
Upregulation of TLRs and IL-6 as a Marker in Human Colorectal Cancer

 and
and
Abstract
:1. Introduction
2. Results
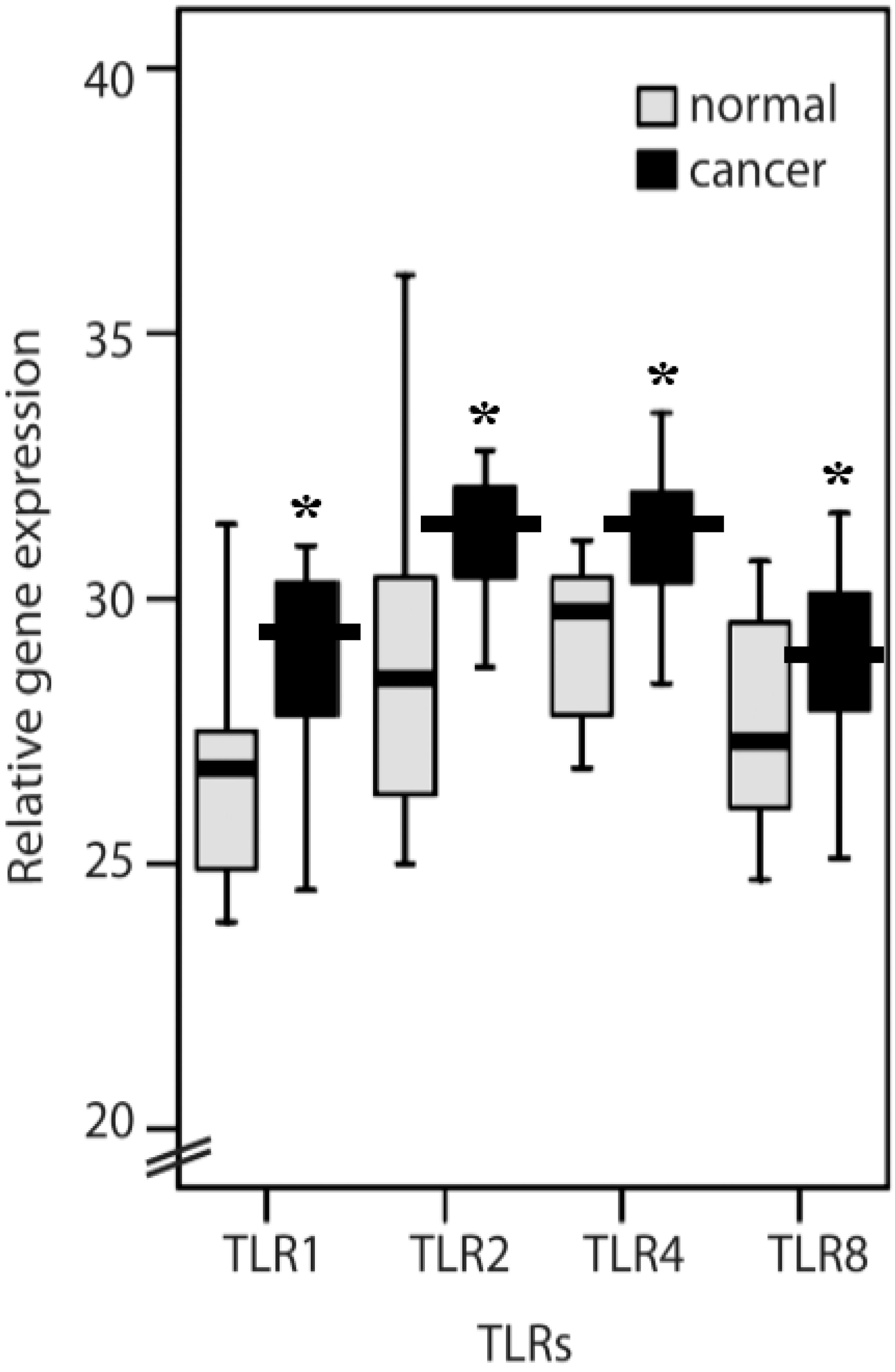
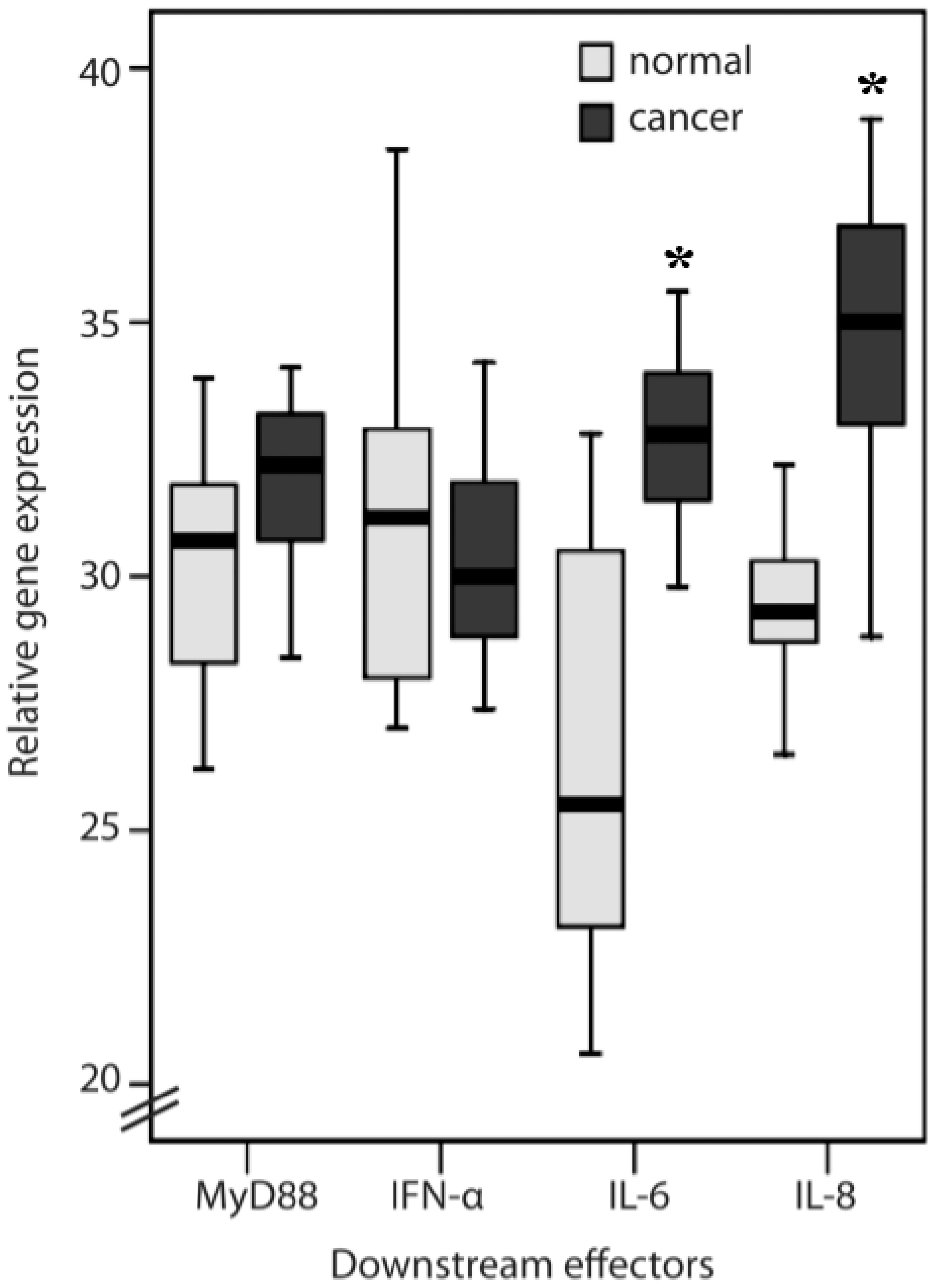
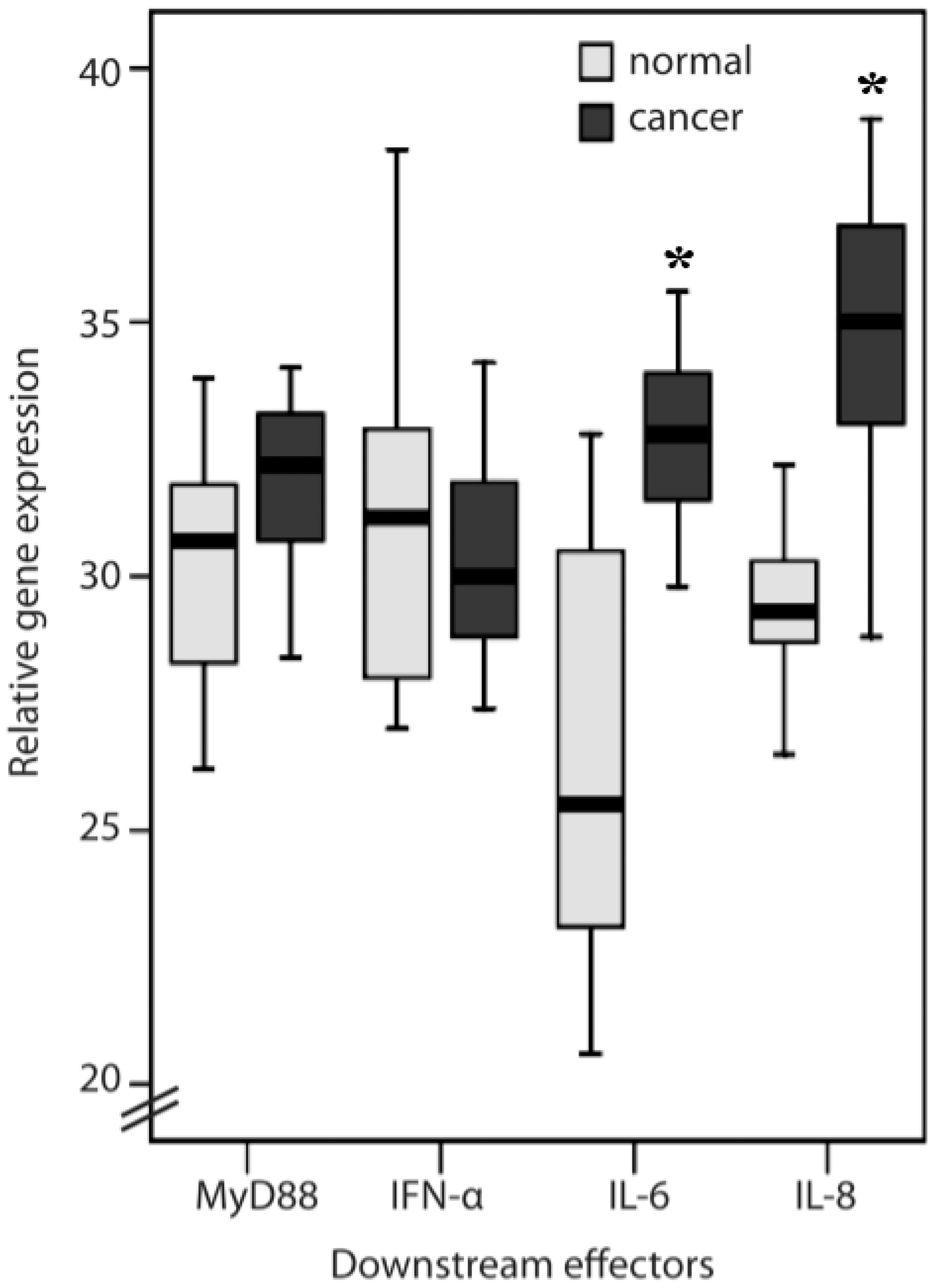
2.1. TLR1, TLR2, TLR4, TLR8, MyD88, IFN-α, IL-6 and IL-8 Expression Levels in Normal Mucosa and Colorectal Cancer Tissues from Patients
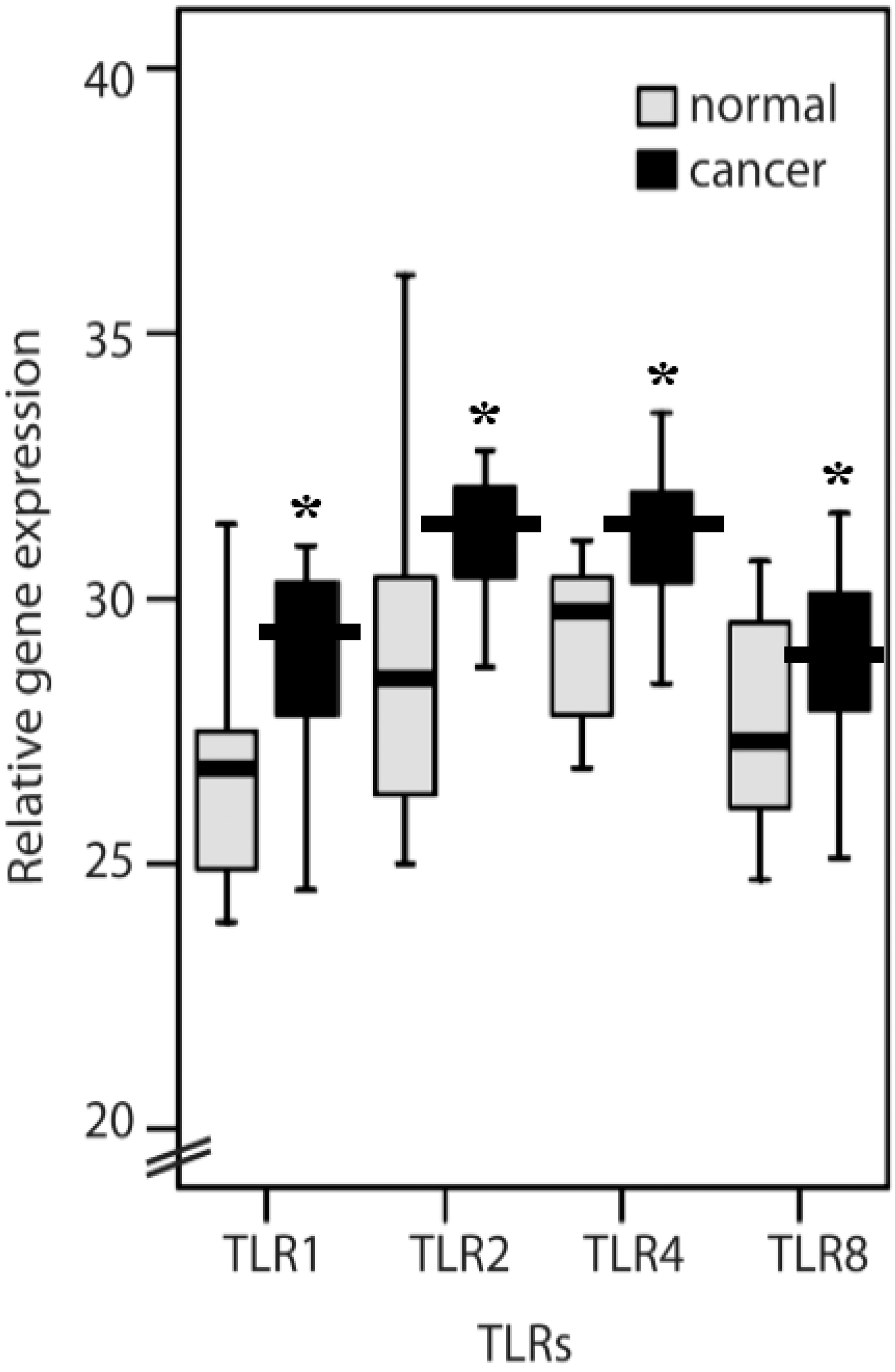
2.2. Clinicopathological Features of TLR1, TLR2, TLR4, TLR8, IL-6, IL-8 and MyD88

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Score * | Normal (n = 16) | Cancer (n = 18) | p-Value |
|---|---|---|---|
| TLR1 | 0.015 | ||
| 1 | 13 | 7 | |
| 2 | 3 | 11 | |
| TLR2 (mucosa) | 0.163 | ||
| 1 | 10 | 15 | |
| 2 | 6 | 3 | |
| TLR2 (lymphocytes) | 0.018 | ||
| 1 | 12 | 6 | |
| 2 | 4 | 12 | |
| TLR4 | 0.571 | ||
| 1 | 13 | 14 | |
| 2 | 3 | 4 | |
| TLR8 | 0.006 | ||
| 1 | 13 | 6 | |
| 2 | 3 | 12 |
| TLR1 | TLR2 (tumor) | TLR4 | TLR8 | IL-6 | IL-8 | MyD88 | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Clinicopathological Factors | High n = 11 | Low n = 7 | p-Value | High n = 9 | Low n = 7 | p-Value | High n = 4 | Low n = 14 | p-Value | High n = 12 | Low n = 6 | p-Value | High n = 8 | Low n = 17 | p-Value | High n = 9 | Low n = 16 | p-Value | High n = 11 | Low n = 7 | p-Value |
| Age (years) | |||||||||||||||||||||
| ≥65 | 7 | 6 | 0.308 | 6 | 4 | 0.696 | 4 | 9 | 0.160 | 8 | 5 | 0.457 | 5 | 11 | 0.915 | 7 | 9 | 0.282 | 6 | 4 | 0.059 |
| <65 | 4 | 1 | 3 | 3 | 0 | 5 | 4 | 1 | 3 | 6 | 2 | 7 | 1 | 6 | |||||||
| Sex | |||||||||||||||||||||
| Male | 9 | 3 | 0.087 | 6 | 5 | 0.838 | 1 | 11 | 0.045 | 8 | 4 | 1.0 | 1 | 7 | 0.152 | 6 | 11 | 0.915 | 5 | 8 | 0.682 |
| Female | 2 | 4 | 3 | 2 | 3 | 3 | 4 | 2 | 7 | 10 | 3 | 5 | 2 | 2 | |||||||
| Tumour Site | |||||||||||||||||||||
| colon | 4 | 3 | 0.783 | 3 | 3 | 0.696 | 3 | 4 | 0.093 | 5 | 2 | 0.732 | 2 | 5 | 0.819 | 2 | 5 | 0.629 | 5 | 2 | 0.034 |
| rectum | 7 | 4 | 6 | 4 | 1 | 10 | 7 | 4 | 6 | 12 | 7 | 11 | 2 | 8 | |||||||
