
The Role of α1-Adrenoceptor Antagonists in the Treatment of Prostate and Other Cancers

 , ,
, ,
Abstract
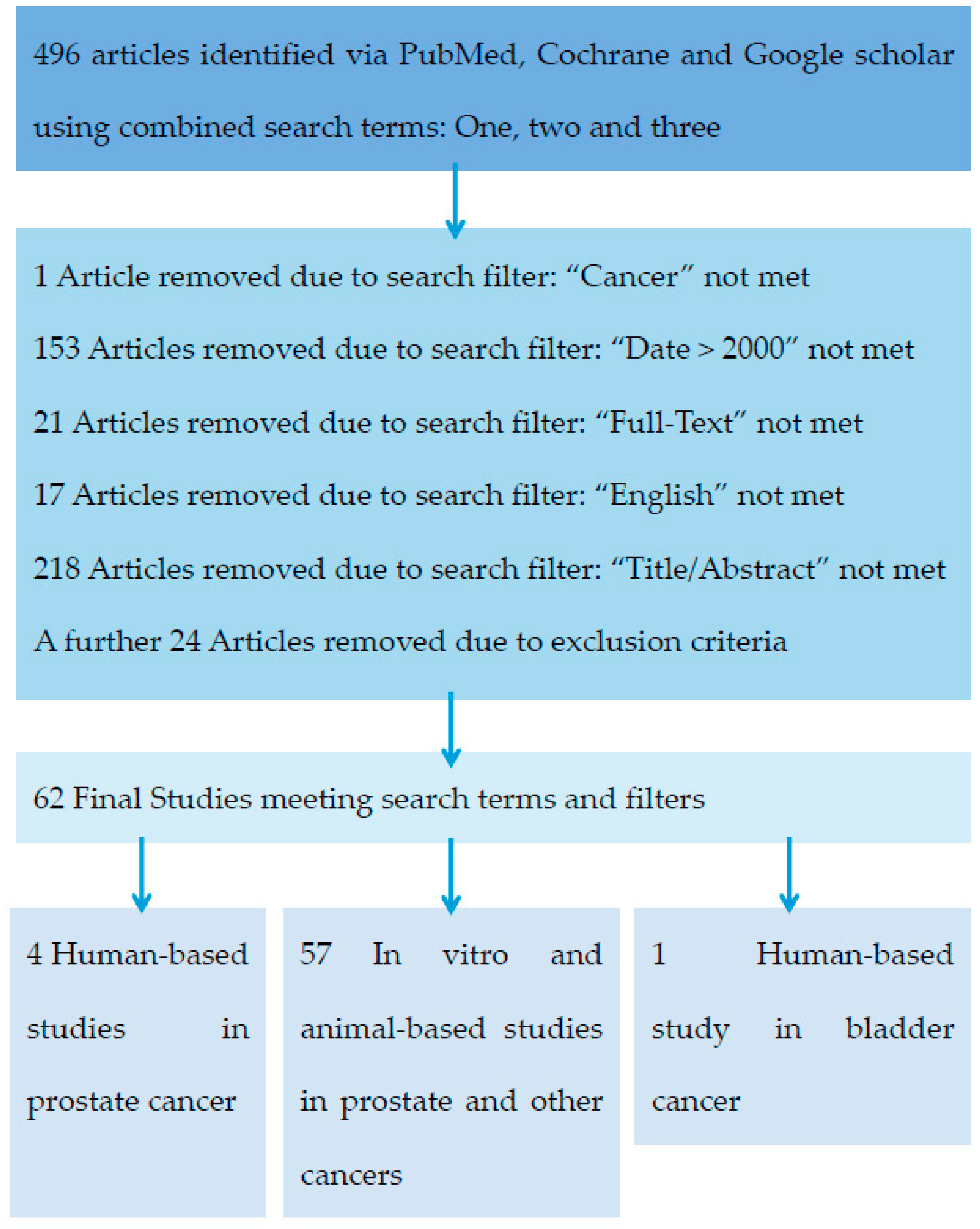
:1. Introduction
2. Results
3. Discussion
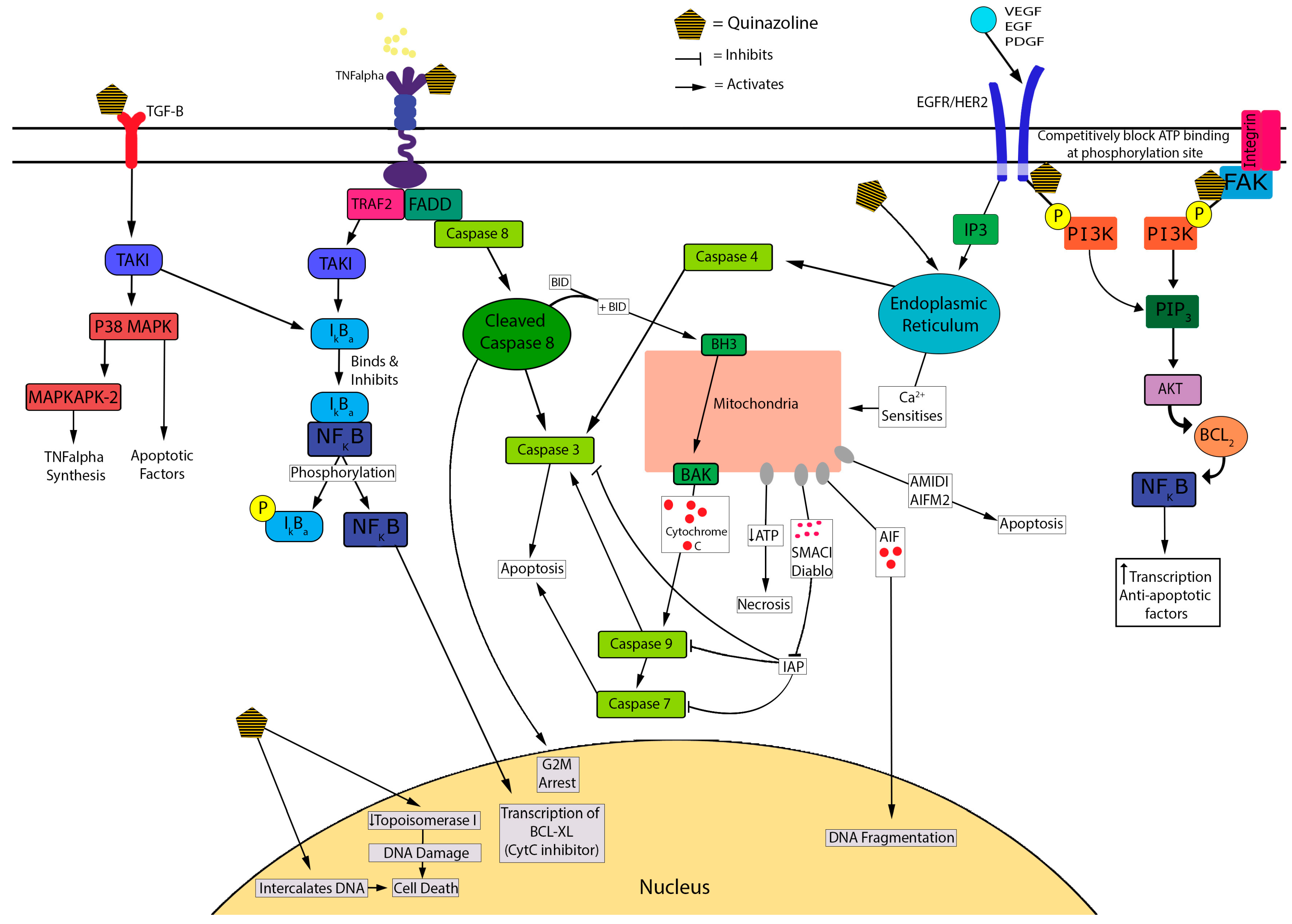
3.1. In Vitro Evidence
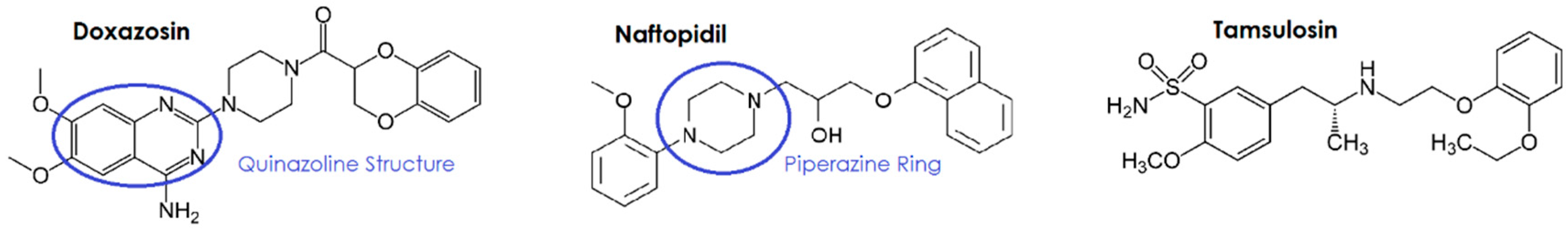
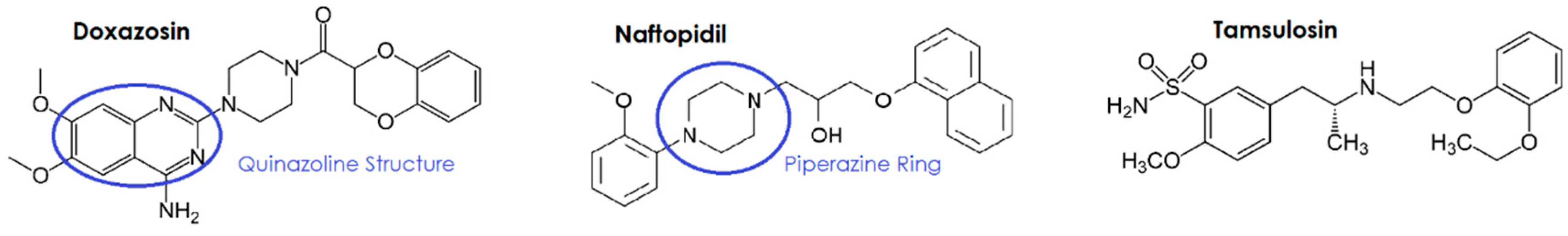
3.1.1. Quinazoline/Piperazine-Dependence
3.1.2. α1-Adrenoceptor-Independence
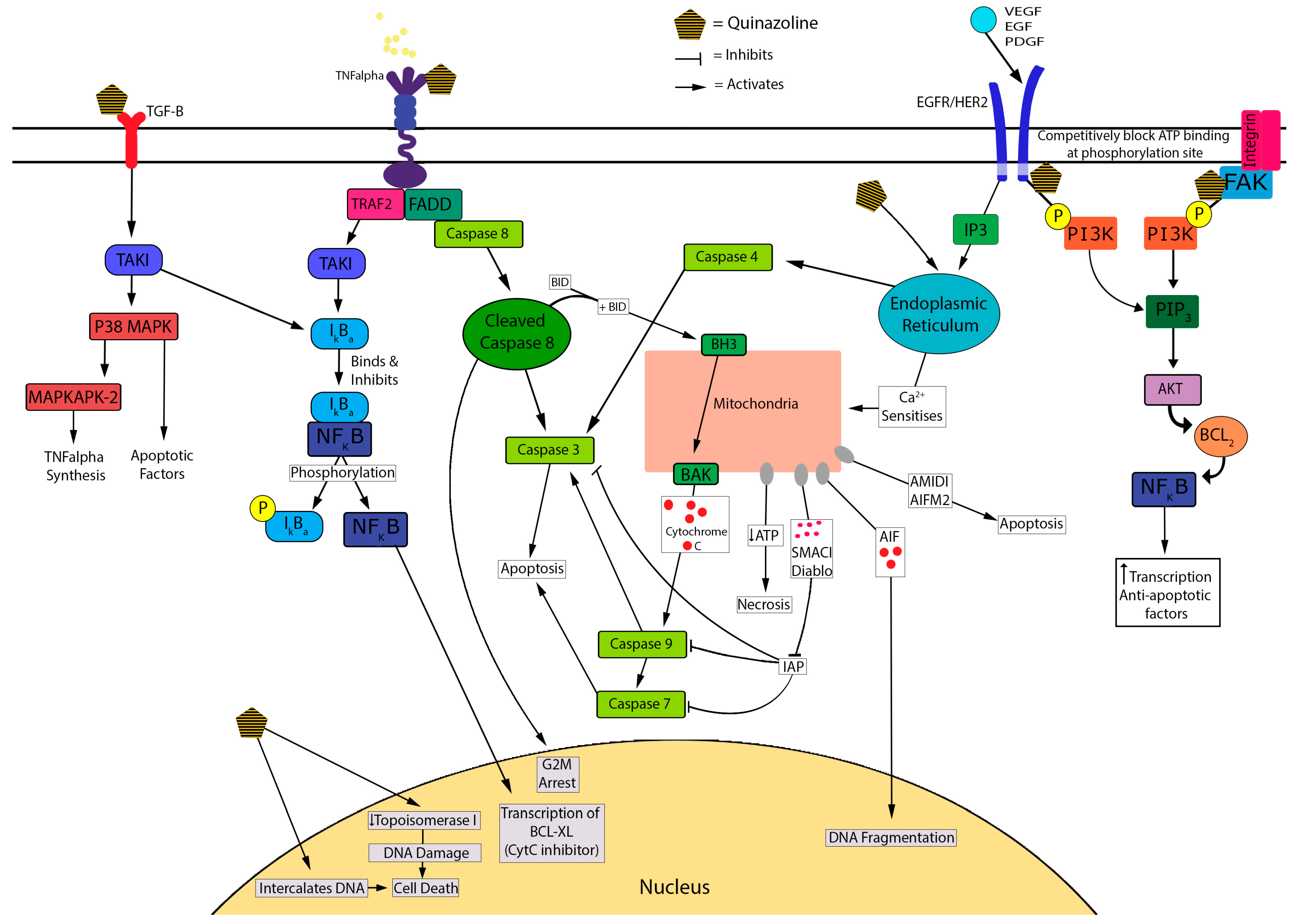
3.1.3. Cell Death Mechanisms
3.2. In Vivo Evidence
3.3. Clinical Evidence
3.4. Anticancer Effects of α-Antagonists in Other Cancers
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| Search Term 1: Agent | Search Term 2: Target Tissue | Search Term 3: Action |
|---|---|---|
| Alfuzosin α Adrenergic antagonist α Adrenoreceptor blockers α Blocker Doxazosin Naftopidil Phentolamine Prazosin Silodosin Terazosin | Adenocarcinoma Cancer Carcinoma Neoplasm Prostate cancer | Anoikis Anti-angiogenic Anti-proliferative Anticancer Antineoplastic Apoptosis Cytotoxic |
| Filters Applied in PubMed | ||
| Text—Full text Publication Date—2000–2016 Language—English Subjects—Cancer Search Fields—Title/Abstract | ||
| Exclusion terms: In Title/Abstract | ||
| AG1478, α-methyl-DL-tryptophan, α-linolenic, α-methyltryptophan, Biscoumarins, BYL719, Calcium channels, Cardiotoxic, Chitosan, Gambogic acid, Glyceollin, Hepatocarcinogens, HhAntag691, Insulin-like growth factor 1 receptor, LSC, Mast cells, PSC833, R482G isoform, Raloxifene, Stapling, Stilbenes, Toremifene, Triphosphate-binding | ||
| Combined Search Terms | ||
| (α adrenergic antagonist OR α adrenoreceptor blocker OR α blocker OR Prazosin OR Doxazosin OR Naftopidil OR Phentolamine OR Alfuzosin OR Terazosin OR Silodosin) AND (Neoplasm OR Cancer OR Prostate cancer OR Carcinoma OR Adenocarcinoma) AND (Anoikis OR Antineoplastic OR Apoptosis OR Anticancer OR Anti-angiogenic OR Anti-proliferative OR Cytotoxic) NOT (Calcium channels OR Cardiotoxic OR LSC OR Biscoumarins OR Glyceollin OR Chitosan OR toremifene OR triphosphate-binding OR α-linolenic OR α-methyl-DL-tryptophan OR HhAntag691 OR α-methyltryptophan OR raloxifene OR AG1478 OR R482G isoform OR PSC833 OR Mast cells OR Gambogic acid OR hepatocarcinogens OR insulin-like growth factor 1 receptor OR stapling OR BYL719 OR stilbenes) | ||

References
- Bray, F.; Ren, J.S.; Masuyer, E.; Ferlay, J. Global estimates of cancer prevalence for 27 sites in the adult population in 2008. Int. J. Cancer 2013, 132, 1133–1145. [Google Scholar] [CrossRef] [PubMed]
- Australian Institute of Health and Welfare. Prostate Cancer in Australia; Cancer Series no. 79. Cat. no. CAN 76; AIHW: Canberra, Australia, 2013.
- Freedland, S.J.; Humphreys, E.B.; Mangold, L.A.; Eisenberger, M.; Dorey, F.J.; Walsh, P.C.; Partin, A.W. Risk of prostate cancer-specific mortality following biochemical recurrence after radical prostatectomy. JAMA 2005, 294, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Zumsteg, Z.S.; Spratt, D.E.; Romesser, P.B.; Pei, X.; Zhang, Z.; Polkinghorn, W.; McBride, S.; Kollmeier, M.; Yamada, Y.; Zelefsky, M.J. The natural history and predictors of outcome following biochemical relapse in the dose escalation era for prostate cancer patients undergoing definitive external beam radiotherapy. Eur. Urol. 2015, 67, 1009–1016. [Google Scholar] [CrossRef] [PubMed]
- Kupelian, P.A.; Buchsbaum, J.C.; Patel, C.; Elshaikh, M.; Reddy, C.A.; Zippe, C.; Klein, E.C. Impact of biochemical failure on overall survival after radiation therapy for localized prostate cancer in the PSA era. Int. J. Radiat. Oncol. Biol. Phys. 2002, 52, 704–711. [Google Scholar] [CrossRef]
- Tombal, B.; Borre, M.; Rathenborg, P.; Werbrouck, P.; van Poppel, H.; Heidenreich, A.; Lversen, P.; Braeckman, J.; Heracek, J.; Baskin-Bey, E.; et al. Enzalutamide monotherapy in hormone-naive prostate cancer: Primary analysis of an open-label, single-arm, phase 2 study. Lancet Oncol. 2014, 15, 592–600. [Google Scholar] [CrossRef]
- Ryan, C.J.; Smith, M.R.; de Bono, J.S.; Molina, A.; Logothetis, C.J.; de Souza, P.; Fizazi, K.; Mainwaring, P.; Piulats, J.M.; Ng, S.; et al. Abiraterone in metastatic prostate cancer without previous chemotherapy. N. Engl. J. Med. 2013, 368, 138–148. [Google Scholar] [CrossRef] [PubMed]
- Mottet, N.; Bellmunt, J.; Bolla, M.; Joniau, S.; Mason, M.; Matveev, V.; Schmid, H.P.; van der Kwast, T.; Wiegel, T.; Zattoni, F.; et al. EAU guidelines on prostate cancer. Part II: Treatment of advanced, relapsing, and castration-resistant prostate cancer. Eur. Urol. 2011, 59, 572–583. [Google Scholar] [CrossRef] [PubMed]
- De Bono, J.S.; Oudard, S.; Ozguroglu, M.; Hansen, S.; Machiels, J.P.; Kocak, I.; Gravis, G.; Bodrogi, I.; Mackenzie, M.J.; Shen, L.; et al. Prednisone plus cabazitaxel or mitoxantrone for metastatic castration-resistant prostate cancer progressing after docetaxel treatment: A randomised open-label trial. Lancet 2010, 376, 1147–1154. [Google Scholar] [CrossRef]
- Schrader, A.J.; Boegemann, M.; Ohlmann, C.H.; Schnoeller, T.J.; Krabbe, L.M.; Hajili, T.; Jentzmik, F.; Stoeckle, M.; Schrader, M.; Herrmann, E.; et al. Enzalutamide in castration-resistant prostate cancer patients progressing after docetaxel and abiraterone. Eur. Urol. 2014, 65, 30–36. [Google Scholar]
- Cotecchia, S. The α1-adrenergic receptors: Diversity of signaling networks and regulation. J. Recept. Signal Transduct. Res. 2010, 30, 410–419. [Google Scholar] [CrossRef] [PubMed]
