Emerging Roles for Intersectin (ITSN) in Regulating Signaling and Disease Pathways

Abstract
:1. Introduction
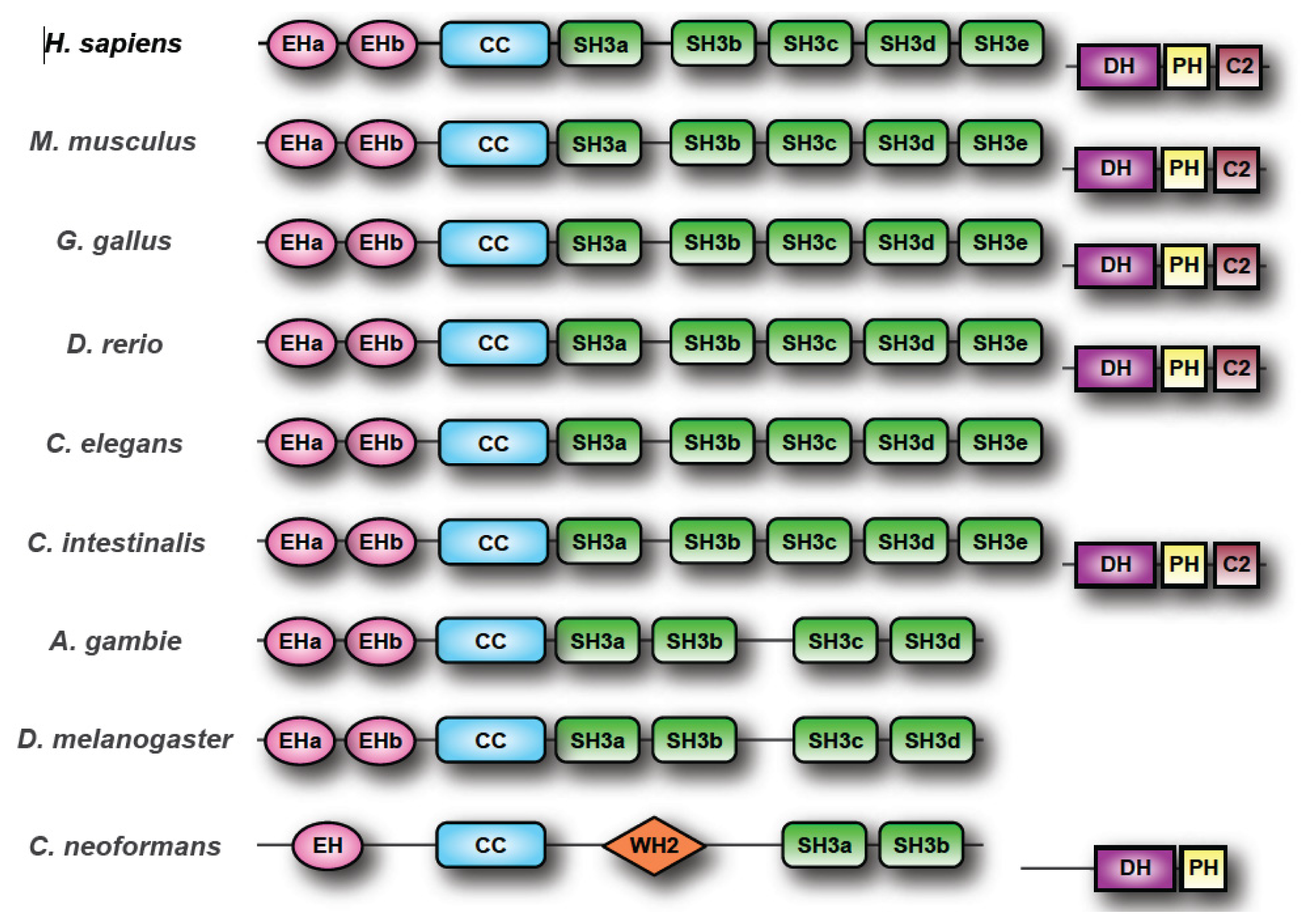
2. The ITSN Gene Family
2.1. ITSN Orthologs
2.2. ITSN Splice Variants and Altered Functions
3. The Role of ITSN in Endocytosis: New Insights
4. A Growing Number of ITSN-Regulated Cell Signaling Pathways
4.1. Identification of Novel ITSN Binding Partners by Yeast Two-Hybrid Screening
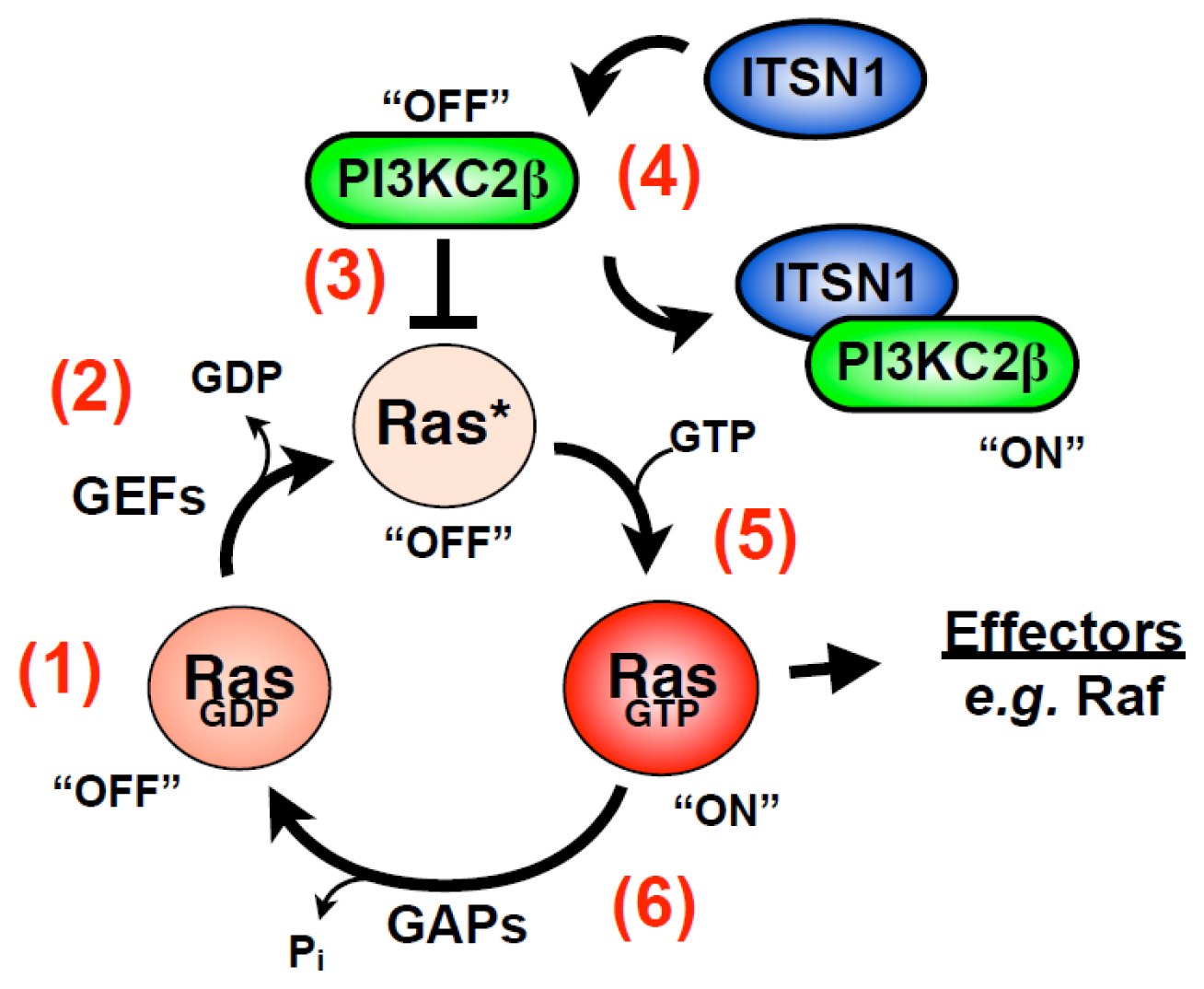
4.2. ITSN Regulates Ras Superfamily Members
4.3. ITSN Regulation of Receptor Tyrosine Kinases (RTK): New Insights
4.4. ITSNs Interact with Herpesvirus Proteins
5. ITSN in Neurobiology: Implications for Down Syndrome (DS) and Alzheimer Disease (AD)
5.1. ITSN1 Regulates Dendritic Spine Morphogenesis and Survival of Neurons
5.2. ITSN1, DS and AD
6. ITSN in Cancer
6.1. ITSN Regulates Oncogenic Signaling Pathways
6.2. ITSN and Asymmetric Cell Division: Maintenance of Cancer Stem Cells
6.3. ITSN’s and Regulation of Receptor Trafficking: Implications for Cancer
7. Future Directions
Acknowledgments
Conflict of Interest
References
- Sparks, A.B.; Hoffman, N.G.; McConnell, S.J.; Fowlkes, D.M.; Kay, B.K. Cloning of ligand targets: Systematic isolation of sh3 domain-containing proteins. Nat. Biotechnol 1996, 14, 741–744. [Google Scholar]
- Yamabhai, M.; Hoffman, N.G.; Hardison, N.L.; McPherson, P.S.; Castagnoli, L.; Cesareni, G.; Kay, B.K. Intersectin, a novel adaptor protein with two eps15 homology and five src homology 3 domains. J. Biol. Chem 1998, 273, 31401–31407. [Google Scholar]
- Yu, Y.; Chu, P.Y.; Bowser, D.N.; Keating, D.J.; Dubach, D.; Harper, I.; Tkalcevic, J.; Finkelstein, D.I.; Pritchard, M.A. Mice deficient for the chromosome 21 ortholog itsn1 exhibit vesicle-trafficking abnormalities. Hum. Mol. Genet 2008, 17, 3281–3290. [Google Scholar]
- Ma, Y.J.; Okamoto, M.; Gu, F.; Obata, K.; Matsuyama, T.; Desaki, J.; Tanaka, J.; Sakanaka, M. Neuronal distribution of ehsh1/intersectin: Molecular linker between clathrin-mediated endocytosis and signaling pathways. J. Neurosci. Res 2003, 71, 468–477. [Google Scholar]
- Guipponi, M.; Scott, H.S.; Chen, H.; Schebesta, A.; Rossier, C.; Antonarakis, S.E. Two isoforms of a human intersectin (itsn) protein are produced by brain-specific alternative splicing in a stop codon. Genomics 1998, 53, 369–376. [Google Scholar]
- Pucharcos, C.; Casas, C.; Nadal, M.; Estivill, X.; de la Luna, S. The human intersectin genes and their spliced variants are differentially expressed. Biochim. Biophys. Acta 2001, 1521, 1–11. [Google Scholar]
- Sengar, A.S.; Wang, W.; Bishay, J.; Cohen, S.; Egan, S.E. The eh and sh3 domain ese proteins regulate endocytosis by linking to dynamin and eps15. EMBO J 1999, 18, 1159–1171. [Google Scholar]
- Pucharcos, C.; Estivill, X.; de la Luna, S. Intersectin 2, a new multimodular protein involved in clathrin-mediated endocytosis. FEBS Lett 2000, 478, 43–51. [Google Scholar]
- Santolini, E.; Salcini, A.E.; Kay, B.K.; Yamabhai, M.; Di Fiore, P.P. The EH network. Exp. Cell Res 1999, 253, 186–209. [Google Scholar]
- Fernandez-Chacon, R.; Achiriloaie, M.; Janz, R.; Albanesi, J.P.; Sudhof, T.C. Scamp1 function in endocytosis. J. Biol. Chem 2000, 275, 12752–12756. [Google Scholar]
- Martina, J.A.; Bonangelino, C.J.; Aguilar, R.C.; Bonifacino, J.S. Stonin 2: An adaptor-like protein that interacts with components of the endocytic machinery. J. Cell Biol 2001, 153, 1111–1120. [Google Scholar]