| Histology | |||||||||||||||||||||
| Well | 0 | 1 | 0.387 | 0 | 0 | 0.09 | 1 | 0 | 0.098 | 0 | 1 | 0.339 | 0 | 2 | 0.534 | 0 | 2 | 0.513 | 0 | 1 | 0.155 |
| Moderate | 8 | 5 | 6 | 7 | 3 | 10 | 9 | 4 | 7 | 14 | 8 | 13 | 5 | 9 | |||||||
| Poor/mucinous | 3 | 1 | 3 | 0 | 0 | 4 | 3 | 1 | 1 | 1 | 1 | 1 | 2 | 0 | |||||||
| Stage | |||||||||||||||||||||
| I | 1 | 1 | 0.985 | 0 | 1 | 0.694 | 1 | 1 | 0.376 | 1 | 1 | 0.896 | 1 | 2 | 0.727 | 1 | 2 | 0.657 | 0 | 1 | 0.285 |
| II | 3 | 2 | 3 | 2 | 2 | 3 | 3 | 2 | 2 | 8 | 5 | 5 | 2 | 3 | |||||||
| III | 2 | 1 | 2 | 1 | 0 | 3 | 2 | 1 | 1 | 2 | 1 | 2 | 2 | 0 | |||||||
| IV | 5 | 3 | 4 | 3 | 1 | 7 | 6 | 2 | 4 | 5 | 2 | 7 | 3 | 6 | |||||||
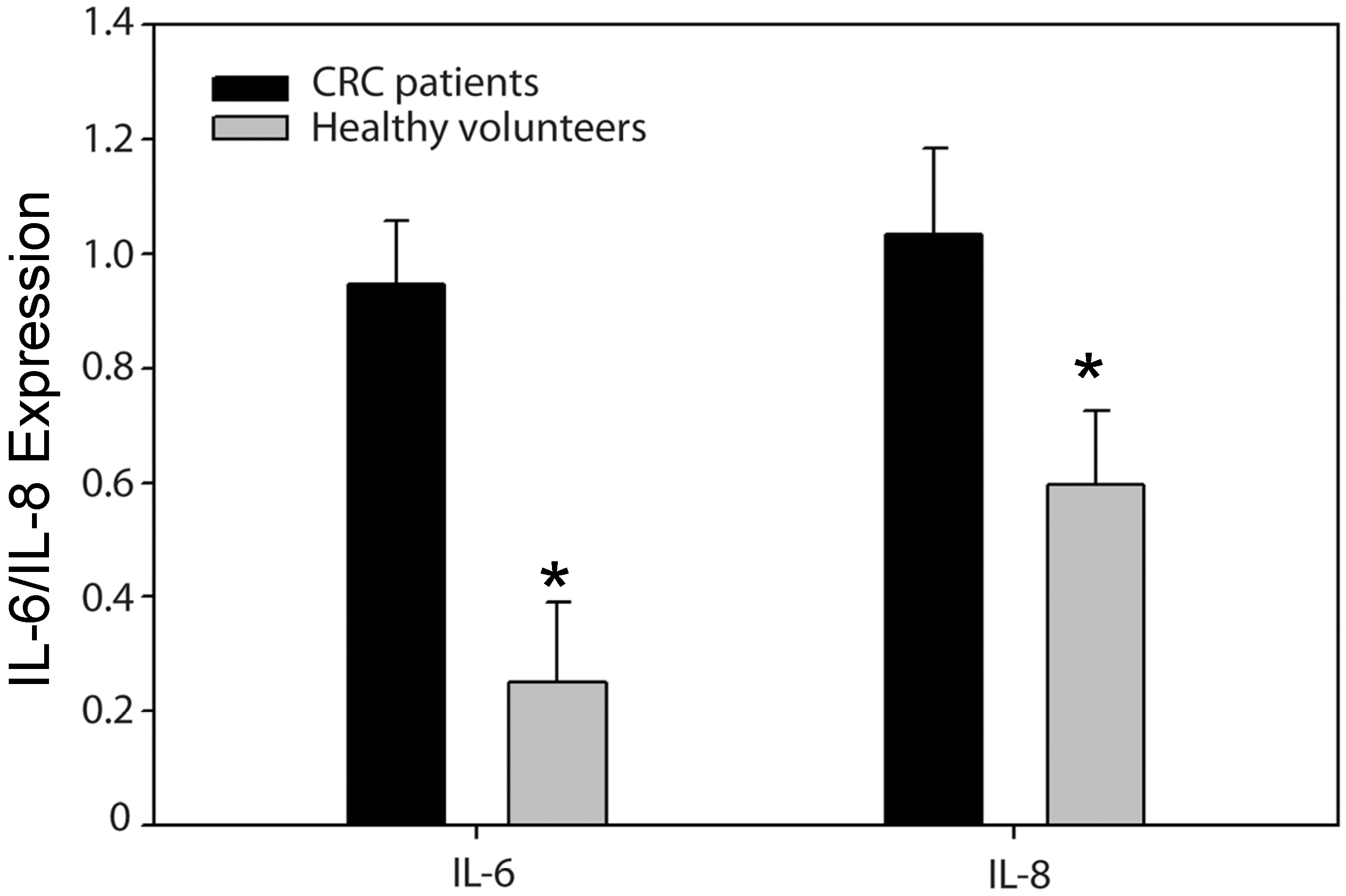
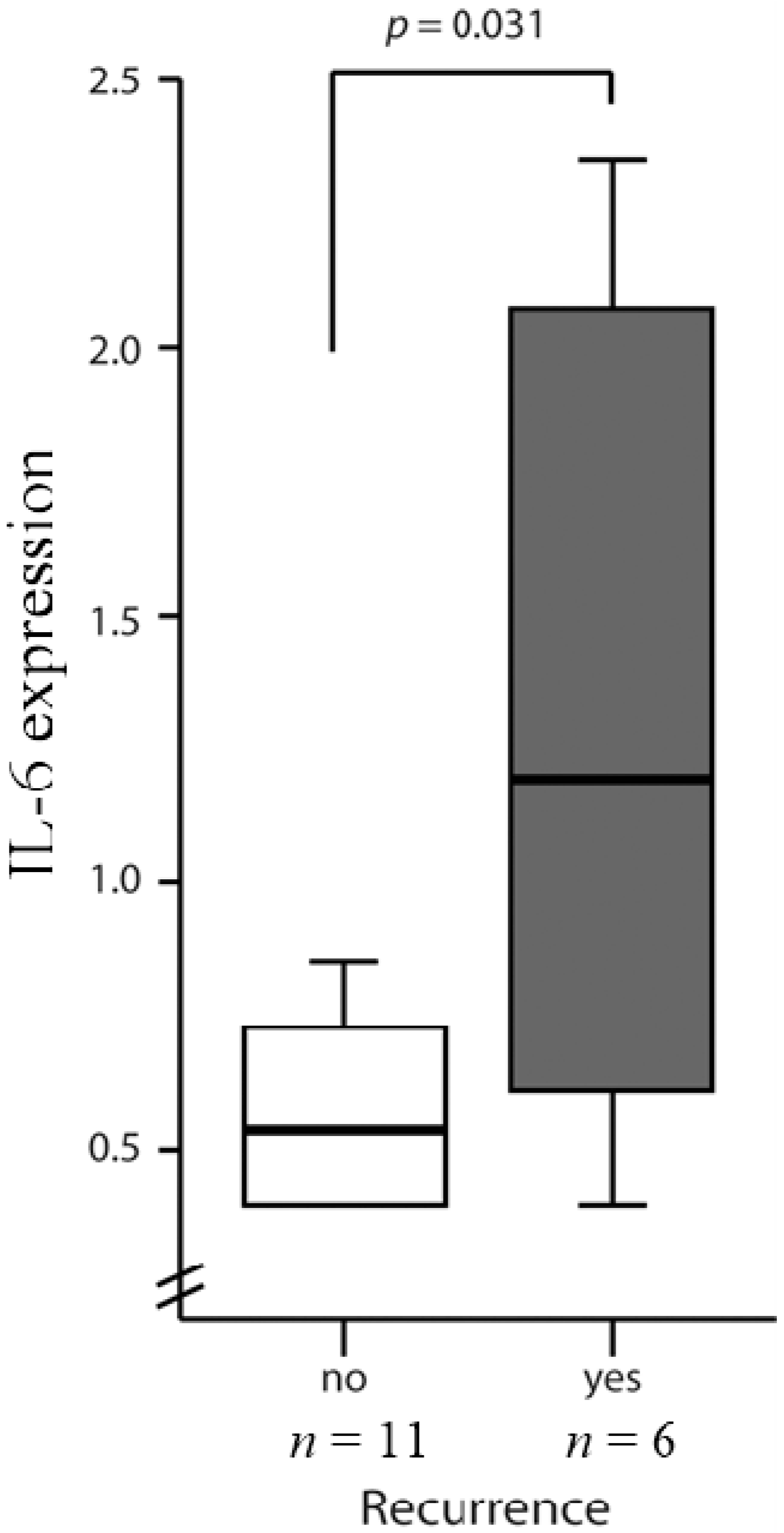
2.3. High Levels of IL-6 and IL-8 Secretion in CRC Patients Correlated with Higher Risks of Recurrence

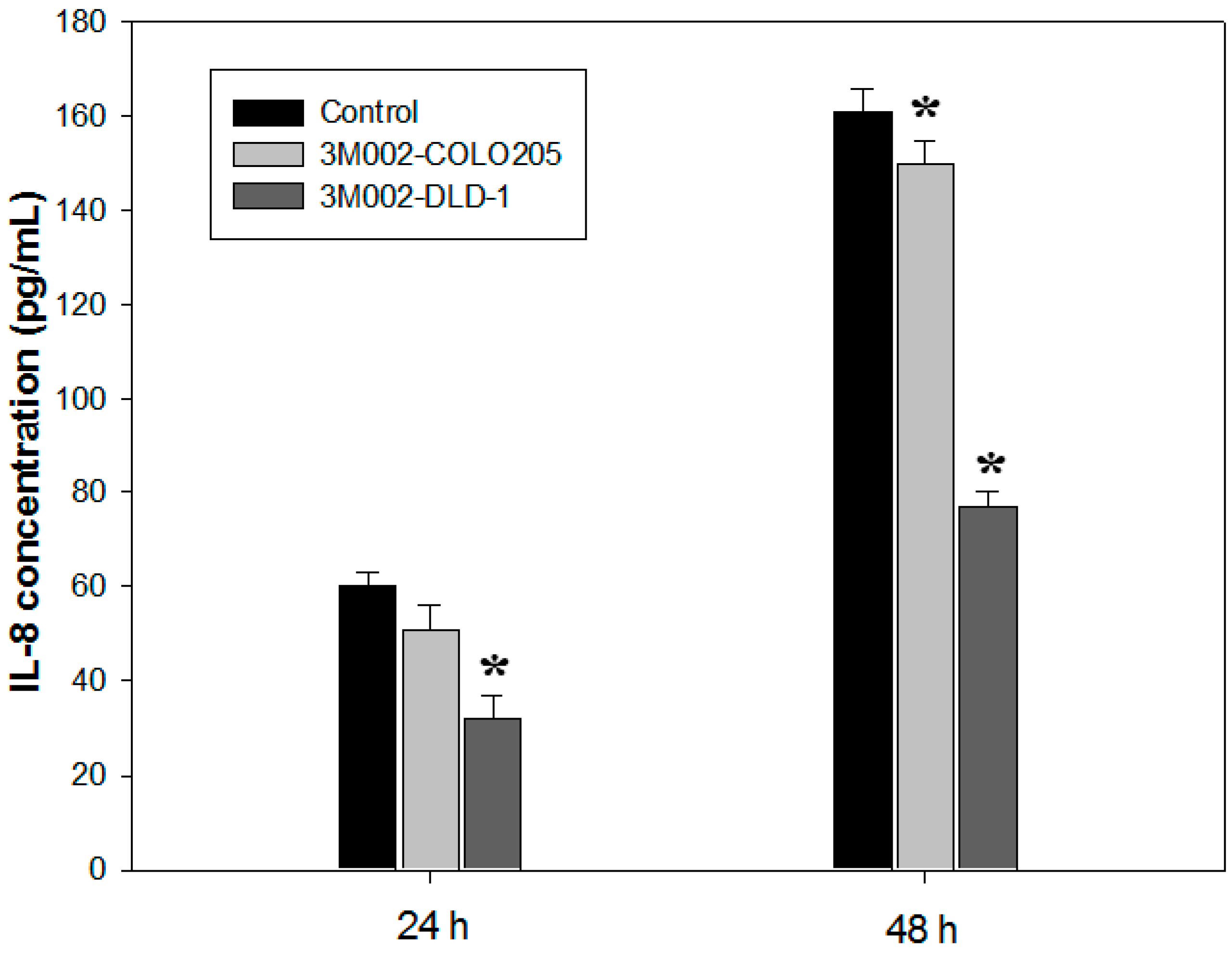
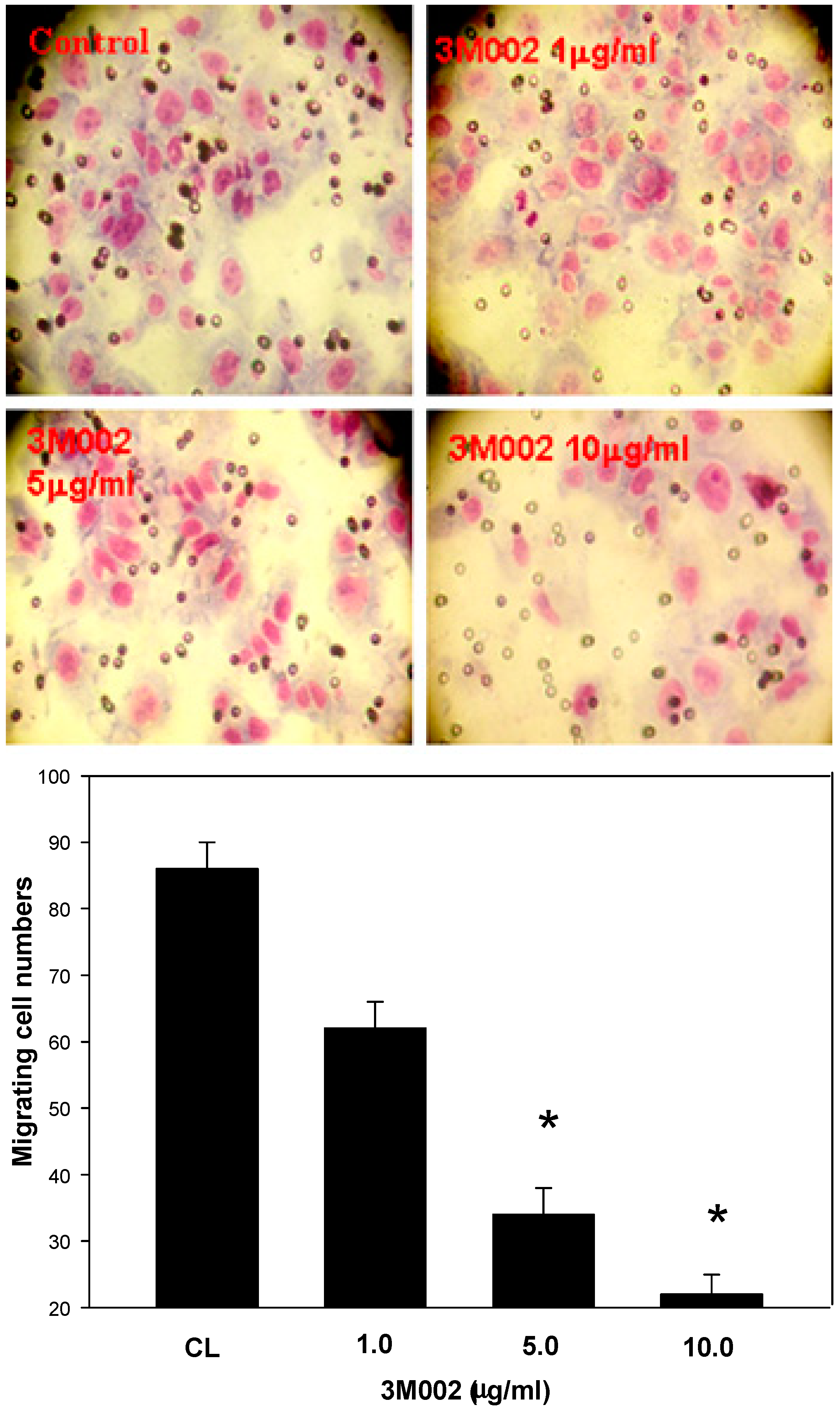
2.4. Release of Proinflammatory Cytokines IL-8 and IL-6 Was Inhibited after 3M002 Treatments

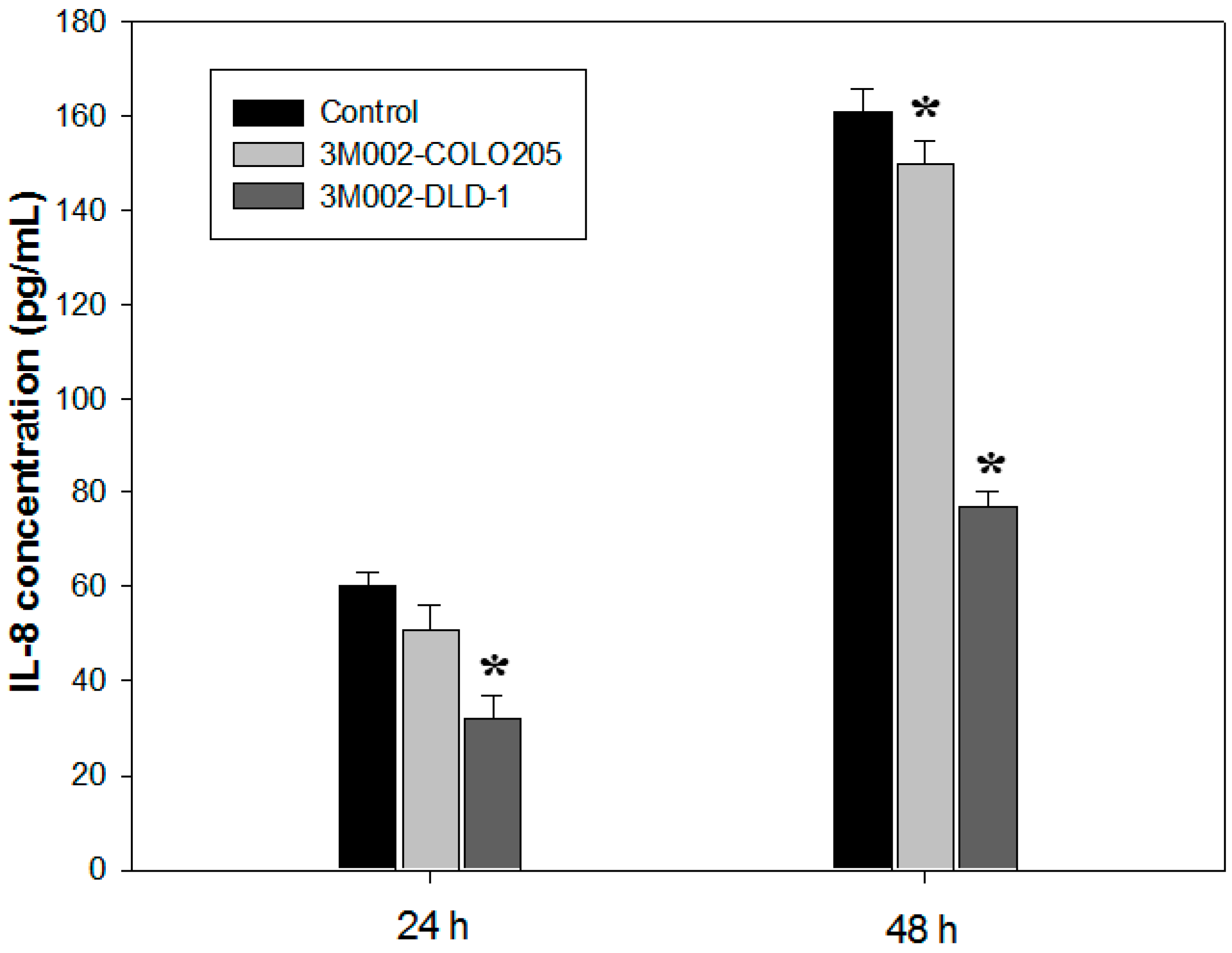
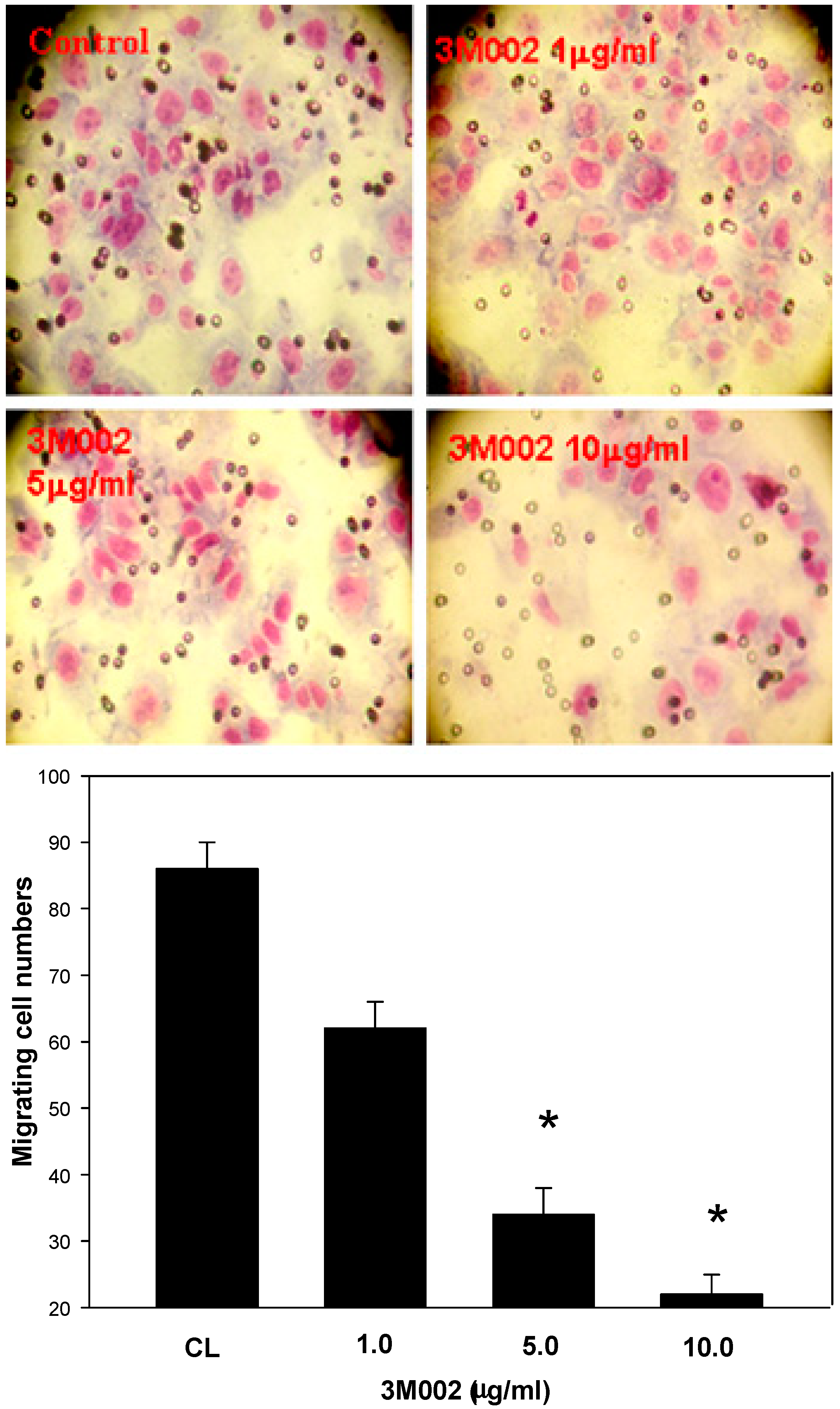
3. Discussion
4. Experimental Section
4.1. Patients and Specimens
4.2. RNA Isolation and cDNA Synthesis
4.3. Real-Time Quantitative PCR
| Name | Sequence | Gene |
|---|---|---|
| TLR1-F | 5'-CAGCAGCCTCAAGCATGTCTA-3' | Rat Toll-like receptor-1 |
| TLR1-R | 5'-CAGCCCTAAGACAACAATACAATAGAAGA-3' | |
| TLR2-F | 5'-GCCAGCAGGTTCAGGATGTC-3' | Human Toll-like receptor-2 |
| TLR2-R | 5'-TGTTCCTGCTGGGAGCTTTC-3' | |
| TLR4-F | 5'-CTTTATTCCCGGTGTGGCCA-3' | Human Toll-like receptor-4 |
| TLR4-R | 5'-GCAGGGTCTTCTCCACCTTC-3' | |
| TLR8-F | 5'-TTTCCCACCTACCCTCTGGCTT-3' | Human Toll-like receptor-8 |
| TLR8-R | 5'-TGCTCTGCATGAGGTTGTCGGATGA-3' | |
| MyD88-F | 5'-CCGCCTGTCTCTGTTCTT-3' | Human MyD88 |
| MyD88-R | 5'-TCCTCCTCAATGCTGGGT-3' | |
| IFN-α-F | 5'-CTATCCCTGTCCTGCATGAGC-3' | interferon-α |
| IFN-α-R | 5'-GGGTTGCATCCCAAGCGT-3' | |
| IL-6-F | 5'-GTCAACTCCATCTGCCCTTCAG-3' | Interleukin-6 |
