New Insights Contributing to the Development of Effective Vaccines and Therapies to Reduce the Pathology Caused by hRSV


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. General Features of Human Respiratory Syncytial Virus (hRSV)
2. hRSV Epidemiology
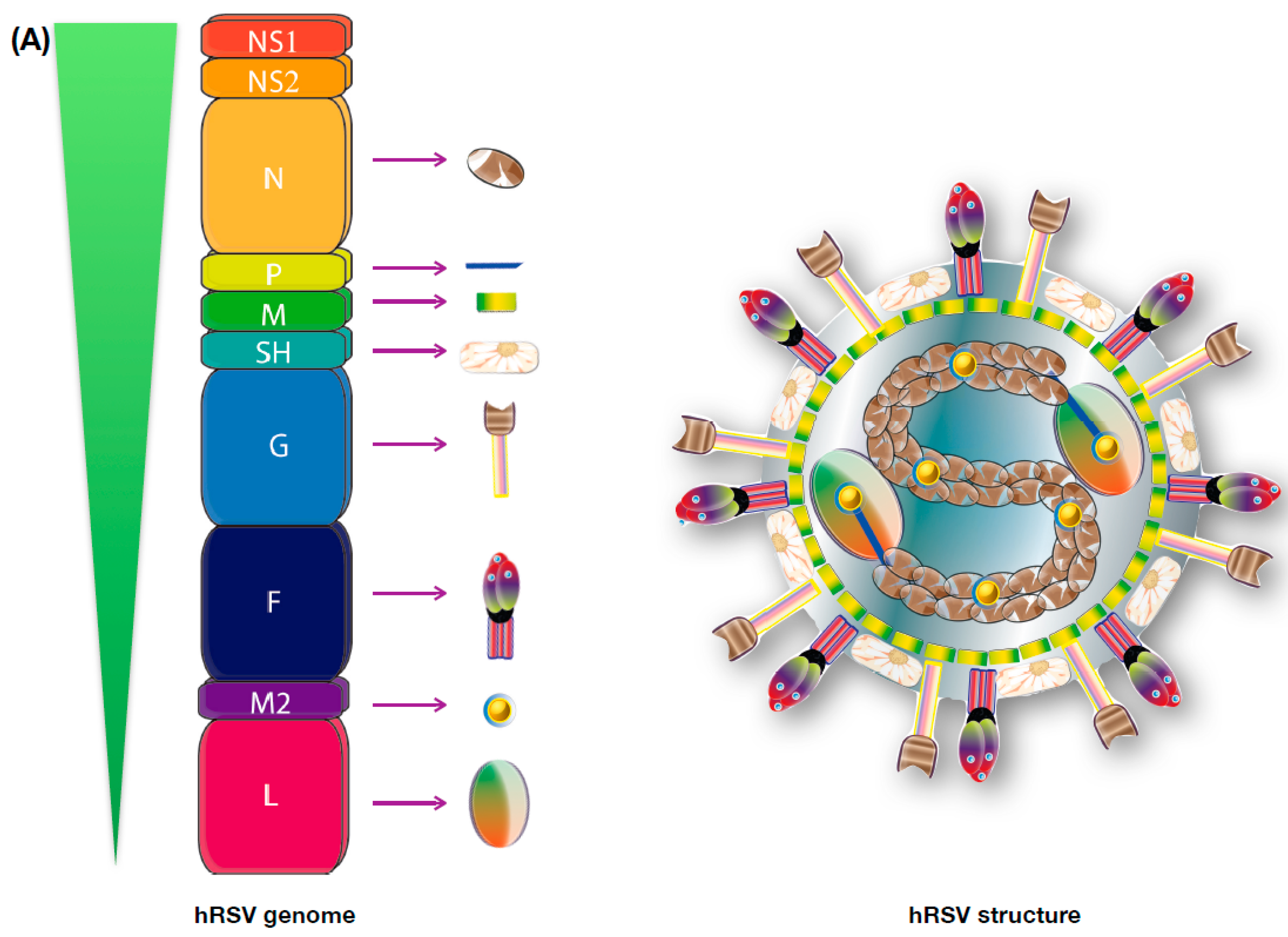
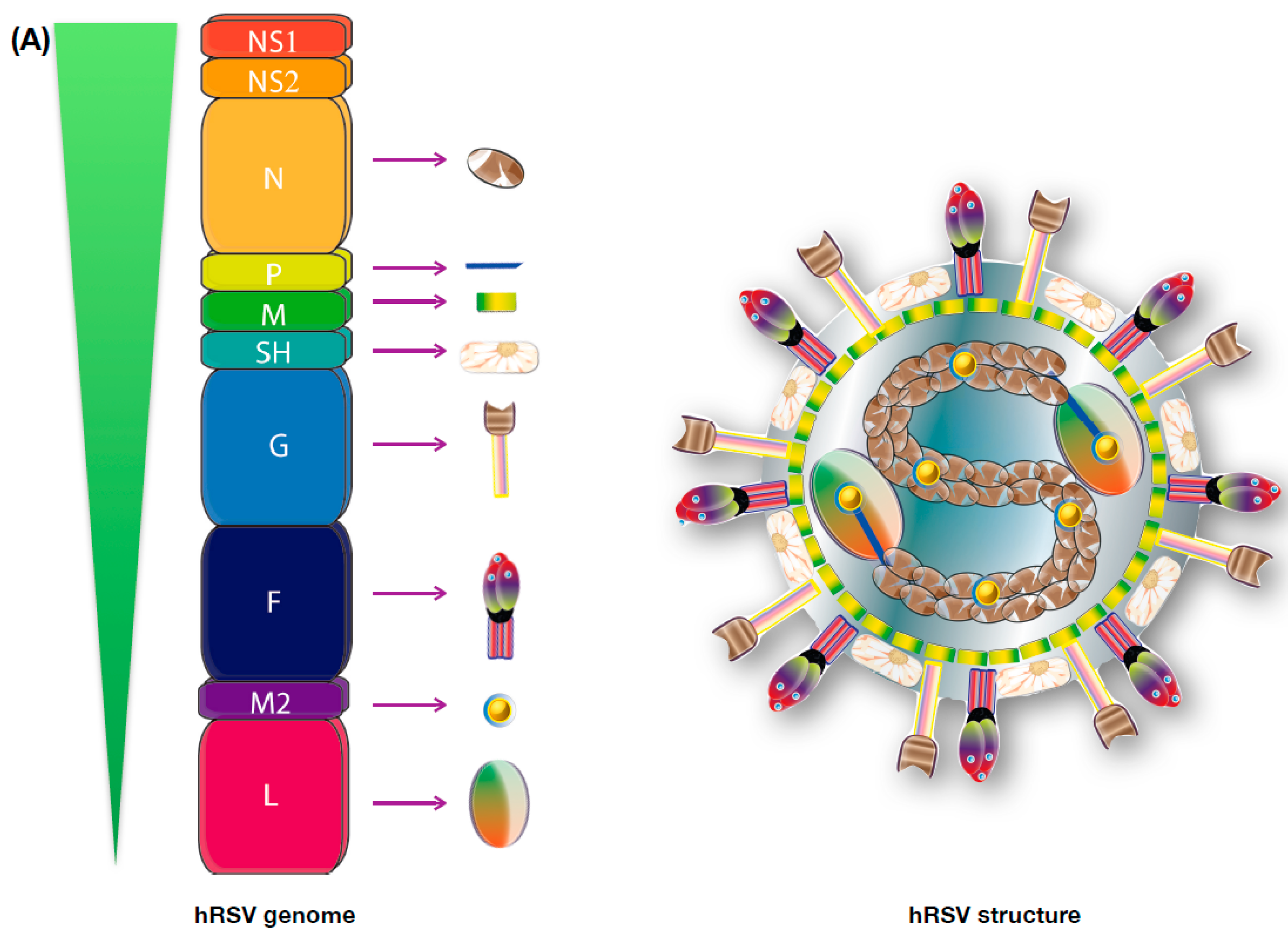
3. Viral Genome and Protein Function
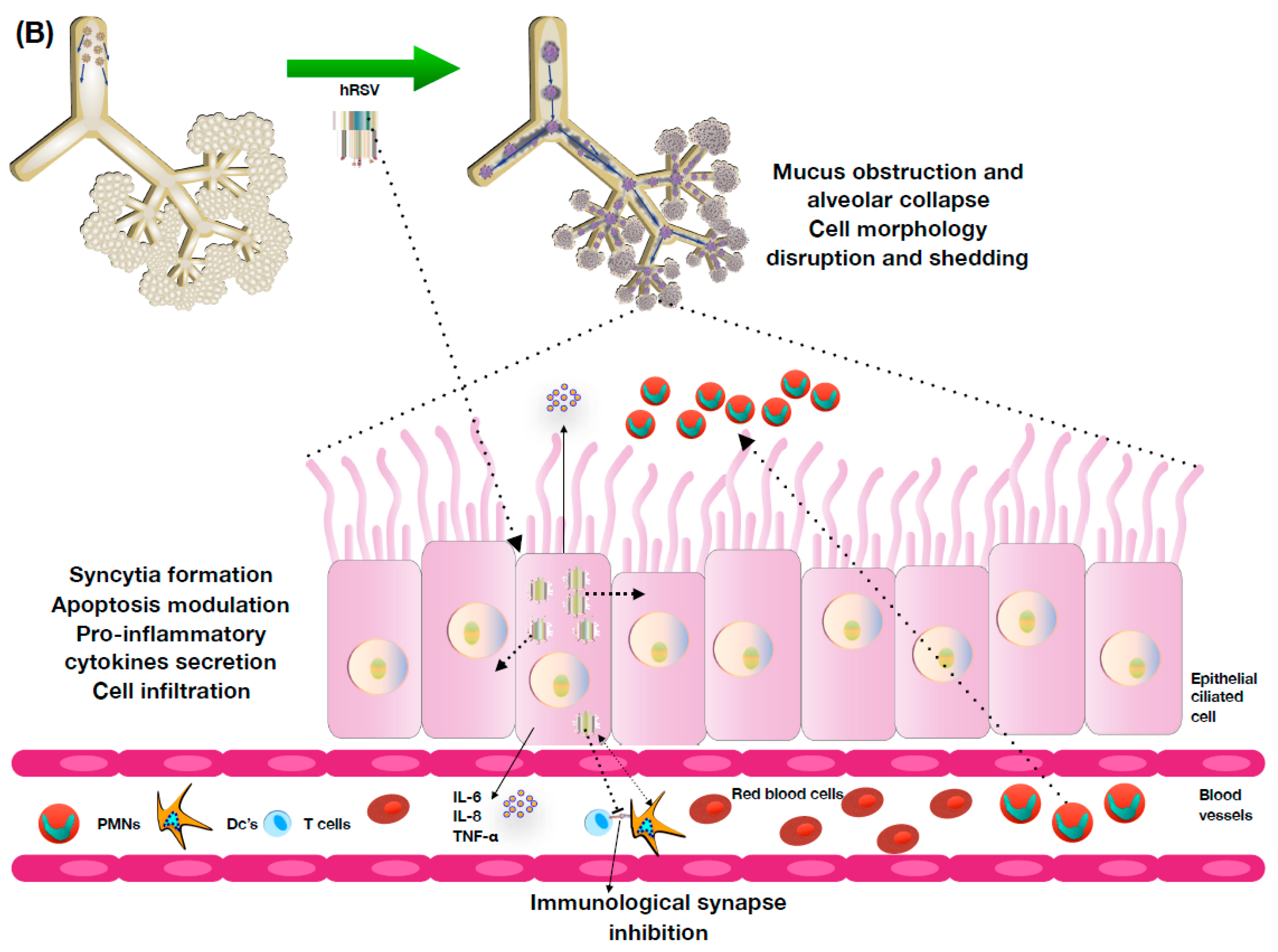
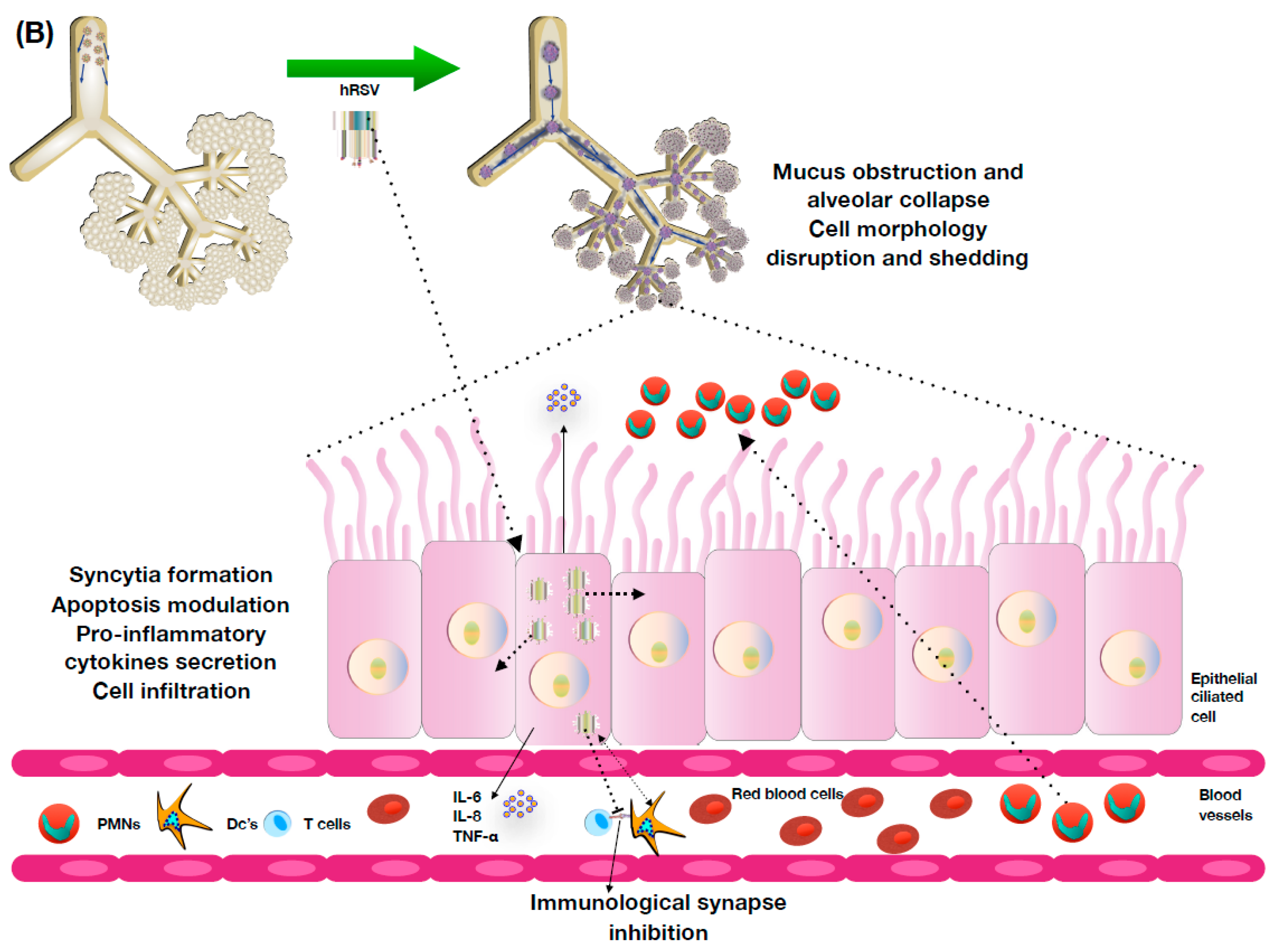
4. hRSV Pathology and Host Immune Response
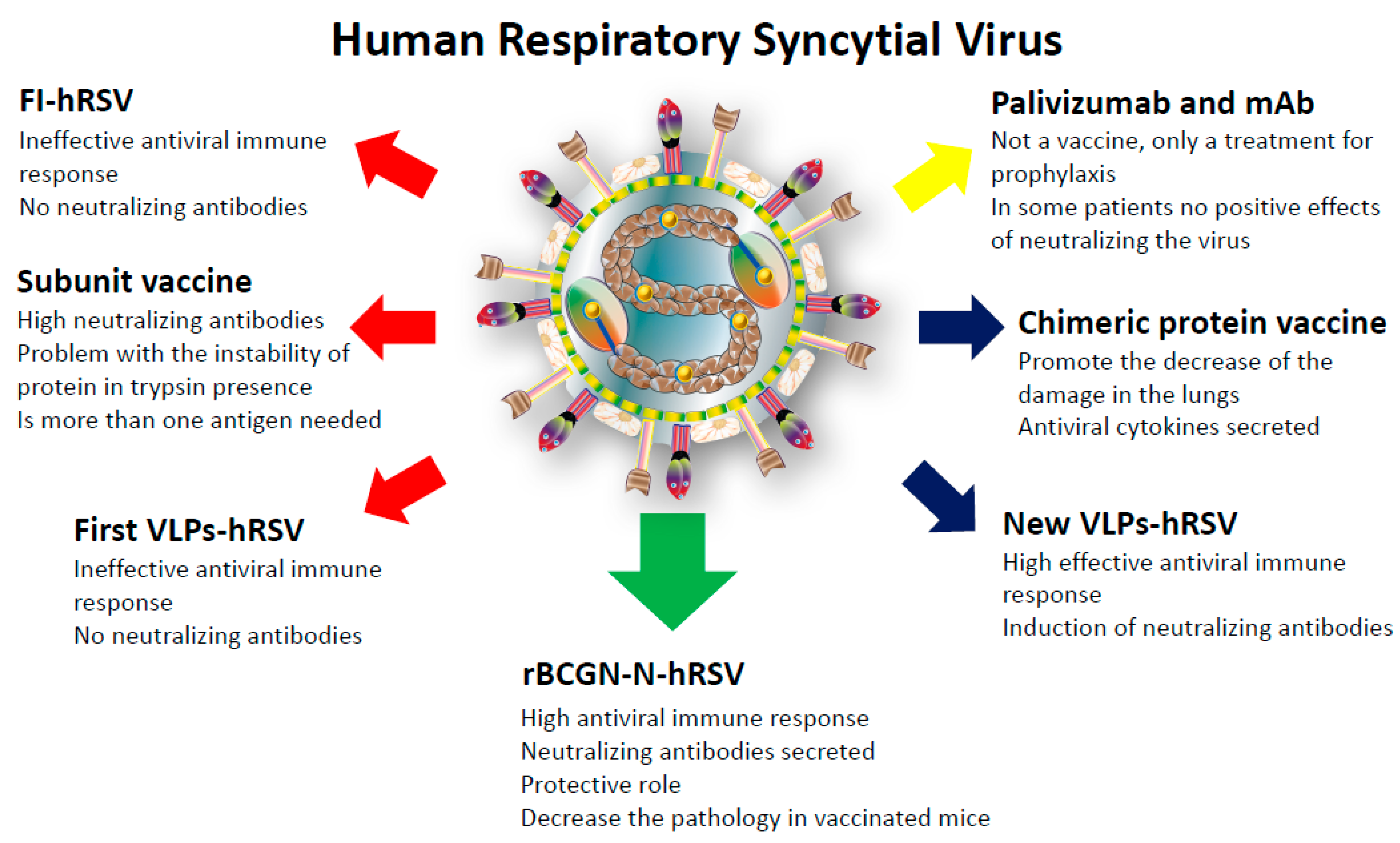
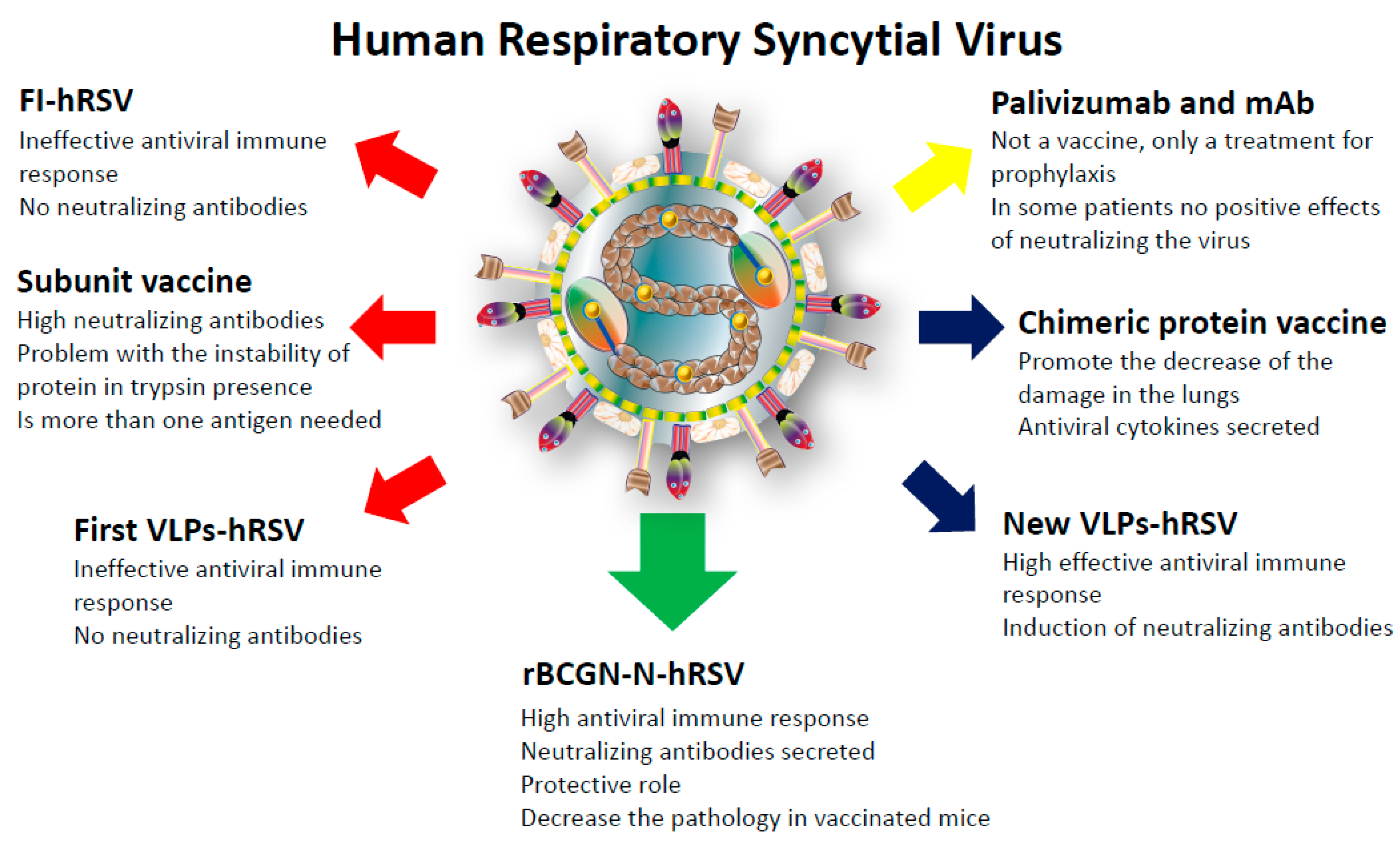
5. Prophylaxis and Vaccine Development
6. The rBCG-N-hRSV Vaccine
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| ALRTI | Acute lower respiratory tract infections |
| BCG | Mycobacterium bovis Bacillus Calmette-Guérin |
| DCs | Dendritic Cells |
| hMPV | Human Metapneumovirus |
| hRSV | Human Respiratory Syncytial Virus |
| IFN | Interferon |
| PMNs | Polymorphonuclear cells |
| rBCG-N | Recombinant Mycobacterium bovis Bacillus Calmette-Guérin |
| ssRNA | Single-stranded RNA |
References
- Afonso, C.L.; Amarasinghe, G.K.; Bányai, K.; Báo, Y.; Basler, C.F.; Bavari, S.; Bejerman, N.; Blasdell, K.R.; Briand, F.X.; Briese, T.; et al. Taxonomy of the order Mononegavirales: Update 2016. Arch. Virol. 2016, 161, 2351–2360. [Google Scholar] [CrossRef] [PubMed]
- Monto, A.S. Epidemiology of viral respiratory infections. Am. J. Med. 2002, 112 (Suppl. S6A), 4S–12S. [Google Scholar] [CrossRef]
- Robinson, R.F. Impact of respiratory syncytial virus in the United States. Am. J. Health Syst. Pharm. 2008, 65, S3–S6. [Google Scholar] [CrossRef] [PubMed]
- Nair, H.; Nokes, D.J.; Gessner, B.D.; Dherani, M.; Madhi, S.A.; Singleton, R.J.; O’Brien, K.L.; Roca, A.; Wright, P.F.; Bruce, N.; et al. Global burden of acute lower respiratory infections due to respiratory syncytial virus in young children: A systematic review and meta-analysis. Lancet 2010, 375, 1545–1555. [Google Scholar] [CrossRef]
- Hall, C.B. Respiratory syncytial virus in young children. Lancet 2010, 375, 1500–1502. [Google Scholar] [CrossRef]
- Chanock, R.; Roizman, B.; Myers, R. Recovery from infants with respiratory illness of a virus related to chimpanzee coryza agent (CCA). I. Isolation, properties and characterization. Am. J. Hyg. 1957, 66, 281–290. [Google Scholar] [PubMed]
