C.; Sarasquete, C. Histochemical characterisation of oocytes of the swordfish Xiphias gladius. Sci. Mar. 2008, 72, 549–564. [Google Scholar]
- Patino, R.; Sullivan, C.V. Ovarian follicle growth, maturation, and ovulation in teleost fish. Fish. Physiol. Biochem. 2002, 26, 57–70. [Google Scholar] [CrossRef]
- Inoue, S.; Iwasaki, M. Isolation of a novel glycoprotein from the eggs of rainbowtrout: Occurrence of disialosyl groups on all carbohydrate chains. Biochem. Biophys. Res. Commun. 1978, 83, 1018–1023. [Google Scholar] [CrossRef]
- Kitajima, K.; Sorimachi, H.; Inoue, S.; Inoue, Y. Comparative structures of the apopolysialoglycoproteins from unfertilized and fertilized eggs of salmonid fishes. Biochemistry 1988, 27, 7141–7145. [Google Scholar] [CrossRef]
- Song, Y.; Kitajima, K.; Inoue, Y. New tandem-repeating peptide structures in polysialoglycoproteins from the unfertilized eggs of kokanee salmon. Arch. Biochem. Biophys. 1990, 283, 167–172. [Google Scholar] [CrossRef]
- Seko, A.; Kitajima, K.; Iwasaki, M.; Inoue, S.; Inoue, Y. Structural studies of fertilization-associated carbohydrate-rich glycoproteins (hyosophorin) isolated from the fertilized and unfertilized eggs of flounder, Paralichthys olivaceus. Presence of a novel penta-antennary N-linked glycan chain in the tandem repeating glycopeptide unit of hyosophorin. J. Biol. Chem. 1989, 264, 15922–15929. [Google Scholar]
- Iwasaki, M.; Inoue, S. Structures of the carbohydrate units of polysialoglycoproteins isolated from the eggs of four species of salmonid fishes. Glycoconj. J. 1985, 2, 209–228. [Google Scholar] [CrossRef]
- Kitajima, K.; Inoue, Y.; Inoue, S. Polysialoglycoproteins of Salmonidae fish eggs. Complete structure of 200-kDa polysialoglycoprotein from the unfertilized eggs of rainbow trout (Salmo gairdneri). J. Biol. Chem. 1986, 261, 5262–5269. [Google Scholar]
- Nadano, D.; Iwasaki, M.; Endo, S.; Kitajima, K.; Inoue, S.; Inoue, Y. A naturally occurring deaminated neuraminic acid, 3-deoxy-D-glycero-D-galacto-nonulosonic acid (KDN). ts unique occurrence at the nonreducing ends of oligosialyl chains in polysialoglycoprotein of rainbow trout eggs. J. Biol. Chem. 1986, 261, 11550–11557. [Google Scholar]
- Asahina, S.; Sato, C.; Matsuno, M.; Matsuda, T.; Colley, K.; Kitajima, K. Involvement of the alpha2,8-polysialyltransferases II/STX and IV/PST in the biosynthesis of polysialic acid chains on the O-linked glycoproteins in rainbow trout ovary. J. Biochem. 2006, 140, 687–701. [Google Scholar] [CrossRef]
- Inoue, S.; Kanamori, A.; Kitajima, K.; Inoue, Y. KDN-glycoprotein: A novel deaminated neuraminic acid-rich glycoprotein isolated from vitelline envelope of rainbow trout eggs. Biochem. Biophys. Res. Commun. 1988, 153, 172–176. [Google Scholar] [CrossRef]
- Accogli, G.; Zizza, S.; Garcia-Lopez, A.; Sarasquete, C.; Desantis, S. Lectin-binding pattern of the Senegalese sole Solea senegalensis oogenesis. Microsc. Res. Tech. 2012, 75, 1124–1135. [Google Scholar] [CrossRef]
- Mandich, A.; Massari, A.; Bottero, S.; Marino, G. Histological and histochemical study of female germ cell development in the dusky grouper Epinephelus marginatus (Lowe, 1834). Eur. J. Histochem. 2002, 46, 87–100. [Google Scholar]
- Longo, F.J. Fertilization: A comparative ultrastructural review. Biol. Reprod. 1973, 9, 149–215. [Google Scholar]
- Schuel, H. Secretory Functions of Egg Cortical Granules. In Biology of Fertilization; Metz, C.B., Monroy, A., Eds.; Academic Press: San Diego, CA, USA, 1985; Volume 3, pp. 1–44. [Google Scholar]
- Zamboni, L. Ultrastructure of mammalian oocytes and ova. Biol. Reprod. 1970, 2, 44–63. [Google Scholar] [CrossRef]
- Wolf, D.P. On the contents of the cortical granules from Xenopus laevis eggs. Dev. Biol. 1974, 38, 14–29. [Google Scholar] [CrossRef]
- Gilkey, J.C. Mechanisms of fertilization in fishes. Am. Zool. 1981, 21, 359–375. [Google Scholar]
- Deits, T.; Farrance, M.; Kay, E.S.; Medill, L.; Turner, E.E.; Weidman, P.J.; Shapiro, B.M. Purification and properties of ovoperoxidase, the enzyme responsible for hardening the fertilization membrane of the sea urchin egg. J. Biol. Chem. 1984, 259, 13525–13533. [Google Scholar]
- Wessel, G.M.; Brooks, J.M.; Green, E.; Haley, S.; Voronina, E.; Wong, J.; Zaydfudim, V.; Conner, S. The biology of cortical granules. Int. Rev. Cytol. 2001, 209, 117–206. [Google Scholar] [CrossRef]
- Anderson, S.L.; Chang, E.S.; Clark, W.H.J. Timing of postvitellogenic ovarian changes in the ridgeback prawn Sicyonia ingentis (Penaeidae) determined by ovarian biopsy. Aquaculture 1984, 42, 257–271. [Google Scholar] [CrossRef]
- Lynn, J.W.; Clark, W.H.J. Physiological and biochemical investigations of the egg jelly release in Penaeus aztecus. Biol. Bull. 1987, 173, 451–460. [Google Scholar] [CrossRef]
- Pongtippatee-Taweepreda, P.; Chavadej, J.; Plodpai, P.; Pratoomchart, B.; Sobhon, P.; Weerachatyanukul, W.; Withyachumnarnkul, B. Egg activation in the black tiger shrimp Penaeus monodon. Aquaculture 2004, 234, 183–198. [Google Scholar] [CrossRef]
- Rankin, S.M.; Davis, R.W. Ultrastructure of oocytes of the shrimp, Penaeus vannamei: Cortical specialization formation. Tissue Cell 1990, 22, 879–893. [Google Scholar] [CrossRef]
- Kruevaisayawan, H.; Vanichviriyakit, R.; Withyachumnarnkul, B.; Chavadej, J.; Sobhon, P. Oogenesis and formation of cortical rods in the black tiger shrimp, Penaeus monodon. Aquaculture 2010, 301, 91–98. [Google Scholar] [CrossRef]
- Clark, J.W.H.; Yudin, A.I.; Lynn, J.W.; Griffin, F.J.; Pillai, M.C. Jelly layer formation in Penaeoidean shrimp eggs. Biol. Bull. 1990, 178, 295–299. [Google Scholar] [CrossRef]
- Tan-Fermin, J.D.; Pudadera, R.A. Ovarian maturation stages of the wild giant tiger prawn, Penaeus monodon Fabricius. Aquaculture 1989, 77, 229–242. [Google Scholar] [CrossRef]
