2.3 Xanthohumol content in hop leaves
The content of xanthohumol in hop leaves and hop cones differ in orders of magnitude [
6]. The economic interest for xanthohumol or other polyphenols in hop leaves would be justified if with a simple stress such as drought or with crossbreeding their contents were increased.
Low contents of xanthohumol were detected in hop leaves from the pot and field experiment (
Tables 1 and
2); the highest in leaves of cv. Taurus (0.08% in DM), followed by cv. Southern Star. For the plants which were regularly watered, xanthohumol content in leaves was mainly lower, compared to plants which were exposed to drought stress and plants watered naturally by the rain outdoors. In the pot experiment, xanthohumol content in leaves was higher at the beginning of August compared to the middle of July, then the trend changed as to cultivar and water treatment.
Similarly, in the field experiment, xanthohumol content in leaves was the highest in cv. Taurus too (
Table 2). In the middle of July xanthohumol content was 2.4 to 8.4-times higher in the leaves of that cultivar compared to other included cultivars. Xanthohumol content in leaves decreased from the middle of July to the beginning of August (the period was dry and hot), except for cv. Merkur and cv. 279D112, where the content increased, and for cv. Taurus, where the content was stable during that period. For all cultivars included in this study xanthohumol content in leaves decreased from the beginning of August to the middle of August (that period was relatively cold and wet, the hop cones were developing intensively).
2.4 Polyphenol content in hop leaves
Polyphenol content in leaves of plants grown in the pots increased from the middle of July to the beginning of August and then decreased for all cultivars and all water treatments (
Tables 3 and
4). There were differences among cultivars; the highest content was found at South African cultivar Southern Star (as high as 14.28 g/kg DM), followed by the new Slovenian cultivar 279D112 (7.24 g/kg DM), Aurora and Cicero (Slovene cultivars) (
