Detecting Proteins in Highly Autofluorescent Cells Using Quantum Dot Antibody Conjugates

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Experimental Section
2.1. Culture Conditions
2.2. Immunolabeling Procedure
3. Results and Discussion
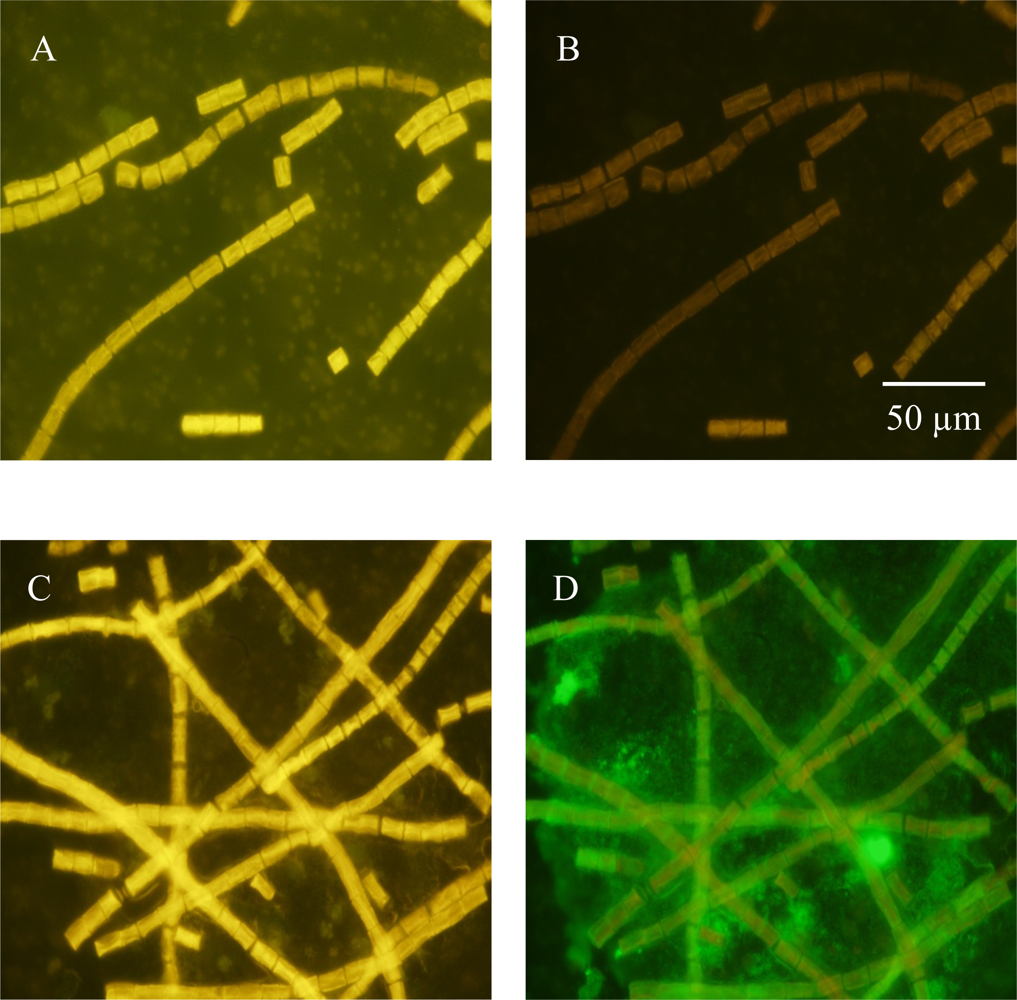
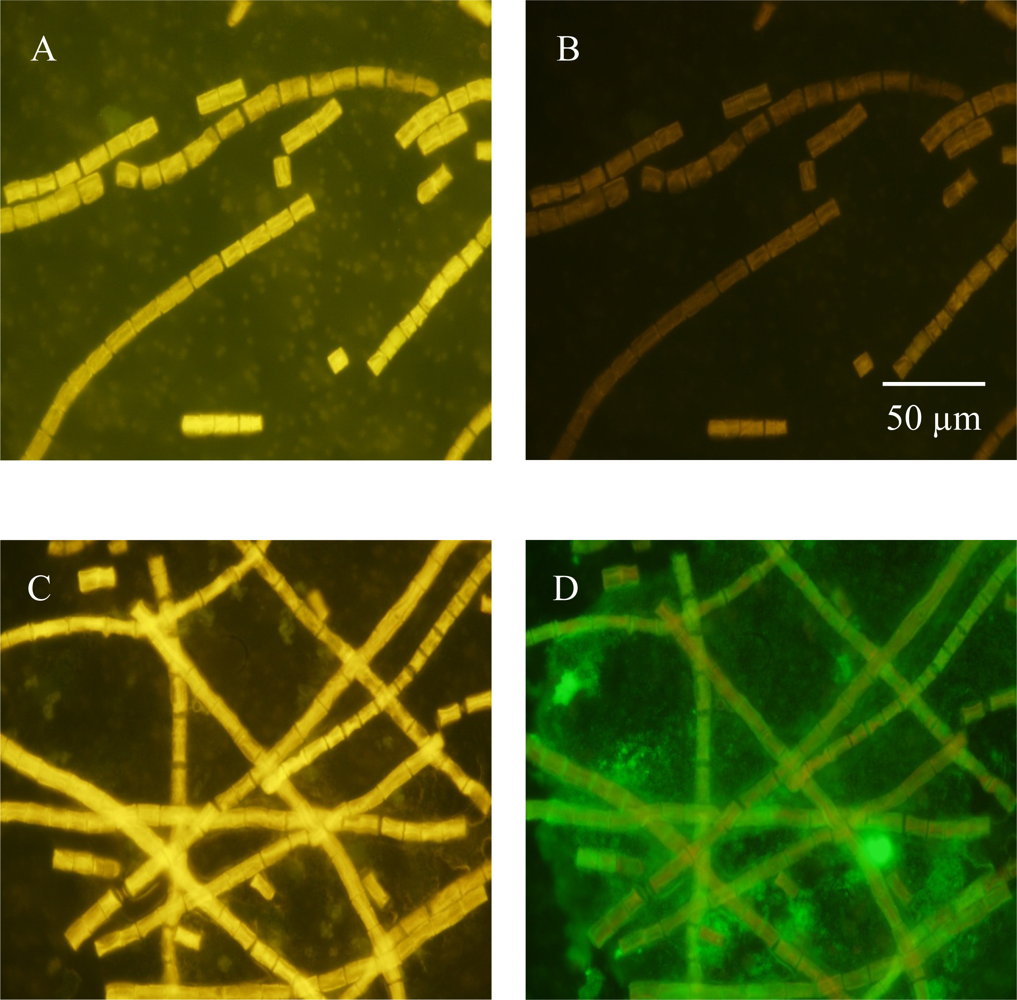
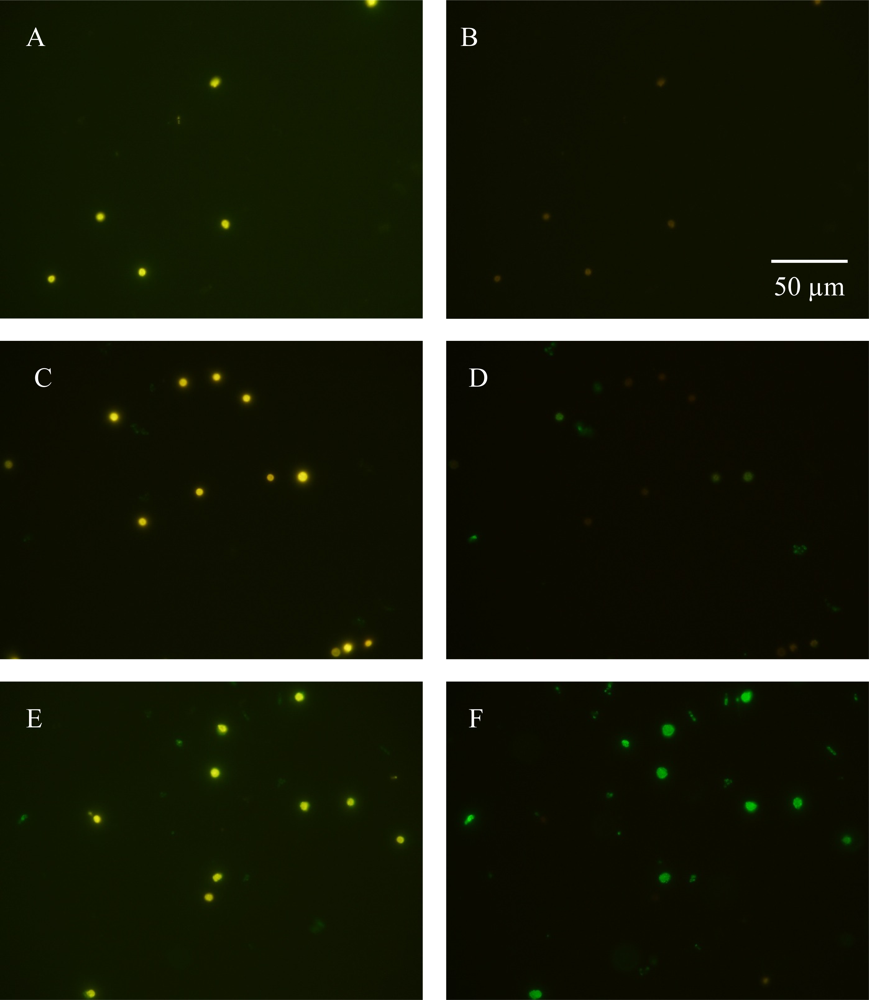
3.1. Detecting in situ Iron Stress with Qdot Antibody Conjugates
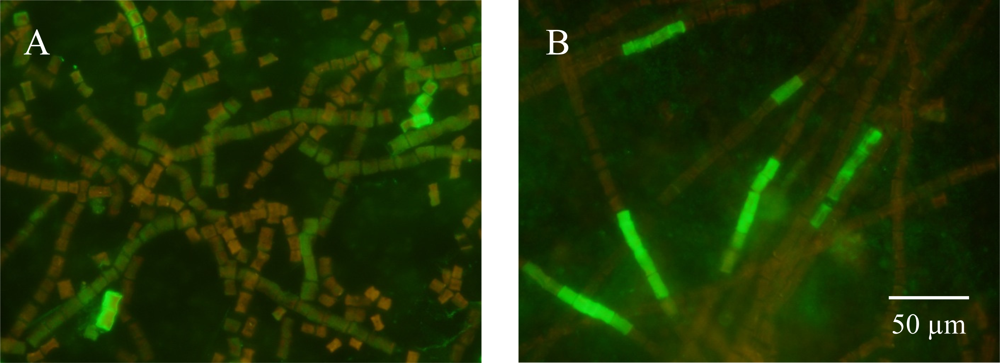
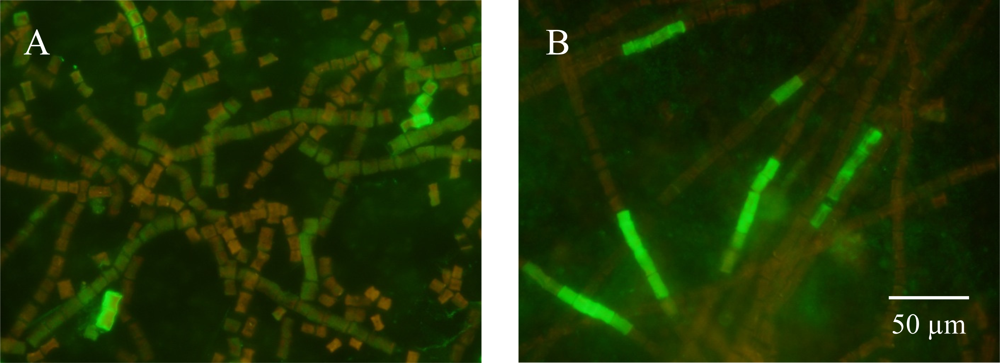
3.2. Detecting Nitrogenase in Highly Autofluorescent Cells
4. Conclusions
Acknowledgments
References
- Taniuchi, Y.; Murakami, A.; Ohki, K. Whole-cell immunocytochemical detection of nitrogenase in cyanobacteria: improved protocol for highly fluorescent cells. Aquat. Microb. Ecol 2008, 51, 237–247. [Google Scholar]
- Ohki, K. Intercellular localization of nitrogenase in a non-heterocystous cyanobacterium (Cyanophyte), Trichodesmium sp. NIBB1067. J. Oceanogr 2008, 64, 211–216. [Google Scholar]
- Ohki, K.; Taniuchi, Y. Detection of nitrogenase in individual cells of a natural population of Trichodesmium using immunocytochemical methods for fluorescent cells. J. Oceanogr 2009, 65, 427–432. [Google Scholar]