| IL-6-R | 5'-GGTCTGTTGTGGGTGGTATCCT-3' | |
| IL-8-F | 5'-TCTCTTGGCAGCCTTCCTGA-3' | Interleukin-8 |
| IL-8-R | 5'-CGCAGTGTGGTCCACTCTCA-3' | |
| β-actin-F | 5'-TCACCCACACTGTGCCCATCTACGA-3' | Human β-actin |
| β-actin-R | 5'-CAGCGGAACCGCTCATTGCCAATGG-3' |
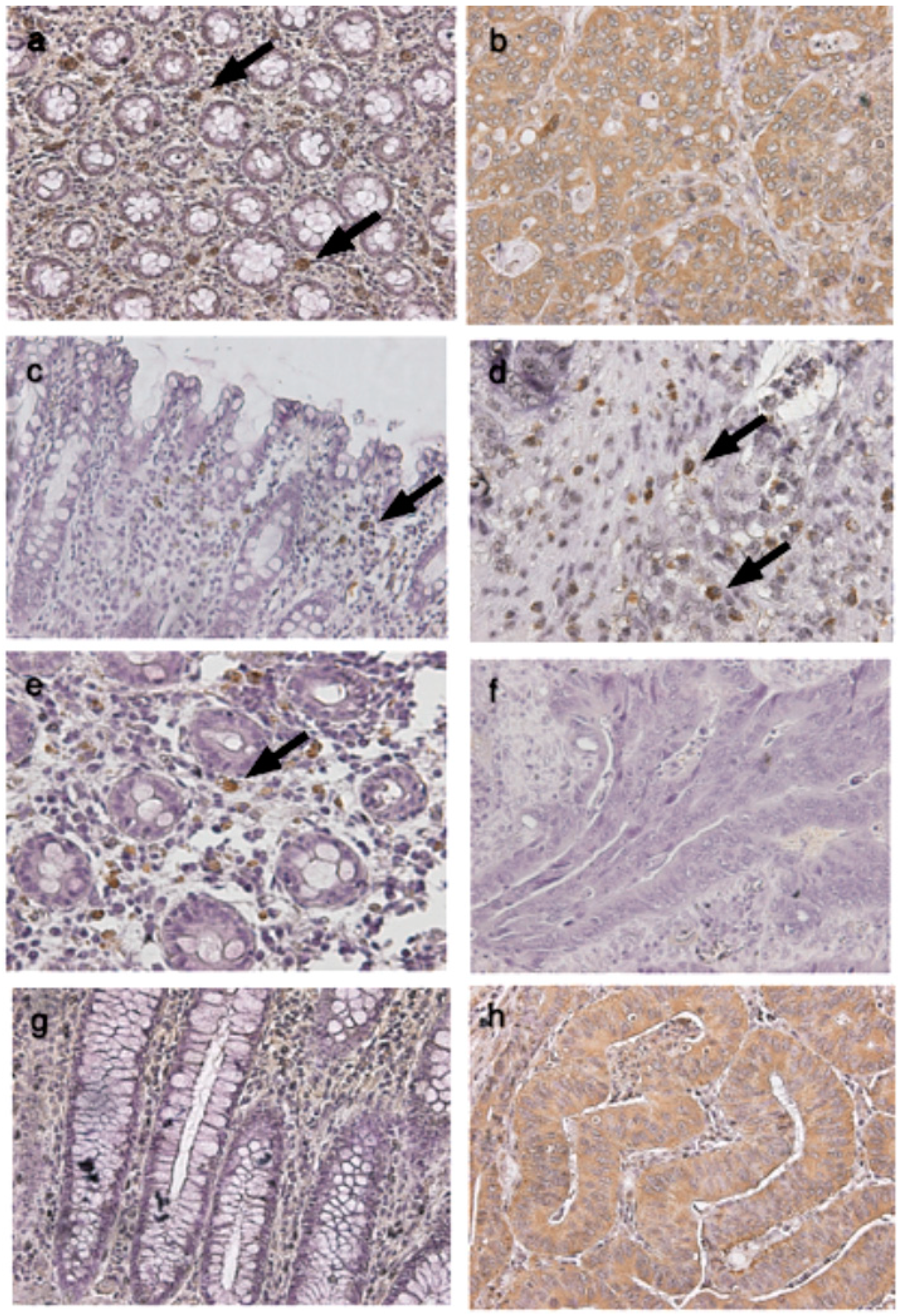
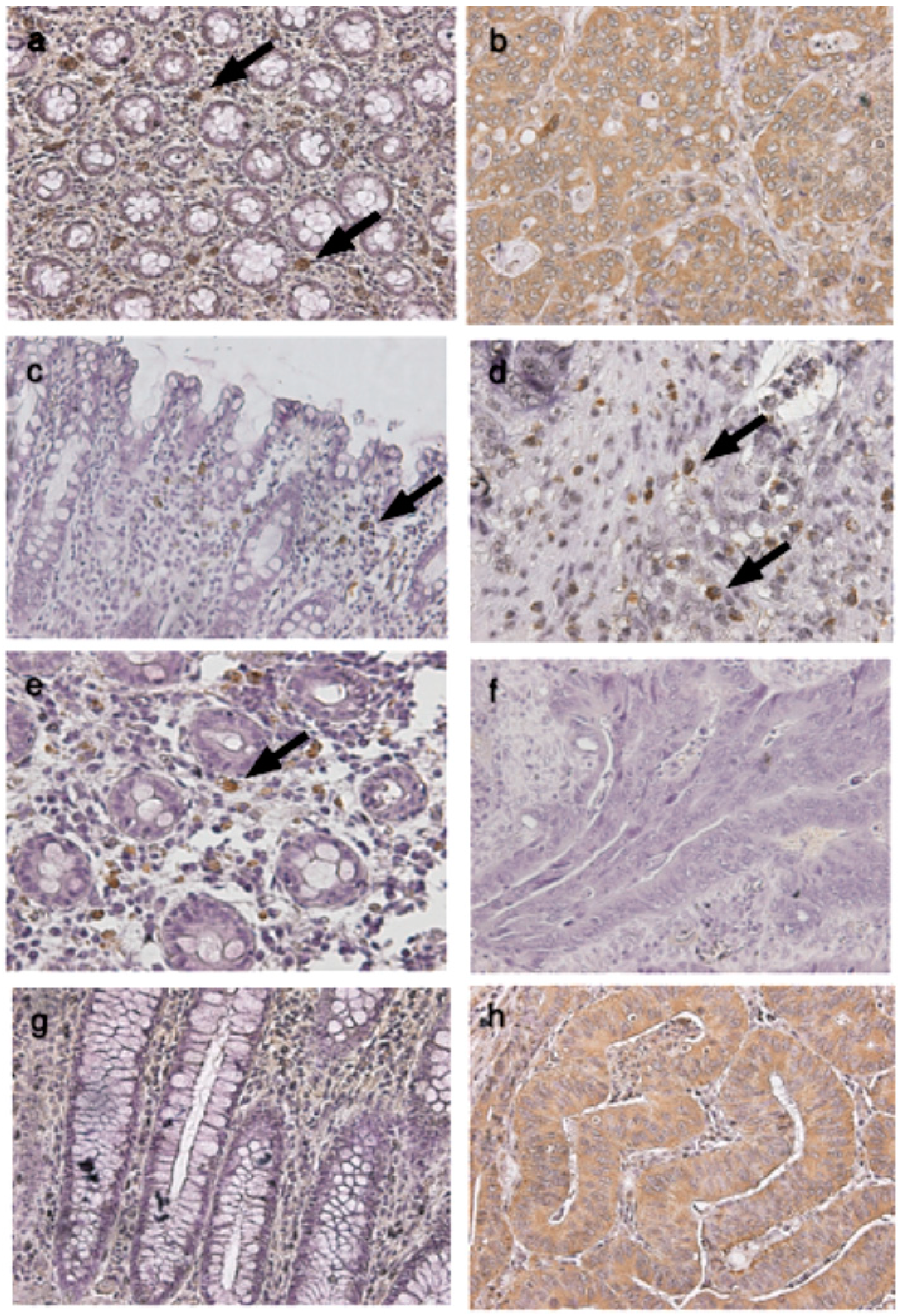
4.4. Immunohistochemical Studies of TLRs in Cancerous and Normal Tissues
4.5. Cell Line, Cell Culture and Chemical Treatment
4.6. Enzyme-Linked Immunosorbent Assay (ELISA)
4.7. Data Analysis and Evaluation
4.8. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Jemal, A.; Siegel, R.; Ward, E.; Hao, Y.; Xu, J.; Thun, M.J. Cancer statistics, 2009. CA Cancer J. Clin. 2009, 59, 225–249. [Google Scholar] [CrossRef]
- Sellon, R.K.; Tonkonogy, S.; Schultz, M.; Dieleman, L.A.; Grenther, W.; Balish, E.; Rennick, D.M.; Sartor, R.B. Resident enteric bacteria are necessary for development of spontaneous colitis and immune system activation in interleukin-10-deficient mice. Infect. Immun. 1998, 66, 5224–5231. [Google Scholar] [PubMed]
- Kado, S.; Uchida, K.; Funabashi, H.; Iwata, S.; Nagata, Y.; Ando, M.; Onoue, M.; Matsuoka, Y.; Ohwaki, M.; Morotomi, M. Intestinal microflora are necessary for development of spontaneous adenocarcinoma of the large intestine in T-cell receptor beta chain and p53 double knockout mice. Cancer Res. 2001, 61, 2395–2398. [Google Scholar] [PubMed]
- Fukata, M.; Hernandez, Y.; Conduah, D.; Cohen, J.; Chen, A.; Breglio, K.; Goo, T.; Hsu, D.; Xu, R.; Abreu, M.T. Innate immune signaling by Toll-like receptor-4 (TLR4) shapes the inflammatory microenvironment in colitis-associated tumors. Inflamm. Bowel Dis. 2009, 15, 997–1006. [Google Scholar] [CrossRef] [PubMed]
- Medzhitov, R.; Preston-Hurlburt, P.; Janeway, C.A., Jr. A human homologue of the Drosophila Toll protein signals activation of adaptive immunity. Nature 1997, 388, 394–397. [Google Scholar] [CrossRef] [PubMed]