Table 3). For the majority of cultivars the highest polyphenol content was determined in the leaves of plants that were not watered. The exceptions were cv. Taurus and crossbread 279/122, where the highest polyphenol content in leaves was detected at plants which were watered as indicated by the precipitation and cv. 279D112 where the highest polyphenol content in leaves was detected at plants watered regularly.
The results in the field experiment were rather similar: polyphenol content in leaves increased from the middle of July to the beginning of August and then decreased for all cultivars. The highest contents in the field were measured in cvs. Southern Star, 279D112 and Taurus. The lowest contents were measured for cvs. Celeia and Aurora. (
Table 4)
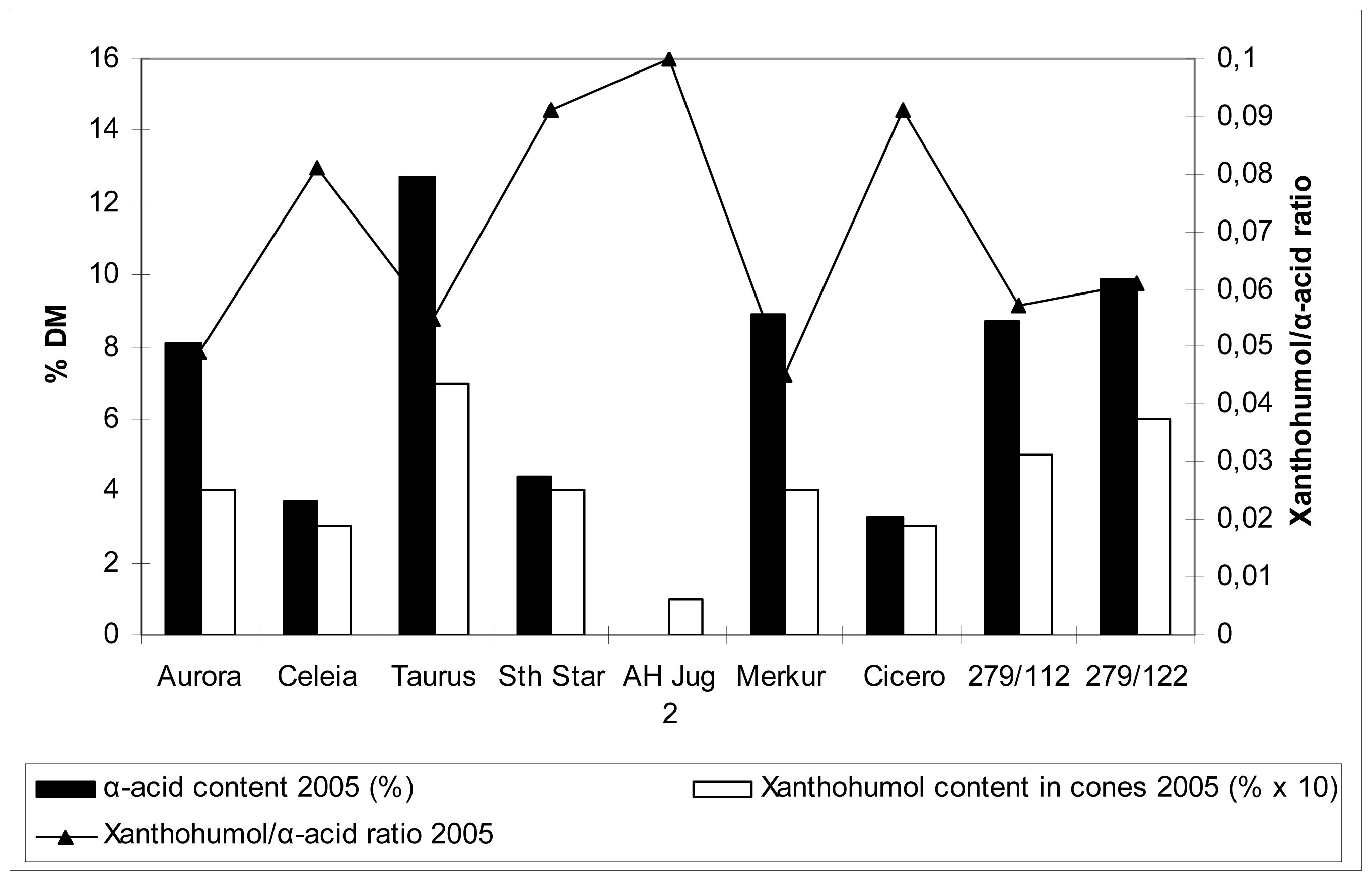
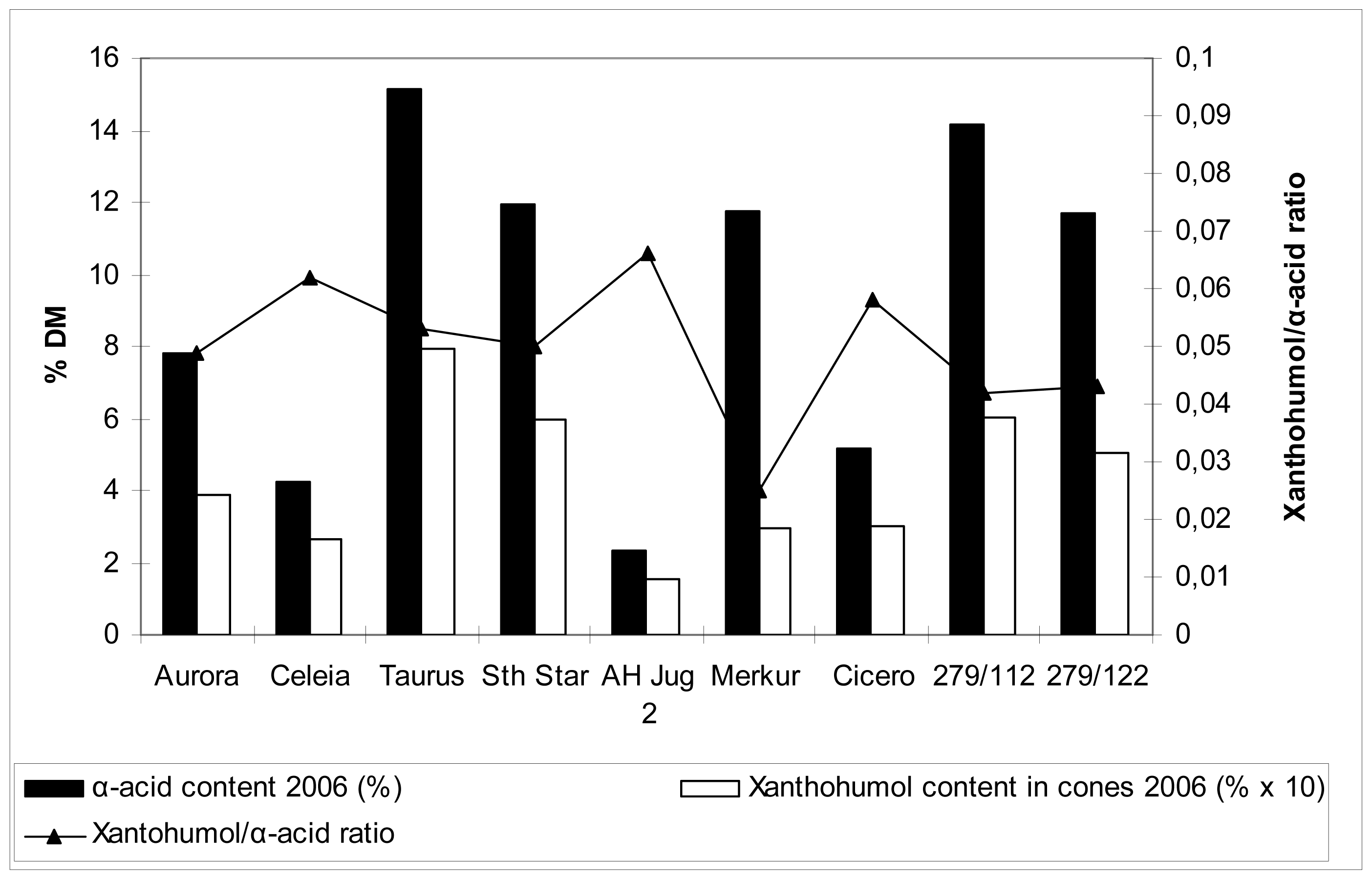
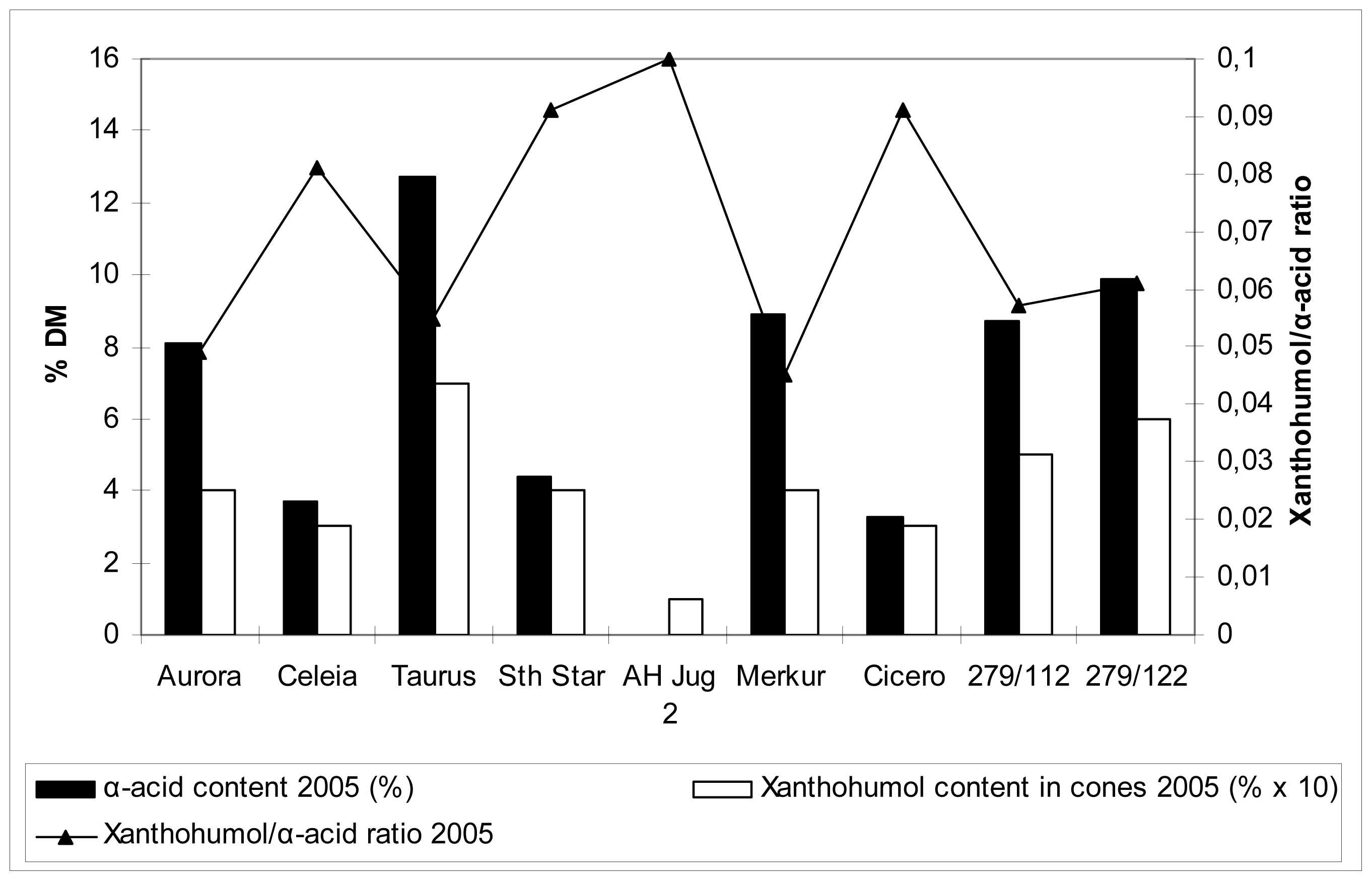
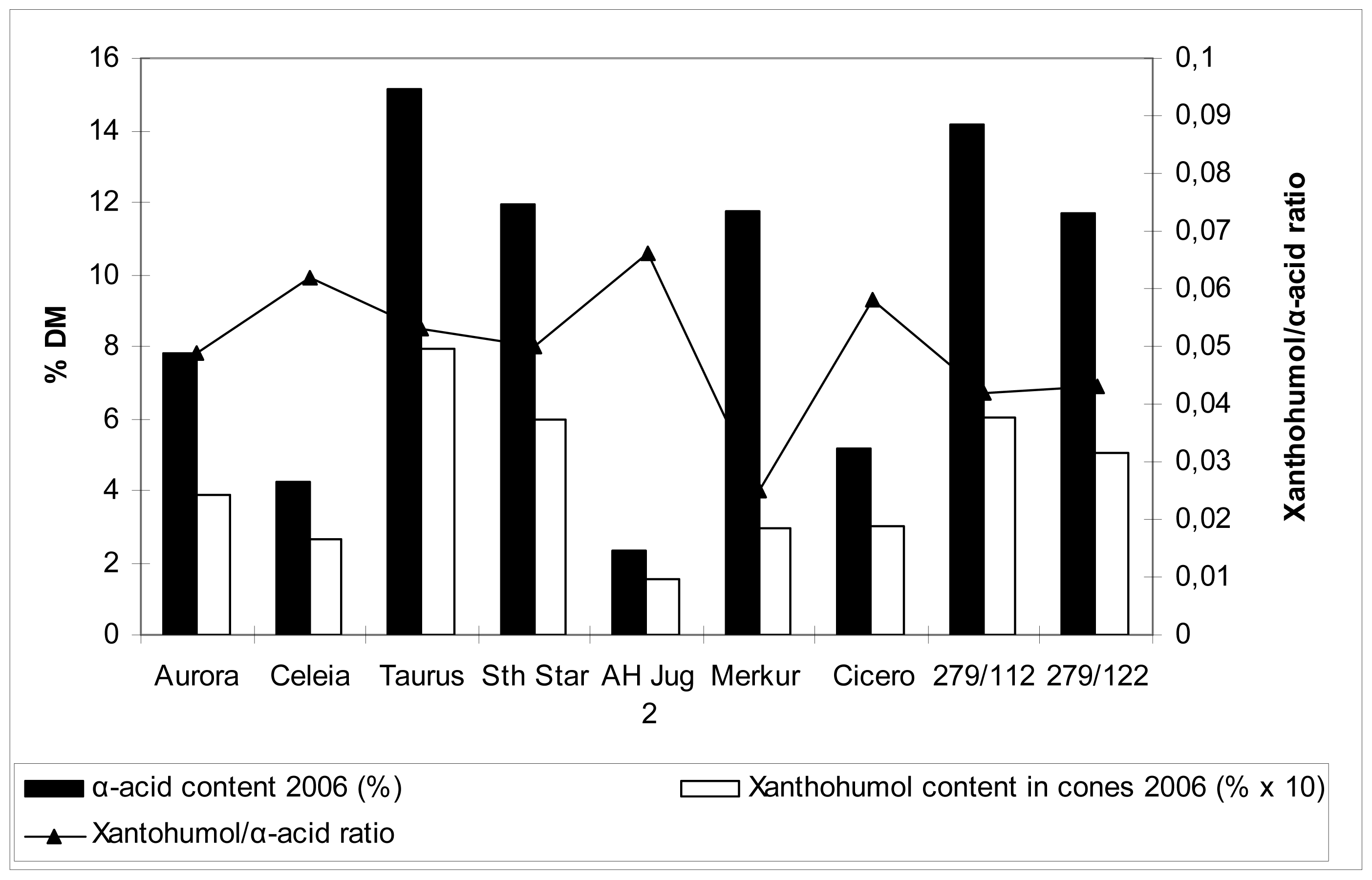
2.5 Xanthohumol, polyphenol and α-acid content in hop cones