- Chanock, R.; Finberg, L. Recovery from infants with respiratory illness of a virus related to chimpanzee coryza agent (CCA). II. Epidemiologic aspects of infection in infants and young children. Am. J. Hyg. 1957, 66, 291–300. [Google Scholar] [PubMed]
- Wright, M.; Piedimonte, G. Respiratory syncytial virus prevention and therapy: Past, present, and future. Pediatr. Pulmonol. 2011, 46, 324–347. [Google Scholar] [CrossRef] [PubMed]
- Garcia, C.G.; Bhore, R.; Soriano-Fallas, A.; Trost, M.; Chason, R.; Ramilo, O.; Mejias, A. Risk Factors in Children Hospitalized With RSV Bronchiolitis Versus Non-RSV Bronchiolitis. Pediatrics 2010, 126, e1453–e1460. [Google Scholar] [CrossRef] [PubMed]
- Van Drunen Littel-Van Den Hurk, S.; Watkiss, E.R. Pathogenesis of respiratory syncytial virus. Curr. Opin. Virol. 2012, 2, 300–305. [Google Scholar] [CrossRef] [PubMed]
- Bendelja, K.; Gagro, A.; Bace, A.; Lokar-Kolbas, R.; Krsulovic-Hresic, V.; Drazenovic, V.; Mlinaric-Galinovic, G.; Rabatic, S. Predominant type-2 response in infants with respiratory syncytial virus (RSV) infection demonstrated by cytokine flow cytometry. Clin. Exp. Immunol. 2000, 121, 332–338. [Google Scholar] [CrossRef] [PubMed]
- Roman, M.; Calhoun, W.J.; Hinton, K.L.; Avendano, L.F.; Simon, V.; Escobar, A.M.; Gaggero, A.; Diaz, P.V. Respiratory syncytial virus infection in infants is associated with predominant Th-2-like response. Am. J. Respir. Crit. Care Med. 1997, 156, 190–195. [Google Scholar] [CrossRef] [PubMed]
- Becker, Y. Respiratory syncytial virus (RSV) evades the human adaptive immune system by skewing the Th1/Th2 cytokine balance toward increased levels of Th2 cytokines and IgE, markers of allergy—A review. Virus Genes 2006, 33, 235–252. [Google Scholar] [CrossRef] [PubMed]
- Puchkov, G.F.; Min’kovich, B.M. Respiratory syncytial infection in a child complicated by interstitial myocarditis with fatal outcome. Arkh. Patol. 1972, 34, 70–73. [Google Scholar] [PubMed]
- Suda, K.; Ito, M.; Abumi, K.; Haba, H.; Armstrong, D.S.; Menahem, S. Cardiac arrhythmias as a manifestation of acquired heart disease in association with paediatric respiratory syncitial virus infection. J. Paediatr. Child Health 1993, 29, 309–311. [Google Scholar]