- Hussain, N.K.; Yamabhai, M.; Ramjaun, A.R.; Guy, A.M.; Baranes, D.; O’Bryan, J.P.; Der, C.J.; Kay, B.K.; McPherson, P.S. Splice variants of intersectin are components of the endocytic machinery in neurons and nonneuronal cells. J. Biol. Chem 1999, 274, 15671–15677. [Google Scholar]
- O’Bryan, J.P. Intersecting pathways in cell biology. Sci. Signal 2010, 3(re 10). [Google Scholar]
- Snyder, J.T.; Worthylake, D.K.; Rossman, K.L.; Betts, L.; Pruitt, W.M.; Siderovski, D.P.; Der, C.J.; Sondek, J. Structural basis for the selective activation of rho gtpases by dbl exchange factors. Nat. Struct. Biol 2002, 9, 468–475. [Google Scholar]
- Hussain, N.K.; Jenna, S.; Glogauer, M.; Quinn, C.C.; Wasiak, S.; Guipponi, M.; Antonarakis, S.E.; Kay, B.K.; Stossel, T.P.; Lamarche-Vane, N.; et al. Endocytic protein intersectin-l regulates actin assembly via cdc42 and n-wasp. Nat. Cell Biol 2001, 3, 927–932. [Google Scholar]
- Zamanian, J.L.; Kelly, R.B. Intersectin 1l guanine nucleotide exchange activity is regulated by adjacent src homology 3 domains that are also involved in endocytosis. Mol. Biol. Cell 2003, 14, 1624–1637. [Google Scholar]
- McGavin, M.K.; Badour, K.; Hardy, L.A.; Kubiseski, T.J.; Zhang, J.; Siminovitch, K.A. The intersectin 2 adaptor links wiskott aldrich syndrome protein (wasp)-mediated actin polymerization to t cell antigen receptor endocytosis. J. Exp. Med 2001, 194, 1777–1787. [Google Scholar]
- Shen, G.; Whittington, A.; Song, K.; Wang, P. Pleiotropic function of intersectin homologue cin1 in cryptococcus neoformans. Mol. Microbiol 2010, 76, 662–676. [Google Scholar]
- Amores, A.; Force, A.; Yan, Y.L.; Joly, L.; Amemiya, C.; Fritz, A.; Ho, R.K.; Langeland, J.; Prince, V.; Wang, Y.L.; et al. Zebrafish hox clusters and vertebrate genome evolution. Science 1998, 282, 1711–1714. [Google Scholar]
- Novokhatska, O.; Dergai, M.; Houssin, N.; Tsyba, L.; Moreau, J.; Rynditch, A. Intersectin 2 nucleotide exchange factor regulates cdc42 activity during xenopus early development. Biochem. Biophys. Res. Commun 2011, 408, 663–668. [Google Scholar]
- Tsyba, L.; Nikolaienko, O.; Dergai, O.; Dergai, M.; Novokhatska, O.; Skrypkina, I.; Rynditch, A. Intersectin multidomain adaptor proteins: Regulation of functional diversity. Gene 2011, 473, 67–75. [Google Scholar]
- Tong, X.K.; Hussain, N.K.; de Heuvel, E.; Kurakin, A.; Abi-Jaoude, E.; Quinn, C.C.; Olson, M.F.; Marais, R.; Baranes, D.; Kay, B.K.; et al. The endocytic protein intersectin is a major binding partner for the ras exchange factor msos1 in rat brain. EMBO J 2000, 19, 1263–1271. [Google Scholar]
- Martin, N.P.; Mohney, R.P.; Dunn, S.; Das, M.; Scappini, E.; O’Bryan, J.P. Intersectin regulates epidermal growth factor receptor endocytosis, ubiquitylation, and signaling. Mol. Pharmacol 2006, 70, 1643–1653. [Google Scholar]
- Tsyba, L.; Gryaznova, T.; Dergai, O.; Dergai, M.; Skrypkina, I.; Kropyvko, S.; Boldyryev, O.; Nikolaienko, O.; Novokhatska, O.; Rynditch, A. Alternative splicing affecting the sh3a domain controls the binding properties of intersectin 1 in neurons. Biochem. Biophys. Res. Commun 2008, 372, 929–934. [Google Scholar]
- Dergai, M.; Tsyba, L.; Dergai, O.; Zlatskii, I.; Skrypkina, I.; Kovalenko, V.; Rynditch, A. Microexon-based regulation of itsn1 and src sh3 domains specificity relies on introduction of charged amino acids into the interaction interface. Biochem. Biophys. Res. Commun 2010, 399, 307–312. [Google Scholar]