- Khayat, M.; Babin, P.J.; Funkenstein, B.; Sammar, M.; Nagasawa, H.; Tietz, A.; Lubzens, E. Molecular characterization and high expression during oocyte development of a shrimp ovarian cortical rod protein homologous to insect intestinal peritrophins. Biol. Reprod. 2001, 64, 1090–1099. [Google Scholar] [CrossRef]
- Avarre, J.C.; Khayat, M.; Michelis, R.; Nagasawa, H.; Tietz, A.; Lubzens, E. Inhibition of de novo synthesis of a jelly layer precursor protein by crustacean hyperglycemic hormone family peptides and posttranscriptional regulation by sinus gland extracts in Penaeus semisulcatus ovaries. Gen. Comp. Endocrinol. 2001, 124, 257–268. [Google Scholar] [CrossRef]
- Kim, Y.K.; Kawazoe, I.; Tsutsui, N.; Jasmani, S.; Wilder, M.N.; Aida, K. Isolation and cDNA cloning of ovarian cortical rod protein in kuruma prawn Marsupenaeus japonicus (Crustacea: Decapoda: Penaeidae). Zool. Sci. 2004, 21, 1109–1119. [Google Scholar] [CrossRef]
- Kim, Y.K.; Tsutsui, N.; Kawazoe, I.; Okumura, T.; Kaneko, T.; Aida, K. Localization and developmental expression of mRNA for cortical rod protein in kuruma prawn Marsupenaeus japonicus. Zool. Sci. 2005, 22, 675–680. [Google Scholar] [CrossRef]
- Kruevaisayawan, H.; Vanichviriyakit, R.; Weerachatyanukul, W.; Withyachumnarnkul, W.; Sobhon, P. Biochemical characterization and physiological role of cortical rods in black tiger shrimp, Penaeus monodon. Aquaculture 2007, 270, 289–298. [Google Scholar]
- Matozzo, V.; Gagne, F.; Marin, M.G.; Ricciardi, F.; Blaise, C. Vitellogenin as a biomarker of exposure to estrogenic compounds in aquatic invertebrates: A review. Environ. Int. 2008, 34, 531–545. [Google Scholar] [CrossRef]
- Fabra, M.; Raldua, D.; Bozzo, M.G.; Deen, P.M.; Lubzens, E.; Cerda, J. Yolk proteolysis and aquaporin-1o play essential roles to regulate fish oocyte hydration during meiosis resumption. Dev. Biol. 2006, 295, 250–262. [Google Scholar] [CrossRef]
- Matsubara, T.; Ohkubo, N.; Andoh, T.; Sullivan, C.V.; Hara, A. Two forms of vitellogenin, yielding two distinct lipovitellins, play different roles during oocyte maturation and early development of barfin flounder, Verasper moseri, a marine teleost that spawns pelagic eggs. Dev. Biol. 1999, 213, 18–32. [Google Scholar] [CrossRef]
- Arukwe, A.; Goksoyr, A. Eggshell and egg yolk proteins in fish: Hepatic proteins for the next generation: Oogenetic, population, and evolutionary implications of endocrine disruption. Comp. Hepatol. 2003, 2, 4. [Google Scholar] [CrossRef] [Green Version]
- Abdu, U.; Davis, C.; Khalaila, I.; Sagi, A. The vitellogenin cDNA of Cherax quadricarinatus encodes a lipoprotein with calcium binding ability, and its expression is induced following the removal of the androgenic gland in a sexually plastic system. Gen. Comp. Endocrinol. 2002, 127, 263–272. [Google Scholar] [CrossRef]
- Khalaila, I.; Peter-Katalinic, J.; Tsang, C.; Radcliffe, C.M.; Aflalo, E.D.; Harvey, D.J.; Dwek, R.A.; Rudd, P.M.; Sagi, A. Structural characterization of the N-glycan moiety and site of glycosylation in vitellogenin from the decapod crustacean Cherax quadricarinatus. Glycobiology 2004, 14, 767–774. [Google Scholar] [CrossRef]
- Jaenicke, R. Protein stability and protein folding. Ciba Found. Symp. 1991, 161, 206–221. [Google Scholar]
- Raikhel, A.S.; Dhadialla, T.S. Accumulation of yolk proteins in insect oocytes. Annu. Rev. Entomol. 1992, 37, 217–251. [Google Scholar] [CrossRef]
- Warrier, S.; Subramoniam, T. Receptor mediated yolk protein uptake in the crab Scylla serrata: Crustacean vitellogenin receptor recognizes related mammalian serum lipoproteins. Mol. Reprod. Dev. 2002, 61, 536–548. [Google Scholar] [CrossRef]
- Shyu, A.B.; Raff, R.A.; Blumenthal, T. Expression of the vitellogenin gene in female and male sea urchin. Proc. Natl. Acad. Sci. USA 1986, 83, 3865–3869. [Google Scholar] [CrossRef]
- Ozaki, H.; Moriya, O.; Harrington, F.E. A glycoprotein in the accessory cell of the echinoid ovary and its role in vitellogenesis. Dev. Genes Evol. 1986, 195, 74–79. [Google Scholar]
- Unuma, T.; Suzuki, T.; Kurokawa, T.; Yamamoto, T.; Akiyama, T. A protein identical to the yolk protein is stored in the testis in male red sea urchin, Pseudocentrotus depressus. Biol. Bull. 1998, 194, 92–97. [Google Scholar] [CrossRef]
- Harrington, F.E.; Ozaki, H. The major yolk glycoprotein precursor in echinoids is secreted by coelomocytes into the coelomic plasma. Cell Differ. 1986, 19, 51–57. [Google Scholar] [CrossRef]
- Degroote, S.; Wolthoorn, J.; van Meer, G. The cell biology of glycosphingolipids. Semin. Cell Dev. Biol. 2004, 15, 375–387. [Google Scholar]
- Fishman, P.H. Recent advances in identifying the functions of gangliosides. Chem. Phys. Lipids 1986, 42, 137–151. [Google Scholar] [CrossRef]
- Hakomori, S. Glycosphingolipids in cellular interaction, differentiation, and oncogenesis. Annu. Rev. Biochem. 1981, 50, 733–764. [Google Scholar] [CrossRef]
- Hakomori, S. Bifunctional role of glycosphingolipids. Modulators for transmembrane signaling and mediators for cellular interactions. J. Biol. Chem. 1990, 265, 18713–18716. [Google Scholar]
- Hakomori, S. Structure and function of sphingoglycolipids in transmembrane signalling and cell-cell interactions. Biochem. Soc. Trans. 1993, 21, 583–595. [Google Scholar]
- Hakomori, S.I. Cell adhesion/recognition and signal transduction through glycosphingolipid microdomain. Glycoconj. J. 2000, 17, 143–151. [Google Scholar] [CrossRef]
- Kojima, H.; Tohsato, Y.; Kabayama, K.; Itonori, S.; Ito, M. Biochemical studies on sphingolipids of Artemia franciscana: Complex neutral glycosphingolipids. Glycoconj. J. 2012. [Google Scholar] [CrossRef]
- Yamamoto, T.; Teshima, T.; Saitoh, U.; Hoshi, M.; Shiba, T. Synthesis of ganglioside M5 from sea urchin egg. Tetrahedron Lett. 1994, 35, 2701–2704. [Google Scholar] [CrossRef]
- Kubo, H.; Irie, A.; Inagaki, F.; Hoshi, M. Gangliosides from the eggs of the sea urchin, Anthocidaris crassispina. J. Biochem. 1990, 108, 185–192. [Google Scholar]