- Mitcham, J.L.; Parnet, P.; Bonnert, T.P.; Garka, K.E.; Gerhart, M.J.; Slack, J.L.; Gayle, M.A.; Dower, S.K.; Sims, J.E. T1/ST2 signaling establishes it as a member of an expanding interleukin-1 receptor family. J. Biol. Chem. 1996, 271, 5777–5783. [Google Scholar] [CrossRef] [PubMed]
- Muzio, M.; Bosisio, D.; Polentarutti, N.; D’amico, G.; Stoppacciaro, A.; Mancinelli, R.; van’t Veer, C.; Penton-Rol, G.; Ruco, L.P.; Allavena, P.; et al. Differential expression and regulation of Toll-like receptors (TLR) in human leukocytes: Selective expression of TLR3 in dendritic cells. J. Immunol. 2000, 164, 5998–6004. [Google Scholar] [CrossRef] [PubMed]
- Hoshino, K.; Takeuchi, O.; Kawai, T.; Sanjo, H.; Ogawa, T.; Takeda, Y.; Takeda, K.; Akira, S. Cutting edge: Toll-like receptor 4 (TLR4)-deficient mice are hyporesponsive to lipopolysaccharide: Evidence for TLR4 as the Lps gene product. J. Immunol. 1999, 162, 3749–3752. [Google Scholar] [PubMed]
- Kobayashi, M.; Saitoh, S.; Tanimura, N.; Takahashi, K.; Kawasaki, K.; Nishijima, M.; Fujimoto, Y.; Fukase, K.; Akashi-Takamura, S.; Miyake, K. Regulatory roles for MD-2 and TLR4 in ligand-induced receptor clustering. J. Immunol. 2006, 176, 6211–6218. [Google Scholar] [CrossRef] [PubMed]
- Akira, S. Toll-like receptor signaling. J. Biol. Chem. 2003, 278, 38105–38108. [Google Scholar] [CrossRef] [PubMed]
- Schön, M.P.; Schön, M. TLR7 and TLR8 as targets in cancer therapy. Oncogene 2008, 27, 190–199. [Google Scholar] [CrossRef] [PubMed]
- Heil, F.; Ahmad-Nejad, P.; Hemmi, H.; Hochrein, H.; Ampenberger, F.; Gellert, T.; Dietrich, H.; Lipford, G.; Takeda, K.; Akira, S.; et al. The Toll-like receptor 7 (TLR7)-specific stimulus loxoribine uncovers a strong relationship within the TLR7, 8 and 9 subfamily. Eur. J. Immunol. 2003, 33, 2987–2997. [Google Scholar] [CrossRef] [PubMed]
- Raeburn, C.D.; Calkins, C.M.; Zimmerman, M.A.; Arya, J.; Barsness, K.A.; Harken, A.H. Toll-like receptors and surgical disease. Surgery 2002, 131, 477–483. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, M.; Sato, M. Toll-like receptor signaling in anti-cancer immunity. J. Med. Investig. 2003, 50, 9–24. [Google Scholar]
