Oxytocin: Old Hormone, New Drug

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Cardiac OT Actions
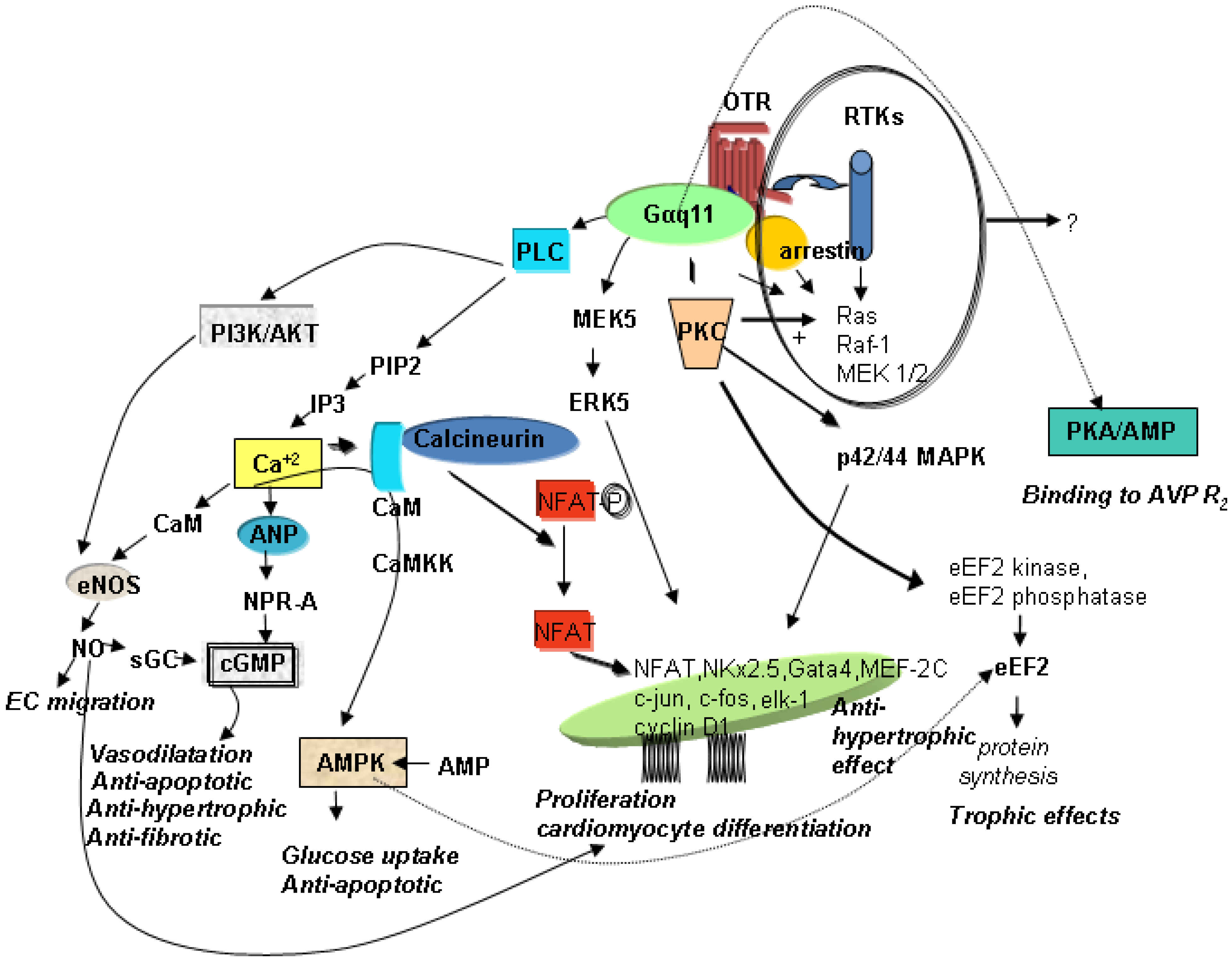
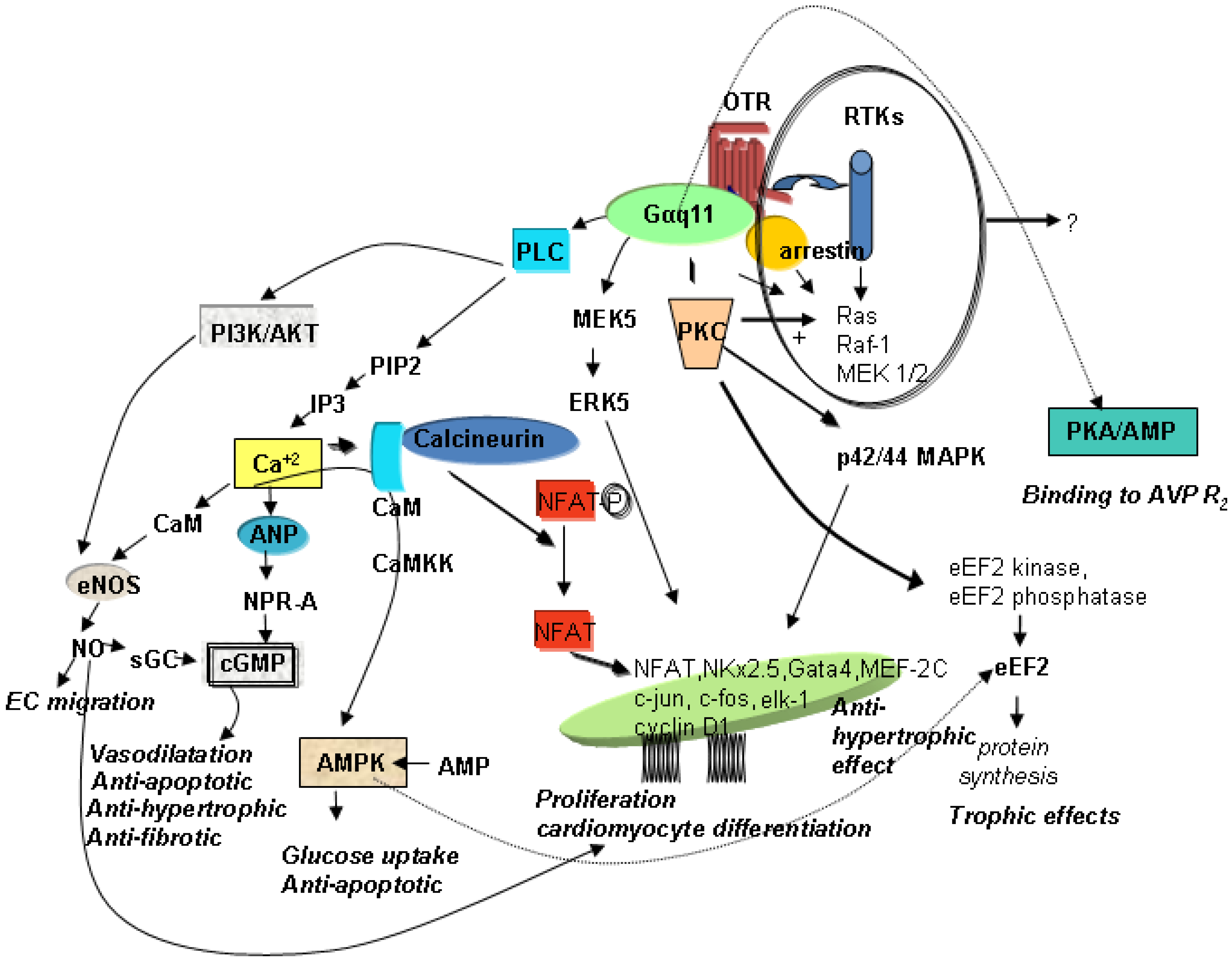
3. Mechanisms of OT-Mediated Cardioprotection
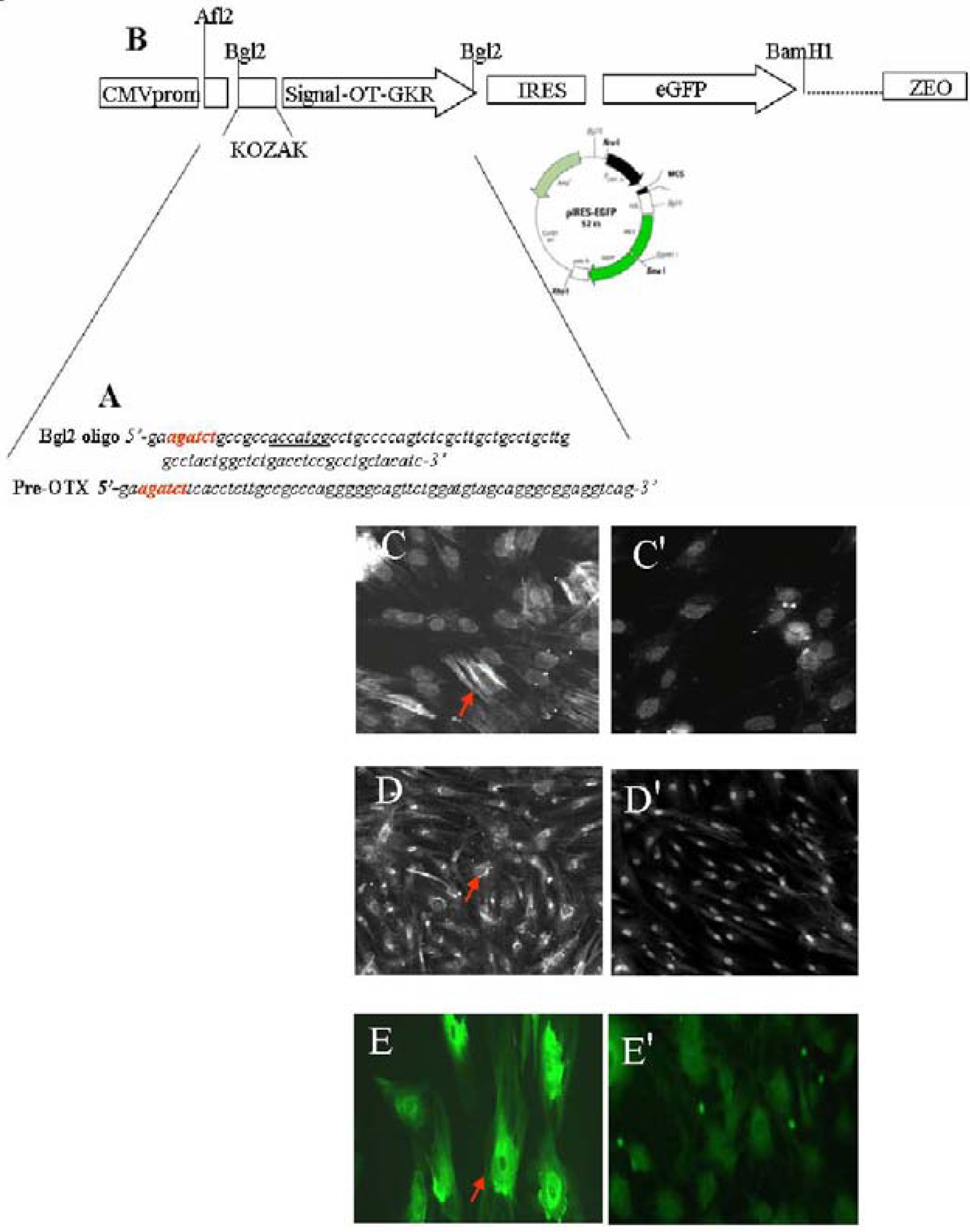
4. OT’s Role in Inflammation
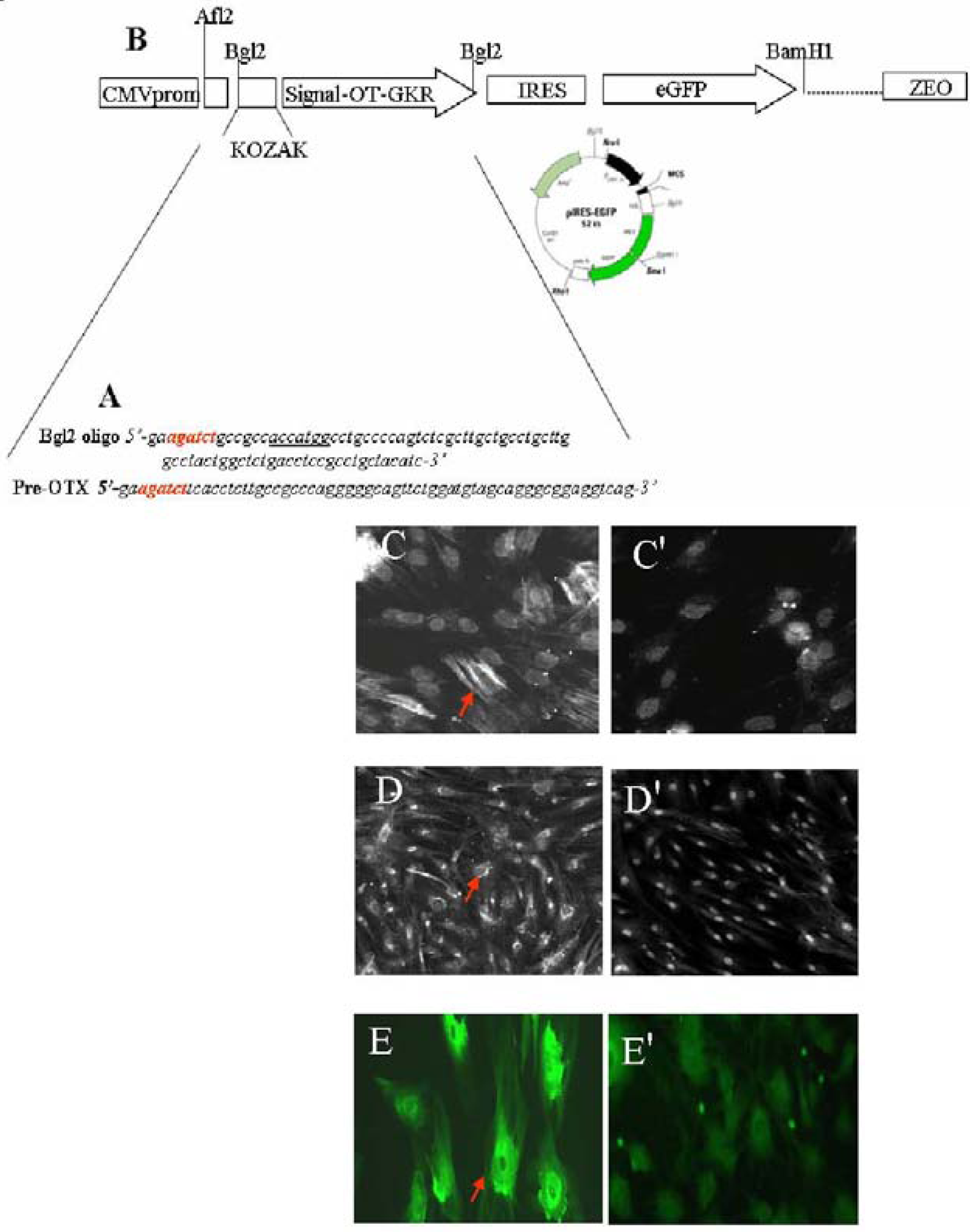
5. OT Induces Stem Cell Differentiation
6. OT Stimulates CMs Differentiation in 3-Dimensional Cultures
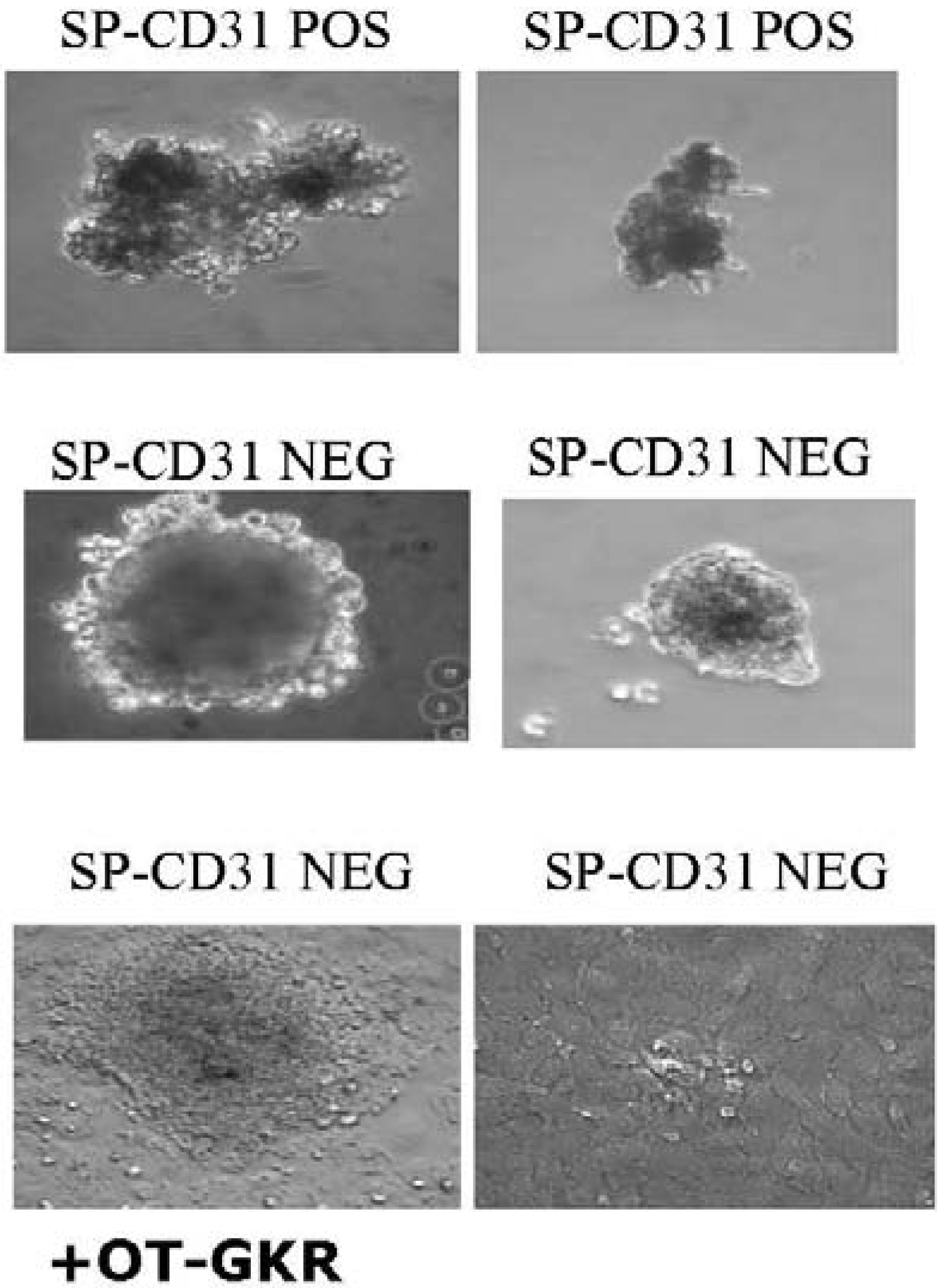
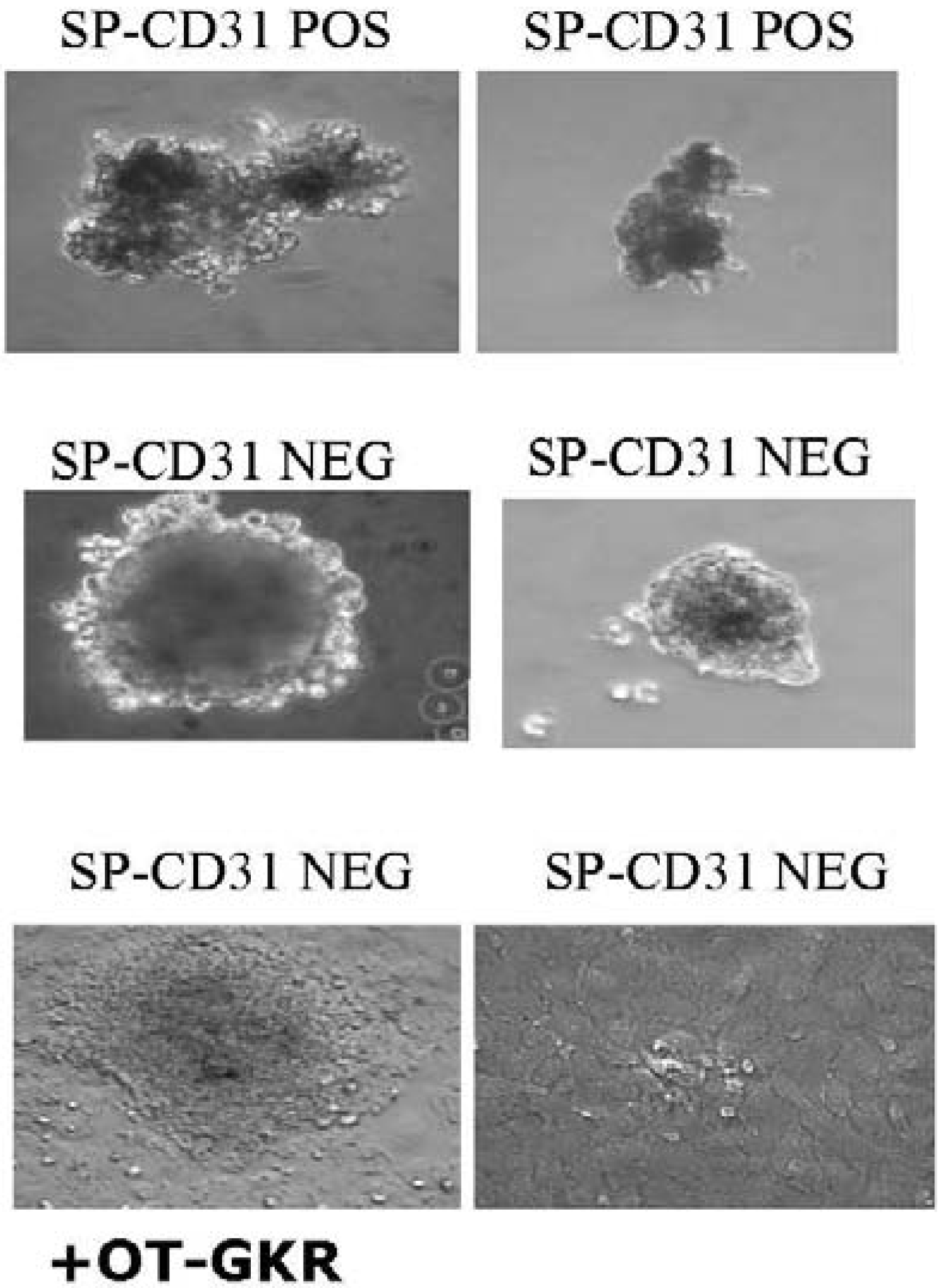
7. Differentiation of Endogenous Stem Cells Isolated from Animal Hearts
8. Summary and Conclusions
Acknowledgements
References and Notes
- Dale, H.H. The actions of extracts of pituitary body. Biochem. J. 1910, 4, 427–447. [Google Scholar]
- Ott, I.; Scott, J.C. The action of infundibulin upon the mammary secretion. Proc. Soc. Exp. Biol. NY 1910, 8, 48–49. [Google Scholar]
- Du Vigneaud, V.; Ressler, C.; Trippett, S. The sequence of amino acids in oxytocin, with a proposal for the structure of oxytocin. J. Biol. Chem. 1953, 205, 949–957. [Google Scholar]
- Gimpl, G.; Fahrenholz, F. The oxytocin receptor system: Structure, function, and regulation. Physiol. Rev. 2001, 81, 629–683. [Google Scholar] [PubMed]
- McCann, S.M.; Antunes-Rodrigues, J.; Jankowski, M.; Gutkowska, J. Oxytocin, vasopressin and atrial natriuretic peptide control body fluid homeostasis by action on their receptors in brain, cardiovascular system and kidney. Prog. Brain. Res. 2002, 139, 309–328. [Google Scholar] [CrossRef] [PubMed]
- Gutkowska, J.; Jankowski, M.; Lambert, C.; Mukaddam-Daher, S.; Zingg, H.H.; McCann, S.M. Oxytocin releases atrial natriuretic peptide by combining with oxytocin receptors in the heart. Proc. Natl. Acad. Sci. USA 1997, 94, 11704–11709. [Google Scholar]
- Jankowski, M.; Hajjar, F.; Al Kawas, S.; Mukaddam-Daher, S.; Hoffman, G.; McCann, S.M.; Gutkowska, J. Rat heart: A site of oxytocin production and action. Proc. Natl. Acad. Sci. USA 1998, 95, 14558–14563. [Google Scholar]
- Danalache, B.A.; Paquin, J.; Donghao, W.; Grygorczyk, R.; Moore, J.C.; Mummery, C.L.; Gutkowska, J.; Jankowski, M. Nitric oxide signaling in oxytocin-mediated cardiomyogenesis. Stem Cells 2007, 25, 679–688. [Google Scholar]
- Paquin, J.; Danalache, B.A.; Jankowski, M.; McCann, S.M.; Gutkowska, J. Oxytocin induces differentiation of P19 embryonic stem cells to cardiomyocytes. Proc. Natl. Acad. Sci. USA 2002, 99, 9550–9555. [Google Scholar]