- Donnerstein, R.L.; Berg, R.A.; Shehab, Z.; Ovadia, M. Complex atrial tachycardias and respiratory syncytial virus infections in infants. J. Pediatr. 1994, 125, 23–28. [Google Scholar] [CrossRef]
- Eisenhut, M.; Thorburn, K. Hepatitis associated with severe respiratory syncytial virus-positive lower respiratory tract infection. Scand. J. Infect. Dis. 2002, 34, 235. [Google Scholar] [CrossRef] [PubMed]
- Eisenhut, M.; Thorburn, K.; Ahmed, T. Transaminase levels in ventilated children with respiratory syncytial virus bronchiolitis. Intensive Care Med. 2004, 30, 931–934. [Google Scholar] [CrossRef] [PubMed]
- Bohmwald, K.; Espinoza, J.A.; González, P.A.; Bueno, S.M.; Riedel, C.A.; Kalergis, A.M. Central nervous system alterations caused by infection with the human respiratory syncytial virus. Rev. Med. Virol. 2014, 24, 407–419. [Google Scholar] [CrossRef] [PubMed]
- Bohmwald, K.; Espinoza, J.A.; Becerra, D.; Rivera, K.; Lay, M.K.; Bueno, S.M.; Riedel, C.A.; Kalergis, A.M. Inflammatory damage on respiratory and nervous systems due to hRSV infection. Curr. Opin. Immunol. 2015, 36, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Eisenhut, M. Extrapulmonary manifestations of severe respiratory syncytial virus infection—A systematic review. Crit. Care 2006, 10, R107. [Google Scholar] [CrossRef] [PubMed]
- Hansbro, P.M.; Starkey, M.R.; Mattes, J.; Horvat, J.C. Pulmonary immunity during respiratory infections in early life and the development of severe asthma. Ann. Am. Thorac. Soc. 2014, 11, S297–S302. [Google Scholar] [CrossRef] [PubMed]
- KIM, H.W.; Canchola, J.G.; Brandt, C.D.; Pyles, G.; Chanock, R.M.; Jensen, K.; Parrott, R.H. Respiratory syncytial virus disease in infants despite prior administration of antigenic inactivated vaccine. Am. J. Epidemiol. 1969, 89, 422–434. [Google Scholar] [CrossRef] [PubMed]
- Welliver, R.C.; Kaul, T.N.; Sun, M.; Ogra, P.L. Defective regulation of immune responses in respiratory syncytial virus infection. J. Immunol. 1984, 133, 1925–1930. [Google Scholar] [PubMed]
- Welliver, R.C.; Wong, D.T.; Sun, M.; Middleton, E.; Vaughan, R.S.; Ogra, P.L. The development of respiratory syncytial virus-specific IgE and the release of histamine in nasopharyngeal secretions after infection. N. Engl. J. Med. 1981, 305, 841–846. [Google Scholar] [CrossRef] [PubMed]