- Nezuo, M.; Shogomori, H.; Hoshi, M.; Yamamoto, T.; Teshima, T.; Shiba, T.; Chiba, K. Developmental changes in localization of the main ganglioside during sea urchin embryogenesis. Glycobiology 2000, 10, 1243–1247. [Google Scholar] [CrossRef]
- Shogomori, H.; Chiba, K.; Kubo, H.; Hoshi, M. Non-plasmalemmal localisation of the major ganglioside in sea urchin eggs. Zygote 1993, 1, 215–223. [Google Scholar]
- Shogomori, H.; Chiba, K.; Hoshi, M. Association of the major ganglioside in sea urchin eggs with yolk lipoproteins. Glycobiology 1997, 7, 391–398. [Google Scholar] [CrossRef]
- Kubo, H.; Hoshi, M. Immunocytochemical study of the distribution of a ganglioside in sea urchin eggs. J. Biochem. 1990, 108, 193–199. [Google Scholar]
- Kubo, H.; Jiang, G.J.; Irie, A.; Morita, M.; Matsubara, T.; Hoshi, M. A novel ceramide trihexoside from the eggs of the sea urchin, Hemicentrotus pulcherrimus. J. Biochem. 1992, 111, 726–731. [Google Scholar]
- Kubo, H.; Jiang, G.J.; Irie, A.; Suzuki, M.; Inagaki, F.; Hoshi, M. A novel difucosylated neutral glycosphingolipid from the eggs of the sea urchin, Hemicentrotus pulcherrimus: I. Purification and structural determination of the glycolipid. J. Biochem. 1992, 112, 281–285. [Google Scholar]
- Kubo, H.; Irie, A.; Inagaki, F.; Hoshi, M. Melibiosylceramide as the sole ceramide dihexoside from the eggs of the sea urchin, Anthocidaris crassispina. J. Biochem. 1988, 104, 755–760. [Google Scholar]
- Inagaki, F.; Tate, S.; Kubo, H.; Hoshi, M. A novel difucosylated neutral glycosphingolipid from the eggs of the sea urchin, Hemicentrotus pulcherrimus: II. Structural determination by two-dimensional NMR. J. Biochem. 1992, 112, 286–289. [Google Scholar]
- Dumont, J.N.; Brummett, A.R. Egg envelopes in vertebrates. Dev. Biol. 1985, 1, 235–288. [Google Scholar]
- Glabe, C.G.; Grabel, L.B.; Vacquier, V.D.; Rosen, S.D. Carbohydrate specificity of sea urchin sperm bindin: A cell surface lectin mediating sperm-egg adhesion. J. Cell Biol. 1982, 94, 123–128. [Google Scholar] [CrossRef]
- Focarelli, R.; Rosa, D.; Rosati, F. Differentiation of the vitelline coat and the polarized site of sperm entrance in the egg of Unio elongatulus (Mollusca, Bivalvia). J. Exp. Zool. 1990, 254, 88–96. [Google Scholar] [CrossRef]
- Wikramanayake, A.H.; Clark, W.H. Two extracellular matrices from oocytes of the marine shrimp Sicyonia ingentis that independently mediate only primary or secondary sperm binding. Dev. Growth Differ. 1994, 36, 89–101. [Google Scholar]
- Honegger, T.G.; Koyanagi, R. The ascidian egg envelope in fertilization: Structural and molecular features. Int. J. Dev. Biol. 2008, 52, 527–533. [Google Scholar] [CrossRef]
- Murata, K.; Sugiyama, H.; Yasumasu, S.; Iuchi, I.; Yasumasu, I.; Yamagami, K. Cloning of cDNA and estrogen-induced hepatic gene expression for choriogenin H, a precursor protein of the fish egg envelope (chorion). Proc. Natl. Acad. Sci. USA 1997, 94, 2050–2055. [Google Scholar]
- Chang, Y.S.; Hsu, C.C.; Wang, S.C.; Tsao, C.C.; Huang, F.L. Molecular cloning, structural analysis, and expression of carp ZP2 gene. Mol. Reprod. Dev. 1997, 46, 258–267. [Google Scholar] [CrossRef]
- Mold, D.E.; Kim, I.F.; Tsai, C.M.; Lee, D.; Chang, C.Y.; Huang, R.C. Cluster of genes encoding the major egg envelope protein of zebrafish. Mol. Reprod. Dev. 2001, 58, 4–14. [Google Scholar] [CrossRef]
- Kanamori, A. Systematic identification of genes expressed during early oogenesis in medaka. Mol. Reprod. Dev. 2000, 55, 31–36. [Google Scholar] [CrossRef]
- Hyllner, S.J.; Westerlund, L.; Olsson, P.E.; Schopen, A. Cloning of rainbow trout egg envelope proteins: Members of a unique group of structural proteins. Biol. Reprod. 2001, 64, 805–811. [Google Scholar] [CrossRef]
- Lee, C.; Jeon, S.H.; Na, J.G.; Park, K. Sequence analysis of choriogenin H gene of medaka (Oryzias latipes) and mRNA expression. Environ. Toxicol. Chem. 2002, 21, 1709–1714. [Google Scholar]
- Miller, D.J.; Ax, R.L. Carbohydrates and fertilization in animals. Mol. Reprod. Dev. 1990, 26, 184–198. [Google Scholar] [CrossRef]
- Kanamori, A.; Inoue, S.; Iwasaki, M.; Kitajima, K.; Kawai, G.; Yokoyama, S.; Inoue, Y. Deaminated neuraminic acid-rich glycoprotein of rainbow trout egg vitelline envelope. Occurrence of a novel alpha-2,8-linked oligo(deaminated neuraminic acid) structure in O-linked glycan chains. J. Biol. Chem. 1990, 265, 21811–21819. [Google Scholar]
- Tezuka, T.; Taguchi, T.; Kanamori, A.; Muto, Y.; Kitajima, K.; Inoue, Y.; Inoue, S. Identification and structural determination of the KDN-containing N-linked glycan chains consisting of bi- and triantennary complex-type units of KDN-glycoprotein previously isolated from rainbow trout vitelline envelopes. Biochemistry 1994, 33, 6495–6502. [Google Scholar] [CrossRef]
- Inoue, S.; Inoue, Y. Fish Glycoprotein. In Glycoprotein II; Montreuil, J., Vliegenthart, J.F.G., Schachter, H., Eds.; Elsevier Science BV: Amsterdam, The Netherlands, 1997; Volume 29, pp. 143–161. [Google Scholar]
- Pillai, M.C.; Clark, W.H., Jr. Development of cortical vesicles in Sicyonia ingentis ova: Their heterogeneity and role in elaboration of the hatching envelope. Mol. Reprod. Dev. 1990, 26, 78–89. [Google Scholar] [CrossRef]
- Dupré, E.; Gòmez, D.; Araya, A.; Gallardo, C. Role of egg surface glycoconjugate in the fertilization of the rock shrimp Rhynchocinetes typus (Milne-Edwards, 1837). Lat. Am. J. Aquat. Res. 2012, 40, 22–29. [Google Scholar] [CrossRef]
- Litscher, E.; Honegger, T.G. Glycoprotein constituents of the vitelline coat of Phallusia mammillata (Ascidiacea) with fertilization inhibiting activity. Dev. Biol. 1991, 148, 536–551. [Google Scholar] [CrossRef]
- Baginski, T.; Hirohashi, N.; Hoshi, M. Sulfated O-linked glycans of the vitelline coat as ligands in gamete interaction in the ascidian, Halocynthia roretzi. Dev. Growth Differ. 1999, 41, 357–364. [Google Scholar]