- Jankowski, M.; Danalache, B.; Wang, D.; Bhat, P.; Hajjar, F.; Marcinkiewicz, M.; Paquin, J.; McCann, S.M.; Gutkowska, J. Oxytocin in cardiac ontogeny. Proc. Natl. Acad. Sci. USA 2004, 101, 13074–13079. [Google Scholar]
- Matsuura, K.; Nagai, T.; Nishigaki, N.; Oyama, T.; Nishi, J.; Wada, H.; Sano, M.; Toko, H.; Akazawa, H.; Sato, T.; Nakaya, H.; Kasanuki, H.; Komuro, I. Adult cardiac Sca-1-positive cells differentiate into beating cardiomyocytes. J. Biol. Chem. 2004, 279, 11384–11391. [Google Scholar]
- Oyama, T.; Nagai, T.; Wada, H.; Naito, A.T.; Matsuura, K.; Iwanaga, K.; Takahashi, T.; Goto, M.; Mikami, Y.; Yasuda, N.; Akazawa, H.; Uezumi, A.; Takeda, S.; Komuro, I. Cardiac side population cells have a potential to migrate and differentiate into cardiomyocytes in vitro and in vivo. J. Cell Biol. 2007, 176, 329–341. [Google Scholar] [CrossRef] [PubMed]
- Florian, M.; Jankowski, M.; Gutkowska, J. Oxytocin increases glucose uptake in neonatal rat cardiomyocytes. Endocrinology 2010, in press. [Google Scholar]
- Soares, T.J.; Coimbra, T.M.; Martins, A.R.; Pereira, A.G.; Carnio, E.C.; Branco, L.G.; Albuquerque-Araujo, W.I.; De Nucci, G.; Favaretto, A.L.; Gutkowska, J.; McCann, S.M.; Antunes-Rodrigues, J. Atrial natriuretic peptide and oxytocin induce natriuresis by release of cGMP. Proc. Natl. Acad. Sci. USA 1999, 96, 278–283. [Google Scholar]
- Favaretto, A.L.; Ballejo, G.O.; Albuquerque-Araujo, W.I.; Gutkowska, J.; Antunes-Rodrigues, J.; McCann, S.M. Oxytocin releases atrial natriuretic peptide from rat atria in vitro that exerts negative inotropic and chronotropic action. Peptides 1997, 18, 1377–1381. [Google Scholar]
- Mukaddam-Daher, S.; Lin, Y.L.; Gutkowska, J.; Cardinal, R. Negative inotropic and chronotropic effects of oxytocin. Hypertension 2001, 38, 292–296. [Google Scholar]
- Thibonnier, M.; Conarty, D.M.; Preston, J.A.; Plesnicher, C.L.; Dweik, R.A.; Erzurum, S.C. Human vascular endothelial cells express oxytocin receptors. Endocrinology 1999, 140, 1301–1309. [Google Scholar]
- Bobbioni-Harsch, E.; Frütiger, S.; Hughes, G.; Panico, M.; Etienne, A.; Zappacosta, F.; Morris, H.R.; Jeanrenaud, B. Physiological concentrations of oxytocin powerfully stimulate insulin secretion in vitro. Endocrine 1995, 3, 55–59. [Google Scholar] [PubMed]
- Reversi, A.; Cassoni, P.; Chini, B. Oxytocin receptor signaling in myoepithelial and cancer cells. J. Mammary Gland. Biol. Neoplasia. 2005, 10, 221–229. [Google Scholar]
- Zingg, H.H.; Laporte, S.A. The oxytocin receptor. Trends Endocrinol. Metab. 2003, 14, 222–227. [Google Scholar]