- Lin, S.; Henze, S.; Lundgren, P.; Bergman, B.; Carpenter, E.J. Whole-cell immunolocalization of nitrogenase in marine diazotrophic cyanobacteria, Trichodesmium spp. Appl. Environ. Microbiol 1998, 64, 3052–3058. [Google Scholar]
- Falcón, L.I.; Carpenter, E.J.; Cipriano, F.; Bergman, B.; Capone, D.G. N2 fixation by unicellular bacteriopplankton from the Atlantic and Pacific Oceans: Phylogeny and in situ rates. Appl. Environ. Microbiol 2004, 70, 765–770. [Google Scholar]
- Michel, K-P.; Thole, H.H.; Pistorius, E.K. IdiA, a 34 kDa protein in the cyanobacteria Synechococcus sp. Strains 6301 and PCC 7942, is required for growth under iron and manganese limitations. Microbiology 1996, 142, 2635–2645. [Google Scholar]
- Michel, K-P.; Exss-Sonne, P.; Scholten-Beck, G.; Kahmann, U.; Ruppel, H.G.; Pistorius, E.K. Immunochemical localization of IdiA, a protein expressed under iron and manganese limitation in the mesophilic cyanobacterium Synechococcus PCC 6301 and the thermophilic cynaobacteria Synechococcus elongates. Planta 1998, 205, 73–81. [Google Scholar]
- Tölle, J.; Michel, K.-P.; Kruip, J.; Kahmann, U.; Preisfeld, A.; Pistorius, E.K. Localization and function of the IdiA homologue Slr1295 in the cyamobacterium Synechocystis sp. Strain PCC 6803. Microbiology 2002, 148, 3293–3305. [Google Scholar]