- De Santis, R.; Pinto, M.R. Gamete Interaction in Ascidians: Sperm Binding and Penetration through the Vitelline Coat. In Mechanism of Fertilization: Plants to Humans; Dale, B., Ed.; Springer-Verlag: Berlin, Germany, 1990; pp. 297–304. [Google Scholar]
- Rosati, F.; Cotelli, F.; de Santis, R.; Monroy, A.; Pinto, M.R. Synthesis of fucosyl-containing glycoproteins of the vitelline coat in oocytes of Ciona intestinalis (Ascidia). Proc. Natl. Acad. Sci. USA 1982, 79, 1908–1911. [Google Scholar]
- SeGall, G.K.; Lennarz, W.J. Chemical characterization of the component of the jelly coat from sea urchin eggs responsible for induction of the acrosome reaction. Dev. Biol. 1979, 71, 33–48. [Google Scholar] [CrossRef]
- Shimizu, T.; Kinoh, H.; Yamaguchi, M.; Suzuki, N. Purification and characterization of the egg jelly macromolecules, sialoglycoprotein and fucose sulfate glycoconjugate, of the sea urchin Hemicentrotus Pulcherrimus. Dev. Growth Differ. 1990, 32, 473–487. [Google Scholar]
- Kitazume, S.; Kitajima, K.; Inoue, S.; Troy, F.A., II.; Cho, J.W.; Lennarz, W.J.; Inoue, Y. Identification of polysialic acid-containing glycoprotein in the jelly coat of sea urchin eggs. Occurrence of a novel type of polysialic acid structure. J. Biol. Chem. 1994, 269, 22712–22718. [Google Scholar]
- Hirohashi, N.; Kamei, N.; Kubo, H.; Sawada, H.; Matsumoto, M.; Hoshi, M. Egg and sperm recognition systems during fertilization. Dev. Growth Differ. 2008, 50, S221–S238. [Google Scholar] [CrossRef]
- Mulloy, B.; Ribeiro, A.C.; Alves, A.P.; Vieira, R.P.; Mourao, P.A. Sulfated fucans from echinoderms have a regular tetrasaccharide repeating unit defined by specific patterns of sulfation at the 0–2 and 0–4 positions. J. Biol. Chem. 1994, 269, 22113–22123. [Google Scholar]
- Alves, A.P.; Mulloy, B.; Diniz, J.A.; Mourao, P.A. Sulfated polysaccharides from the egg jelly layer are species-specific inducers of acrosomal reaction in sperms of sea urchins. J. Biol. Chem. 1997, 272, 6965–6971. [Google Scholar] [CrossRef]
- Vacquier, V.D.; Moy, G.W. The fucose sulfate polymer of egg jelly binds to sperm REJ and is the inducer of the sea urchin sperm acrosome reaction. Dev. Biol. 1997, 192, 125–135. [Google Scholar] [CrossRef]
- Alves, A.P.; Mulloy, B.; Moy, G.W.; Vacquier, V.D.; Mourao, P.A. Females of the sea urchin Strongylocentrotus purpuratus differ in the structures of their egg jelly sulfated fucans. Glycobiology 1998, 8, 939–946. [Google Scholar] [CrossRef]
- Vilela-Silva, A.C.; Castro, M.O.; Valente, A.P.; Biermann, C.H.; Mourao, P.A. Sulfated fucans from the egg jellies of the closely related sea urchins Strongylocentrotus droebachiensis and Strongylocentrotus pallidus ensure species-specific fertilization. J. Biol. Chem. 2002, 277, 379–387. [Google Scholar]
- Endo, T.; Hoshi, M.; Endo, S.; Arata, Y.; Kobata, A. Structures of the sugar chains of a major glycoprotein present in the egg jelly coat of a starfish, Asterias amurensis. Arch. Biochem. Biophys. 1987, 252, 105–112. [Google Scholar] [CrossRef]
- Hoshi, M.; Nishigaki, T.; Ushiyama, A.; Okinaga, T.; Chiba, K.; Matsumoto, M. Egg-jelly signal molecules for triggering the acrosome reaction in starfish spermatozoa. Int. J. Dev. Biol. 1994, 38, 167–174. [Google Scholar]
- Uno, Y.; Hoshi, M. Separation of the sperm agglutinin and the acrosome reaction-inducing substance in egg jelly of starfish. Science 1978, 200, 58–59. [Google Scholar]
- Naruse, M.; Suetomo, H.; Matsubara, T.; Sato, T.; Yanagawa, H.; Hoshi, M.; Matsumoto, M. Acrosome reaction-related steroidal saponin, Co-ARIS, from the starfish induces structural changes in microdomains. Dev. Biol. 2010, 347, 147–153. [Google Scholar] [CrossRef]
- Koyota, S.; Wimalasiri, K.M.; Hoshi, M. Structure of the main saccharide chain in the acrosome reaction-inducing substance of the starfish, Asterias amurensis. J. Biol. Chem. 1997, 272, 10372–10376. [Google Scholar] [CrossRef]
- Gunaratne, H.M.; Yamagaki, T.; Matsumoto, M.; Hoshi, M. Biochemical characterization of inner sugar chains of acrosome reaction-inducing substance in jelly coat of starfish eggs. Glycobiology 2003, 13, 567–580. [Google Scholar] [CrossRef]
- Hoshi, M.; Moriyama, H.; Matsumoto, M. Structure of acrosome reaction-inducing substance in the jelly coat of starfish eggs: A mini review. Biochem. Biophys. Res. Commun. 2012, 425, 595–598. [Google Scholar] [CrossRef]
- Suphamungmee, W.; Chansela, P.; Weerachatyanukul, W.; Poomtong, T.; Vanichviriyakit, R.; Sobhon, P. Ultrastructure, composition, and possible roles of the egg coats in Haliotis asinin. J. Shellfish Res. 2010, 29, 687–697. [Google Scholar] [CrossRef]
- Shigekawa, K.; Clark, W.H., Jr. Spermiogenesis in the marine shrimp, Sicyonia ingentis. Dev. Growth Differ. 1986, 28, 95–112. [Google Scholar]
- Nagao, J.; Munehara, H. Annual cycle of testicular maturation in the helmet crab Telmessus cheiragonus. Fish. Sci. 2003, 69, 1200–1208. [Google Scholar] [CrossRef]
- Okumura, T.; Hara, M. Androgenic gland cell structure and spermatogenesis during the molt cycle and correlation to morphotypic differentiation in the giant freshwater prawn, Macrobrachium rosenbergii. Zool. Sci. 2004, 21, 621–628. [Google Scholar] [CrossRef]
- Garcia, T.M.; Silva, J.R.F. Testis and vas deferens morphology of the red-clawed mangrove tree crab (Goniopsis cruentata) (Latreille, 1803). Braz. Arch. Biol. Technol. 2006, 49, 339–345. [Google Scholar]
- Jones, R.; Brown, C.R.; Lancaster, R.T. Carbohydrate-binding properties of boar sperm proacrosin and assessment of its role in sperm-egg recognition and adhesion during fertilization. Dev. Genes Evol. 1988, 102, 781–779. [Google Scholar]
- Anakwe, O.O.; Gerton, G.L. Acrosome biogenesis begins during meiosis: Evidence from the synthesis and distribution of an acrosomal glycoprotein, acrogranin, during guinea pig spermatogenesis. Biol. Reprod. 1990, 42, 317–328. [Google Scholar] [CrossRef]