- Kropyvko, S.; Gerasymchuk, D.; Skrypkina, I.; Dergai, M.; Dergai, O.; Nikolaienko, O.; Rynditch, A.; Tsyba, L. Structural diversity and differential expression of novel human intersectin 1 isoforms. Mol. Biol. Rep 2010, 37, 2789–2796. [Google Scholar]
- Shen, G.; Zhou, E.; Alspaugh, J.A.; Wang, P. Wsp1 is downstream of cin1 and regulates vesicle transport and actin cytoskeleton as an effector of cdc42 and rac1 in cryptococcus neoformans. Eukaryot. Cell 2012, 11, 471–481. [Google Scholar]
- Wang, W.; Bouhours, M.; Gracheva, E.O.; Liao, E.H.; Xu, K.; Sengar, A.S.; Xin, X.; Roder, J.; Boone, C.; Richmond, J.E.; et al. Itsn-1 controls vesicle recycling at the neuromuscular junction and functions in parallel with dab-1. Traffic 2008, 9, 742–754. [Google Scholar]
- Rose, S.; Malabarba, M.G.; Krag, C.; Schultz, A.; Tsushima, H.; Di Fiore, P.P.; Salcini, A.E. Caenorhabditis elegans intersectin: A synaptic protein regulating neurotransmission. Mol. Biol. Cell 2007, 18, 5091–5099. [Google Scholar]
- Roos, J.; Kelly, R.B. Dap160, a neural-specific eps15 homology and multiple sh3 domain-containing protein that interacts with drosophila dynamin. J. Biol. Chem 1998, 273, 19108–19119. [Google Scholar]
- Koh, T.W.; Verstreken, P.; Bellen, H.J. Dap160/intersectin acts as a stabilizing scaffold required for synaptic development and vesicle endocytosis. Neuron 2004, 43, 193–205. [Google Scholar]
- Marie, B.; Sweeney, S.T.; Poskanzer, K.E.; Roos, J.; Kelly, R.B.; Davis, G.W. Dap160/intersectin scaffolds the periactive zone to achieve high-fidelity endocytosis and normal synaptic growth. Neuron 2004, 43, 207–219. [Google Scholar]
- Thomas, S.; Ritter, B.; Verbich, D.; Sanson, C.; Bourbonniere, L.; McKinney, R.A.; McPherson, P.S. Intersectin regulates dendritic spine development and somatodendritic endocytosis but not synaptic vesicle recycling in hippocampal neurons. J. Biol. Chem 2009, 284, 12410–12419. [Google Scholar]
- Predescu, S.A.; Predescu, D.N.; Knezevic, I.; Klein, I.K.; Malik, A.B. Intersectin-1s regulates the mitochondrial apoptotic pathway in endothelial cells. J. Biol. Chem 2007, 282, 17166–17178. [Google Scholar]
- Predescu, S.A.; Predescu, D.N.; Timblin, B.K.; Stan, R.V.; Malik, A.B. Intersectin regulates fission and internalization of caveolae in endothelial cells. Mol. Biol. Cell 2003, 14, 4997–5010. [Google Scholar]
- Okamoto, M.; Schoch, S.; Sudhof, T.C. Ehsh1/intersectin, a protein that contains eh and sh3 domains and binds to dynamin and snap-25. A protein connection between exocytosis and endocytosis? J. Biol. Chem 1999, 274, 18446–18454. [Google Scholar]
- Solomaha, E.; Szeto, F.L.; Yousef, M.A.; Palfrey, H.C. Kinetics of src homology 3 domain association with the proline-rich domain of dynamins: Specificity, occlusion, and the effects of phosphorylation. J. Biol. Chem 2005, 280, 23147–23156. [Google Scholar]
- Oh, P.; McIntosh, D.P.; Schnitzer, J.E. Dynamin at the neck of caveolae mediates their budding to form transport vesicles by gtp-driven fission from the plasma membrane of endothelium. J. Cell Biol 1998, 141, 101–114. [Google Scholar]
- Knezevic, I.; Predescu, D.; Bardita, C.; Wang, M.; Sharma, T.; Keith, B.; Neamu, R.; Malik, A.; Predescu, S. Regulation of dynamin-2 assembly-disassembly and function through the sh3a domain of intersectin-1s. J. Cell. Mol. Med 2011, 15, 2364–2376. [Google Scholar]