- Gassanov, N.; Jankowski, M.; Danalache, B.; Wang, D.; Grygorczyk, R.; Hoppe, U.C.; Gutkowska, J. Arginine Vasopressin-mediated Cardiac Differentiation: insights into the role of its receptors and nitric oxide signaling. J. Biol. Chem. 2007, 282, 11255–11265. [Google Scholar]
- Gutkowska, J.; Miszkurka, M.; Danalache, B.; Gassanov, N.; Wang, D.; Jankowski, M. Functional arginine-vasopressin system in early heart maturation. Am. J. Physiol. Heart Circ. Physiol. 2007, 293, H2262–H2270. [Google Scholar]
- Takayanagi, Y.; Kasahara, Y.; Onaka, T.; Takahashi, N.; Kawada, T.; Nishimori, K. Oxytocin receptor-deficient mice developed late-onset obesity. Neuroreport 2008, 19, 951–955. [Google Scholar]
- Bernatova, I.; Rigatto, K.V.; Key, M.P.; Morris, M. Stress-induced pressor and corticosterone responses in oxytocin-deficient mice. Exp. Physiol. 2004, 89, 549–557. [Google Scholar]
- Michelini, L.C.; Marcelo, M.C.; Amico, J.; Morris, M. Oxytocinergic regulation of cardiovascular function: Studies in oxytocin-deficient mice. Am. J. Physiol. Heart Circ. Physiol. 2003, 284, H2269–H2276. [Google Scholar]
- Iseri, S.O.; Sener, G.; Saglam, B.; Gedik, N.; Ercan, F.; Yegen, B.C. Oxytocin ameliorates oxidative colonic inflammation by a neutrophil-dependent mechanism. Peptides 2005, 26, 483–491. [Google Scholar]
- Szeto, A.; Nation, D.A.; Mendez, A.J.; Dominguez-Bendala, J.; Brooks, L.G.; Schneiderman, N.; McCabe, P.M. Oxytocin attenuates NADPH-dependent superoxide activity and IL-6 secretion in macrophages and vascular cells. Am. J. Physiol. Endocrinol. Metab. 2008, 295, E1495–E1501. [Google Scholar]
- Houshmand, F.; Faghihi, M.; Zahediasl, S. Biphasic protective effect of oxytocin on cardiac ischemia/reperfusion injury in anaesthetized rats. Peptides 2009. Epub ahead of print. [Google Scholar]
- Kobayashi, H.; Yasuda, S.; Bao, N.; Iwasa, M.; Kawamura, I.; Yamada, Y.; Yamaki, T.; Sumi, S.; Ushikoshi, H.; Nishigaki, K.; Takemura, G.; Fujiwara, T.; Fujiwara, H.; Minatoguchi, S. Post-infarct treatment with oxytocin improves cardiac function and remodeling via activating cell-survival signals and angiogenesis. J. Cardiovasc. Pharmacol. 2009. Sept 12, Epub ahead of print.. [Google Scholar]
- Ondrejcakova, M.; Ravingerova, T.; Bakos, J.; Pancza, D.; Jezova, D. Oxytocin exerts protective effects on in vitro myocardial injury induced by ischemia and reperfusion. Can. J. Physiol. Pharmacol. 2009, 87, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Authier, S.; Tanguay, J.F.; Geoffroy, P.; Gauvin, D.; Bichot, S.; Ybarra, N.; Otis, C.; Troncy, E. Cardiovascular effects of oxytocin infusion in a porcine model of myocardial infarct. J. Cardiovasc. Pharmacol. 2009. Epub ahead of print. [Google Scholar]