- Price, D.T.; Schwinn, D.A.; Lomasney, J.W.; Allen, L.F.; Caron, M.G.; Lefkowitz, R.J. Identification, quantification, and localization of mRNA for three distinct α1 adrenergic receptor subtypes in human prostate. J. Urol. 1993, 150 Pt 1, 546–551. [Google Scholar] [PubMed]
- White, C.W.; Xie, J.H.; Ventura, S. Age-related changes in the innervation of the prostate gland: Implications for prostate cancer initiation and progression. Organogenesis 2013, 9, 206–215. [Google Scholar] [CrossRef] [PubMed]
- Thebault, S.; Roudbaraki, M.; Sydorenko, V.; Shuba, Y.; Lemonnier, L.; Slomianny, C.; Deasilly, E.; Bonnal, J.L.; Mauroy, B.; Skryma, R.; et al. α1-Adrenergic receptors activate Ca2+-permeable cationic channels in prostate cancer epithelial cells. J. Clin. Investig. 2003, 111, 1691–1701. [Google Scholar] [CrossRef] [PubMed]
- Jensen, B.C.; Swigart, P.M.; Simpson, P.C. Ten commercial antibodies for α-1-adrenergic receptor subtypes are nonspecific. Naunyn Schmiedebergs Arch. Pharmacol. 2009, 379, 409–412. [Google Scholar] [CrossRef] [PubMed]
- Tseng-Crank, J.; Kost, T.; Goetz, A.; Hazum, S.; Roberson, K.M.; Haizlip, J.; Godinot, N.; Robertson, C.N.; Saussy, D. The α 1C-adrenoceptor in human prostate: Cloning, functional expression, and localization to specific prostatic cell types. Br. J. Pharmacol. 1995, 115, 1475–1485. [Google Scholar] [CrossRef] [PubMed]
- Thebault, S.; Flourakis, M.; Vanoverberghe, K.; Vandermoere, F.; Roudbaraki, M.; Lehennkyi, V.; Slomianny, C.; Beck, B.; Mariot, P.; Bonnal, J.L.; et al. Differential role of transient receptor potential channels in Ca2+ entry and proliferation of prostate cancer epithelial cells. Cancer Res. 2006, 66, 2038–2047. [Google Scholar] [CrossRef] [PubMed]
- Munaron, L.; Antoniotti, S.; Lovisolo, D. Intracellular calcium signals and control of cell proliferation: How many mechanisms? J. Cell. Mol. Med. 2004, 8, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Liou, S.F.; Lin, H.H.; Liang, J.C.; Chen, I.J.; Yeh, J.L. Inhibition of human prostate cancer cells proliferation by a selective α1-adrenoceptor antagonist labedipinedilol-A involves cell cycle arrest and apoptosis. Toxicology 2009, 256, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Jepsen, J.V.; Bruskewitz, R.C. Comprehensive patient evaluation for benign prostatic hyperplasia. Urology 1998, 51, 13–18. [Google Scholar] [CrossRef]
- Gillenwater, J.Y.; Conn, R.L.; Chrysant, S.G.; Roy, J.; Gaffney, M.; Ice, K.; Dias, N. Doxazosin for the treatment of benign prostatic hyperplasia in patients with mild to moderate essential hypertension: A double-blind, placebo-controlled, dose-response multicenter study. J. Urol. 1995, 154, 110–115. [Google Scholar] [CrossRef]
- Vincent, J.; Elliott, H.L.; Meredith, P.A.; Reid, J.I. Doxazosin, an α1-adrenoceptor antagonist: Pharmacokinetics and concentration-effect relationships in man. Br. J. Clin. Pharmacol. 1983, 15, 719–725. [Google Scholar] [CrossRef] [PubMed]
- Sonders, R.C. Pharmacokinetics of terazosin. Am. J. Med. 1986, 80, 20–24. [Google Scholar] [CrossRef]
- Lepor, H.; Kazzazi, A.; Djavan, B. α-Blockers for benign prostatic hyperplasia: The new era. Curr. Opin. Urol. 2012, 22, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Michel, M.C.; Schafers, R.F.; Goepel, M. α-Blockers and lower urinary tract function: More than smooth muscle relaxation? BJU Int. 2000, 86, 23–28. [Google Scholar] [CrossRef] [PubMed]
- McConnell, J.D.; Bruskewitz, R.; Walsh, P.; Andriole, G.; Lieber, M.; Holtgrewe, H.L.; Albertsen, P.; Roehrborn, C.G.; Nickel, J.C.; Wang, D.C.; et al. The effect of finasteride on the risk of acute urinary retention and the need for surgical treatment among men with benign prostatic hyperplasia. Finasteride Long-Term Efficacy and Safety Study Group. N. Engl. J. Med. 1998, 338, 557–563. [Google Scholar] [CrossRef] [PubMed]
- Lukacs, B.; Grange, J.C.; Comet, D.; Mc Carthy, C. Prospective follow-up of 3228 patients suffering from clinical benign prostatic hyperplasia (BPH) treated for 3 years wi alfuzosin in general practice. BPH Group in General Practice. Prog. Urol. J. Assoc. Fr. D’urol. Soc. Fr. Durol. 1999, 9, 271–280. [Google Scholar]
- Yokoyama, T.; Watanabe, T.; Saika, T.; Nasu, Y.; Kumon, H.; Miyaji, Y.; Nagai, A. Natural course of lower urinary tract symptoms following discontinuation of α-1-adrenergic blockers in patients with benign prostatic hyperplasia. Int. J. Urol. 2007, 14, 598–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishizaki, T.; Kanno, T.; Tsuchiya, A.; Kaku, Y.; Shimizu, T.; Tanaka, A. 1-[2-(2-Methoxyphenylamino)ethylamino]-3-(naphthalene-1-yloxy)propan-2-ol may be a promising anticancer drug. Molecules 2014, 19, 21462–21472. [Google Scholar] [CrossRef] [PubMed]
- Kyprianou, N.; Vaughan, T.B.; Michel, M.C. Apoptosis induction by doxazosin and other quinazoline α1-adrenoceptor antagonists: A new mechanism for cancer treatment? Naunyn Schmiedebergs Arch. Pharmacol. 2009, 380, 473–477. [Google Scholar] [CrossRef] [PubMed]
- Patane, S. Insights into cardio-oncology: Polypharmacology of quinazoline-based α1-adrenoceptor antagonists. World J. Cardiol. 2015, 7, 238–242. [Google Scholar] [CrossRef] [PubMed]
- Desiniotis, A.; Kyprianou, N. Advances in the design and synthesis of prazosin derivatives over the last ten years. Expert Opin. Ther. Targets 2011, 15, 1405–1418. [Google Scholar] [CrossRef] [PubMed]
- Tahmatzopoulos, A.; Rowland, R.G.; Kyprianou, N. The role of α-blockers in the management of prostate cancer. Expert Opin. Pharm. 2004, 5, 1279–1285. [Google Scholar] [CrossRef] [PubMed]