- Munoz, F.M.; Piedra, P.A.; Glezen, W.P. Safety and immunogenicity of respiratory syncytial virus purified fusion protein-2 vaccine in pregnant women. Vaccine 2003, 21, 3465–3467. [Google Scholar] [CrossRef]
- Rivera, C.A.; Gomez, R.S.; Diaz, R.A.; Cespedes, P.F.; Espinoza, J.A.; Gonzalez, P.A.; Riedel, C.A.; Bueno, S.M.; Kalergis, A.M. Novel therapies and vaccines against the human respiratory syncytial virus. Expert Opin. Investig. Drugs 2015, 24, 1613–1630. [Google Scholar] [CrossRef] [PubMed]
- Raghunandan, R.; Lu, H.; Zhou, B.; Xabier, M.G.; Massare, M.J.; Flyer, D.C.; Fries, L.F.; Smith, G.E.; Glenn, G.M. An insect cell derived respiratory syncytial virus (RSV) F nanoparticle vaccine induces antigenic site II antibodies and protects against RSV challenge in cotton rats by active and passive immunization. Vaccine 2014, 32, 6485–6492. [Google Scholar] [CrossRef] [PubMed]
- Bueno, S.M.; González, P.A.; Cautivo, K.M.; Mora, J.E.; Leiva, E.D.; Tobar, H.E.; Fennelly, G.J.; Eugenin, E.A.; Jacobs, W.R.; Riedel, C.A.; et al. Protective T cell immunity against respiratory syncytial virus is efficiently induced by recombinant BCG. Proc. Natl. Acad. Sci. USA 2008, 105, 20822–20827. [Google Scholar] [CrossRef] [PubMed]
- Céspedes, P.F.; Rey-Jurado, E.; Espinoza, J.A.; Rivera, C.A.; Canedo-Marroquín, G.; Bueno, S.M.; Kalergis, A.M. A single, low dose of a cGMP recombinant BCG vaccine elicits protective T cell immunity against the human respiratory syncytial virus infection and prevents lung pathology in mice. Vaccine 2017, 35, 757–766. [Google Scholar] [CrossRef] [PubMed]
- Sande, C.J.; Mutunga, M.N.; Medley, G.F.; Cane, P.A.; Nokes, D.J. Group and Genotype-Specific Neutralizing Antibody Responses Against Respiratory Syncytial Virus in Infants and Young Children With Severe Pneumonia. J. Infect. Dis. 2013, 207, 489–492. [Google Scholar] [CrossRef] [PubMed]
- RSV | Research and Surveillance | Respiratory Syncytial Virus | CDC. Available online: https://www.cdc.gov/rsv/research/index.html (accessed on 29 June 2017).
- (WHO) WHO Global Respiratory Syncytial Virus Surveillance. Available online: http://www.who.int/influenza/rsv/en/ (accessed on 29 June 2017).