- Dietz, J.R. Mechanisms of atrial natriuretic peptide secretion from the atrium. Cardiovasc. Res. 2005, 68, 8–17. [Google Scholar]
- Toth, M.; Vuorinen, K.H.; Vuolteenaho, O.; Hassinen, I.E.; Uusimaa, P.A.; Leppaluoto, J.; Ruskoaho, H. Hypoxia stimulates release of ANP and BNP from perfused rat ventricular myocardium. Am. J. Physiol. 1994, 266, H1572–H1580. [Google Scholar]
- Ruskoaho, H. Atrial natriuretic peptide: Synthesis, release, and metabolism. Pharmacol. Rev. 1992, 44, 479–602. [Google Scholar]
- Oliver, P.M.; Fox, J.E.; Kim, R.; Rockman, H.A.; Kim, H.S.; Reddick, R.L.; Pandey, K.N.; Milgram, S.L.; Smithies, O.; Maeda, N. Hypertension, cardiac hypertrophy, and sudden death in mice lacking natriuretic peptide receptor A. Proc. Natl. Acad. Sci. USA 1997, 94, 14730–14735. [Google Scholar]
- Calderone, A.; Thaik, C.M.; Takahashi, N.; Chang, D.L.F.; Colucci, W.S. Nitric oxide, atrial natriuretic peptide, and cyclic GMP inhibit the growth-promoting effects of norepinephrine in cardiac myocytes and fibroblasts. J. Clin. Invest. 1998, 101, 812–818. [Google Scholar] [CrossRef] [PubMed]
- De Vito, P.; Di Nardo, P.; Palmery, M.; Peluso, I.; Luly, P.; Baldini, P.M. Oxidant-induced pHi/Ca2+ changes in rat aortic smooth muscle cells. The role of atrial natriuretic peptide. Mol. Cell Biochem. 2003, 252, 353–362. [Google Scholar] [CrossRef] [PubMed]
- Kiemer, A.K.; Vollmar, A.M. The atrial natriuretic peptide regulates the production of inflammatory mediators in macrophages. Ann. Rheum. Dis. 2001, 60 (Suppl. 3), iii68–iii70. [Google Scholar] [PubMed]
- Birkenfeld, A.L.; Budziarek, P.; Boschmann, M.; Moro, C.; Adams, F.; Franke, G.; Berlan, M.; Marques, M.A.; Sweep, F.C.; Luft, F.C.; Lafontan, M.; Jordan, J. Atrial natriuretic peptide induces postprandial lipid oxidation in humans. Diabetes 2008, 57, 3199–3204. [Google Scholar]
- Khairallah, R.J.; Khairallah, M.; Gεlinas, R.; Bouchard, B.; Young, M.E.; Allen, B.G.; Lopaschuk, G.D.; Deschepper, C.F.; Des Rosiers, C. Cyclic GMP signaling in cardiomyocytes modulates fatty acid trafficking and prevents triglyceride accumulation. J. Mol. Cell. Cardiol. 2008, 45, 230–239. [Google Scholar]
- Mehra, M.R.; Uber, P.A.; Park, M.H.; Scott, R.L.; Ventura, H.O.; Harris, B.C.; Frohlich, E.D. Obesity and suppressed B-type natriuretic peptide levels in heart failure. J. Am. Coll. Cardiol. 2004, 43, 1590–1595. [Google Scholar]