- Salaun, B.; Coste, I.; Rissoan, M.C.; Lebecque, S.J.; Renno, T. TLR3 can directly trigger apoptosis in human cancer cells. J. Immunol. 2006, 176, 4894–4901. [Google Scholar] [CrossRef] [PubMed]
- Fukata, M.; Chen, A.; Vamadevan, A.S.; Cohen, J.; Breglio, K.; Krishnareddy, S.; Hsu, D.; Xu, R.; Harpaz, N.; Dannenberg, A.J.; et al. Toll-like receptor-4 promotes the development of colitis-associated colorectal tumors. Gastroenterology 2007, 133, 1869–1881. [Google Scholar] [CrossRef] [PubMed]
- Simiantonaki, N.; Kurzik-Dumke, U.; Karyofylli, G.; Jayasinghe, C.; Michel-Schmidt, R.; Kirkpatrick, C.J. Reduced expression of TLR4 is associated with the metastatic status of human colorectal cancer. Int. J. Mol. Med. 2007, 20, 21–29. [Google Scholar] [PubMed]
- Eyking, A.; Ey, B.; Rünzi, M.; Roig, A.I.; Reis, H.; Schmid, K.W.; Gerken, G.; Podolsky, D.K.; Cario, E. Toll-like receptor 4 variant D299G induces features of neoplastic progression in Caco-2 intestinal cells and is associated with advanced human colon cancer. Gastroenterology 2011, 141, 2154–2165. [Google Scholar] [CrossRef] [PubMed]
- Galizia, G.; Orditura, M.; Romano, C.; Lieto, E.; Castellano, P.; Pelosio, L.; Imperatore, V.; Catalano, G.; Pignatelli, C.; de Vita, F. Prognostic significance of circulating IL-10 and IL-6 serum levels in colon cancer patients undergoing surgery. Clin. Immunol. 2002, 102, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Waugh, D.J.; Wilson, C. The interleukin-8 pathway in cancer. Clin. Cancer Res. 2008, 14, 6735–6741. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Sato, S.; Ishii, K.J.; Coban, C.; Hemmi, H.; Yamamoto, M.; Terai, K.; Matsuda, M.; Inoue, J.; Uematsu, S.; et al. Interferon-α induction through Toll-like receptors involves a direct interaction of IRF7 with MyD88 and TRAF6. Nature Immunol. 2004, 5, 1061–1068. [Google Scholar] [CrossRef]