- Predescu, D.N.; Neamu, R.; Bardita, C.; Wang, M.; Predescu, S.A. Impaired caveolae function and upregulation of alternative endocytic pathways induced by experimental modulation of intersectin-1s expression in mouse lung endothelium. Biochem. Res. Int 2012, 2012, 672705. [Google Scholar]
- Evergren, E.; Gad, H.; Walther, K.; Sundborger, A.; Tomilin, N.; Shupliakov, O. Intersectin is a negative regulator of dynamin recruitment to the synaptic endocytic zone in the central synapse. J. Neurosci 2007, 27, 379–390. [Google Scholar]
- Pechstein, A.; Bacetic, J.; Vahedi-Faridi, A.; Gromova, K.; Sundborger, A.; Tomlin, N.; Krainer, G.; Vorontsova, O.; Schafer, J.G.; Owe, S.G.; et al. Regulation of synaptic vesicle recycling by complex formation between intersectin 1 and the clathrin adaptor complex ap2. Proc. Natl. Acad. Sci. USA 2010, 107, 4206–4211. [Google Scholar]
- Dergai, O.; Novokhatska, O.; Dergai, M.; Skrypkina, I.; Tsyba, L.; Moreau, J.; Rynditch, A. Intersectin 1 forms complexes with sgip1 and reps1 in clathrin-coated pits. Biochem. Biophys. Res. Commun 2010, 402, 408–413. [Google Scholar]
- Henne, W.M.; Boucrot, E.; Meinecke, M.; Evergren, E.; Vallis, Y.; Mittal, R.; McMahon, H.T. Fcho proteins are nucleators of clathrin-mediated endocytosis. Science 2010, 328, 1281–1284. [Google Scholar]
- Wong, K.A.; Wilson, J.; Russo, A.; Wang, L.; Okur, M.N.; Wang, X.; Martin, N.P.; Scappini, E.; Carnegie, G.K.; O’Bryan, J.P. Intersectin (itsn) family of scaffolds function as molecular hubs in protein interaction networks. PLoS One 2012, 7, e36023. [Google Scholar]
- Morderer, D.; Nikolaienko, O.; Skrypkina, I.; Cherkas, V.; Tsyba, L.; Belan, P.; Rynditch, A. Endocytic adaptor protein intersectin 1 forms a complex with microtubule stabilizer stop in neurons. Gene 2012, 505, 360–364. [Google Scholar]
- Soares, D.C.; Carlyle, B.C.; Bradshaw, N.J.; Porteous, D.J. Disc1: Structure, function, and therapeutic potential for major mental illness. ACS Chem. Neurosci 2011, 2, 609–632. [Google Scholar]
- Jenna, S.; Hussain, N.K.; Danek, E.I.; Triki, I.; Wasiak, S.; McPherson, P.S.; Lamarche-Vane, N. The activity of the gtpase-activating protein cdgap is regulated by the endocytic protein intersectin. J. Biol. Chem 2002, 277, 6366–6373. [Google Scholar]
- Ahmad, K.F.; Lim, W.A. The minimal autoinhibited unit of the guanine nucleotide exchange factor intersectin. PLoS One 2010, 5, e11291. [Google Scholar]
- Kintscher, C.; Wuertenberger, S.; Eylenstein, R.; Uhlendorf, T.; Groemping, Y. Autoinhibition of gef activity in intersectin 1 is mediated by the short sh3-dh domain linker. Protein Sci 2010, 19, 2164–2174. [Google Scholar]
- Jaffe, A.B.; Kaji, N.; Durgan, J.; Hall, A. Cdc42 controls spindle orientation to position the apical surface during epithelial morphogenesis. J. Cell Biol 2008, 183, 625–633. [Google Scholar]
- Martin-Belmonte, F.; Gassama, A.; Datta, A.; Yu, W.; Rescher, U.; Gerke, V.; Mostov, K. Pten-mediated apical segregation of phosphoinositides controls epithelial morphogenesis through cdc42. Cell 2007, 128, 383–397. [Google Scholar]
- Rodriguez-Fraticelli, A.E.; Vergarajauregui, S.; Eastburn, D.J.; Datta, A.; Alonso, M.A.; Mostov, K.; Martin-Belmonte, F. The cdc42 gef intersectin 2 controls mitotic spindle orientation to form the lumen during epithelial morphogenesis. J. Cell Biol 2010, 189, 725–738. [Google Scholar]