- Martinez-Menarguez, J.A.; Ballesta, J.; Aviles, M.; Castells, M.T.; Madrid, J.F. Cytochemical characterization of glycoproteins in the developing acrosome of rats. An ultrastructural study using lectin histochemistry, enzymes and chemical deglycosylation. Histochemistry 1992, 97, 439–449. [Google Scholar] [CrossRef]
- Pratt, S.A.; Scully, N.F.; Shur, B.D. Cell surface beta 1,4 galactosyltransferase on primary spermatocytes facilitates their initial adhesion to Sertoli cells in vitro. Biol. Reprod. 1993, 49, 470–482. [Google Scholar] [CrossRef]
- Raychoudhury, S.S.; Millette, C.F. Multiple fucosyltransferases and their carbohydrate ligands are involved in spermatogenic cell-Sertoli cell adhesion in vitro in rats. Biol. Reprod. 1997, 56, 1268–1273. [Google Scholar]
- Schulz, R.W.; Miura, T. Spermatogenesis and its endocrine regulation. Fish. Physiol. Biochem. 2002, 26, 43–56. [Google Scholar] [CrossRef]
- Nobrega, R.H.; Batlouni, S.R.; Franca, L.R. An overview of functional and stereological evaluation of spermatogenesis and germ cell transplantation in fish. Fish. Physiol. Biochem. 2009, 35, 197–206. [Google Scholar] [CrossRef]
- Parenti, L.R.; Grier, H.J. Evolution and phylogeny of gonad morphology in bony fishes. Integr. Comp. Biol. 2004, 44, 333–348. [Google Scholar] [CrossRef]
- Desantis, S.; Zizza, S.; Garcia-Lopez, A.; Sciscioli, V.; Mananos, E.; de Metrio, V.G.; Sarasquete, C. Lectin-binding pattern of Senegalese sole Solea senegalensis (Kaup) testis. Histol. Histopathol. 2010, 25, 205–216. [Google Scholar]
- Liguoro, A.; Prisco, M.; Mennella, C.; Ricchiari, L.; Angelini, F.; Andreuccetti, P. Distribution of terminal sugar residues in the testis of the spotted ray Torpedo marmorata. Mol. Reprod. Dev. 2004, 68, 524–530. [Google Scholar] [CrossRef]
- Tokalov, S.V.; Gutzeit, H.O. Lectin-binding pattern as tool to identify and enrich specific primary testis cells of the tilapia (Oreochromis niloticus) and medaka (Oryzias latipes). J. Exp. Zool. Part A 2007, 308, 127–138. [Google Scholar]
- Varki, A.; Cummings, R.; Esko, J.; Freeze, H.; Hart, G.; Marth, J. Nuclear and Cytoplasmic Glycosylation. In Essentials of Glycobiology; Varki, A., Cummings, R., Esko, J., Freeze, H., Hart, G., Marth, J., Eds.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1999; pp. 171–183. [Google Scholar]
- Martinez-Menarguez, J.A.; Aviles, M.; Madrid, J.F.; Castells, M.T.; Ballesta, J. Glycosylation in Golgi apparatus of early spermatids of rat. A high resolution lectin cytochemical study. Eur. J. Cell Biol. 1993, 61, 21–33. [Google Scholar]
- Unuma, T.; Yamamoto, T.; Akiyama, T.; Shiraishi, M.; Ohta, H. Quantitative changes in yolk protein and other components in the ovary and testis of the sea urchin Pseudocentrotus depressus. J. Exp. Biol. 2003, 206, 365–372. [Google Scholar] [CrossRef]
- Verhey, C.A.; Moyer, F.H. Fine structural changes during sea urchin oogenesis. J. Exp. Zool. 1967, 162, 195–225. [Google Scholar] [CrossRef]
- Marsh, A.G.; Watts, S.A. Energy metabolism and gonad development. Dev. Aquac. Fish. Sci. 2001, 32, 27–42. [Google Scholar] [CrossRef]
- Miyata, S.; Sato, C.; Kitamura, S.; Toriyama, M.; Kitajima, K. A major flagellum sialoglycoprotein in sea urchin sperm contains a novel polysialic acid, an alpha2,9-linked poly-N-acetylneuraminic acid chain, capped by an 8-O-sulfated sialic acid residue. Glycobiology 2004, 14, 827–840. [Google Scholar] [CrossRef]
- Miyata, S.; Sato, C.; Kumita, H.; Toriyama, M.; Vacquier, V.D.; Kitajima, K. Flagellasialin: A novel sulfated alpha2,9-linked polysialic acid glycoprotein of sea urchin sperm flagella. Glycobiology 2006, 16, 1229–1241. [Google Scholar] [CrossRef]
- Kambara, Y.; Shiba, K.; Yoshida, M.; Sato, C.; Kitajima, K.; Shingyoji, C. Mechanism regulating Ca2+-dependent mechanosensory behaviour in sea urchin spermatozoa. Cell Struct. Funct. 2011, 36, 69–82. [Google Scholar] [CrossRef]
- Ijuin, T.; Kitajima, K.; Song, Y.; Kitazume, S.; Inoue, S.; Haslam, S.M.; Morris, H.R.; Dell, A.; Inoue, Y. Isolation and identification of novel sulfated and nonsulfated oligosialyl glycosphingolipids from sea urchin sperm. Glycoconj. J. 1996, 13, 401–413. [Google Scholar] [CrossRef]
- Miyata, S.; Yamakawa, N.; Toriyama, M.; Sato, C.; Kitajima, K. Co-expression of two distinct polysialic acids, alpha2,8- and alpha2,9-linked polymers of N-acetylneuraminic acid, in distinct glycoproteins and glycolipids in sea urchin sperm. Glycobiology 2011, 21, 1596–1605. [Google Scholar] [CrossRef]
- Desantis, S.; Labate, M.; Maiorano, P.; Tursi, A. An ultrastructural and histochemical study of the germinal cells contained in hemispermatophores of males of the Aristaeomorpha foliacea (Risso, 1827). Hydrobiologia 2006, 557, 41–49. [Google Scholar] [CrossRef]
- Subramonian, T. Spermatophore formation in two intertidal crabs Albunea symnista and Emerita asiatica (Decapoda: Anomura). Biol. Bull. 1984, 166, 78–95. [Google Scholar] [CrossRef]
- Radha, T.; Subramonian, T. Origin and nature of spermatophoric mass of spiny lobster Panulirus homarus. Mar. Biol. 1985, 86, 13–19. [Google Scholar] [CrossRef]
- Yufeng, W.; Nanshan, D.; Wei, L. A histochemical study on the male reproductive system of Macrobrachium rosenbergii. J. Northwest Atl. Fish. Sci. 1997, 4, 13–16. [Google Scholar]
- Sasikala, S.L.; Subramoniam, T. On the occurrence of acid mucopolysaccharides in the spermatophores of two marine prawns, Penaeus indiens (Milne-Edwards) and Metapenaeus monoceros (Fabricius) (Crustacea: Macrura). J. Exp. Mar. Biol. Ecol. 1987, 113, 145–153. [Google Scholar] [CrossRef]
- Subramonian, T. Chemical Composition of Spermatophores in Decapod Crustaceans. In Crustacean Sexual Biology; Bauer, R.T., Martin, J.A., Eds.; Columbia University Press: New York, NY, USA, 1991; pp. 308–321. [Google Scholar]
- Desantis, S.; Labate, M.; Cirillo, F.; Labate, G.M. Testicular activity and sperm glycoproteins in giant red shrimp (Aristaeomorpha foliacea). J. Northwest Atl. Fish. Sci. 2003, 31, 205–212. [Google Scholar]