- Uronis, J.M.; Jobin, C. Microbes and colorectal cancer: Is there a relationship? Curr. Oncol. 2009, 16, 22–24. [Google Scholar] [PubMed]
- Doan, H.Q.; Bowen, K.A.; Jackson, L.A.; Evers, B.M. Toll-like receptor 4 activation increases Akt phosphorylation in colon cancer cells. Anticancer Res. 2009, 29, 2473–2478. [Google Scholar] [PubMed]
- Kigerl, K.A.; Lai, W.; Rivest, S.; Hart, R.P.; Satoskar, A.R.; Popovich, P.G. Toll-like receptor (TLR)-2 and TLR-4 regulate inflammation, gliosis, and myelin sparing after spinal cord injury. J. Neurochem. 2007, 102, 37–50. [Google Scholar] [CrossRef] [PubMed]
- Garay, R.P.; Viens, P.; Bauer, J.; Normier, G.; Bardou, M.; Jeannin, J.F.; Chiavaroli, C. Cancer relapse under chemotherapy: Why TLR2/4 receptor agonists can help. Eur. J. Pharmacol. 2007, 563, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Sussman, D.A.; Santaolalla, R.; Bejarano, P.A.; Garcia-Buitrago, M.T.; Perez, M.T.; Abreu, M.T.; Clarke, J. In silico and Ex vivo approaches identify a role for toll-like receptor 4 in colorectal cancer. J. Exp. Clin. Cancer Res. 2014, 33, 45–57. [Google Scholar] [CrossRef] [PubMed]
- Wang, E.L.; Qian, Z.R.; Nakasono, M.; Tanahashi, T.; Yoshimoto, K.; Bando, Y.; Kudo, E.; Shimada, M.; Sano, T. High expression of Toll-like receptor 4/myeloid differentiation factor 88 signals correlates with poor prognosis in colorectal cancer. Br. J. Cancer 2010, 102, 908–915. [Google Scholar] [CrossRef] [PubMed]
- Spitzer, J.H.; Visintin, A.; Mazzoni, A.; Kennedy, M.N.; Segal, D.M. Toll-like receptor 1 inhibits Toll-like receptor 4 signaling in endothelial cells. Eur. J. Immunol. 2002, 32, 1182–1187. [Google Scholar] [CrossRef] [PubMed]
- Uehori, J.; Matsumoto, M.; Tsuji, S.; Akazawa, T.; Takeuchi, O.; Akira, S.; Kawata, T.; Azuma, I.; Toyoshima, K.; Seya, T. Simultaneous blocking of human Toll-like receptor 2 and 4 suppresses myeloid dendritic cell activation induced by Mycobacterium bovis bacillus Calmette-Gue’rin (BCG)-peptidoglycan (PGN). Infect. Immun. 2003, 71, 4238–4249. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, M.; Uematsu, S.; Okamoto, T.; Matsuura, Y.; Sato, S.; Kumar, H.; Satoh, T.; Saitoh, T.; Takeda, K.; Ishii, K.J.; et al. Enhanced TLR-mediated NF-IL6 dependent gene expression by Trib1 deficiency. J. Exp. Med. 2007, 204, 2233–2239. [Google Scholar] [CrossRef] [PubMed]
- Schneider, M.R.; Hoeflich, A.; Fischer, J.R.; Wolf, E.; Sordat, B.; Lahm, H. Interleukin-6 stimulates clonogenic growth of primary and metastatic human colon carcinoma cells. Cancer Lett. 2000, 151, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Wu, Z.; Ren, S.; Wei, Y.; Gao, M.; Randolph, G.J.; Qu, C. TLR8 agonists stimulate newly recruited monocyte-derived cells into potent APCs that enhance HBsAg immunogenicity. Vaccine 2010, 28, 6273–6281. [Google Scholar] [CrossRef] [PubMed]
- Beck, B.; Dörfel, D.; Lichtenegger, F.S.; Geiger, C.; Lindner, L.; Merk, M.; Schendel, D.J.; Subklewe, M. Effects of TLR agonists on maturation and function of 3-day dendritic cells from AML patients in complete remission. J. Transl. Med. 2011, 9, 151–165. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yang, H.; Barnie, P.A.; Yang, P.; Su, Z.; Chen, J.; Jiao, Z.; Lu, L.; Wang, S.; Xu, H. The expression of Toll-like receptor 8 and its relationship with VEGF and Bcl-2 in cervical cancer. Int. J. Med. Sci. 2014, 11, 608–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grimm, M.; Kim, M.; Rosenwald, A.; Heemann, U.; Germer, C.T.; Waaga-Gasser, A.M.; Gasser, M. Toll-like receptor (TLR) 7 and TLR8 expression on CD133+ cells in colorectal cancer points to a specific role for inflammation-induced TLRs in tumourigenesis and tumour progression. Eur. J. Cancer 2010, 46, 2849–2857. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Precopio, M.; Lan, T.; Yu, D.; Tang, J.X.; Kandimalla, E.R.; Agrawal, S. Antitumor activity and immune response induction of a dual agonist of Toll-like receptors 7 and 8. Mol. Cancer Ther. 2010, 9, 1788–1797. [Google Scholar] [CrossRef] [PubMed]