Table 5 and
Figures 1 and
2 show the data on xanthohumol and polyphenol content in cones, together with relevant comparison with α-acid content. These values are far more familiar compared to those for the content of the same substances in the leaves, so no detailed commentary is included. Generally speaking, there were differences among cultivars in polyphenol content in hop cones. Higher content was detected for cvs. Merkur, Aurora, Celeia and Southern Star, lower for cvs. Taurus, Cicero and at the autochtonous hop Jug2. Higher content of polyphenols in leaves did not necessarily reflect in higher content of polyphenols in cones.
Xanthohumol is the principal simple prenylated chalcone that occurs only in the hop plant. It is secreted as part of the hop resin and is accompanied by at least 13 related chalcones [
13]. Although prenylflavonoids have a restricted distribution, the biosynthesis of flavonoids is well characterized at both genetic and enzymatic levels. It has been proven in many plants that flavonoids protect them from UV light [
14].
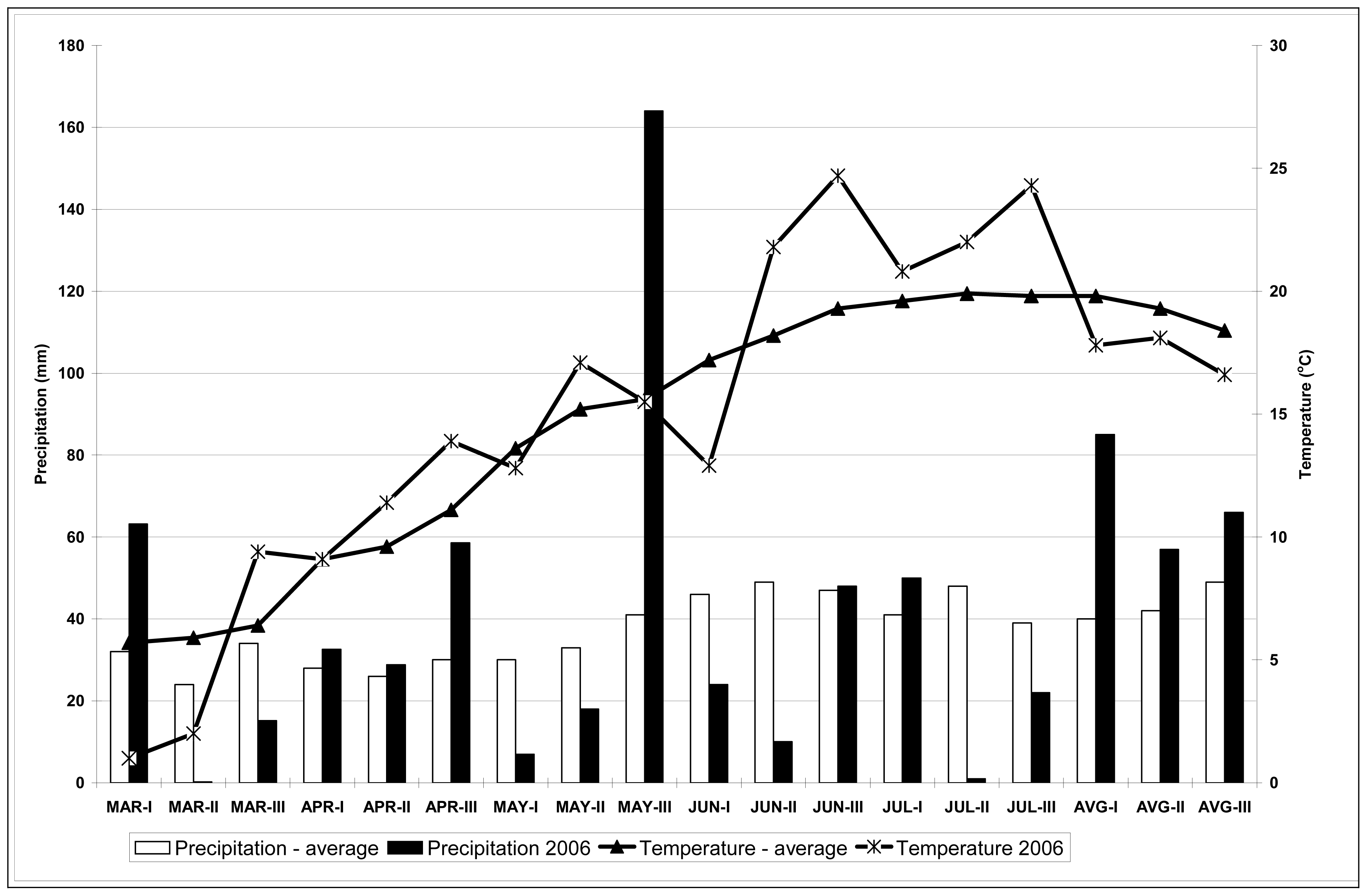
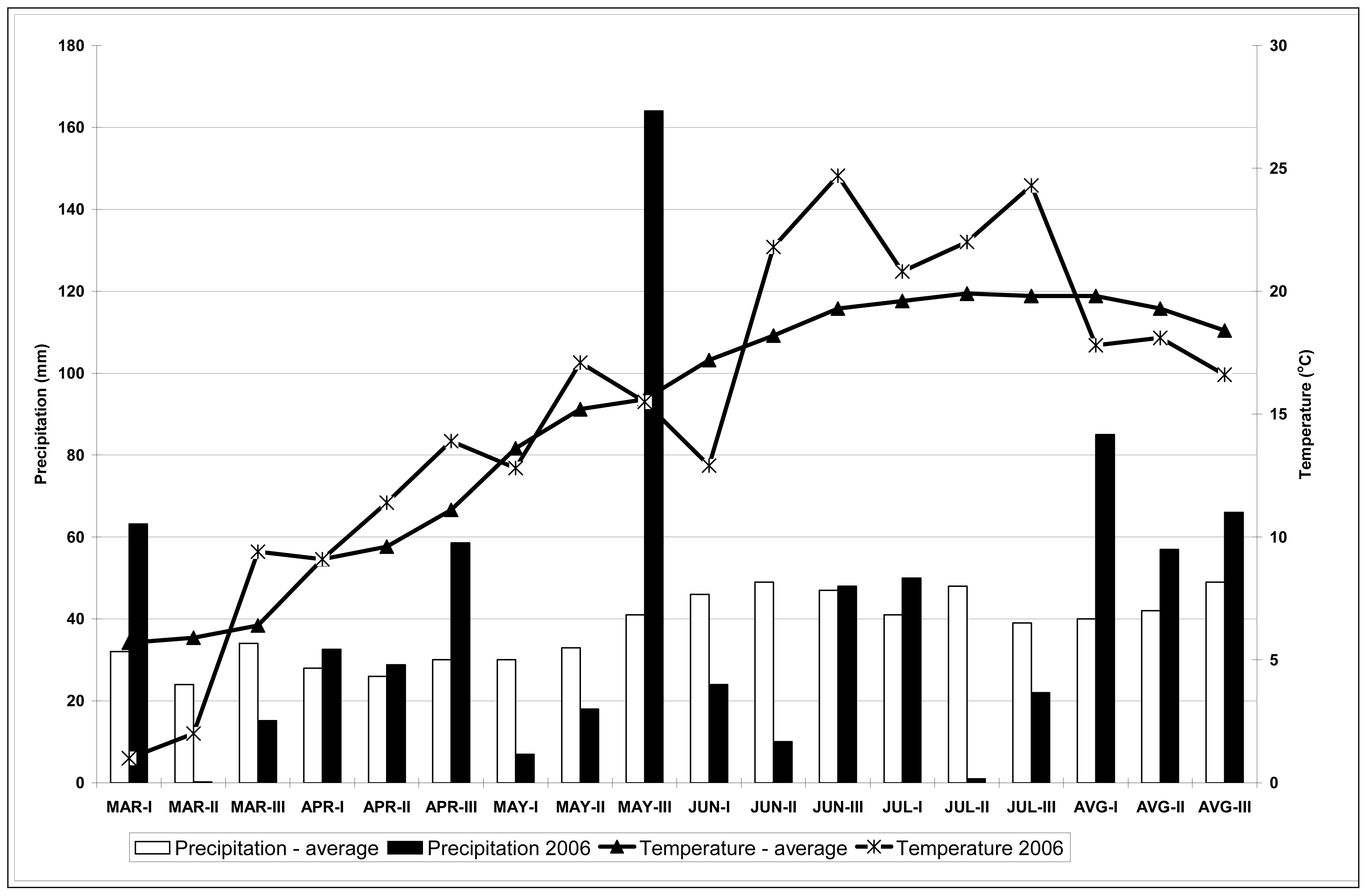
The 2006 season was a good one to investigate drought stress impact on the content of secondary metabolites in hops grown in the field (in our case the content of polyphenols and xanthohumol) in hop leaves and cones. Precipitation was low in July, accompanied by temperatures over 30 °C. In the period from the middle of July to the beginning of August the content of polyphenols in leaves increased for all cultivars included in this study, while that of xanthohumol decreased. The only exceptions being the cv. 279D112 and cv. Merkur, where xanthohumol content in leaves increased during the period in question. Xanthohumol in leaves reached values up to 0.009% DM, while polyphenol content in leaves ranged from 0.57 g/kg DM to 14.28 g/kg DM. Higher values were reached at plants in pots compared to the plants on the field because drought stress was certainly higher