- Tong, X.K.; Hussain, N.K.; Adams, A.G.; O’Bryan, J.P.; McPherson, P.S. Intersectin can regulate the ras/map kinase pathway independent of its role in endocytosis. J. Biol. Chem 2000, 275, 29894–29899. [Google Scholar]
- Mohney, R.P.; Das, M.; Bivona, T.G.; Hanes, R.; Adams, A.G.; Philips, M.R.; O’Bryan, J.P. Intersectin activates ras but stimulates transcription through an independent pathway involving jnk. J. Biol. Chem 2003, 278, 47038–47045. [Google Scholar]
- Adams, A.; Thorn, J.M.; Yamabhai, M.; Kay, B.K.; O’Bryan, J.P. Intersectin, an adaptor protein involved in clathrin-mediated endocytosis, activates mitogenic signaling pathways. J. Biol. Chem 2000, 275, 27414–27420. [Google Scholar]
- Das, M.; Scappini, E.; Martin, N.P.; Wong, K.A.; Dunn, S.; Chen, Y.J.; Miller, S.L.; Domin, J.; O’Bryan, J.P. Regulation of neuron survival through an intersectin-phosphoinositide 3′-kinase c2beta-akt pathway. Mol. Cell Biol 2007, 27, 7906–7917. [Google Scholar]
- Wong, K.A.; Russo, A.; Wang, X.; Chen, Y.-J.; Lavie, A.; O’Bryan, J.P. A new dimension to Ras function: A novel role for nucleotide-free Ras in class II phosphatidylinositol 3-kinase beta (PI3KC2β) regulation. PLoS One 2012, 7, e45360. [Google Scholar]
- Okur, M.N.; Zhu, J.O.; Fong, C.W.; Martinez, N.; Garcia-Dominguez, C.; Rojas, J.M.; Guy, G.; O’Bryan, J.P. Intersectin 1 enhances cbl ubiquitylation of epidermal growth factor receptor through regulation of sprouty2-cbl interaction. Mol. Cell Biol 2012, 32, 817–825. [Google Scholar]
- Nikolaienko, O.; Skrypkina, I.; Tsyba, L.; Fedyshyn, Y.; Morderer, D.; Buchman, V.; de la Luna, S.; Drobot, L.; Rynditch, A. Intersectin 1 forms a complex with adaptor protein ruk/cin85 in vivo independently of epidermal growth factor stimulation. Cell. Signal 2009, 21, 753–759. [Google Scholar]
- Frosi, Y.; Anastasi, S.; Ballaro, C.; Varsano, G.; Castellani, L.; Maspero, E.; Polo, S.; Alema, S.; Segatto, O. A two-tiered mechanism of egfr inhibition by ralt/mig6 via kinase suppression and receptor degradation. J. Cell Biol 2010, 189, 557–571. [Google Scholar]
- Anastasi, S.; Sala, G.; Huiping, C.; Caprini, E.; Russo, G.; Iacovelli, S.; Lucini, F.; Ingvarsson, S.; Segatto, O. Loss of ralt/mig-6 expression in erbb2-amplified breast carcinomas enhances erbb-2 oncogenic potency and favors resistance to herceptin. Oncogene 2005, 24, 4540–4548. [Google Scholar]
- Xu, D.; Makkinje, A.; Kyriakis, J.M. Gene 33 is an endogenous inhibitor of epidermal growth factor (egf) receptor signaling and mediates dexamethasone-induced suppression of egf function. J. Biol. Chem 2005, 280, 2924–2933. [Google Scholar]
- Anastasi, S.; Baietti, M.F.; Frosi, Y.; Alema, S.; Segatto, O. The evolutionarily conserved ebr module of ralt/mig6 mediates suppression of the egfr catalytic activity. Oncogene 2007, 26, 7833–7846. [Google Scholar]
- Lim, C.S.; Seet, B.T.; Ingham, R.J.; Gish, G.; Matskova, L.; Winberg, G.; Ernberg, I.; Pawson, T. The k15 protein of kaposi’s sarcoma-associated herpesvirus recruits the endocytic regulator intersectin 2 through a selective sh3 domain interaction. Biochemistry 2007, 46, 9874–9885. [Google Scholar]
- Dergai, O.; Dergai, M.; Skrypkina, I.; Matskova, L.; Tsyba, L.; Gudkova, D.; Rynditch, A. The lmp2a protein of epstein-barr virus regulates phosphorylation of itsn1 and shb adaptors by tyrosine kinases. Cell. Signal 2013, 25, 33–40. [Google Scholar]