- Webb, E.A.; Moffett, M.T.; Waterbury, J.B. Iron stress in open-ocean cyanobacteria (Synechococcus, Trichodesmium, and Crocosphaera spp.): Identification of the IdiA protein. Appl. Environ. Microbiol 2001, 67, 5444–5452. [Google Scholar]
- Zehr, J.P.; Waterbury, J.B.; Turner, P.J.; Montoya, J.P.; Omoregie, E.; Steward, G.F.; Hansen, A.; Karl, D.M. Unicellular cyanobacteria fix N2 in the subropical North Pacific Ocean. Nature 2001, 412, 635–638. [Google Scholar]
- Montoya, J.P.; Holl, C.M.; Zehr, J.P.; Hansen, A.; Villareal, T.A.; Capone, D.G. High rates of N2 fixation by unicellular diazotrophs in the oliggotrophic Pacific Ocean. Nature 2004, 430, 1027–1031. [Google Scholar]
- Waterbury, J.B.; Watson, S.B.; Valois, F.W.; Franks, D.G. Biological and ecological characterization of the marine cyanobacterium Synechococcus. Can. Bull. Fish. Aquat. Sci 1986, 214, 71–120. [Google Scholar]
- Chen, Y.-B.; Zehr, J.P.; Mellon, M. Growth and nitrogen fixation of the diazotrophic filamentous nonheterocystous cyanobacterium Trichodesmium spp. IMS101 in defined media: Evidence of a circadium rhythm. J. Phycol 1996, 32, 916–923. [Google Scholar]