- Peris, K.; Micantonio, T.; Fargnoli, M.C.; Lozzi, G.P.; Chimenti, S. Imiquimod 5% cream in the treatment of Bowen’s disease and invasive squamous cell carcinoma. J. Am. Acad. Dermatol. 2006, 55, 324–327. [Google Scholar] [CrossRef] [PubMed]
- Ning, Y.; Manegold, P.C.; Hong, Y.K.; Zhang, W.; Pohl, A.; Lurje, G.; Winder, T.; Yang, D.; LaBonte, M.J.; Wilson, P.M.; et al. Interleukin-8 is associated with proliferation, migration, angiogenesis and chemosensitivity in vitro and in vivo in colon cancer cell line models. Int. J. Cancer 2010, 1, 2038–2049. [Google Scholar]
- Huang, W.S.; Chin, C.C.; Chen, C.N.; Kuo, Y.H.; Chen, T.C.; Yu, H.R.; Tung, S.Y.; Shen, C.H.; Hsieh, Y.Y.; Guo, S.E.; et al. Stromal cell-derived factor-1/CXC receptor 4 and β1 integrin interaction regulates urokinase-type plasminogen activator expression in human colorectal cancer cells. J. Cell. Physiol. 2012, 227, 1114–1122. [Google Scholar] [CrossRef] [PubMed]
- Chiu, Y.W.; Lin, T.H.; Huang, W.S.; Teng, C.Y.; Liou, Y.S.; Kuo, W.H.; Lin, W.L.; Huang, H.I.; Tung, J.N.; Huang, C.Y.; et al. Baicalein inhibits the migration and invasive properties of human hepatoma cells. Toxicol. Appl. Pharmacol. 2011, 255, 316–326. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, C.-C.; Kuo, H.-C.; Wang, F.-S.; Jou, M.-H.; Lee, K.-C.; Chuang, J.-H. Upregulation of TLRs and IL-6 as a Marker in Human Colorectal Cancer. Int. J. Mol. Sci. 2015, 16, 159-177. https://doi.org/10.3390/ijms16010159
Lu C-C, Kuo H-C, Wang F-S, Jou M-H, Lee K-C, Chuang J-H. Upregulation of TLRs and IL-6 as a Marker in Human Colorectal Cancer. International Journal of Molecular Sciences. 2015; 16(1):159-177. https://doi.org/10.3390/ijms16010159
Chicago/Turabian StyleLu, Chien-Chang, Hsing-Chun Kuo, Feng-Sheng Wang, Ming-Huey Jou, Ko-Chao Lee, and Jiin-Haur Chuang. 2015. "Upregulation of TLRs and IL-6 as a Marker in Human Colorectal Cancer" International Journal of Molecular Sciences 16, no. 1: 159-177. https://doi.org/10.3390/ijms16010159