- Nishimura, T.; Yamaguchi, T.; Tokunaga, A.; Hara, A.; Hamaguchi, T.; Kato, K.; Iwamatsu, A.; Okano, H.; Kaibuchi, K. Role of numb in dendritic spine development with a cdc42 gef intersectin and ephb2. Mol. Biol. Cell 2006, 17, 1273–1285. [Google Scholar]
- Irie, F.; Yamaguchi, Y. Ephb receptors regulate dendritic spine development via intersectin, cdc42 and n-wasp. Nat. Neurosci 2002, 5, 1117–1118. [Google Scholar]
- Chabu, C.; Doe, C.Q. Dap160/intersectin binds and activates apkc to regulate cell polarity and cell cycle progression. Development 2008, 135, 2739–2746. [Google Scholar]
- Morfini, G.A.; Burns, M.; Binder, L.I.; Kanaan, N.M.; LaPointe, N.; Bosco, D.A.; Brown, R.H., Jr; Brown, H.; Tiwari, A.; Hayward, L.; et al. Axonal transport defects in neurodegenerative diseases. J. Neurosci. 2009, 29, 12776–12786. [Google Scholar]
- Van der Crabben, S.; van Binsbergen, E.; Ausems, M.; Poot, M.; Bierings, M.; Buijs, A. Constitutional runx1 deletion presenting as non-syndromic thrombocytopenia with myelodysplasia: 21q22 itsn1 as a candidate gene in mental retardation. Leuk. Res 2010, 34, e8–e12. [Google Scholar]
- Lindstrand, A.; Malmgren, H.; Sahlen, S.; Schoumans, J.; Nordgren, A.; Ergander, U.; Holm, E.; Anderlid, B.M.; Blennow, E. Detailed molecular and clinical characterization of three patients with 21q deletions. Clin. Genet 2010, 77, 145–154. [Google Scholar]
- Yao, G.; Chen, X.N.; Flores-Sarnat, L.; Barlow, G.M.; Palka, G.; Moeschler, J.B.; McGillivray, B.; Morse, R.P.; Korenberg, J.R. Deletion of chromosome 21 disturbs human brain morphogenesis. Genet. Med 2006, 8, 1–7. [Google Scholar]
- Canfield, M.A.; Honein, M.A.; Yuskiv, N.; Xing, J.; Mai, C.T.; Collins, J.S.; Devine, O.; Petrini, J.; Ramadhani, T.A.; Hobbs, C.A.; et al. National estimates and race/ethnic-specific variation of selected birth defects in the united states, 1999–2001. Birth Defects Res. A Clin. Mol. Teratol 2006, 76, 747–756. [Google Scholar]
- Zigman, W.B.; Lott, I.T. Alzheimer’s disease in down syndrome: Neurobiology and risk. Ment. Retard Dev. Disabil. Res. Rev 2007, 13, 237–246. [Google Scholar]
- Pucharcos, C.; Fuentes, J.J.; Casas, C.; de la Luna, S.; Alcantara, S.; Arbones, M.L.; Soriano, E.; Estivill, X.; Pritchard, M. Alu-splice cloning of human intersectin (itsn), a putative multivalent binding protein expressed in proliferating and differentiating neurons and overexpressed in down syndrome. Eur. J. Hum. Genet 1999, 7, 704–712. [Google Scholar]
- Hunter, M.P.; Nelson, M.; Kurzer, M.; Wang, X.; Kryscio, R.J.; Head, E.; Pinna, G.; O’Bryan, J.P. Intersectin 1 contributes to phenotypes in vivo: Implications for down’s syndrome. Neuroreport 2011, 22, 767–772. [Google Scholar]
- Wilmot, B.; McWeeney, S.K.; Nixon, R.R.; Montine, T.J.; Laut, J.; Harrington, C.A.; Kaye, J.A.; Kramer, P.L. Translational gene mapping of cognitive decline. Neurobiol. Aging 2008, 29, 524–541. [Google Scholar]
- Dunckley, T.; Beach, T.G.; Ramsey, K.E.; Grover, A.; Mastroeni, D.; Walker, D.G.; LaFleur, B.J.; Coon, K.D.; Brown, K.M.; Caselli, R.; et al. Gene expression correlates of neurofibrillary tangles in alzheimer’s disease. Neurobiol. Aging 2006, 27, 1359–1371. [Google Scholar]
- Blalock, E.M.; Geddes, J.W.; Chen, K.C.; Porter, N.M.; Markesbery, W.R.; Landfield, P.W. Incipient alzheimer’s disease: Microarray correlation analyses reveal major transcriptional and tumor suppressor responses. Proc. Natl. Acad. Sci. USA 2004, 101, 2173–2178. [Google Scholar]