- Yanagimachi, R. Fertility of mammalian spermatozoa: Its development and relativity. Zygote 1994, 2, 371–372. [Google Scholar]
- Snell, W.J.; White, J.M. The molecules of mammalian fertilization. Cell 1996, 85, 629–637. [Google Scholar] [CrossRef]
- Vacquier, V.D. Evolution of gamete recognition proteins. Science 1998, 281, 1995–1998. [Google Scholar] [CrossRef]
- Tosti, E. Sperm activation in species with external fertilisation. Zygote 1994, 2, 359–361. [Google Scholar]
- Wassarman, P.M.; Litscher, E.S. Towards the molecular basis of sperm and egg interaction during mammalian fertilization. Cells Tissues Organs 2001, 168, 36–45. [Google Scholar] [CrossRef]
- Monroy, A.; Rosati, F. A comparative analysis of sperm-egg interaction. Gamete Res. 1983, 7, 85–102. [Google Scholar] [CrossRef]
- Wassarman, P.M. The biology and chemistry of fertilization. Science 1987, 235, 553–560. [Google Scholar]
- Moy, G.W.; Mendoza, L.M.; Schulz, J.R.; Swanson, W.J.; Glabe, C.G.; Vacquier, V.D. The sea urchin sperm receptor for egg jelly is a modular protein with extensive homology to the human polycystic kidney disease protein, PKD1. J. Cell Biol. 1996, 133, 809–817. [Google Scholar] [CrossRef]
- Mengerink, K.J.; Vacquier, V.D. Glycobiology of sperm-egg interactions in deuterostomes. Glycobiology 2001, 11, 37R–43R. [Google Scholar] [CrossRef]
- Vacquier, V.D.; Swanson, W.J.; Hellberg, M.E. What have we learned about sea urchin sperm bindin? Dev. Growth Differ. 1995, 37, 1–10. [Google Scholar]
- Ohlendieck, K.; Partin, J.S.; Stears, R.L.; Lennarz, W.J. Developmental expression of the sea urchin egg receptor for sperm. Dev. Biol. 1994, 165, 53–62. [Google Scholar] [CrossRef]
- Dhume, S.T.; Lennarz, W.J. The involvement of O-linked oligosaccharide chains of the sea urchin egg receptor for sperm in fertilization. Glycobiology 1995, 5, 11–17. [Google Scholar] [CrossRef]
- Hirohashi, N.; Lennarz, W.J. Role of a vitelline layer-associated 350 kDa glycoprotein in controlling species-specific gamete interaction in the sea urchin. Dev. Growth Differ. 2001, 43, 247–255. [Google Scholar] [CrossRef]
- Hoshi, M.; Matsui, T.; Nishiyama, I.; Amano, T.; Okita, Y. Physiological inducers of the acrosome reaction. Cell Differ. Dev. 1988, 25 Suppl., 19–24. [Google Scholar] [CrossRef]
- Longo, F.J.; Ushiyama, A.; Chiba, K.; Hoshi, M. Ultrastructural localization of acrosome reaction-inducing substance (ARIS) on sperm of the starfish Asterias amurensis. Mol. Reprod. Dev. 1995, 41, 91–99. [Google Scholar] [CrossRef]
- Ushiyama, A.; Araki, T.; Chiba, K.; Hoshi, M. Specific binding of acrosome-reaction-inducing substance to the head of starfish spermatozoa. Zygote 1993, 1, 121–127. [Google Scholar]
- Hoshi, M.; Amano, T.; Okita, Y.; Okinaga, T.; Matsui, T. Egg signals for triggering the acrosome reaction in starfish spermatozoa. J. Reprod. Fertil. 1990, 42, 23–31. [Google Scholar]
- Nishiyama, I.; Matsui, T.; Fujimoto, Y.; Ikekawa, N.; Hoshi, M. Correlation between the molecular structure and the biological activity of Co-ARIS, a cofactor for acrosome reaction-inducing substance. Dev. Growth Differ. 1987, 29, 171–176. [Google Scholar]
- Nishigaki, T.; Chiba, K.; Miki, W.; Hoshi, M. Structure and function of asterosaps, sperm-activating peptides from the jelly coat of starfish eggs. Zygote 1996, 4, 237–245. [Google Scholar]
- Nishigaki, T.; Chiba, K.; Hoshi, M. A 130-kDa membrane protein of sperm flagella is the receptor for asterosaps, sperm-activating peptides of starfish Asterias amurensis. Dev. Biol. 2000, 219, 154–162. [Google Scholar] [CrossRef]
- Matsumoto, M.; Hirata, J.; Hirohashi, N.; Hoshi, M. Sperm-egg binding mediated by sperm alpha-D-fucosidase in the ascidian, Halocynthia roretzi. Zool. Sci. 2002, 19, 43–48. [Google Scholar] [CrossRef]
- Matsumoto, M.; Kawase, O.; Islam, M.S.; Naruse, M.; Watanabe, S.N.; Ishikawa, R.; Hoshi, M. Regulation of the starfish sperm acrosome reaction by cGMP, pH, cAMP and Ca2+. Int. J. Dev. Biol. 2008, 52, 523–526. [Google Scholar] [CrossRef]
- Swanson, W.J.; Vacquier, V.D. The abalone egg vitelline envelope receptor for sperm lysin is a giant multivalent molecule. Proc. Natl. Acad. Sci. USA 1997, 94, 6724–6729. [Google Scholar] [CrossRef]
- Vacquier, V.D.; Carner, K.R.; Stout, C.D. Species-specific sequences of abalone lysin, the sperm protein that creates a hole in the egg envelope. Proc. Natl. Acad. Sci. USA 1990, 87, 5792–5796. [Google Scholar] [CrossRef]
- Kresge, N.; Vacquier, V.D.; Stout, C.D. Abalone lysin: The dissolving and evolving sperm protein. Bioessays 2001, 23, 95–103. [Google Scholar]
- Swanson, W.J.; Aquadro, C.F.; Vacquier, V.D. Polymorphism in abalone fertilization proteins is consistent with the neutral evolution of the egg’s receptor for lysin (VERL) and positive darwinian selection of sperm lysin. Mol. Biol. Evol. 2001, 18, 376–383. [Google Scholar] [CrossRef]
- Hoshi, M.; de Santis, R.; Pinto, M.R.; Cotelli, F.; Rosati, F. Sperm glycosidases as mediators of sperm-egg binding in the ascidians. Zool. Sci. 1985, 2, 65–69. [Google Scholar]
- Hoshi, M. Sperm glycosidase as a plausible mediator of sperm binding to the vitelline envelope in Ascidians. Adv. Exp. Med. Biol. 1986, 207, 251–260. [Google Scholar]
- Godknecht, A.; Honegger, T.G. Isolation, characterization, and localization of a sperm-bound N-acetylglucosaminidase that is indispensable for fertilization in the ascidian, Phallusia mammilla. Dev. Biol. 1991, 143, 398–407. [Google Scholar] [CrossRef]
- Godknecht, A.J.; Honegger, T.G. Specific inhibition of sperm β-N-acetylglucosaminidase by the synthetic inhibitor N-acetylglucosaminono-1,5-lactone O-(phenylcarbamoyl)oxime inhibits fertilization in the ascidian, Phallusia mammillata. Dev. Growth Differ. 1995, 37, 183–189. [Google Scholar]
- Xie, M.; Honegger, T.G. Ultrastructural investigations on sperm penetration and gamete fusion in the ascidians Boltenia villosa and Phallusia mammillata. Mar. Biol. 1993 116, 117–127.