- Keledjian, K.; Borkowski, A.; Kim, G.; Isaacs, J.T.; Jacobs, S.C.; Kyprianou, N. Reduction of human prostate tumor vascularity by the α1-adrenoceptor antagonist terazosin. Prostate 2001, 48, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Harris, A.M.; Warner, B.W.; Wilson, J.M.; Becker, A.; Rowland, R.G.; Conner, W.; Lane, M.; Kimbler, K.; Durbin, E.B.; Baron, A.T.; et al. Effect of α1-adrenoceptor antagonist exposure on prostate cancer incidence: An observational cohort study. J. Urol. 2007, 178, 2176–2180. [Google Scholar] [CrossRef] [PubMed]
- Yamada, D.; Nishimatsu, H.; Kumano, S.; Hirano, Y.; Suzuki, M.; Fujimura, T.; Fukuhara, H.; Enomoto, Y.; Kume, H.; Homma, Y. Reduction of prostate cancer incidence by naftopidil, an α1-adrenoceptor antagonist and transforming growth factor-β signaling inhibitor. Int. J. Urol. 2013, 20, 1220–1227. [Google Scholar] [CrossRef] [PubMed]
- Bilbro, J.; Mart, M.; Kyprianou, N. Therapeutic value of quinazoline-based compounds in prostate cancer. AntiCancer Res. 2013, 33, 4695–4700. [Google Scholar] [PubMed]
- Kyprianou, N.; Benning, C.M. Suppression of human prostate cancer cell growth by α1-adrenoceptor antagonists doxazosin and terazosin via induction of apoptosis. Cancer Res. 2000, 60, 4550–4555. [Google Scholar] [PubMed]
- Pan, S.L.; Guh, J.H.; Huang, Y.W.; Chern, J.W.; Chou, J.Y.; Teng, C.M. Identification of apoptotic and antiangiogenic activities of terazosin in human prostate cancer and endothelial cells. J. Urol. 2003, 169, 724–729. [Google Scholar] [CrossRef]
- Walden, P.D.; Globina, Y.; Nieder, A. Induction of anoikis by doxazosin in prostate cancer cells is associated with activation of caspase-3 and a reduction of focal adhesion kinase. Urol. Res. 2004, 32, 261–265. [Google Scholar] [CrossRef] [PubMed]
- Benning, C.M.; Kyprianou, N. Quinazoline-derived α1-adrenoceptor antagonists induce prostate cancer cell apoptosis via an α1-adrenoceptor-independent action. Cancer Res. 2002, 62, 597–602. [Google Scholar] [PubMed]
- Kyprianou, N. Doxazosin and terazosin suppress prostate growth by inducing apoptosis: Clinical significance. J. Urol. 2003, 169, 1520–1525. [Google Scholar] [CrossRef] [PubMed]
- Arencibia, J.M.; del Rio, M.; Bonnin, A.; Lopes, R.; Lemoine, N.R.; Lopez-Barahona, M. Doxazosin induces apoptosis in LNCaP prostate cancer cell line through DNA binding and DNA-dependent protein kinase down-regulation. Int. J. Oncol. 2005, 27, 1617–1623. [Google Scholar] [PubMed]
- Siddiqui, E.J.; Del Rio, M.; Bonnin, A.; Lopes, R.; Lemoine, N.R.; Lopez-Barahona, M. Growth inhibitory effect of doxazosin on prostate and bladder cancer cells. Is the serotonin receptor pathway involved? Anticancer Res. 2005, 25, 4281–4286. [Google Scholar] [PubMed]
- Garrison, J.B.; Kyprianou, N. Doxazosin induces apoptosis of benign and malignant prostate cells via a death receptor-mediated pathway. Cancer Res. 2006, 66, 464–472. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.C.; Chueh, S.C.; Hsiao, C.J.; Li, T.K.; Chen, T.H.; Liao, C.H.; Lyu, P.C.; Guh, J.H. Prazosin displays anticancer activity against human prostate cancers: Targeting DNA and cell cycle. Neoplasia 2007, 9, 830–839. [Google Scholar] [CrossRef] [PubMed]
- Forbes, A.; Anoopkumar-Dukie, S.; Chess-Williams, R.; McDermott, C. Relative cytotoxic potencies and cell death mechanisms of α1-adrenoceptor antagonists in prostate cancer cell lines. Prostate 2016, 76, 757–766. [Google Scholar] [CrossRef] [PubMed]
- Fernando, M.A.; Heaney, A.P. α1-Adrenergic receptor antagonists: Novel therapy for pituitary adenomas. Mol. Endocrinol. 2005, 19, 3085–3096. [Google Scholar] [CrossRef] [PubMed]
- Youm, Y.H.; Yang, H.; Yoon, Y.D.; Kim, D.Y.; Lee, C.; Yoo, T.K. Doxazosin-induced clusterin expression and apoptosis in prostate cancer cells. Urol. Oncol. 2007, 25, 483–488. [Google Scholar] [CrossRef] [PubMed]
- Tahmatzopoulos, A.; Sheng, S.; Kyprianou, N. Maspin sensitizes prostate cancer cells to doxazosin-induced apoptosis. Oncogene 2005, 24, 5375–5383. [Google Scholar] [CrossRef] [PubMed]
- Partin, J.V.; Anglin, I.E.; Kyprianou, N. Quinazoline-based α 1-adrenoceptor antagonists induce prostate cancer cell apoptosis via TGF-β signalling and I κB α induction. Br. J. Cancer 2003, 88, 1615–1621. [Google Scholar] [CrossRef] [PubMed]
- Keledjian, K.; Kyprianou, N. Anoikis induction by quinazoline based α1-adrenoceptor antagonists in prostate cancer cells: Antagonistic effect of Bcl-2. J. Urol. 2003, 169, 1150–1156. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.H.; Guh, J.H.; Chueh, S.C.; Yu, H.J. Anti-angiogenic effects and mechanism of prazosin. Prostate 2011, 71, 976–984. [Google Scholar] [CrossRef] [PubMed]
- Shaw, Y.J.; Yang, Y.T.; Garrison, J.B.; Kyprianou, N.; Chen, C.S. Pharmacological exploitation of the α1-adrenoreceptor antagonist doxazosin to develop a novel class of antitumor agents that block intracellular protein kinase B/Akt activation. J. Med. Chem. 2004, 47, 4453–4462. [Google Scholar] [CrossRef] [PubMed]
- Garrison, J.B.; Shaw, Y.J.; Chen, C.S.; Kyprianou, N. Novel quinazoline-based compounds impair prostate tumorigenesis by targeting tumor vascularity. Cancer Res. 2007, 67, 11344–11352. [Google Scholar] [CrossRef] [PubMed]
- Cell Signalling Technology. Regulation of Apoptosis: Overview. 11. 2012. Available online: http://www.cellsignal.com/contents/science-pathway-research-apoptosis/regulation-of-apoptosis-signaling-pathway/pathways-apoptosis-regulation (accessed on 9 August 2016).