- Cataldo, A.M.; Peterhoff, C.M.; Troncoso, J.C.; Gomez-Isla, T.; Hyman, B.T.; Nixon, R.A. Endocytic pathway abnormalities precede amyloid beta deposition in sporadic alzheimer’s disease and down syndrome: Differential effects of apoe genotype and presenilin mutations. Am. J. Pathol 2000, 157, 277–286. [Google Scholar]
- Keating, D.J.; Dubach, D.; Zanin, M.P.; Yu, Y.; Martin, K.; Zhao, Y.F.; Chen, C.; Porta, S.; Arbones, M.L.; Mittaz, L.; et al. Dscr1/rcan1 regulates vesicle exocytosis and fusion pore kinetics: Implications for down syndrome and alzheimer’s disease. Hum. Mol. Genet 2008, 17, 1020–1030. [Google Scholar]
- Guipponi, M.; Scott, H.S.; Hattori, M.; Ishii, K.; Sakaki, Y.; Antonarakis, S.E. Genomic structure, sequence, and refined mapping of the human intersectin gene (itsn), which encompasses 250 kb on chromosome 21q22.1→q22.2. Cytogenet. Cell Genet 1998, 83, 218–220. [Google Scholar]
- Scappini, E.; Koh, T.W.; Martin, N.P.; O’Bryan, J.P. Intersectin enhances huntingtin aggregation and neurodegeneration through activation of c-jun-nh2-terminal kinase. Hum. Mol. Genet 2007, 16, 1862–1871. [Google Scholar]
- Wang, J.B.; Wu, W.J.; Cerione, R.A. Cdc42 and ras cooperate to mediate cellular transformation by intersectin-l. J. Biol. Chem 2005, 280, 22883–22891. [Google Scholar]
- Russo, A.; O’Bryan, J.P. Intersectin 1 is required for neuroblastoma tumorigenesis. Oncogene 2012, 31, 4828–4834. [Google Scholar]
- Ma, Y.; Wang, B.; Li, W.; Liu, X.; Wang, J.; Ding, T.; Zhang, J.; Ying, G.; Fu, L.; Gu, F. Intersectin1-s is involved in migration and invasion of human glioma cells. J. Neurosci. Res 2011, 89, 1079–1090. [Google Scholar]
- Ma, Y.; Wang, B.; Li, W.; Ying, G.; Fu, L.; Niu, R.; Gu, F. Reduction of intersectin1-s induced apoptosis of human glioblastoma cells. Brain Res 2010, 1351, 222–228. [Google Scholar]
- Marras, E.; Concari, P.; Cortellezzi, L.; Dondi, D.; de Eguileor, M.; Perletti, G. Involvement of pi3k in pkcepsilon-mediated oncogenic signal in rat colonic epithelial cells. Int. J. Oncol 2001, 19, 395–399. [Google Scholar]
- Koutros, S.; Schumacher, F.R.; Hayes, R.B.; Ma, J.; Huang, W.Y.; Albanes, D.; Canzian, F.; Chanock, S.J.; Crawford, E.D.; Diver, W.R.; et al. Pooled analysis of phosphatidylinositol 3-kinase pathway variants and risk of prostate cancer. Cancer Res 2010, 70, 2389–2396. [Google Scholar]
- Sato, N.; Fukushima, N.; Maitra, A.; Iacobuzio-Donahue, C.A.; van Heek, N.T.; Cameron, J.L.; Yeo, C.J.; Hruban, R.H.; Goggins, M. Gene expression profiling identifies genes associated with invasive intraductal papillary mucinous neoplasms of the pancreas. Am. J. Pathol 2004, 164, 903–914. [Google Scholar]
- Armstrong, S.A.; Staunton, J.E.; Silverman, L.B.; Pieters, R.; den Boer, M.L.; Minden, M.D.; Sallan, S.E.; Lander, E.S.; Golub, T.R.; Korsmeyer, S.J. Mll translocations specify a distinct gene expression profile that distinguishes a unique leukemia. Nat. Genet 2002, 30, 41–47. [Google Scholar]
- Qian, Z.; Fernald, A.A.; Godley, L.A.; Larson, R.A.; Le Beau, M.M. Expression profiling of cd34+ hematopoietic stem/progenitor cells reveals distinct subtypes of therapy-related acute myeloid leukemia. Proc. Natl. Acad. Sci. USA 2002, 99, 14925–14930. [Google Scholar]