- Hoshi, M.; de Santis, R.; Pinto, M.R.; Cotelli, F.; Rosati, F. Is Sperm L-fucosidase Responsible for Sperm-Egg Binding in Ciona Intestinalis. In The Sperm Cell; André, J., Ed.; Martinus Nijhoff: Leiden, NL, USA, 1983; pp. 107–110. [Google Scholar]
- Fukumoto, M. Fertilization in ascidians apical processes and gamete fusion in Ciona intestinalis spermatozoa. J. Cell Sci. 1988, 89, l89–l96. [Google Scholar]
- Fukumoto, M. Morphological aspects of ascidian fertilization: Acrosome reaction, apical processes and gamete fusion in Ciona intestinalis. Invertebr. Reprod. Dev. 1990, 17, 147–154. [Google Scholar] [CrossRef]
- Fukumoto, M.; Numakunai, T. Morphological aspects of fertilization in Halocynthia roretzi (Ascidiacea, Tunicata). J. Struct. Biol. 1995, 114, 157–166. [Google Scholar] [CrossRef]
- Sawada, H.; Sakai, N.; Abe, Y.; Tanaka, E.; Takahashi, Y.; Fujino, J.; Kodama, E.; Takizawa, S.; Yokosawa, H. Extracellular ubiquitination and proteasome-mediated degradation of the ascidian sperm receptor. Proc. Natl. Acad. Sci. USA 2002, 99, 1223–1228. [Google Scholar]
- Sawada, H.; Yokosawa, H.; Ishii, S. Purification and characterization of two types of trypsin-like enzymes from sperm of the ascidian (Prochordata) Halocynthia roretzi. Evidence for the presence of spermosin, a novel acrosin-like enzyme. J. Biol. Chem. 1984, 259, 2900–2904. [Google Scholar]
- Lambert, C.C.; Someno, T.; Sawada, H. Sperm surface proteases in ascidian fertilization. J. Exp. Zool. 2002, 292, 88–95. [Google Scholar] [CrossRef]
- Dupre, E.M.; Barros, C. In vitro fertilization of the rock shrimp, Rhynchocinetes typus (Decapoda, Caridea): A review. Biol. Res. 2011, 44, 125–133. [Google Scholar] [CrossRef]
- Barros, C.; Dupré, E.; Viveros, L. Sperm-egg interactions in the shrimp Rhynchocinetes typus. Gamete Res. 1986, 14, 171–180. [Google Scholar] [CrossRef]
- Rios, M.; Barros, C. Trypsin-like enzymes during fertilization in the shrimp Rhynchocinetes typus. Mol. Reprod. Dev. 1997, 46, 581–586. [Google Scholar] [CrossRef]
- Bustamante, E.; Palomino, J.; Amoroso, A.; Moreno, R.D.; Barros, C. Purification and biochemical characterization of a trypsin-like enzyme present in the sperm of the rock shrimp, Rhynchocinetes typus. Invertebr. Reprod. Dev. 2001, 39, 175–181. [Google Scholar] [CrossRef]
- Gilbert, S.F. Developmental Biology, 5th ed; Sinauer Associates: Sunderland, MA, USA, 1997. [Google Scholar]
- Foltz, K.R.; Shilling, F.M. Receptor-mediated signal transduction and egg activation. Zygote 1993, 1, 276–279. [Google Scholar]
- Hirohashi, N.; Lennarz, W.J. Sperm-egg binding in the sea urchin: A high level of intracellular ATP stabilizes sperm attachment to the egg receptor. Dev. Biol. 1998, 201, 270–279. [Google Scholar] [CrossRef]
- Kamei, N.; Glabe, C.G. The species-specific egg receptor for sea urchin sperm adhesion is EBR1, a novel ADAMTS protein. Genes Dev. 2003, 17, 2502–2507. [Google Scholar] [CrossRef]
- Kamei, N.; Swanson, W.J.; Glabe, C.G. A rapidly diverging EGF protein regulates species-specific signal transduction in early sea urchin development. Dev. Biol. 2000, 225, 267–276. [Google Scholar] [CrossRef]
- Hagstrom, B.E. Further studies on cross fertilization in sea urchins. Exp. Cell Res. 1956, 11, 507–510. [Google Scholar] [CrossRef]
- Adelson, D.L.; Alliegro, M.C.; McClay, D.R. On the ultrastructure of hyalin, a cell adhesion protein of the sea urchin embryo extracellular matrix. J. Cell Biol. 1992, 116, 1283–1289. [Google Scholar] [CrossRef]
- Shapiro, B.M.; Somers, C.; Weidman, P.J. Extracellular Remodeling during Fertilization. In Cell Biology of Fertilization; Schatten, H., Schatten, G., Eds.; Academic Press: San Diego, CA, USA, 1989; pp. 251–276. [Google Scholar]
- Somers, C.E.; Shapiro, B.M. Functional domains of proteoliaisin, the adhesive protein that orchestrates fertilization envelope assembly. J. Biol. Chem. 1991, 266, 16870–16875. [Google Scholar]
- Nomura, K.; Suzuki, N. Sea urchin ovoperoxidase: Solubilization and isolation from the fertilization envelope, some structural and functional properties, and degradation by hatching enzyme. Arch. Biochem. Biophys. 1995, 319, 525–534. [Google Scholar] [CrossRef]
- Inoue, S.; Inoue, Y. Fertilization (activation)-induced 200- to 9-kDa depolymerization of polysialoglycoprotein, a distinct component of cortical alveoli of rainbow trout eggs. J. Biol. Chem. 1986, 261, 5256–5261. [Google Scholar]
- Inoue, S.; Kitajima, K.; Inoue, Y.; Kudo, S. Localization of polysialoglycoprotein as a major glycoprotein component in cortical alveoli of the unfertilized eggs of Salmo gairdneri. Dev. Biol. 1987, 123, 442–454. [Google Scholar] [CrossRef]
- Laale, H.W. The perivitelline space and egg envelopes of bony fishes: A review. Copeia 1980, 1980, 210–226. [Google Scholar] [CrossRef]
- Rudy, P.P., Jr.; Potts, W.T. Sodium balance in the eggs of the Atlantic salmon, Salmo salar. J. Exp. Biol. 1969, 50, 239–246. [Google Scholar]
- Eddy, F.B. Osmotic properties of the perivitelline fluid and some properties of the chorion of Atlantic salmon eggs (Salmo salar). J. Zool. 1974, 174, 237–243. [Google Scholar] [CrossRef]
- Peterson, R.H.; Martin-Robichaud, D.J. Rates of ionic diffusion across the egg chorion of Atlantic salmon (Salmo salar). Physiol. Zool. 1993, 66, 289–306. [Google Scholar]
- Li, X.; Jenssen, E.; Fyhn, H.J. Effects of salinity on egg swelling in Atlantic salmon (Salmo salar). Aquaculture 1989, 76, 317–334. [Google Scholar] [CrossRef]
- Kudo, S. Fertilization, cortical reaction, polyspermy-preventing and anti-microbial mechanisms in fish eggs. Bull. Inst. Zool. Acad. Sci. Monogr. 1991, 16, 313–340. [Google Scholar]
- Kobayashi, W.; Yamamoto, T.S. Factors inducing closure of the micropylar canal in the chum salmon egg. J. Fish. Biol. 1993, 42, 385–394. [Google Scholar] [CrossRef]
- Kudo, S.; Teshima, C. Assembly in vitro of vitelline envelope components induced by a cortical alveolus sialoglycoprotein of eggs of the fish Tribolodon hakonensis. Zygote 1998, 6, 193–201. [Google Scholar] [CrossRef]