- Liu, C.M.; Lo, Y.C.; Tai, M.H.; Wu, B.N.; Wu, W.J.; Chou, Y.H.; Chai, C.Y.; Huang, C.H.; Chen, I.J. Piperazine-designed α1A/α1D-adrenoceptor blocker KMUP-1 and doxazosin provide down-regulation of androgen receptor and PSA in prostatic LNCaP cells growth and specifically in xenografts. Prostate 2009, 69, 610–623. [Google Scholar] [CrossRef] [PubMed]
- Ho, C.H.; Hsu, J.L.; Liu, S.P.; Hsu, L.C.; Chang, W.L.; Chao, C.C.; Guh, J.H. Repurposing of phentolamine as a potential anticancer agent against human castration-resistant prostate cancer: A central role on microtubule stabilization and mitochondrial apoptosis pathway. Prostate 2015, 75, 1454–1466. [Google Scholar] [CrossRef] [PubMed]
- Abal, M.; Andreu, J.M.; Barasoain, I. Taxanes: Microtubule and centrosome targets, and cell cycle dependent mechanisms of action. Curr. Cancer Drug Targets 2003, 3, 193–203. [Google Scholar] [CrossRef] [PubMed]
- Anglin, I.E.; Glassman, D.T.; Kyprianou, N. Induction of prostate apoptosis by α1-adrenoceptor antagonists: Mechanistic significance of the quinazoline component. Prostate Cancer Prostatic Dis. 2002, 5, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Keledjian, K.; Garrison, J.B.; Kyprianou, N. Doxazosin inhibits human vascular endothelial cell adhesion, migration, and invasion. J. Cell. Biochem. 2005, 94, 374–388. [Google Scholar] [CrossRef] [PubMed]
- Petty, A.; Myshkin, E.; Qin, H.; Guo, H.; Miao, H.; Tochtrop, G.P.; Hsieh, J.T.; Page, P.; Liu, L.L.; Lindner, D.J.; et al. A small molecule agonist of EphA2 receptor tyrosine kinase inhibits tumor cell migration in vitro and prostate cancer metastasis in vivo. PLoS ONE 2012, 7, e42120. [Google Scholar] [CrossRef] [PubMed]
- Justulin, L.A., Jr.; Acquaro, C.; Carvalho, R.F.; Silva, M.D.; Felisbino, S.L. Combined effect of the finasteride and doxazosin on rat ventral prostate morphology and physiology. Int. J. Androl. 2010, 33, 489–499. [Google Scholar] [CrossRef] [PubMed]
- Tahmatzopoulos, A.; Kyprianou, N. Apoptotic impact of α1-blockers on prostate cancer growth: A myth or an inviting reality? Prostate 2004, 59, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Hui, H.; Fernando, M.A.; Heaney, A.P. The α1-adrenergic receptor antagonist doxazosin inhibits EGFR and NF-κB signalling to induce breast cancer cell apoptosis. Eur. J. Cancer 2008, 44, 160–166. [Google Scholar] [CrossRef] [PubMed]
- Park, M.S.; Kim, B.R.; Kang, S.; Kim, D.Y.; Rho, S.B. The antihypertension drug doxazosin suppresses JAK/STATs phosphorylation and enhances the effects of IFN-α/γ-induced apoptosis. Genes Cancer 2014, 5, 470–479. [Google Scholar] [PubMed]
- Kawahara, T.; Aljarah, A.K.; Shareef, H.K.; Inoue, S.; Ide, H.; Patterson, J.D.; Kashiwagi, E.J.; Han, B.; Li, Y.; Zheng, Y.C.; et al. Silodosin inhibits prostate cancer cell growth via ELK1 inactivation and enhances the cytotoxic activity of gemcitabine. Prostate 2016, 76, 744–756. [Google Scholar] [CrossRef] [PubMed]
- Kawahara, T.; Ide, H.; Kashiwagi, E.; Patterson, J.D.; Inoue, S.; Shareef, H.K.; Aljarah, A.K.; Zheng, Y.C.; Baras, A.S.; Miyamto, H. Silodosin inhibits the growth of bladder cancer cells and enhances the cytotoxic activity of cisplatin via ELK1 inactivation. Am. J. Cancer Res. 2015, 5, 2959–2968. [Google Scholar] [PubMed]
- Iwamoto, Y.; Ishii, K.; Sasaki, T.; Kato, M.; Kanda, H.; Yamada, Y.; Arima, K.; Shiraishi, T.; Sugimura, Y. Oral naftopidil suppresses human renal-cell carcinoma by inducing G1 cell-cycle arrest in tumor and vascular endothelial cells. Cancer Prev. Res. 2013, 6, 1000–1006. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, S.; Schwarze, S.; Kyprianou, N. Anoikis disruption of focal adhesion-Akt signaling impairs renal cell carcinoma. Eur. Urol. 2011, 59, 734–744. [Google Scholar] [CrossRef] [PubMed]
- Takara, K.; Sakaeda, T.; Kakumoto, M.; Tanigawara, Y.; Kobayashi, H.; Okumura, K.; Noriaki, O.; Teruyoshi, Y. Effects of α-adrenoceptor antagonist doxazosin on MDR1-mediated multidrug resistance and transcellular transport. Oncol. Res. 2009, 17, 527–533. [Google Scholar] [CrossRef] [PubMed]
- Powe, D.G.; Voss, M.J.; Habashy, H.O.; Zanker, K.S.; Green, A.R.; Ellis, I.O.; Entschladen, F. α- And β-adrenergic receptor (AR) protein expression is associated with poor clinical outcome in breast cancer: An immunohistochemical study. Breast Cancer Res. Treat. 2011, 130, 457–463. [Google Scholar] [CrossRef] [PubMed]
- El Sharkawi, F.Z.; El Shemy, H.A.; Khaled, H.M. Possible anticancer activity of rosuvastatine, doxazosin, repaglinide and oxcarbazepin. Asian Pac. J. Cancer Prev. 2014, 15, 199–203. [Google Scholar] [CrossRef] [PubMed]
- Kanno, T.; Tanaka, A.; Shimizu, T.; Nakano, T.; Nishizaki, T. 1-[2-(2-Methoxyphenylamino)ethylamino]-3-(naphthalene-1-yloxy)propan-2-ol as a potential anticancer drug. Pharmacology 2013, 91, 339–345. [Google Scholar] [CrossRef] [PubMed]
- Kaku, Y.; Tsuchiya, A.; Kanno, T.; Nakao, S.; Shimizu, T.; Tanaka, A.; Nishizaki, T. The newly synthesized anticancer drug HUHS1015 is useful for treatment of human gastric cancer. Cancer Chemother. Pharmacol. 2015, 75, 527–535. [Google Scholar] [CrossRef] [PubMed]
- Kaku, Y.; Tsuchiya, A.; Shimizu, T.; Tanaka, A.; Nishizaki, T. HUHS1015 Suppresses colonic cancer growth by inducing necrosis and apoptosis in association with mitochondrial damage. Anticancer Res. 2016, 36, 39–48. [Google Scholar] [PubMed]
- Shen, S.G.; Zhang, D.; Hu, H.T.; Li, J.H.; Wang, Z.; Ma, Q.Y. Effects of α-adrenoreceptor antagonists on apoptosis and proliferation of pancreatic cancer cells in vitro. World J. Gastroenterol. 2008, 14, 2358–2363. [Google Scholar] [CrossRef] [PubMed]
- Masachika, E.; Kanno, T.; Nakano, T.; Gotoh, A.; Nishizaki, T. Naftopidil induces apoptosis in malignant mesothelioma cell lines independently of α1-adrenoceptor blocking. Anticancer Res. 2013, 33, 887–894. [Google Scholar] [PubMed]
- Fuchs, R.; Stracke, A.; Ebner, N.; Zeller, C.W.; Raninger, A.M.; Schittmayer, M.; Kueznik, T.; Absenger-Novak, M.; Birner-Gruenberger, R. The cytotoxicity of the α1-adrenoceptor antagonist prazosin is linked to an endocytotic mechanism equivalent to transport-P. Toxicology 2015, 338, 17–29. [Google Scholar] [CrossRef] [PubMed]
- Albinana, V.; Villar, Gomez de Las Heras, K.; Serrano-Heras, G.; Segura, T.; Perona-Moratalla, A.B.; Mota-Perez, M.; de Campos, J.M.; Botella, L.M. Propranolol reduces viability and induces apoptosis in hemangioblastoma cells from von Hippel-Lindau patients. Orphanet J. Rare Dis. 2015, 10, 118. [Google Scholar] [CrossRef] [PubMed]
- Staudacher, I.; Jehle, J.; Staudacher, K.; Pledl, H.W.; Lemke, D.; Schweizer, P.A.; Becker, R.; Katus, H.A.; Thomas, D. HERG K+ channel-dependent apoptosis and cell cycle arrest in human glioblastoma cells. PLoS ONE 2014, 9, e88164. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, R.; Schwach, G.; Stracke, A.; Meier-Allard, N.; Absenger, M.; Ingolic, E.; Haas, H.S.; Pfragner, R.; Sadjak, A. The anti-hypertensive drug prazosin induces apoptosis in the medullary thyroid carcinoma cell line TT. Anticancer Res. 2015, 35, 31–38. [Google Scholar] [PubMed]
- Tahmatzopoulos, A.; Lagrange, C.A.; Zeng, L.; Mitchell, B.L.; Conner, W.T.; Kyprianou, N. Effect of terazosin on tissue vascularity and apoptosis in transitional cell carcinoma of bladder. Urology 2005, 65, 1019–1023. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.S.; Cho, H.J.; Cho, J.M.; Kang, J.Y.; Yang, H.W.; Yoo, T.K. Dual silencing of Hsp27 and c-FLIP enhances doxazosin-induced apoptosis in PC-3 prostate cancer cells. Sci. World J. 2013, 2013, 174392. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.W.; Lee, J.W.; Chung, J.H.; Jo, J.K. Expression of heat shock protein 27 in prostate cancer cell lines according to the extent of malignancy and doxazosin treatment. World J. Men’s Health 2013, 31, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Cal, C.; Uslu, R.; Gunaydin, G.; Ozyurt, C.; Omay, S.B. Doxazosin: A new cytotoxic agent for prostate cancer? BJU Int. 2000, 85, 672–675. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.L.; Cheng, H.L.; Huang, L.W.; Hsieh, B.S.; Hu, Y.C.; Chih, T.T.; Shyu, H.W.; Su, S.J. Combined effects of terazosin and genistein on a metastatic, hormone-independent human prostate cancer cell line. Cancer Lett. 2009, 276, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Hori, Y.; Ishii, K.; Kanda, H; Iwamoto, Y.; Nishikawa, K.; Soga, N. Naftopidil, a selective α1-adrenoceptor antagonist, suppresses human prostate tumor growth by altering interactions between tumor cells and stroma. Cancer Prev. Res. 2011, 4, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; He, F.; Huang, M.; Liu, X.; Xiong, Y.; Huang, Y.; Zhu, L.; Yang, Y.; Xu, X.J.; Yuan, M. Novel naftopidil-related derivatives and their biological effects as α1-adrenoceptors antagonists and antiproliferative agents. Eur. J. Med. Chem. 2015, 96, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Hensley, P.J.; Desiniotis, A.; Wang, C.; Stromberg, A.; Chen, C.S.; Kyprianou, N. Novel pharmacologic targeting of tight junctions and focal adhesions in prostate cancer cells. PLoS ONE 2014, 9, e86238. [Google Scholar] [CrossRef] [PubMed]
- Kaku, Y.; Tsuchiya, A.; Kanno, T.; Nishizaki, T. HUHS1015 induces necroptosis and caspase-independent apoptosis of MKN28 human gastric cancer cells in association with AMID accumulation in the nucleus. Anti-Cancer Agents Med. Chem. 2015, 15, 242–247. [Google Scholar] [CrossRef]
| Author | Title | Drug | Results | Study Type |
|---|---|---|---|---|