- Iwane, M.K.; Farnon, E.C.; Gerber, S.I. Importance of Global Surveillance for Respiratory Syncytial Virus. J. Infect. Dis. 2013, 208, S165–S166. [Google Scholar] [CrossRef] [PubMed]
- Paramore, L.C.; Ciuryla, V.; Ciesla, G.; Liu, L. Economic impact of respiratory syncytial virus-related illness in the US: An analysis of national databases. Pharmacoeconomics 2004, 22, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Gomez, R.S.; Guisle-Marsollier, I.; Bohmwald, K.; Bueno, S.M.; Kalergis, A.M. Respiratory Syncytial Virus: Pathology, therapeutic drugs and prophylaxis. Immunol. Lett. 2014, 162, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Palavecino, C.E.; Céspedes, P.F.; Gómez, R.S.; Kalergis, A.M.; Bueno, S.M.; Cespedes, P.F.; Gomez, R.S.; Kalergis, A.M.; Bueno, S.M. Immunization with a recombinant bacillus Calmette-Guerin strain confers protective Th1 immunity against the human metapneumovirus. J. Immunol. 2014, 192, 214–223. [Google Scholar] [CrossRef] [PubMed]
- Hacking, D.; Hull, J. Respiratory syncytial virus—Viral biology and the host response. J. Infect. 2002, 45, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Collins, P.L.; Chanock, R.M.; Murphy, B.R. Respiratory syncytial virus. Fields Virol. 2016, 1, 1443–1486. [Google Scholar] [CrossRef]
- Pretel, E.; Camporeale, G.; de Prat-Gay, G. The Non-Structural NS1 Protein Unique to Respiratory Syncytial Virus: A Two-State Folding Monomer in Quasi-Equilibrium with a Stable Spherical Oligomer. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Ling, Z.; Tran, K.C.; Teng, M.N. Human Respiratory Syncytial Virus Nonstructural Protein NS2 Antagonizes the Activation of Beta Interferon Transcription by Interacting with RIG-I. J. Virol. 2009, 83, 3734–3742. [Google Scholar] [CrossRef] [PubMed]
- Lo, M.S.; Brazas, R.M.; Holtzman, M.J. Respiratory Syncytial Virus Nonstructural Proteins NS1 and NS2 Mediate Inhibition of Stat2 Expression and α/β Interferon Responsiveness. J. Virol. 2005, 79, 9315–9319. [Google Scholar] [CrossRef] [PubMed]
- Liesman, R.M.; Buchholz, U.J.; Luongo, C.L.; Yang, L.; Proia, A.D.; DeVincenzo, J.P.; Collins, P.L.; Pickles, R.J. RSV-encoded NS2 promotes epithelial cell shedding and distal airway obstruction. J. Clin. Investig. 2014, 124, 2219–2233. [Google Scholar] [CrossRef] [PubMed]
- Ramaswamy, M.; Shi, L.; Varga, S.M.; Barik, S.; Behlke, M.A.; Look, D.C. Respiratory syncytial virus nonstructural protein 2 specifically inhibits type I interferon signal transduction. Virology 2006, 344, 328–339. [Google Scholar] [CrossRef] [PubMed]
- Lay, M.K.; Bueno, S.M.; Gálvez, N.; Riedel, C.A.; Kalergis, A.M. New insights on the viral and host factors contributing to the airway pathogenesis caused by the respiratory syncytial virus. Crit. Rev. Microbiol. 2015, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Liljeroos, L.; Krzyzaniak, M.A.; Helenius, A.; Butcher, S.J. Architecture of respiratory syncytial virus revealed by electron cryotomography. Proc. Natl. Acad. Sci. USA 2013, 110, 11133–11138. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, P.A.; Prado, C.E.; Leiva, E.D.; Carreno, L.J.; Bueno, S.M.; Riedel, C.A.; Kalergis, A.M. Respiratory syncytial virus impairs T cell activation by preventing synapse assembly with dendritic cells. Proc. Natl. Acad. Sci. USA 2008, 105, 14999–15004. [Google Scholar] [CrossRef] [PubMed]
- Céspedes, P.F.; Bueno, S.M.; Ramírez, B.A.; Gomez, R.S.; Riquelme, S.A.; Palavecino, C.E.; Mackern-Oberti, J.P.; Mora, J.E.; Depoil, D.; Sacristán, C.; et al. Surface expression of the hRSV nucleoprotein impairs immunological synapse formation with T cells. Proc. Natl. Acad. Sci. USA 2014, 111, E3214–E3223. [Google Scholar] [CrossRef] [PubMed]
- Collins, P.L.; Melero, J.A. Progress in understanding and controlling respiratory syncytial virus: Still crazy after all these years. Virus Res. 2011, 162, 80–99. [Google Scholar] [CrossRef] [PubMed]
- Ogra, P.L. Respiratory syncytial virus: The virus, the disease and the immune response. Paediatr. Respir. Rev. 2004, 5 (Suppl. A), S119–S126. [Google Scholar] [CrossRef]
- Mastrangelo, P.; Hegele, R.G. RSV Fusion: Time for a new model. Viruses 2013, 5, 873–885. [Google Scholar] [CrossRef] [PubMed]
- Gould, P.S.; Easton, A.J. Coupled Translation of the Second Open Reading Frame of M2 mRNA Is Sequence Dependent and Differs Significantly within the Subfamily Pneumovirinae. J. Virol. 2007, 81, 8488–8496. [Google Scholar] [CrossRef] [PubMed]
- Batonick, M.; Wertz, G.W. Requirements for human respiratory syncytial virus glycoproteins in assembly and egress from infected cells. Adv. Virol. 2011, 2011. [Google Scholar] [CrossRef] [PubMed]
- Domurat, F.; Roberts, N.J.; Walsh, E.E.; Dagan, R. Respiratory syncytial virus infection of human mononuclear leukocytes in vitro and in vivo. J. Infect. Dis. 1985, 152, 895–902. [Google Scholar] [CrossRef] [PubMed]
- Yui, I.; Hoshi, A.; Shigeta, Y.; Takami, T.; Nakayama, T. Detection of human respiratory syncytial virus sequences in peripheral blood mononuclear cells. J. Med. Virol. 2003, 70, 481–489. [Google Scholar] [CrossRef] [PubMed]
- Tayyari, F.; Marchant, D.; Moraes, T.J.; Duan, W.; Mastrangelo, P.; Hegele, R.G. Identification of nucleolin as a cellular receptor for human respiratory syncytial virus. Nat. Med. 2011, 17, 1132–1135. [Google Scholar] [CrossRef] [PubMed]
- Lay, M.K.; Gonzalez, P.A.; Leon, M.A.; Cespedes, P.F.; Bueno, S.M.; Riedel, C.A.; Kalergis, A.M. Advances in understanding respiratory syncytial virus infection in airway epithelial cells and consequential effects on the immune response. Microbes Infect. 2013, 15, 230–242. [Google Scholar] [CrossRef] [PubMed]
- McLellan, J.S.; Chen, M.; Leung, S.; Graepel, K.W.; Du, X.; Yang, Y.; Zhou, T.; Baxa, U.; Yasuda, E.; Beaumont, T.; et al. Structure of RSV fusion glycoprotein trimer bound to a prefusion-specific neutralizing antibody. Science 2013, 340, 1113–1117. [Google Scholar] [CrossRef] [PubMed]
- Corti, D.; Bianchi, S.; Vanzetta, F.; Minola, A.; Perez, L.; Agatic, G.; Guarino, B.; Silacci, C.; Marcandalli, J.; Marsland, B.J.; et al. Cross-neutralization of four paramyxoviruses by a human monoclonal antibody. Nature 2013, 501, 439–443. [Google Scholar] [CrossRef] [PubMed]
- Satake, M.; Coligan, J.E.; Elango, N.; Norrby, E.; Venkatesan, S. Respiratory syncytial virus envelope glycoprotein (G) has a novel structure. Nucleic Acids Res. 1985, 13, 7795–7812. [Google Scholar] [CrossRef] [PubMed]
- Wertz, G.W.; Collins, P.L.; Huang, Y.; Gruber, C.; Levine, S.; Ball, L.A. Nucleotide sequence of the G protein gene of human respiratory syncytial virus reveals an unusual type of viral membrane protein. Proc. Natl. Acad. Sci. USA 1985, 82, 4075–4079. [Google Scholar] [CrossRef] [PubMed]
- Magro, M.; Mas, V.; Chappell, K.; Vazquez, M.; Cano, O.; Luque, D.; Terron, M.C.; Melero, J.A.; Palomo, C. Neutralizing antibodies against the preactive form of respiratory syncytial virus fusion protein offer unique possibilities for clinical intervention. Proc. Natl. Acad. Sci. USA 2012, 109, 3089–3094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, B.R.; Walsh, E.E. Formalin-inactivated respiratory syncytial virus vaccine induces antibodies to the fusion glycoprotein that are deficient in fusion-inhibiting activity. J. Clin. Microbiol. 1988, 26, 1595–1597. [Google Scholar] [PubMed]
- Freitas, G.R.O.; Silva, D.A.O.; Yokosawa, J.; Paula, N.T.; Costa, L.F.; Carneiro, B.M.; Ribeiro, L.Z.G.; Oliveira, T.F.M.; Mineo, J.R.; Queiróz, D.A.O. Antibody response and avidity of respiratory syncytial virus-specific total IgG, IgG1, and IgG3 in young children. J. Med. Virol. 2011, 83, 1826–1833. [Google Scholar] [CrossRef] [PubMed]
- Quan, F.-S.; Kim, Y.; Lee, S.; Yi, H.; Kang, S.-M.; Bozja, J.; Moore, M.L.; Compans, R.W. Viruslike particle vaccine induces protection against respiratory syncytial virus infection in mice. J. Infect. Dis. 2011, 204, 987–995. [Google Scholar] [CrossRef] [PubMed]