- Taylor, J.A.; Christenson, R.H.; Rao, K.; Jorge, M.; Gottlieb, S.S. B-type natriuretic peptide and N-terminal pro B-type natriuretic peptide are depressed in obesity despite higher left ventricular end diastolic pressures. Am. Heart J. 2006, 152, 1071–1076. [Google Scholar]
- Aoyama, E.; Yoshihara, R.; Tai, A.; Yamamoto, I.; Gohda, E. PKC- and PI3K-dependent but ERK-independent proliferation of murine splenic B cells stimulated by chondroitin sulfate B. Immunol. Lett. 2005, 99, 80–84. [Google Scholar]
- Miki, T.; Miura, T.; Tanno, M.; Nishihara, M.; Naitoh, K.; Sato, T.; Takahashi, A.; Shimamoto, K. Impairment of cardioprotective PI3K-Akt signaling by post-infarct ventricular remodeling is compensated by an ERK-mediated pathway. Basic Res. Cardiol. 2007, 102, 163–170. [Google Scholar]
- Lee, E.S.; Uhm, K.O.; Lee, Y.M.; Kwon, J.; Park, S.H.; Soo, K.H. Oxytocin stimulates glucose uptake in skeletal muscle cells through the calcium-CaMKK-AMPK pathway. Regul. Pept. 2008, 151, 71–74. [Google Scholar]
- Marsin, A.S.; Bouzin, C.; Bertrand, L.; Hue, L. The stimulation of glycolysis by hypoxia in activated monocytes is mediated by AMP-activated protein kinase and inducible 6-phosphofructo-2-kinase. J. Biol. Chem. 2002, 277, 30778–30783. [Google Scholar]
- Russell, R.R., III; Li, J.; Coven, D.L.; Pypaert, M.; Zechner, C.; Palmeri, M.; Giordano, F.J.; Mu, J.; Birnbaum, M.J.; Young, L.H. AMP-activated protein kinase mediates ischemic glucose uptake and prevents postischemic cardiac dysfunction, apoptosis, and injury. J. Clin. Invest. 2004, 114, 495–503. [Google Scholar]
- Kewalramani, G.; Puthanveetil, P.; Wang, F.; Kim, M.S.; Deppe, S.; Abrahani, A.; Luciani, D.S.; Johnson, J.D.; Rodrigues, B. AMP-activated protein kinase confers protection against TNF-{alpha}-induced cardiac cell death. Cardiovasc. Res. 2009, 84, 42–53. [Google Scholar]
- Horman, S.; Browne, G.; Krause, U.; Patel, J.; Vertommen, D.; Bertrand, L.; Lavoinne, A.; Hue, L.; Proud, C.; Rider, M. Activation of AMP-activated protein kinase leads to the phosphorylation of elongation factor 2 and an inhibition of protein synthesis. Curr. Biol. 2002, 12, 1419–1423. [Google Scholar]
- Chan, A.Y.; Soltys, C.L.; Young, M.E.; Proud, C.G.; Dyck, J.R. Activation of AMP-activated protein kinase inhibits protein synthesis associated with hypertrophy in the cardiac myocyte. J. Biol. Chem. 2004, 279, 32771–32779. [Google Scholar]
- Cattaneo, M.G.; Lucci, G.; Vicentini, L.M. Oxytocin stimulates in vitro angiogenesis via a Pyk-2/Src-dependent mechanism. Exp. Cell Res. 2009, 315, 3210–3219. [Google Scholar] [CrossRef] [PubMed]