| Keledjian, K. et al. [34] | Reduction of human prostate tumor vascularity by the α1-adrenoceptor antagonist terazosin | Terazosin | Increased apoptotic index in prostate carcinoma after terazosin treatment. Reduction in prostate tumour vascularity in terazosin-treated BPH patients. Patients were treated for 6–11 months | Retrospective Cohort study |
| Harris, A. et al. [35] | Effect of α1-adrenoceptor antagonist exposure on prostate cancer incidence: an observational cohort study | Doxazosin & Terazosin | 4070 men were treated with α-antagonists for Benign prostatic hyperplasia or hypertension or HTN. The incidence of PCa among treated men vs. untreated men was 1.65% and 2.41% respectively. Data showed 7.6 fewer cases developed per 1000 exposed men | Observational Cohort study |
| Yamada, D. et al. [36] | Reduction of prostate cancer incidence by naftopidil, an α1-adrenoceptor antagonist and transforming growth factor-β signalling inhibitor | Naftopidil & Tamsulosin | PCa incidence was significantly lower in men treated with naftopidil for ≥3 months compared to men treated with tamsulosin. (p = 0.035) | Retrospective Cohort study |
| Bilbro, J. et al. [37] | Therapeutic value of quinazoline-based compounds in prostate cancer | Doxazosin, terazosin and other quinazolines | Patients treated with α1-antagonists: doxazosin and terazosin, at the Markey Cancer centre had reduced risk of developing PCa | Retrospective Cohort study |
| Ref. | Author | Title | Study Type | Cancer Type | Drugs | Findings (Original Studies) |
|---|---|---|---|---|---|---|
| [29] | Nishizaki, T. et al. | 1-[2-(2-Methoxyphenylamino) ethylamino]-3-(naphthalene-1-yloxy) propan-2-ol may be a promising anticancer drug | Review | NA | NA | |
| [30] | Kyprianou, N. et al. | Apoptosis induction by doxazosin and other quinazoline α1-adrenoceptor antagonists: a new mechanism for cancer treatment? | Review | NA | NA | |
| [31] | Patane, S. et al. | Insights into cardio-oncology: Polypharmacology of quinazoline-based α1-adrenoceptor antagonists | Review | NA | NA | |
| [32] | Desiniotis, A. et al. | Advances in the design and synthesis of prazosin derivatives over the last ten years | Review | NA | NA | |
| [33] | Tahmatzopoulos, A. et al. | The role of α-blockers in the management of prostate cancer | Review | NA | NA | |
| [34] | Kyprianou, N. et al. | Suppression of human prostate cancer cell growth by α1-adrenoceptor antagonists Doxazosin and terazosin via induction of apoptosis | In vitro, in vivo (mice) | Prostate Cancer (PC-3 and DU145) | Doxazosin, terazosin, tamsulosin, phenoxy-benzamine | Doxazosin and terazosin induced apoptosis in prostate epithelial and smooth muscle cells in patients with BPH, without affecting rate of cell proliferation in PCa cells. This effect could not be prevented by irreversible inhibition of α1-adrenoceptors (phenoxybenzamine), indicating an in vitro toxicitity occurs via an α-receptor independent mechanism. Doxazosin administration (at tolerated pharmacologically relevant doses) in SCID mice resulted in a significant inhibition of PC-3 tumour growth, presumably via induction of apoptosis. |
| [35] | Pan, S. et al. | Identification of apoptotic and antiangiogenic activities of terazosin in human prostate cancer and endothelial cells | In vitro, in vivo (mice) | PCa PC-3 & endothelial HUVEC cells | Terazosin | It was found that terazosin induced apoptosis in PC-3 and human benign prostatic cells (IC50 > 100 µM), and possessed potent anti-angiogenic effect in endothelial cells compared to PCa cells. Terazosin (IC50 of 7.9 µM) significantly inhibited VEGF-induced angiogenesis and endothelial tube formation in nude mice, demonstrating that terazosin had a more potent anti-angiogenic than cytotoxic effects. Terazosin also effectively inhibited vascular endothelial growth factor induced proliferation and tube formation in cultured human umbilical vein endothelial cells (IC50 9.9 and 6.8 µM, respectively). Doxazosin, but not tamsulosin, mimicked these effects and the anti-tumour effects of these drugs were determined to occur independent of α1-adrenoceptor antagonizing activity. |
| [36] | Walden, P. et al. | Induction of anoikis by Doxazosin in prostate cancer cells is associated with activation of caspase-3 and a reduction of focal adhesion kinase | In vitro | PCa (PC-3 and LNCaP) | Doxazosin | Doxazosin induced changes in morphology consistent with anoikis in both benign and cancerous prostatic cells (rounding up of cells, DNA-degradation in the nucleus, cell shrinkage, the appearance of vacuoles, and cell detachment from the tissue culture plate) and increased caspase-3 activity. The increase of caspase-3 activity by doxazosin promotes anoikis and, subsequently, apoptosis of cancer cells. Treatment of PC-3 cells with doxazosin significantly reduced the protein levels of anti-anoikis kinase, FAK, but did not significantly affect the levels of ILK. Norepinephrine had no effect on doxazosin-induced cell morphology or caspase-3 activity, indicating that the apoptotic/anoikis effects of doxazosin result from mechanism that is a1-adrenoceptor independent. |
| [37] | Benning, C. et al. | Quinazoline-derived α1-adrenoceptor antagonists induce prostate cancer cell apoptosis via an α1-adrenoceptor-independent action | In vitro | Prostate cancer cells | Doxazosin, terazosin | Transfection-mediated overexpression of α1-adrenoreceptors in human prostate cancer cells, DU-145 (AR-independent, and reportedly lack of adrenoceptors), did not alter the ability of prostate cancer cells to undergo apoptosis in response to quinazolines. These findings indicate that apoptotic activity of quinazoline-based α1 adrenoceptor antagonists (doxazosin and terazosin) in prostate cancer cells is independent of α1-adrenoceptor antagonism. |
| [38] | Kyprianou, N. | Doxazosin and terazosin suppress prostate growth by inducing apoptosis: clinical significance | Review, in vitro, in vivo (mice) | PC-3, DU-145 and SMC-1 | Doxazosin, terazosin | Doxazosin and terazosin significantly reduced the viability of PC-3 and LNCaP cells by inducing caspase-3 mediated apoptosis in a dose dependent manner, however only doxazosin induced significant death of PCa cells. Doxazosin (and not terazosin) significantly affect the rate of proliferation of PCa cells. Irreversible inhibition with phenoxybenzamine did not abolish the apoptotic effect of doxazosin or terazosin against PCa or SMC cells, indicating the cytotoxic effects occurred via an α1-independent mechanism. Oral treatment with doxazosin resulted in significant decrease in tumour volume of PCa xenografts compared to controls, presumably via induction of apoptosis. |
| [39] | Arencibia, J. et al. | Doxazosin induces apoptosis in LNCaP prostate cancer cell line through DNA binding and DNA-dependent protein kinase down-regulation | In vitro | LNCaP | Doxazosin | Doxazosin induced dose-dependent LNCaP cytotoxicity and apoptosis, which could not be prevented by phenoxybenzamine, indicating an α1-adrenoceptor independent cytotoxicity. Microarray analysis following doxazosin treatment (8–24 h, 20 µM) identified 70–92 deregulated genes, including several involved in cell-cycle control and drug response, and a few related to other cellular processes such as apoptosis or angiogenesis. An inverse correlation was observed with doxazosin concentration and topoisomers, suggesting that topoisomerase I is inhibited by the binding of doxazosin to DNA. Thus, doxazosin could cause DNA damage, resulting in apoptotic cell death. |
| [40] | Siddiqui, E. et al. | Growth inhibitory effect of Doxazosin on prostate and bladder cancer cells. Is the serotonin receptor pathway involved? | In vitro | PCa PC-3, bladder cancer HT1376 | Doxazosin | Doxazosin was found to significantly reduce PCa PC-3 and bladder cancer HT1376 cell growth, which was partially prevented through pre-treatment with 5HT or 5-HT1B. These findings may be related to the structural similarity between subtype 1 serotonin and adrenergic receptors, and the authors suggests that doxazosin displaces 5-HT from 5-HT receptors. |
| [41] | Garrison, J. et al. | Doxazosin induces apoptosis of benign and malignant prostate cells via a death receptor-mediated pathway | In vitro | PC-3 and BPH1 | Doxazosin | Doxazosin (24 h) causes a dose dependent loss of cell viability and induces apoptosis in PC-3 and BPH1 cells 24 h after treatment. After a short doxazosin treatment (6 h), several genes that play a critical role in apoptosis were upregulated in PC-3 cells. In particular, doxazosin was found to upregulate Bax mRNA transcription and induced caspase-8 mediated apoptosis. |
| [42] | Lin, S. et al. | Prazosin displays anticancer activity against human prostate cancers: targeting DNA and cell cycle | In vitro, in vivo (mice) | Prostate Cancer | Prazosin | Prazosin exhibited anti-proliferative activity superior to that of other α-blockers. It induced G2 checkpoint arrest and subsequent apoptosis. In PC-3 cells, prazosin increase in DNA strand breakage leading to Cdk1 inactivation and subsequent cell cycle arrest. In mice, prazosin significantly reduced tumour mass in PC-3 derived cancer xenografts. |
| [43] | Forbes, A. et al. | Relative cytotoxic potencies and cell death mechanisms of α-adrenoceptor antagonists in prostate cancer cell lines | In vitro | PCa PC-3, LNCaP | Prazosin Doxazosin, terazosin, silodosin, alfuzosin, tamsulosin | The relative potency order was prazosin = doxazosin > terazosin = silodosin = alfuzosin> tamsulosin on both cell types, but LNCaP cells were significantly more sensitive to these effects that PC-3 cells. Prazosin and doxazosin increased levels of apoptosis and autophagy in both cell lines. However, autophagy was found to play a paradoxical role by contributing to survival of LNCaP and cytotoxicity of PC-3 cells. Treatment with prazosin (30 µM) altered the expression of several cell stress-related proteins: elevating phospho-p38α and reducing S6 kinase in both cell lines. The expression of some proteins were differentially affected in PC-3 and LNCaP cell; Akt and p27 increasing and HIF-1α decreasing in LNCaP cells but not PC-3, while ADAMTS1 was increased in PC-3 cells only. Phosphorylation of EphA2 was also reported to play a role in doxazosin, but not prazosin, induced PC-3 cytotoxicity. |