- Ravanel, K.; Castelle, C.; Defrance, T.; Wild, T.F.; Charron, D.; Lotteau, V.; Rabourdin-Combe, C. Measles virus nucleocapsid protein binds to FcgammaRII and inhibits human B cell antibody production. J. Exp. Med. 1997, 186, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Griffin, D.E. Measles virus-induced suppression of immune responses. Immunol. Rev. 2010, 236, 176–189. [Google Scholar] [CrossRef] [PubMed]
- Tregoning, J.S.; Schwarze, J. Respiratory viral infections in infants: Causes, clinical symptoms, virology, and immunology. Clin. Microbiol. Rev. 2010, 23, 74–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, R.K.; Foster, P.S.; Rosenberg, H.F. Respiratory viral infection, epithelial cytokines, and innate lymphoid cells in asthma exacerbations. J. Leukoc. Biol. 2014, 96, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Munywoki, P.K.; Koech, D.C.; Agoti, C.N.; Bett, A.; Cane, P.A.; Medley, G.F.; Nokes, D.J. Frequent Asymptomatic Respiratory Syncytial Virus Infections during an Epidemic in a Rural Kenyan Household Cohort. J. Infect. Dis. 2015, 212, 1711–1718. [Google Scholar] [CrossRef] [PubMed]
- Kusel, M.M.H.; de Klerk, N.H.; Holt, P.G.; Kebadze, T.; Johnston, S.L.; Sly, P.D. Role of respiratory viruses in acute upper and lower respiratory tract illness in the first year of life: A birth cohort study. Pediatr. Infect. Dis. J. 2006, 25, 680–686. [Google Scholar] [CrossRef] [PubMed]
- Bulkow, L.R.; Singleton, R.J.; DeByle, C.; Miernyk, K.; Redding, G.; Hummel, K.B.; Chikoyak, L.; Hennessy, T.W. Risk Factors for Hospitalization With Lower Respiratory Tract Infections in Children in Rural Alaska. Pediatrics 2012, 129, e1220–e1227. [Google Scholar] [CrossRef] [PubMed]
- Borchers, A.T.; Chang, C.; Gershwin, M.E.; Gershwin, L.J. Respiratory syncytial virus—A comprehensive review. Clin. Rev. Allergy Immunol. 2013, 45, 331–379. [Google Scholar] [CrossRef] [PubMed]
- Hall, C.B.; Simoes, E.A.F.; Anderson, L.J. Clinical and epidemiologic features of respiratory syncytial virus. Curr. Top. Microbiol. Immunol. 2013, 372, 39–57. [Google Scholar] [PubMed]
- Lee, H.-C.; Headley, M.B.; Loo, Y.-M.; Berlin, A.; Gale, M.; Debley, J.S.; Lukacs, N.W.; Ziegler, S.F. Thymic stromal lymphopoietin is induced by respiratory syncytial virus-infected airway epithelial cells and promotes a type 2 response to infection. J. Allergy Clin. Immunol. 2012, 130, 1187–1196. [Google Scholar] [CrossRef] [PubMed]
- Glezen, W.P.; Taber, L.H.; Frank, A.L.; Kasel, J.A. Risk of primary infection and reinfection with respiratory syncytial virus. Am. J. Dis. Child. 1986, 140, 543–546. [Google Scholar] [CrossRef] [PubMed]
- Hall, C.B.; Walsh, E.E.; Long, C.E.; Schnabel, K.C. Immunity to and frequency of reinfection with respiratory syncytial virus. J. Infect. Dis. 1991, 163, 693–698. [Google Scholar] [CrossRef] [PubMed]
- Hall, C.B.; Long, C.E.; Schnabel, K.C. Respiratory syncytial virus infections in previously healthy working adults. Clin. Infect. Dis. 2001, 33, 792–796. [Google Scholar] [CrossRef] [PubMed]
- Falsey, A.R.; Walsh, E.E. Respiratory syncytial virus infection in adults. Clin. Microbiol. Rev. 2000, 13, 371–384. [Google Scholar] [CrossRef] [PubMed]
- Jansen, A.G.S.C.; Sanders, E.A.M.; Hoes, A.W.; Van Loon, A.M.; Hak, E. Influenza- and respiratory syncytial virus-associated mortality and hospitalisations. Eur. Respir. J. 2007, 30, 1158–1166. [Google Scholar] [CrossRef] [PubMed]
- Venkatesh, M.P.; Weisman, L.E. Prevention and treatment of respiratory syncytial virus infection in infants: An update. Expert Rev. Vaccines 2006, 5, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Pullan, C.R.; Hey, E.N. Wheezing, asthma, and pulmonary dysfunction 10 years after infection with respiratory syncytial virus in infancy. Br. Med. J. (Clin. Res. Ed.) 1982, 284, 1665–1669. [Google Scholar] [CrossRef]
- Henderson, F.W.; Collier, A.M.; Clyde, W.A.; Denny, F.W. Respiratory-syncytial-virus infections, reinfections and immunity. A prospective, longitudinal study in young children. N. Engl. J. Med. 1979, 300, 530–534. [Google Scholar] [CrossRef] [PubMed]
- Sigurs, N.; Aljassim, F.; Kjellman, B.; Robinson, P.D.; Sigurbergsson, F.; Bjarnason, R.; Gustafsson, P.M. Asthma and allergy patterns over 18 years after severe RSV bronchiolitis in the first year of life. Thorax 2010, 65, 1045–1052. [Google Scholar] [CrossRef] [PubMed]
- Klein Wolterink, R.G.; Hendriks, R.W. Type 2 innate lymphocytes in allergic airway inflammation. Curr. Allergy Asthma Rep. 2013, 13, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Schauer, U.; Hoffjan, S.; Rothoeft, T.; Bartz, H.; Konig, S.; Fuchs, E.; Bittscheidt, J.; Kochling, A.; Stephan, V. Severe respiratory syncytial virus infections and reduced interferon-gamma generation in vitro. Clin. Exp. Immunol. 2004, 138, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Openshaw, P.J.; Chiu, C. Protective and dysregulated T cell immunity in RSV infection. Curr. Opin. Virol. 2013, 3, 468–474. [Google Scholar] [CrossRef] [PubMed]
- Openshaw, P.J.M.; Chiu, C.; Culley, F.J.; Johansson, C. Protective and Harmful Immunity to RSV Infection. Annu. Rev. Immunol. 2017, 35, 501–532. [Google Scholar] [CrossRef] [PubMed]
- Cully, M. Lung disease: IL-25 blockade could reduce virus-associated asthma attacks. Nat. Rev. Drug Discov. 2014, 13, 810–811. [Google Scholar] [CrossRef] [PubMed]
- De Graaff, P.M.A.; de Jong, E.C.; van Capel, T.M.; van Dijk, M.E.A.; Roholl, P.J.M.; Boes, J.; Luytjes, W.; Kimpen, J.L.L.; van Bleek, G.M. Respiratory syncytial virus infection of monocyte-derived dendritic cells decreases their capacity to activate CD4 T cells. J. Immunol. 2005, 175, 5904–5911. [Google Scholar] [CrossRef] [PubMed]
- Bangham, C.R.; Openshaw, P.J.; Ball, L.A.; King, A.M.; Wertz, G.W.; Askonas, B.A. Human and murine cytotoxic T cells specific to respiratory syncytial virus recognize the viral nucleoprotein (N), but not the major glycoprotein (G), expressed by vaccinia virus recombinants. J. Immunol. 1986, 137, 3973–3977. [Google Scholar] [PubMed]
- Cherrie, A.H.; Anderson, K.; Wertz, G.W.; Openshaw, P.J. Human cytotoxic T cells stimulated by antigen on dendritic cells recognize the N, SH, F, M, 22K, and 1b proteins of respiratory syncytial virus. J. Virol. 1992, 66, 2102–2110. [Google Scholar] [PubMed]
- Zeng, R.; Cui, Y.; Hai, Y.; Liu, Y. Pattern recognition receptors for respiratory syncytial virus infection and design of vaccines. Virus Res. 2012, 167, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Qin, L.; Hu, C.P.; Feng, J.T.; Xia, Q. Activation of lymphocytes induced by bronchial epithelial cells with prolonged RSV infection. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [PubMed]
- Spann, K.M.; Tran, K.C.; Chi, B.; Rabin, R.L.; Collins, P.L. Suppression of the induction of α, β, and λ interferons by the NS1 and NS2 proteins of human respiratory syncytial virus in human epithelial cells and macrophages. J. Virol. 2004, 78, 4363–4369. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, R.; Tsutsumi, H.; Osaki, M.; Sone, S.; Imai, S.; Chiba, S. Respiratory syncytial virus infection of neonatal monocytes stimulates synthesis of interferon regulatory factor 1 and interleukin-1β (IL-1β)-converting enzyme and secretion of IL-1β. J. Virol. 1998, 72, 837–840. [Google Scholar] [PubMed]
- Espinoza, J.A.; Bohmwald, K.; Céspedes, P.F.; Riedel, C.A.; Bueno, S.M.; Kalergis, A.M. Modulation of host adaptive immunity by hRSV proteins. Virulence 2014, 5, 740–751. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.; Choi, S.Y.; Jin, H.T.; Sung, Y.C.; Braciale, T.J. Improved effector activity and memory CD8 T cell development by IL-2 expression during experimental respiratory syncytial virus infection. J. Immunol. 2004, 172, 503–508. [Google Scholar] [CrossRef] [PubMed]