- Devost, D.; Wrzal, P.; Zingg, H.H. Oxytocin receptor signalling. Prog. Brain. Res. 2008, 170, 167–176. [Google Scholar]
- Hausenloy, D.J.; Yellon, D.M. New directions for protecting the heart against ischaemia-reperfusion injury: Targeting the Reperfusion Injury Salvage Kinase (RISK)-pathway. Cardiovasc. Res. 2004, 61, 448–460. [Google Scholar]
- Jezdinsky, J.; Stojan, B.; Queisnerova, M. Demonstration of the existance of an anti-inflammatory factor situated in the posterior pituitary. Arch. Int. Pharmacodyn. Ther. 1963, 146, 561–578. [Google Scholar]
- Hansenne, I.; Rasier, G.; Pequeux, C.; Brilot, F.; Renard, C.; Breton, C.; Greimers, R.; Legros, J.J.; Geenen, V.; Martens, H.J. Ontogenesis and functional aspects of oxytocin and vasopressin gene expression in the thymus network. J. Neuroimmunol. 2005, 158, 67–75. [Google Scholar]
- Ndiaye, K.; Poole, D.H.; Pate, J.L. Expression and regulation of functional oxytocin receptors in bovine T lymphocytes. Biol Reprod. 2008, 78, 786–793. [Google Scholar] [CrossRef] [PubMed]
- Kimura, T. Regulation of the human oxytocin receptor in the uterus: A molecular approach. Hum. Reprod. Update. 1998, 4, 615–624. [Google Scholar]
- Petersson, M.; Wiberg, U.; Lundeberg, T.; Uvnas-Moberg, K. Oxytocin decreases carrageenan induced inflammation in rats. Peptides 2001, 22, 1479–1484. [Google Scholar]
- Asad, M.; Shewade, D.G.; Koumaravelou, K.; Abraham, B.K.; Vasu, S.; Ramaswamy, S. Gastric antisecretory and antiulcer activity of oxytocin in rats and guinea pigs. Life Sci. 2001, 70, 17–24. [Google Scholar]
- Iseri, S.O.; Sener, G.; Saglam, B.; Gedik, N.; Ercan, F.; Yegen, B.C. Oxytocin protects against sepsis-induced multiple organ damage: Role of neutrophils. J. Surg. Res. 2005, 126, 73–81. [Google Scholar]
- Iseri, S.O.; Gedik, I.E.; Erzik, C.; Uslu, B.; Arbak, S.; Gedik, N.; Yegen, B.C. Oxytocin ameliorates skin damage and oxidant gastric injury in rats with thermal trauma. Burns 2008, 34, 361–369. [Google Scholar]
- Dusunceli, F.; Iseri, S.O.; Ercan, F.; Gedik, N.; Yegen, C.; Yegen, B.C. Oxytocin alleviates hepatic ischemia-reperfusion injury in rats. Peptides 2008, 29, 1216–1222. [Google Scholar]
- Detillion, C.E.; Craft, T.K.; Glasper, E.R.; Prendergast, B.J.; DeVries, A.C. Social facilitation of wound healing. Psychoneuroendocrinology 2004, 29, 1004–1011. [Google Scholar]
- Ozlem, I.S.; Dusunceli, F.; Erzik, C.; Uslu, B.; Arbak, S.; Yegen, B.C. Oxytocin or social housing alleviates local burn injury in rats. J. Surg. Res. 2009. Epub ahead of print. [Google Scholar]