- Currin, C.A.; Paerl, H.W.; Suba, G.K.; Alberte, R.S. Immunofluorescence detection and characterization of N2-fixing microorganisms from aquatic environments. Limnol. Oceanogr 1990, 35, 59–71. [Google Scholar]
- Biegala, I.C.; Raimbault, P. High abundance of diazotrophic picocyanobacteria (<3μm) in a Southwest Pacific coral lagoon. Aquat. Microb. Ecol 2008, 51, 45–53. [Google Scholar]
- El-Shehawy, R.; Lugomela, C.; Ernst, A.; Bergman, B. Diurnal expression of hetR and diazocyte development in the filamentous non-heterocystous cyanobacteria Trichodesmium erythraeum. Microbiology 2003, 149, 1169–1146. [Google Scholar]
- Berman-Frank, I.; Lundgren, P.; Chen, Y-B.; Küpper, H.; Kolber, K.; Bergman, B.; Falkowski, P. Segregation of nitrogen fixation and oxygenic photosynthesis in the marine cyanobacterium Trichodesmium. Science 2001, 294, 1534–1537. [Google Scholar]
- Janson, S.; Carpenter, E.J.; Bergman, B. Compartmentalization of nitrogenase in a non-heterocystous cyanobacterium: Trichodesmium contortum. FEMS Microbiol. Lett 1994, 118, 9–14. [Google Scholar]