- Vallbracht, S.; Unsöld, H.; Ehl, S. Functional impairment of cytotoxic T cells in the lung airways following respiratory virus infections. Eur. J. Immunol. 2006, 36, 1434–1442. [Google Scholar] [CrossRef] [PubMed]
- Olson, M.R.; Varga, S.M. Pulmonary immunity and immunopathology: Lessons from respiratory syncytial virus. Expert Rev. Vaccines 2008, 7, 1239–1255. [Google Scholar] [CrossRef] [PubMed]
- Varga, S.M.; Wang, X.; Welsh, R.M.; Braciale, T.J. Immunopathology in RSV infection is mediated by a discrete oligoclonal subset of antigen-specific CD4 + T cells. Immunity 2001, 15, 637–646. [Google Scholar] [CrossRef]
- Medeiros, M.A.; Armôa, G.R.G.; Dellagostin, O.A.; McIntosh, D. Induction of humoral immunity in response to immunization with recombinant Mycobacterium bovis BCG expressing the S1 subunit of Bordetella pertussis toxin. Can. J. Microbiol. 2005, 51, 1015–1020. [Google Scholar] [CrossRef] [PubMed]
- Snapper, C.M.; Paul, W.E. Interferon-gamma and B cell stimulatory factor-1 reciprocally regulate Ig isotype production. Science 1987, 236, 944–947. [Google Scholar] [CrossRef] [PubMed]
- Espinoza, J.A.; Bohmwald, K.; Cespedes, P.F.; Gomez, R.S.; Riquelme, S.A.; Cortes, C.M.; Valenzuela, J.A.; Sandoval, R.A.; Pancetti, F.C.; Bueno, S.M.; et al. Impaired learning resulting from respiratory syncytial virus infection. Proc. Natl. Acad. Sci. USA 2013, 110, 9112–9117. [Google Scholar] [CrossRef] [PubMed]
- McBride, J.T. Pulmonary function changes in children after respiratory syncytial virus infection in infancy. J. Pediatr. 1999, 135, 28–32. [Google Scholar] [PubMed]
- De Almeida Nagata, D.E.; Demoor, T.; Ptaschinski, C.; Ting, H.A.; Jang, S.; Reed, M.; Mukherjee, S.; Lukacs, N.W. IL-27R-mediated regulation of IL-17 controls the development of respiratory syncytial virus-associated pathogenesis. Am. J. Pathol. 2014, 184, 1807–1818. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.; Incognito, L.; Cheng, X.; Ulbrandt, N.D.; Wu, H. Respiratory Syncytial Virus-Neutralizing Monoclonal Antibodies Motavizumab and Palivizumab Inhibit Fusion. J. Virol. 2010, 84, 8132–8140. [Google Scholar] [CrossRef] [PubMed]
- Hamelin, M.E.; Couture, C.; Sackett, M.; Kiener, P.; Suzich, J.; Ulbrandt, N.; Boivin, G. The prophylactic administration of a monoclonal antibody against human metapneumovirus attenuates viral disease and airways hyperresponsiveness in mice. Antivir. Ther. 2008, 13, 39–46. [Google Scholar] [PubMed]
- Gill, M.A.; Welliver, R.C. Motavizumab for the prevention of respiratory syncytial virus infection in infants. Expert Opin. Biol. Ther. 2009, 9, 1335–1345. [Google Scholar] [CrossRef] [PubMed]
- Weisman, L.E. Motavizumab, a second-generation humanized mAb for the prevention of respiratory syncytial virus infection in high-risk populations. Curr. Opin. Mol. Ther. 2009, 11, 208–218. [Google Scholar] [PubMed]
- Fernández, P.; Trenholme, A.; Abarca, K.; Griffin, M.P.; Hultquist, M.; Harris, B.; Losonsky, G.A. A phase 2, randomized, double-blind safety and pharmacokinetic assessment of respiratory syncytial virus (RSV) prophylaxis with motavizumab and palivizumab administered in the same season. BMC Pediatr. 2010, 10, 38. [Google Scholar] [CrossRef] [PubMed]
- Lagos, R.; DeVincenzo, J.P.; Muñoz, A.; Hultquist, M.; Suzich, J.; Connor, E.M.; Losonsky, G.A. Safety and antiviral activity of motavizumab, a respiratory syncytial virus (RSV)-specific humanized monoclonal antibody, when administered to RSV-infected children. Pediatr. Infect. Dis. J. 2009, 28, 835–837. [Google Scholar] [CrossRef] [PubMed]
- Robbie, G.J.; Criste, R.; Dall’Acqua, W.F.; Jensen, K.; Patel, N.K.; Losonsky, G.A.; Griffin, M.P. A novel investigational Fc-modified humanized monoclonal antibody, motavizumab-YTE, has an extended half-life in healthy adults. Antimicrob. Agents Chemother. 2013, 57, 6147–6153. [Google Scholar] [CrossRef] [PubMed]
- Abarca, K.; Jung, E.; Fernández, P.; Zhao, L.; Harris, B.; Connor, E.M.; Losonsky, G.A. Safety, Tolerability, Pharmacokinetics, and Immunogenicity of Motavizumab, a Humanized, Enhanced-Potency Monoclonal Antibody for the Prevention of Respiratory Syncytial Virus Infection in At-Risk Children. Pediatr. Infect. Dis. J. 2009, 28, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Ramilo, O.; Lagos, R.; Sáez-Llorens, X.; Suzich, J.; Wang, C.K.; Jensen, K.M.; Harris, B.S.; Losonsky, G.A.; Griffin, M.P. Motavizumab Treatment of Infants Hospitalized With Respiratory Syncytial Virus Infection Does Not Decrease Viral Load or Severity of Illness. Pediatr. Infect. Dis. J. 2014, 33, 703–709. [Google Scholar] [CrossRef] [PubMed]
- Knudson, C.J.; Hartwig, S.M.; Meyerholz, D.K.; Varga, S.M. RSV Vaccine-Enhanced Disease Is Orchestrated by the Combined Actions of Distinct CD4 T Cell Subsets. PLoS Pathog. 2015, 11, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Bell, J.A.; Sundberg, J.P.; Ghim, S.J.; Newsome, J.; Jenson, A.B.; Schlegel, R. A formalin-inactivated vaccine protects against mucosal papillomavirus infection: A canine model. Pathobiology 1994, 62, 194–198. [Google Scholar] [CrossRef] [PubMed]
- Prince, G.A.; Curtis, S.J.; Yim, K.C.; Porter, D.D. Vaccine-enhanced respiratory syncytial virus disease in cotton rats following immunization with Lot 100 or a newly prepared reference vaccine. J. Gen. Virol. 2001, 82, 2881–2888. [Google Scholar] [CrossRef] [PubMed]
- Loebbermann, J.; Durant, L.; Thornton, H.; Johansson, C.; Openshaw, P.J. Defective immunoregulation in RSV vaccine-augmented viral lung disease restored by selective chemoattraction of regulatory T cells. Proc. Natl. Acad. Sci. USA 2013, 110, 2987–2992. [Google Scholar] [CrossRef] [PubMed]
- Swanson, K.A.; Balabanis, K.; Xie, Y.; Aggarwal, Y.; Palomo, C.; Mas, V.; Metrick, C.; Yang, H.; Shaw, C.A.; Melero, J.A.; et al. A Monomeric Uncleaved Respiratory Syncytial Virus F Antigen Retains Prefusion-Specific Neutralizing Epitopes. J. Virol. 2014, 88, 11802–11810. [Google Scholar] [CrossRef] [PubMed]
- Herfst, S.; Schrauwen, E.J.A.; de Graaf, M.; van Amerongen, G.; van den Hoogen, B.G.; de Swart, R.L.; Osterhaus, A.D.M.E.; Fouchier, R.A.M. Immunogenicity and efficacy of two candidate human metapneumovirus vaccines in cynomolgus macaques. Vaccine 2008, 26, 4224–4230. [Google Scholar] [CrossRef] [PubMed]
- Gilman, M.S.A.; Moin, S.M.; Mas, V.; Chen, M.; Patel, N.K.; Kramer, K.; Zhu, Q.; Kabeche, S.C.; Kumar, A.; Palomo, C.; et al. Characterization of a Prefusion-Specific Antibody That Recognizes a Quaternary, Cleavage-Dependent Epitope on the RSV Fusion Glycoprotein. PLoS Pathog. 2015, 11. [Google Scholar] [CrossRef] [PubMed]
- Krarup, A.; Truan, D.; Furmanova-Hollenstein, P.; Bogaert, L.; Bouchier, P.; Bisschop, I.J.M.; Widjojoatmodjo, M.N.; Zahn, R.; Schuitemaker, H.; McLellan, J.S.; et al. A highly stable prefusion RSV F vaccine derived from structural analysis of the fusion mechanism. Nat. Commun. 2015, 6, 8143. [Google Scholar] [CrossRef] [PubMed]
- Lambert, S.L.; Aslam, S.; Stillman, E.; MacPhail, M.; Nelson, C.; Ro, B.; Sweetwood, R.; Lei, Y.M.; Woo, J.C.; Tang, R.S. A novel Respiratory Syncytial Virus (RSV) F subunit vaccine adjuvanted with GLA-SE elicits robust protective TH1-type humoral and cellular immunity in rodent models. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.N.Y.T.; Suk Hwang, H.; Kim, M.C.; Lee, Y.N.Y.T.; Cho, M.K.; Kwon, Y.M.; Seok Lee, J.; Plemper, R.K.; Kang, S.M. Recombinant influenza virus carrying the conserved domain of respiratory syncytial virus (RSV) G protein confers protection against RSV without inflammatory disease. Virology 2015, 476, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Russell, R.F.; McDonald, J.U.; Ivanova, M.; Zhong, Z.; Bukreyev, A.; Tregoning, J.S. Partial Attenuation of Respiratory Syncytial Virus with a Deletion of a Small Hydrophobic Gene Is Associated with Elevated Interleukin-1β Responses. J. Virol. 2015, 89, 8974–8981. [Google Scholar] [CrossRef] [PubMed]