- Hoshinaga, K.; Mohanakumar, T.; Goldman, M.H.; Wolfgang, T.C.; Szentpetery, S.; Lee, H.M.; Lower, R.R. Clinical significance of in situ detection of T lymphocyte subsets and monocyte/macrophage lineages in heart allografts. Transplantation 1984, 38, 634–637. [Google Scholar] [CrossRef] [PubMed]
- Jankowski, M.; Bissonauth, V.; Gao, L.; Gangal, M.; Wang, D.; Danalache, B.; Wang, Y.; Stoyanova, E.; Clouthier, G.; Blaise, G.; Gutkowska, J. Anti-inflammatory effect of oxytocin in rat myocardial infarction. Basic Res. Cardiol. 2009, in press. [Google Scholar]
- Geenen, V. Thymus-dependent T cell tolerance of neuroendocrine functions: Principles, reflections, and implications for tolerogenic/negative self-vaccination. Ann. NY Acad. Sci. 2006, 1088, 284–296. [Google Scholar] [CrossRef]
- Bodi, V.; Sanchis, J.; Nunez, J.; Mainar, L.; Minana, G.; Benet, I.; Solano, C.; Chorro, F.J.; Llacer, A. Uncontrolled immune response in acute myocardial infarction: Unraveling the thread. Am. Heart J. 2008, 156, 1065–1073. [Google Scholar]
- Frangogiannis, N.G. The immune system and cardiac repair. Pharmacol. Res. 2008, 58, 88–111. [Google Scholar]
- Spangelo, B.L.; deHoll, P.D.; Kalabay, L.; Bond, B.R.; Arnaud, P. Neurointermediate pituitary lobe cells synthesize and release interleukin-6 in vitro: Effects of lipopolysaccharide and interleukin-1 beta. Endocrinology 1994, 135, 556–563. [Google Scholar] [PubMed]
- Chaouat, G.; Ledee-Bataille, N.; Dubanchet, S. Immune cells in uteroplacental tissues throughout pregnancy: A brief review. Reprod. Biomed. Online. 2007, 14, 256–266. [Google Scholar]
- Tabata, C.; Ogita, K.; Sato, K.; Nakamura, H.; Qing, Z.; Negoro, H.; Kumasawa, K.; Temma-Asano, K.; Tsutsui, T.; Nishimori, K.; Kimura, T. Calcineurin/NFAT pathway: A novel regulator of parturition. Am. J. Reprod. Immunol. 2009, 62, 44–50. [Google Scholar]
- Fathi, F.; Murasawa, S.; Hasegawa, S.; Asahara, T.; Kermani, A.J.; Mowla, S.J. Cardiac differentiation of P19CL6 cells by oxytocin. Int. J. Cardiol. 2009, 134, 75–81. [Google Scholar]
- Hatami, L.; Valojerdi, M.R.; Mowla, S.J. Effects of oxytocin on cardiomyocyte differentiation from mouse embryonic stem cells. Int. J. Cardiol. 2007, 117, 80–89. [Google Scholar]
- Uchida, S.; Fuke, S.; Tsukahara, T. Upregulations of Gata4 and oxytocin receptor are important in cardiomyocyte differentiation processes of P19CL6 cells. J. Cell. Biochem. 2007, 100, 629–641. [Google Scholar]