| [44] | Fernando, M. et al. | α1-Adrenergic receptor antagonists: novel therapy for pituitary adenomas | In vitro, in vivo (mice) | Pituitary tumour | Doxazosin | Treatment with Doxazosin results in reduced phosphorylation and down-regulation of NF-κB. Decreased phosphorylated retinoblastoma and PCNA expression, which resulted in cell cycle arrest at G0-G1. Doxazosin treatment also increased cleaved caspase 3. In mice, the tumour mass was lower in the doxazosin treated group. In contrast to current literature, this study suggested that the cytotoxic activity of quinazoline-antagonists was greater in cells that express α1a and 1b. In addition to apoptosis, doxazosin treatment appeared to reduce the circulating ACTH level and therefore may be useful for symptomatic relief. |
| [45] | Youm, Y. et al. | Doxazosin-induced clusterin expression and apoptosis in prostate cancer cells | In vitro | PCa PC-3 | Doxazosin | Doxazosin-induced DNA fragmentation after 24 h treatment, and was statistically significant after 48 h treatment of PC-3 cells. Clusterin expression in PC-3 cells was 3-fold higher in doxazosin treated cells (9 h) compared to untreated controls, and was maintained over 48 h. These findings were found to be consistent with doxazosin-induced apoptosis. Immunocytochemistry analysis (after 9 and 12 h treatment) demonstrated the presence of clusterin in 7% and 18% of total cells respectively. At 24 h treatment, clusterin protein was mainly observed in the cytoplasm and rarely in the nuclei of healthy cells. |
| [46] | Tahmatzopoulos, A. et al. | Maspin sensitizes prostate cancer cells to Doxazosin-induced apoptosis | In vitro | PCa DU-145 | Doxazosin | Maspin (tumour suppressor protein) was shown to increase sensitivity of PCa DU-145 cells to doxazosin, by affect the migration and attachment of malignant prostate cells to the ECM. Also caused mammary MDA-MB-435 cells to undergo apoptosis via increased Bax and caspase-3 activation. |
| [47] | Partin, J. et al. | Quinazoline-based α1-adrenoceptor antagonists induce prostate cancer cell apoptosis via TGF-β signalling and I κB α induction | In vitro | PCa PC-3 | Doxazosin, tamsulosin | Doxazosin, but not tamsulosin, was found to induce PC-3 apoptosis by enhancing TGF-β1 signalling, and subsequently, downstream 1κBα. |
| [48] | Keledjian, K. et al. | Anoikis induction by quinazoline based α1-adrenoceptor antagonists in prostate cancer cells: antagonistic effect of bcl-2 | In vitro | PCa PC-3 | Doxazosin, terazosin, tamsulosin (at therapeutic doses) | Treatment of PC-3 cells with doxazosin or terazosin, but not tamsulosin, resulted in significant down regulation of VEGF. Doxazosin also promoted anoikis. However, these effect was reduced in PC-3s that over-expressed Bcl-2 (an anoikis inhibiting factor). In these experimental conditions, these drugs did not have any effect on HIF1-α expression. |
| [49] | Liao, C. et al. | Anti-angiogenic effects and mechanism of prazosin | In vitro | PCa and HUVEC | Prazosin | Prazosin induced apoptosis in PCa and normal HUVEC cells via different mechanisms, suggesting that prazosin-mediated anti-angiogenic activity and differential modulation of apoptotic pathways are cell-type specific. |
| [50] | Kim, S. et al. | Dual silencing of Hsp27 and c-FLIP enhances doxazosin-induced apoptosis in PC-3 prostate cancer cells | In vitro | PCa PC-3 | Doxazosin | Apoptotic indices increased in a dose-dependent manner when doxazosin was added. In basal conditions (+Hsp27/+c-FLIP), doxazosin (25 µM) induced apoptosis in 52% of cells. In −Hsp27/+c-FLIP cells, apoptotic activity increased to 68% of PC-3 cells. In the opposite case (+Hsp27/−c-FLIP) the apoptotic index was 78%. Even greater number of apoptotic cells were observed (92%) when both Hsp27 and c-Flip were silence. These findings indicate that Hsp27 and c-FLIP play a protective role against doxazosin induced cytotoxicity of PC-3 cells. |
| [51] | Lee, S. et al. | Expression of heat shock protein 27 in prostate cancer cell lines according to the extent of malignancy and doxazosin treatment | In vitro | PCa LNCaP, PC-3 | Doxazosin | RT-PCR studies identified Hsp27 expression to be related to PCa malignancy potential in vitro (e.g., Hsp27 > in PC-3 than LNCaP cells), and was dose-dependently enhanced in some cell lines following doxazosin treatment. Apoptotic cell death triggered by HSP27 siRNA is greater in the androgen receptor-negative cell line PC-3 than in the androgen receptor-positive cell line LNCaP. |
| [52] | Cal, C. et al. | Doxazosin: a new cytotoxic agent for prostate cancer? | In vitro | PCa DU145, PC-3 | Doxazosin adriamycin, etoposide, paclitaxel. | DU-145 and PC3 were sensitive to doxazosin-mediated cytotoxicity, which occurred in a dose- and time-dependent fashion. The combination of doxazosin and adriamycin or etoposide resulted in significant dose-dependent cytotoxic synergism. In contrast, the combination of doxazosin and paclitaxel resulted in antagonistic activity, which was enhanced with increasing concentrations of the drugs. |
| [53] | Chang, K. et al. | Combined effects of terazosin and genistein on a metastatic, hormone-independent human prostate cancer cell line | In vitro | PCa DU-145 | Terazosin, genistein | Terazosin or genistein alone inhibited cell growth in a dose-dependent manner genistein (5 µ/mL) being more effective than terazosin (1 µ/mL—nontoxic dose). Combination treatment significantly increased apoptosis in cells compared to genistein alone. The synergistic effects of these drugs had a greater inhibitory effect the pro-survival Bcl-XL protein, compared to either drug along. Genistein and the combination also were reported to have an effect on angiogenesis-related proteins, causing a significant decrease in VEGF165 mRNA and VEGF121 mRNA levels. |
| [54] | Harris, A. et al. | Effect of α1-adrenoceptor antagonist exposure on prostate cancer incidence: an observational cohort study | Observational cohort | PCa | Doxazosin and Terazosin | Incidence of PCa in men exposed to quinazoline-based α-blockers (for BPH or hypertension) was 1.65% whereas in unexposed men, incidence was 2.41%. This indicates men who were exposed to quinazolines were 1.46 times lower relative risk of developing PCa, compared to unexposed men. However, there was no association between quinazoline exposure and overall patient survival. |
| [55] | Liu, C. et al. | Piperazine-designed α1A/α1D-adrenoceptor blocker KMUP-1 and Doxazosin provide down-regulation of androgen receptor and PSA in prostatic LNCaP cells growth and specifically in xenografts | In vitro, in vivo (mice) | PCa: LNCaP, PC-3 and DU-145 | Doxazosin and KMUP-1 | KMUP-1 and Doxazosin both inhibit LNCaP cell growth and downregulate expression of AR and PSA. KMUP-1 is a Xanthine derivative PDE inhibitor with α-blocking features. It also has a piperazine moiety very similar to that seen in Doxazosin, naftopidil which is reported to lead to its activity. KMUP-1 significantly inhibited LNCaP cell growth and induced apoptosis in time and dose-dependent manner. KMUP-1 and doxazosin further inhibited the expression of AR and PSA. Treatment of LNCaP cells with KMUP-1 resulted in cell cycle arrest and apoptotic activities, increasing p21 and p27 and decreasing expressions of cyclin D1, cyclin E, cyclin dependent kinase (CDK) 4, CDK2 and CDK6. Moreover, KMUP-1 activated p53, cleaved poly (ADP-ribose) polymerase and caspase-3, but reduced the expression of Bcl-2. Regular administration of KMUP-1 suppressed the LNCaP xenograft tumour growth in nude mice. These evidences indicate that KMUP-1 and doxazosin inhibit LNCaP cell growth and downregulate expression of AR and PSA. KMUP-1 might be used as a chemoprevention agent for preventing the development of prostate cancer without cardiovascular adverse effect of doxazosin. |
| [56] | Ho, C. et al. | Repurposing of phentolamine as a potential anticancer agent against human castration-resistant prostate cancer: A central role on microtubule stabilization and mitochondrial apoptosis pathway | In vitro | PCa DU145, PC-3 | Phentolamine, paclitaxel | Phentolamine induced anti-proliferative effects in PC-3 and Du-145, two CRPC cell lines and p-glycoprotein overexpressing cells. This effect was not significantly reduced in paclitaxel resistant cells. Phentolamine induced mitotic arrest of the cell cycle and formation of hyperdiploid cells, followed by an increase of apoptosis. Mitotic arrest was confirmed by cyclin B1 up regulation, CDK1 activation and a dramatic increase of mitotic protein phosphorylation. In vitro cellular identification demonstrated that phentolamine, similar to paclitaxel, induced tubulin polymerization and formation of multiple nuclei. The Data suggests that phentolamine is a potential anti-cancer agent. It induces microtubule assembly, leading to mitotic arrest of the cell cycle, which ‘in turn’ induces subsequent mitochondrial damage, and activation of related apoptotic signalling pathways in CRPC cells. Furthermore, the combination between phentolamine and paclitaxel induces a synergistic apoptotic cell death. Phentolamine has a simple chemical structure and is not P-gp substrate. Optimization of phentolamine structure may also be a potential approach for further development. |