for plants in pots compared to the plants in the field where absorption of water was possible from lower layers of soil; there was also 22 mm of precipitation in the second and the third decade of July and the wind lowered the temperatures of the leaves.
Cvs. Merkur and Cicero, which are cultivars less adapted to drought, did not differ in contents of polyphenols and xanthohumol in leaves compared to the other investigated cultivars that are known to be more drought adapted (i. e. less susceptible to drought).
To our best knowledge this is the first systematic study of the effect of water supply on the content of polyphenols and especially on the content of xanthohumol in hops. As water supply (i. e. water or drought stress) is hard to define under field conditions, we combined the field experiment with a pot experiment. Consequently, we had to face the fact, that cones are not always obtainable under such (namely the pot experiment) conditions. We had to compromise and so results presented in this study give xanthohumol and total polyphenol content for hop leaves as well as for hop cones. Leaves were also the only alternative if any data on the time dependence of these substances should be obtained. Some analytical problems emerged especially because the substances under investigation are present in leaves only in rather low concentrations.
Nevertheless, one could conclude that the metabolism and above all the relative content of secondary metabolites in hops depends more on the cultivar than on the growing conditions. In this experiment we noticed that such a simple stress as drought cannot increase much the content of xanthohumol and total polyphenolics. In other words when higher content of xanthohumol or other polyphenols of interest in hop leaves are an aim then metabolic engineering of prenylflavonoid biosynthesis for developing such hop varieties is a necessity. It looks like we are on a right track with the new cultivar 279D112 and our crossbread 279/122.


{kind=link}
{kind=link}
{kind=link}