- Fredriksson, C.; Bergman, B. Ultrastructural characterization of cells specialized for nitrogen fixation in a non-heterocystous cyanobaterium Trichodesmium spp. Protoplasma 1997, 197, 76–85. [Google Scholar]
- Paerl, H.W.; Priscu, J.C.; Brawner, D.L. Immunochemical localization of nitrogenase in marine Trichodesmium aggregates: Relationship to N2 fixation potential. Appl. Environ. Microbiol 1989, 55, 2965–2975. [Google Scholar]
- Orcutt, K.M.; Gundersen, K.; Wells, M.L.; Poulton, N.; Sieracki, M.E.; Smith, G.J. Lighting up phytoplankton cells with quantum dots. Limnol. Oceanogr 2008, 6, 653–658. [Google Scholar]


© 2009 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Orcutt, K.M.; Ren, S.; Gundersen, K. Detecting Proteins in Highly Autofluorescent Cells Using Quantum Dot Antibody Conjugates. Sensors 2009, 9, 7540-7549. https://doi.org/10.3390/s90907540
Orcutt KM, Ren S, Gundersen K. Detecting Proteins in Highly Autofluorescent Cells Using Quantum Dot Antibody Conjugates. Sensors. 2009; 9(9):7540-7549. https://doi.org/10.3390/s90907540
Chicago/Turabian StyleOrcutt, Karen M., Shanshan Ren, and Kjell Gundersen. 2009. "Detecting Proteins in Highly Autofluorescent Cells Using Quantum Dot Antibody Conjugates" Sensors 9, no. 9: 7540-7549. https://doi.org/10.3390/s90907540