| [57] | Anglin, I. et al. | Induction of prostate apoptosis by α1-adrenoceptor antagonists: mechanistic significance of the quinazoline component | Review | |||
| [58] | Keledjian, K. et al. | Doxazosin inhibits human vascular endothelial cell adhesion, migration, and invasion | In vitro | HUVEC, endothelial cells | Doxazosin | Doxazosin results in a dose-dependent loss of cell viability after 24 h of treatment. At concentrations as low as 1 mM, 10% loss of cell viability is observed and at 15 mM there is more than 30% cell death. There is also significant increase in the number of apoptotic cells within 24 h of exposure to doxazosin and a further increase after 48 h. Increased protein expression of pro-caspase-3 was observed after 6 and 12 h of doxazosin treatment. Doxazosin markedly suppresses VEGF—mediated endothelial cell adhesion to fibronectin. HUVEC cells were wounded and 24 h post-wounding, doxazosin treatment (15 mM) resulted in a dramatic decrease in HUVEC cell migration in the absence or presence of exogenous VEGF compared to control. Thus doxazosin can cause suppression of VEGF-mediated cell migration. FGF-2, a potent angiogenic factor, results in significant stimulation of HUVEC angiogenic response that was suppressed by doxazosin treatment. TGF-b had no significant impact on HUVEC-tube formation. Doxazosin treatment for 24 h resulted in a significant downregulation of VEGF mRNA. |
| [59] | Petty et al. | A small moleculre agonist of EphA2 receptor tyrosine kinase inhibits tumor cell migration in vitro and prostate cancer in vivo | In vitro, in vivo (mice) | PC-3 | Doxazosin | Doxazosin induced cell rounding and detachment via agonistic actions on EphA2 in vito. Animal studies found that doxazosin reduced number of tumour metastasis and increased survival in PC-3 xenograft nude mice |
| [60] | Justulin, L. et al. | Combined effect of the finasteride and doxazosin on rat ventral prostate morphology and physiology | In vivo | PCa | Doxazosin and finasteride | Wistar rats were treated with finasteride and doxazosin and the ventral prostate was excised at day 3 and day 30. The combination induced a transient increase in testosterone plasma concentration and a permanent reduction in DHT. The ventral prostate and epithelial cell proliferation were reduced and the collagen fibre volume fraction and apoptosis of the epithelial cell were increased. Transcription of MMP-2, TIMPs-1 and -2 mRNA was decreased after 30 days of treatment. |
| [61] | Keledjian, K. et al. | Reduction of human prostate tumor vascularity by the α1-adrenoceptor antagonist terazosin | In vitro, retro-spective | PCa | Terazosin | A significant induction of apoptosis was observed among the cancerous prostatic epithelial cells in the terazosin-treated, as compared to the untreated prostate cancer specimens, while there was no significant change in the proliferative index of the same tumour cell populations after treatment. Furthermore, terazosin resulted in a significant decrease in prostate tissue MVD compared with the untreated group, which correlated with the increased apoptotic index of the cancerous areas. Tissue PSA expression in the prostatic tumour was also markedly reduced after terazosin treatment, while no significant changes in VEGF expression were detected. These findings provide the first evidence that terazosin; a quinazoline-based α-blocker decreases prostate tumour vascularity. Our study has significant clinical implications in identifying selected α-adrenoceptor antagonists as potential anti-tumour agents with apoptotic and anti-angiogenic effects in the human prostate that can be exploited for the treatment of advanced prostate cancer. |
| [62] | Yamada, D. et al. | Reduction of prostate cancer incidence by naftopidil, an α1-adrenoceptor antagonist and transforming growth factor-β signaling inhibitor | In vitro, retro-spective | PCa | Naftopidil and tamsulosin | Prostate cancer incidence was significantly lower in men who received naftopidil for 3 months or longer compared with tamsulosin (p = 0.035). Immunohistochemically positivity for Bcl2, a marker for resistance to apoptosis, was less frequently detected in prostate cancer cells of men who received naftopidil compared with tamsulosin. Naftopidil induced apoptosis and blocked Smad2 phosphorylation activated by transforming growth factor-B in cell lines. |
| [63] | Tahmat-zopoulos, A. | Apoptotic impact of α1-blockers on prostate cancer growth: a myth or an inviting reality? | Review, in vitro, retrospective | PCa | Terazosin, doxazosin and tamsulosin | Description of α-antagonist induced anoikis and angiogenesis. Discusses retrospective study of patients using terazosin, and marked increase in tumour vascularity on autopsy. |
| [64] | Bilbro, J. et al. | Therapeutic value of quinazoline-based compounds in prostate cancer | Review | PCa | NA | NA |
| [65] | Hui, F. et al. | The α1-adrenergic receptor antagonist Doxazosin inhibits EGFR and NF-κB signalling to induce breast cancer cell apoptosis | In vitro | Breast cancer | Doxazosin | Doxazosin reduces phosphorylation of EGFR and decreases pERK1/2 levels, NF-κB, AP-1, SRE, E2F and CRE-mediated transcriptional activity. Doxazosin also decreased phosphorylated retinoblastoma (pRb) protein expression, providing a potential mechanism for the doxazosin-mediated G0/G1 cell cycle arrest. Quinazoline ring structure is similar to the EGFR tyrosine kinase inhibitors. Doxazosin appears to be safe in normal cells due to the main target being EGFR and NF-κB signalling which has greater activation in cancer cells. |
| [66] | Park, M. et al. | The antihypertension drug Doxazosin suppresses JAK/STATs phosphorylation and enhances the effects of IFN-α/γ-induced apoptosis | In vitro, In vivo (mice) | Ovarian cancer | Doxazosin | Doxazosin significantly suppressed tumour growth in an ovarian cancer cell xenograft mouse model (50%–65% reduction in tumour size), inducing apoptotic cell death by up-regulating the expression of p53. There was no additional liver toxicity or loss of body weight. In vitro identified JAK/STAT signaling as potential mediators underlying the anti-tumour effect of doxazosin. |
| [67] | Kawahara, T. et al. | Silodosin inhibits prostate cancer cell growth via ELK1 inactivation and enhances the cytotoxic activity of gemcitabine | In vitro | Prostate Cancer | Silodosin and gemcitabine | Silodosin treatment reduced the expression/activity of ELK1 in these cells as well as viability of AR-positive cells, but not the viability of AR-negative or ELK1 negative cells. Interestingly silodosin significantly increased the sensitivity to gemcitabine, but not cisplatin or docetaxel. ELK1 is likely activated in prostate cancer cells and promote tumour progression. Furthermore, silodosin that inactivates ELK1 in prostate cancer cells not only inhibits their growth but also enhances the cytotoxic activity of gemcitabine. Thus, ELK1 inhibition has the potential of being a therapeutic approach or prostate cancer. |
| [68] | Kawahara, T. et al. | Silodosin inhibits the growth of bladder cancer cells and enhances the cytotoxic activity of cisplatin via ELK1 inactivation | In vitro | Bladder Cancer (ELK-1 positive urothelial carcinoma) | Silodosin + cisplatin | Involvement of ELK1 in bladder cancer progression via modulation cell proliferation/apoptosis, migration and invasion. In bladder and prostate cancers, ELK1 was shown to induce the proliferation of cells only with an activated androgen receptor). Silodosin was found to not only inhibit cell viability and migration, but also enhance the cytotoxic activity of cisplatin in bladder cancer lines via inactivating ELK1. The results suggest that combined treatment with silodosin is useful for overcoming chemoresistance in patients with ELK-1 positive urothelial carcinoma receiving cisplatin. |
| [69] | Iwamoto, Y. et al. | Oral naftopidil suppresses human renal-cell carcinoma by inducing G(1) cell-cycle arrest in tumor and vascular endothelial cells | In vitro, in vivo (mice) | Renal Cell Carcinoma (ACHN, Caki-2) ACHN | Naftopidil | Naftopidil, but not tamsulosin, was found to inhibit proliferation of renal cancer cells via induction G1 cell cycle arrest in in vitro studies. |
| [70] | Sakamoto, S. et al. | Anoikis disruption of focal adhesion-Akt signaling impairs renal cell carcinoma | In vitro | Renal cancer 786-0, Caki | Doxazosin and derivatives | Quinazoline-based drugs trigger anoikis in renal cancer cells by targeting the focal adhesion survival signalling. |
| [71] | Takara, K. et al. | Effects of α-adrenoceptor antagonist Doxazosin on MDR1-mediated multidrug resistance and transcellular transport | In vitro | Human cervical carcinoma (HeLa, Hvr100-6) | Doxazosin, prazosin, terazosin | Co-treatment of chemotheraputics (vinblastine and paclitaxel) with doxazosin (1 µM) enhanced chemosensitivity of overexpressing multi-drug resistant HeLa cells, Hvr100-6. On the other hand, prazosin (1 µM) was found to partially reverse cells sensitivity to vinblastine when used in combination, by dose-dependently increasing intracellular accumulation of chemotheraputics. Whereas terazosin had no effect. All other combinations of chemotheraputic and α1-antagonists were found to have little or no effect on chemosensitivity. Over all this study suggests that doxazosin thus may partly reverse drug resistance by inhibiting MDR-1-mediated drug efflux, and in turn, contribute to maintenance of intracellular cytotoxic concentrations. |