- Taylor, G.; Wyld, S.; Valarcher, J.F.; Guzman, E.; Thom, M.; Widdison, S.; Buchholz, U.J. Recombinant bovine respiratory syncytial virus with deletion of the SH gene induces increased apoptosis and pro-inflammatory cytokines in vitro, and is attenuated and induces protective immunity in calves. J. Gen. Virol. 2014, 95, 1244–1254. [Google Scholar] [CrossRef] [PubMed]
- Blodörn, K.; Hägglund, S.; Fix, J.; Dubuquoy, C.; Makabi-Panzu, B.; Thom, M.; Karlsson, P.; Roque, J.-L.; Karlstam, E.; Pringle, J.; et al. Vaccine Safety and Efficacy Evaluation of a Recombinant Bovine Respiratory Syncytial Virus (BRSV) with Deletion of the SH Gene and Subunit Vaccines Based On Recombinant Human RSV Proteins: N-nanorings, P and M2-1, in Calves with Maternal Antibodies. PLoS ONE 2014, 9, e100392. [Google Scholar] [CrossRef] [PubMed]
- Yamaji, Y.; Nakayama, T. Recombinant measles viruses expressing respiratory syncytial virus proteins induced virus-specific CTL responses in cotton rats. Vaccine 2014, 32, 4529–4536. [Google Scholar] [CrossRef] [PubMed]
- Murawski, M.R.; McGinnes, L.W.; Finberg, R.W.; Kurt-Jones, E.A.; Massare, M.J.; Smith, G.; Heaton, P.M.; Fraire, A.E.; Morrison, T.G. Newcastle Disease Virus-Like Particles Containing Respiratory Syncytial Virus G Protein Induced Protection in BALB/c Mice, with No Evidence of Immunopathology. J. Virol. 2010, 84, 1110–1123. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.M.; Pottage, L.; Thomas, E.R.; Leishman, A.J.; Doig, T.N.; Xu, D.; Liew, F.Y.; Garside, P. Th1 and Th2 CD4+ T cells provide help for B cell clonal expansion and antibody synthesis in a similar manner in vivo. J. Immunol. 2000, 165, 3136–3144. [Google Scholar] [CrossRef] [PubMed]
- Cullen, L.M.; Blanco, J.C.G.; Morrison, T.G. Cotton rat immune responses to virus-like particles containing the pre-fusion form of respiratory syncytial virus fusion protein. J. Transl. Med. 2015, 13, 350. [Google Scholar] [CrossRef] [PubMed]
- McGinnes Cullen, L.; Schmidt, M.R.; Kenward, S.A.; Woodland, R.T.; Morrison, T.G. Murine Immune Responses to Virus-Like Particle-Associated Pre- and Postfusion Forms of the Respiratory Syncytial Virus F Protein. J. Virol. 2015, 89, 6835–6847. [Google Scholar] [CrossRef] [PubMed]
- Ghildyal, R.; Mills, J.; Murray, M.; Vardaxis, N.; Meanger, J. Respiratory syncytial virus matrix protein associates with nucleocapsids in infected cells. J. Gen. Virol. 2002, 83, 753–757. [Google Scholar] [CrossRef] [PubMed]
- Kurt-Jones, E.A.; Popova, L.; Kwinn, L.; Haynes, L.M.; Jones, L.P.; Tripp, R.A.; Walsh, E.E.; Freeman, M.W.; Golenbock, D.T.; Anderson, L.J.; et al. Pattern recognition receptors TLR4 and CD14 mediate response to respiratory syncytial virus. Nat. Immunol. 2000, 1, 398–401. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Y.-Y.; Fu, Y.-H.; Yan, Y.-F.; Hua, Y.; Ma, Y.; Zhang, X.-J.; Song, J.-D.; Peng, X.-L.; Huang, J.; Hong, T.; et al. A single intranasal administration of virus-like particle vaccine induces an efficient protection for mice against human respiratory syncytial virus. Antivir. Res. 2017, 144, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Eick, A.; Karron, R.; Shaw, J.; Thumar, B.; Reid, R.; Santosham, M.; O’Brien, K.L. The role of neutralizing antibodies in protection of American Indian infants against respiratory syncytial virus disease. Pediatr. Infect. Dis. J. 2008, 27, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, H.E.; Siersma, V.; Andersen, S.; Gahrn-Hansen, B.; Mordhorst, C.H.; Nørgaard-Pedersen, B.; Røder, B.; Sørensen, T.L.; Temme, R.; Vestergaard, B.F. Respiratory syncytial virus infection--risk factors for hospital admission: A case-control study. Acta Paediatr. 2003, 92, 1314–1321. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.W.; Arrobio, J.O.; Brandt, C.D.; Jeffries, B.C.; Pyles, G.; Reid, J.L.; Chanock, R.M.; Parrott, R.H. Epidemiology of respiratory syncytial virus infection in Washington, D.C. I. Importance of the virus in different respiratory tract disease syndromes and temporal distribution of infection. Am. J. Epidemiol. 1973, 98, 216–225. [Google Scholar] [CrossRef] [PubMed]
- Stensballe, L.G.; Ravn, H.; Kristensen, K.; Agerskov, K.; Meakins, T.; Aaby, P.; Simões, E.A.F. Respiratory syncytial virus neutralizing antibodies in cord blood, respiratory syncytial virus hospitalization, and recurrent wheeze. J. Allergy Clin. Immunol. 2009, 123, 398–403. [Google Scholar] [CrossRef] [PubMed]
- Shinoff, J.J.; O’Brien, K.L.; Thumar, B.; Shaw, J.B.; Reid, R.; Hua, W.; Santosham, M.; Karron, R.A. Young Infants Can Develop Protective Levels of Neutralizing Antibody after Infection with Respiratory Syncytial Virus. J. Infect. Dis. 2008, 198, 1007–1015. [Google Scholar] [CrossRef] [PubMed]
- Kaaijk, P.; Luytjes, W.; Rots, N.Y. Vaccination against RSV: Is maternal vaccination a good alternative to other approaches? Hum. Vaccines Immunother. 2013, 9, 1263–1267. [Google Scholar] [CrossRef] [PubMed]
- Leentjens, J.; Kox, M.; Stokman, R.; Gerretsen, J.; Diavatopoulos, D.A.; Van Crevel, R.; Rimmelzwaan, G.F.; Pickkers, P.; Netea, M.G. BCG vaccination enhances the immunogenicity of subsequent influenza vaccination in healthy volunteers: A randomized, placebo-controlled pilot study. J. Infect. Dis. 2015, 212, 1930–1938. [Google Scholar] [CrossRef] [PubMed]
- Rezende, C.A.F.; De Moraes, M.T.B.; Matos, D.C.D.S.; Mcintoch, D.; Armoa, G.R.G. Humoral response and genetic stability of recombinant BCG expressing hepatitis B surface antigens. J. Virol. Methods 2005, 125, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Cautivo, K.M.; Bueno, S.M.; Cortes, C.M.; Wozniak, A.; Riedel, C.A.; Kalergis, A.M. Efficient lung recruitment of respiratory syncytial virus-specific Th1 cells induced by recombinant bacillus Calmette-Guérin promotes virus clearance and protects from infection. J. Immunol. 2010, 185, 7633–7645. [Google Scholar] [CrossRef] [PubMed]
- Fucs, R.; Jesus, J.T.; Souza Junior, P.H.; Franco, L.; Vericimo, M.; Bellio, M.; Nobrega, A. Frequency of natural regulatory CD4+CD25+ T lymphocytes determines the outcome of tolerance across fully mismatched MHC barrier through linked recognition of self and allogeneic stimuli. J. Immunol. 2006, 176, 2324–2329. [Google Scholar] [CrossRef] [PubMed]
- Morse, K.; Norimine, J.; Palmer, G.H.; Sutten, E.L.; Baszler, T.V.; Brown, W.C. Association and evidence for linked recognition of type IV secretion system proteins VirB9-1, VirB9-2, and VirB10 in Anaplasma marginale. Infect. Immun. 2012, 80, 215–227. [Google Scholar] [CrossRef] [PubMed]
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gálvez, N.M.S.; Soto, J.A.; Kalergis, A.M. New Insights Contributing to the Development of Effective Vaccines and Therapies to Reduce the Pathology Caused by hRSV. Int. J. Mol. Sci. 2017, 18, 1753. https://doi.org/10.3390/ijms18081753
Gálvez NMS, Soto JA, Kalergis AM. New Insights Contributing to the Development of Effective Vaccines and Therapies to Reduce the Pathology Caused by hRSV. International Journal of Molecular Sciences. 2017; 18(8):1753. https://doi.org/10.3390/ijms18081753
Chicago/Turabian StyleGálvez, Nicolás M. S., Jorge A. Soto, and Alexis M. Kalergis. 2017. "New Insights Contributing to the Development of Effective Vaccines and Therapies to Reduce the Pathology Caused by hRSV" International Journal of Molecular Sciences 18, no. 8: 1753. https://doi.org/10.3390/ijms18081753
APA StyleGálvez, N. M. S., Soto, J. A., & Kalergis, A. M. (2017). New Insights Contributing to the Development of Effective Vaccines and Therapies to Reduce the Pathology Caused by hRSV. International Journal of Molecular Sciences, 18(8), 1753. https://doi.org/10.3390/ijms18081753