- Gassanov, N.; Devost, D.; Danalache, B.; Noiseux, N.; Jankowski, M.; Zingg, H.H.; Gutkowska, J. Functional activity of the carboxyl-terminally extended oxytocin precursor peptide during cardiac differentiation of embryonic stem cells. Stem Cells 2008, 26, 45–54. [Google Scholar]
- Bisping, E.; Ikeda, S.; Kong, S.W.; Tarnavski, O.; Bodyak, N.; McMullen, J.R.; Rajagopal, S.; Son, J.K.; Ma, Q.; Springer, Z.; Kang, P.M.; Izumo, S.; Pu, W.T. Gata4 is required for maintenance of postnatal cardiac function and protection from pressure overload-induced heart failure. Proc. Natl. Acad. Sci. USA 2006, 103, 14471–14476. [Google Scholar]
- Heineke, J.; Auger-Messier, M.; Xu, J.; Oka, T.; Sargent, M.A.; York, A.; Klevitsky, R.; Vaikunth, S.; Duncan, S.A.; Aronow, B.J.; Robbins, J.; Crombleholme, T.M.; Molkentin, J.D. Cardiomyocyte GATA4 functions as a stress-responsive regulator of angiogenesis in the murine heart. J. Clin. Invest. 2007, 117, 3198–3210. [Google Scholar]
- Abdelalim, E.M.; Tooyama, I. BNP signaling is crucial for embryonic stem cell proliferation. PLoS. ONE. 2009, 4, e5341. [Google Scholar]
- van der Heyden, M.A.; Defize, L.H. Twenty one years of P19 cells: What an embryonal carcinoma cell line taught us about cardiomyocyte differentiation. Cardiovasc. Res. 2003, 58, 292–302. [Google Scholar]
- Van der Heyden, M.A.; van Kempen, M.J.; Tsuji, Y.; Rook, M.B.; Jongsma, H.J.; Opthof, T. P19 embryonal carcinoma cells: A suitable model system for cardiac electrophysiological differentiation at the molecular and functional level. Cardiovasc. Res. 2003, 58, 410–422. [Google Scholar]
- Pfister, O.; Mouquet, F.; Jain, M.; Summer, R.; Helmes, M.; Fine, A.; Colucci, W.S.; Liao, R. CD31- but Not CD31+ cardiac side population cells exhibit functional cardiomyogenic differentiation. Circ. Res. 2005, 97, 52–61. [Google Scholar]
- Martin, C.M.; Meeson, A.P.; Robertson, S.M.; Hawke, T.J.; Richardson, J.A.; Bates, S.; Goetsch, S.C.; Gallardo, T.D.; Garry, D.J. Persistent expression of the ATP-binding cassette transporter, Abcg2, identifies cardiac SP cells in the developing and adult heart. Dev. Biol. 2004, 265, 262–275. [Google Scholar] [CrossRef] [PubMed]
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Gutkowska, J.; Jankowski, M. Oxytocin: Old Hormone, New Drug. Pharmaceuticals 2009, 2, 168-183. https://doi.org/10.3390/ph203168
Gutkowska J, Jankowski M. Oxytocin: Old Hormone, New Drug. Pharmaceuticals. 2009; 2(3):168-183. https://doi.org/10.3390/ph203168
Chicago/Turabian StyleGutkowska, Jolanta, and Marek Jankowski. 2009. "Oxytocin: Old Hormone, New Drug" Pharmaceuticals 2, no. 3: 168-183. https://doi.org/10.3390/ph203168