| [72] | Powe, D. et al. | α- And β-adrenergic receptor (AR) protein expression is associated with poor clinical outcome in breast cancer: an immunohistochemical study | In vitro | Breast cancer | α-Antagonists | α Antagonists were found to inhibit proliferation and induce apoptosis in vitro. |
| [73] | El Sharkawi, F. et al. | Possible anticancer activity of rosuvastatine, Doxazosin, repaglinide and oxcarbazepin | In vitro | MCF7, HeLa, HepG2,EACC | Doxazosin | Doxazosin was most effective in the EACC line exhibiting 100% inhibition of cell proliferation. Specific mechanisms of action are not reported or discussed. |
| [74] | Kanno, T. et al. | 1-[2-(2-Methoxyphenylamino) ethylamino]-3-(naphthalene-1-yloxy)propan-2-ol as a potential anticancer drug | In vitro | Bladder, prostate, MPM, lung, hepatoma, gastric, renal and colorectal cancer cell lines Caco-2 and CW2 | Naftopidil | This study is discussed in review above. Discuses caspase activation and cell death. |
| [75] | Kaku, Y. et al. | The newly synthesized anticancer drug HUHS1015 is useful for treatment of human gastric cancer | In vitro, in vivo (mice) | Gastric cancer (MKN45 and MKN28) | HUHS1015 (naftopidil analogue) | HUHS1015 treatment caused upregulation of TNFα receptor and apoptosis was observed in both MKN28 and MKN45. However, no caspase activation was observed in MKN28, indicating that HUHS1015 resulted in caspase-dependent and independent apoptosis activity. Mice bearing MKN45 tumours had higher survival rates when treated with HUHS1015 compared to those treated with cisplatin, paclitaxel and irinotecan. |
| [76] | Kaku, Y. et al. | HUHS1015 Suppresses Colonic Cancer Growth by Inducing Necrosis and Apoptosis in Association with Mitochondrial Damage | In vitro, in vivo (mice) | Colon cancer (Caco-2, CW2 cells) | HUHS1015 (naftopidil analogue) | HUHS1015 triggered apoptosis in colon cancer Caco-2 and CW2 cells by disrupting the mitochondrial membrane potential, lowering ATP levels, cytochrome c release, and initiation of the caspase cascade. In addition, HUHS1015 increased the number of cells in sub-G1 phase of cell cycling, which corresponded to apoptosis in both cell lines. In vivo mice studies demonstrated that treatment with HUHS1015, but not naftopidil, delayed colonic tumour growth compared to untreated controls. Furthermore, the authors report 100% survival rate for mice with colonic xenograft tumours treated with HUHS1015 or naftopidil, which was higher than control (89% survival). |
| [77] | Shen, S. et al. | Effects of α-adrenoreceptor antagonists on apoptosis and proliferation of pancreatic cancer cells in vitro | In vitro | Pancreatic cancer (PC-2 and PC-3) | Yohimbine and urapidil (α1- and α2-adrenoreceptor antagonists) | Yohimbine induced apoptotic cytotoxicity of both pancreatic PC-3 and PC-3 pancreatic cancer. In contrast, urapidil was only cytotoxic to PC-2 cells. However, the positive control 5-FU, was more cytotoxic than yohimbine in the conditions tested. |
| [78] | Masachika, E. et al. | Naftopidil induces apoptosis in malignant mesothelioma cell lines independently of α1-adrenoceptor blocking | In vitro | Meso-thelial cancer | Naftopidil, prazosin | Naftopidil and prazosin both have the potential to induce apoptosis via activating caspase-3 and caspase-8, but not caspase-9, independent of α1 blocking activity in mesothelioma cells. |
| [79] | Fuchs, R. et al. | The cytotoxicity of the α1-adrenoceptor antagonist prazosin is linked to an endocytotic mechanism equivalent to transport-P | In vitro | K562 cells erythroleukemia, LNCaP (PCa) | Prazosin/QAPB (fluorescent analogue of prazosin) | Prazosin has been shown to be a substrate for an amine uptake mechanism called transport-P. The fluorescent analogue of prazosin, QAPB was associated with endocytic mechanism of prazosin/QAPB similar transport-P. Prazosin/QAPB was able to induce caspase 8 activation (apoptosis) and tabulation of lysosomes in LNCaP cells. The cytotoxic actions of prazosin was inhibited by chloroquine (a lysomototropic drug) and bafilomycin (transport-P inhibitor). This indicates that transport-p-mediated uptake, and subsequent endosome/lysosome accumulation and caspase activation underlies prazosin-induced LNCaP and/or K562 toxicity. |
| [80] | Albinana, V. et al. | Propranolol reduces viability and induces apoptosis in hemanglioblastoma cells from von Hippel-Lindau patients | In vitro | Hemanglio-blastoma, cervical cancer HeLa9XHRE | Propranolol (β-blocker) | Propranolol treatment resulted in cytotoxicity and caspase-mediated apoptosis (50–100 µM, 48 h treatment) of hypoxia response element-transfected HeLa 9XHRE cells. Similar findings were also observed in hemanglioblastoma cells. Overall, the authors suggests these effects may due in part to the inhibitory effect of HIF1 transcription and protein expression in HeLA9XHRE and hemanglioblastoma cells. |
| [81] | Staudacher, I. et al. | HERG K+ channel-dependent apoptosis and cell cycle arrest in human glioblastoma cells | In vitro | Glioblastoma (LNT-229, U87MG) | Doxazosin, terazosin | Doxazosin was found to induce apoptosis and G0/G1 cell cycle arrest of glioblastoma LNT-229 and U87MG cells in a time and concentration dependent manner. Also, blocking of doxazosin binding to hERG by the non-apoptotic hERG ligand, terazosin, rescued glioblastoma cells from doxazosin-induced apoptosis. The apoptotic effect of doxazosin was marked by the activation of pro-apoptotic factors/signalling (phospho-erythropoietin-producing human hepatocellular carcinoma receptor tyrosine kinase A2, phospho-p38 mitogen-activated protein kinase, growth arrest and DNA damage inducible gene 153, cleaved caspases 9, 7, and 3), and by inactivation of anti- apoptotic poly-ADP-ribose-polymerase, respectively. Overall, this study suggests doxazosin is a hERG antagonist, which results in the activation of apoptotic signaling cascade. |
| [82] | Fuchs, R. et al. | The anti-hypertensive drug prazosin induces apoptosis in the medullary thyroid carcinoma cell line TT | In vitro | Medullary thyroid carcinoma | Prazosin | Prazosin (24 h, ≥15 µM) was found to induce caspase-3/7 activation and apoptosis of medullary thyroid carcinoma cells (α1A and α1B adrenoceptors-positive). This cytotoxicity was associated with morphological changes such as long polar needle-shaped polar protrusion fibers, an increased in number of intracellular vacuoles and detachment. The fibres present in treated cells seem to impair mobility of the cell and were associated with prazosin-mediated caspase activation. Prazosin was also found to have a similar morphological effect on normal human fibroblasts, suggesting a lack of specificity and risk of cytotoxicity to non-cancerous cells. |
| [83] | Tahmatzopoulos, A. et al. | Effect of terazosin on tissue vascularity and apoptosis in transitional cell carcinoma of bladder | Observational Cohort | Transitional cell carcinoma (TCC) of the bladder | Terazosin | Pathological specimens of 24 men who underwent radical cystectomy for transitional cell carcinoma of the bladder were evaluated for terazosin-induced anti-cancer effects. For this study, patients with a history of 5a-reductase inhibitor use were excluded. For men who were never exposed to terazosin (15 men), markers of apoptosis were limited in the tumour specimens of these men. In contrast, terazosin exposure prior to cystectomy (9 men, 2–10 mg/day; 3–60 months) was associated with a statistically significant increase in tumour apoptosis. Terazosin treatment also significantly decreased microvascular density (MVD) in approximately 27% of specimens compared to specimens of unexposed men. |
| N/A | Bajek, A. et al. (2011) | Prostate epithelial stem cells are resistant to apoptosis after α1-antagonist treatment. The impact for BPH patients | In vitro | Prostate cancer | Doxazosin | Doxazosin induced apoptosis in co-cultures of progenitor (type of stem cell) and differentiated epithelial cells. However, progenitor cells were not susceptible to apoptosis, which can be a reason of treatment failure in BPH patients. |
| N/A | Minarini, A. et al. (2006) | Recent advances in the design and synthesis of prazosin derivatives | - | - | Found to be irrelevant to our research but still of interest. |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Batty, M.; Pugh, R.; Rathinam, I.; Simmonds, J.; Walker, E.; Forbes, A.; Anoopkumar-Dukie, S.; McDermott, C.M.; Spencer, B.; Christie, D.; et al. The Role of α1-Adrenoceptor Antagonists in the Treatment of Prostate and Other Cancers. Int. J. Mol. Sci. 2016, 17, 1339. https://doi.org/10.3390/ijms17081339
Batty M, Pugh R, Rathinam I, Simmonds J, Walker E, Forbes A, Anoopkumar-Dukie S, McDermott CM, Spencer B, Christie D, et al. The Role of α1-Adrenoceptor Antagonists in the Treatment of Prostate and Other Cancers. International Journal of Molecular Sciences. 2016; 17(8):1339. https://doi.org/10.3390/ijms17081339
Chicago/Turabian StyleBatty, Mallory, Rachel Pugh, Ilampirai Rathinam, Joshua Simmonds, Edwin Walker, Amanda Forbes, Shailendra Anoopkumar-Dukie, Catherine M. McDermott, Briohny Spencer, David Christie, and et al. 2016. "The Role of α1-Adrenoceptor Antagonists in the Treatment of Prostate and Other Cancers" International Journal of Molecular Sciences 17, no. 8: 1339. https://doi.org/10.3390/ijms17081339
APA StyleBatty, M., Pugh, R., Rathinam, I., Simmonds, J., Walker, E., Forbes, A., Anoopkumar-Dukie, S., McDermott, C. M., Spencer, B., Christie, D., & Chess-Williams, R. (2016). The Role of α1-Adrenoceptor Antagonists in the Treatment of Prostate and Other Cancers. International Journal of Molecular Sciences, 17(8), 1339. https://doi.org/10.3390/ijms17081339