- Oppen-Berntsen, D.O.; Helvik, J.V.; Walther, B.T. The major structural proteins of cod (Gadus morhua) eggshells and protein crosslinking during teleost egg hardening. Dev. Biol. 1990, 137, 258–265. [Google Scholar] [CrossRef]
- Koyanagi, R.; Honegger, T.G. Molecular cloning and sequence analysis of an ascidian egg beta-N-acetylhexosaminidase with a potential role in fertilization. Dev. Growth Differ. 2003, 45, 209–218. [Google Scholar] [CrossRef]
- Lambert, C.C. Fertilization-induced modification of chorion N-acetylglucosamine groups blocks polyspermy in ascidian eggs. Dev. Biol. 1986, 116, 168–173. [Google Scholar] [CrossRef]
- Lambert, C.C. Ascidian eggs release glycosidase activity which aids in the block against polyspermy. Development 1989, 105, 415–420. [Google Scholar]
- Schneider, E.G.; Nguyen, H.T.; Lennarz, W.J. The effect of tunicamycin, an inhibitor of protein glycosylation, on embryonic development in the sea urchin. J. Biol. Chem. 1978, 253, 2348–2355. [Google Scholar]
- Heifetz, A.; Lennarz, W.J. Biosynthesis of N-glycosidically linked glycoproteins during gastrulation of sea urchin embryos. J. Biol. Chem. 1979, 254, 6119–6127. [Google Scholar]
- Lennarz, W.J. Glycoprotein synthesis and embryonic development. CRC Crit. Rev. Biochem. 1983, 14, 257–272. [Google Scholar] [CrossRef]
- Carson, D.D.; Lennarz, W.J. Inhibition of polyisoprenoid and glycoprotein biosynthesis causes abnormal embryonic development. Proc. Natl. Acad. Sci. USA 1979, 76, 5709–5713. [Google Scholar]
- Carson, D.D.; Lennarz, W.J. Relationship of dolichol synthesis to glycoprotein synthesis during embryonic development. J. Biol. Chem. 1981, 256, 4679–4686. [Google Scholar]
- Carson, D.D.; Farach, M.C.; Earles, D.S.; Decker, G.L.; Lennarz, W.J. A monoclonal antibody inhibits calcium accumulation and skeleton formation in cultured embryonic cells of the sea urchin. Cell 1985, 41, 639–648. [Google Scholar] [CrossRef]
- Farach, M.C.; Valdizan, M.; Park, H.R.; Decker, G.L.; Lennarz, W.J. Developmental expression of a cell-surface protein involved in calcium uptake and skeleton formation in sea urchin embryos. Dev. Biol. 1987, 122, 320–331. [Google Scholar] [CrossRef]
- Farach-Carson, M.C.; Carson, D.D.; Collier, J.L.; Lennarz, W.J.; Park, H.R.; Wright, G.C. A calcium-binding, asparagine-linked oligosaccharide is involved in skeleton formation in the sea urchin embryo. J. Cell Biol. 1989, 109, 1289–1299. [Google Scholar] [CrossRef]
- Decker, G.L.; Valdizan, M.C.; Wessel, G.M.; Lennarz, W.J. Developmental distribution of a cell surface glycoprotein in the sea urchin Strongylocentrotus purpuratus. Dev. Biol. 1988, 129, 339–349. [Google Scholar] [CrossRef]
- Anstrom, J.A.; Chin, J.E.; Leaf, D.S.; Parks, A.L.; Raff, R.A. Localization and expression of msp130, a primary mesenchyme lineage-specific cell surface protein in the sea urchin embryo. Development 1987, 101, 255–265. [Google Scholar]
- McClay, D.R.; Matranga, V.; Wessel, G. Expression and Appearance of Germ-Layer Specific Antigens on the Surface of Embryonic Sea Urchin Cells. In The Cellular and Molecular Biology of Invertebrate Development; Sawyer, R.H., Showman, R.M., Eds.; University of South Carolina Press: Columbia, SC, USA, 1985; pp. 171–186. [Google Scholar]
- Kabakoff, B.; Lennarz, W.J. Inhibition of glycoprotein processing blocks assembly of spicules during development of the sea urchin embryo. J. Cell Biol. 1990, 111, 391–400. [Google Scholar] [CrossRef]
- Hoshi, M.; Nagai, Y. Biochemistry of mucolipids of sea urchin gametes and embryos. 3. Mucolipids during early development. Jpn. J. Exp. Med. 1970, 40, 361–365. [Google Scholar]
- Cheresh, D.A.; Klier, F.G. Disialoganglioside GD2 distributes preferentially into substrate-associated microprocesses on human melanoma cells during their attachment to fibronectin. J. Cell Biol. 1986, 102, 1887–1897. [Google Scholar] [CrossRef]
- Spiegel, S.; Yamada, K.M.; Hom, B.E.; Moss, J.; Fishman, P.H. Fluorescent gangliosides as probes for the retention and organization of fibronectin by ganglioside-deficient mouse cells. J. Cell Biol. 1985, 100, 721–726. [Google Scholar] [CrossRef]
- Wessel, G.M.; Marchase, R.B.; McClay, D.R. Ontogeny of the basal lamina in the sea urchin embryo. Dev. Biol. 1984, 103, 235–245. [Google Scholar] [CrossRef]
- Jackson, R.L.; Busch, S.J.; Cardin, A.D. Glycosaminoglycans: Molecular properties, protein interactions, and role in physiological processes. Physiol. Rev. 1991, 71, 481–539. [Google Scholar]
- Vilela-Silva, A.C.; Werneck, C.C.; Valente, A.P.; Vacquier, V.D.; Mourao, P.A. Embryos of the sea urchin Strongylocentrotus purpuratus synthesize a dermatan sulfate enriched in 4-O- and 6-O-disulfated galactosamine units. Glycobiology 2001, 11, 433–440. [Google Scholar] [CrossRef]
- Oguri, K.; Yamagata, T. Appearance of a proteoglycan in developing sea urchin embryos. Biochim. Biophys. Acta 1978, 541, 385–393. [Google Scholar] [CrossRef]
- Kinoshita, S.; Yoshii, K. The role of proteoglycan synthesis in the development of sea urchins. II. The effect of administration of exogenous proteoglycan. Exp. Cell Res. 1979, 124, 361–369. [Google Scholar] [CrossRef]
- Akasaka, K.; Amemiya, S.; Terayama, H. Scanning electron microscopical study of the inside of sea urchin embryos (Pseudocentotus depressus). Effects of Aryl beta-xyloside, tunicamycin and deprivation of sulfate tions. Exp. Cell Res. 1980, 129, 1–13. [Google Scholar] [CrossRef]
- Solursh, M.; Mitchell, S.L.; Katow, H. Inhibition of cell migration in sea urchin embryos by beta-D-xyloside. Dev. Biol. 1986, 118, 325–332. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Gallo, A.; Costantini, M. Glycobiology of Reproductive Processes in Marine Animals: The State of the Art. Mar. Drugs 2012, 10, 2861-2892. https://doi.org/10.3390/md10122861
Gallo A, Costantini M. Glycobiology of Reproductive Processes in Marine Animals: The State of the Art. Marine Drugs. 2012; 10(12):2861-2892. https://doi.org/10.3390/md10122861
Chicago/Turabian StyleGallo, Alessandra, and Maria Costantini. 2012. "Glycobiology of Reproductive Processes in Marine Animals: The State of the Art" Marine Drugs 10, no. 12: 2861-2892. https://doi.org/10.3390/md10122861