- Boller, D.; Doepfner, K.T.; de Laurentiis, A.; Guerreiro, A.S.; Marinov, M.; Shalaby, T.; Depledge, P.; Robson, A.; Saghir, N.; Hayakawa, M.; et al. Targeting pi3kc2beta impairs proliferation and survival in acute leukemia, brain tumours and neuroendocrine tumours. Anticancer Re 2012, 32, 3015–3027. [Google Scholar]
- Ramaswamy, S.; Tamayo, P.; Rifkin, R.; Mukherjee, S.; Yeang, C.H.; Angelo, M.; Ladd, C.; Reich, M.; Latulippe, E.; Mesirov, J.P.; et al. Multiclass cancer diagnosis using tumor gene expression signatures. Proc. Natl. Acad. Sci. USA 2001, 98, 15149–15154. [Google Scholar]
- Su, A.I.; Welsh, J.B.; Sapinoso, L.M.; Kern, S.G.; Dimitrov, P.; Lapp, H.; Schultz, P.G.; Powell, S.M.; Moskaluk, C.A.; Frierson, H.F., Jr; et al. Molecular classification of human carcinomas by use of gene expression signatures. Cancer Res 2001, 61, 7388–7393. [Google Scholar]
- Karnoub, A.E.; Weinberg, R.A. Ras oncogenes: Split personalities. Nat. Rev. Mol. Cell Biol 2008, 9, 517–531. [Google Scholar]
- Bremner, R.; Balmain, A. Genetic changes in skin tumor progression: Correlation between presence of a mutant ras gene and loss of heterozygosity on mouse chromosome 7. Cell 1990, 61, 407–417. [Google Scholar]
- Modrek, B.; Ge, L.; Pandita, A.; Lin, E.; Mohan, S.; Yue, P.; Guerrero, S.; Lin, W.M.; Pham, T.; Modrusan, Z.; et al. Oncogenic activating mutations are associated with local copy gain. Mol. Cancer Res 2009, 7, 1244–1252. [Google Scholar]
- Spandidos, D.A.; Frame, M.; Wilkie, N.M. Expression of the normal h-ras1 gene can suppress the transformed and tumorigenic phenotypes induced by mutant ras genes. Anticancer Res 1990, 10, 1543–1554. [Google Scholar]
- Spandidos, A.; Wilkie, N.M. The normal human h-ras1 gene can act as an onco-suppressor. Br. J. Cancer Suppl 1988, 9, 67–71. [Google Scholar]
- Feig, L.A.; Cooper, G.M. Inhibition of nih 3t3 cell proliferation by a mutant ras protein with preferential affinity for gdp. Mol. Cell Biol 1988, 8, 3235–3243. [Google Scholar]
- Zhang, Z.; Wang, Y.; Vikis, H.G.; Johnson, L.; Liu, G.; Li, J.; Anderson, M.W.; Sills, R.C.; Hong, H.L.; Devereux, T.R.; et al. Wildtype kras2 can inhibit lung carcinogenesis in mice. Nat. Genet 2001, 29, 25–33. [Google Scholar]
- Diaz, R.; Lue, J.; Mathews, J.; Yoon, A.; Ahn, D.; Garcia-Espana, A.; Leonardi, P.; Vargas, M.P.; Pellicer, A. Inhibition of ras oncogenic activity by ras protooncogenes. Int. J. Cancer 2005, 113, 241–248. [Google Scholar]
- Diaz, R.; Ahn, D.; Lopez-Barcons, L.; Malumbres, M.; Perez de Castro, I.; Lue, J.; Ferrer-Miralles, N.; Mangues, R.; Tsong, J.; Garcia, R.; et al. The n-ras proto-oncogene can suppress the malignant phenotype in the presence or absence of its oncogene. Cancer Res 2002, 62, 4514–4518. [Google Scholar]
- To, M.D.; Perez-Losada, J.; Mao, J.H.; Hsu, J.; Jacks, T.; Balmain, A. A functional switch from lung cancer resistance to susceptibility at the pas1 locus in kras2la2 mice. Nat. Genet 2006, 38, 926–930. [Google Scholar]
- To, M.D.; Wong, C.E.; Karnezis, A.N.; del Rosario, R.; di Lauro, R.; Balmain, A. Kras regulatory elements and exon 4a determine mutation specificity in lung cancer. Nat. Genet 2008, 40, 1240–1244. [Google Scholar]
- To, M.D.; Rosario, R.D.; Westcott, P.M.; Banta, K.L.; Balmain, A. Interactions between wild-type and mutant rasgenes in lung and skin carcinogenesis. Oncogene 2012. [Google Scholar] [CrossRef]
- Gonczy, P. Mechanisms of asymmetric cell division: Flies and worms pave the way. Nat. Rev. Mol. Cell Biol 2008, 9, 355–366. [Google Scholar]
- Neumuller, R.A.; Knoblich, J.A. Dividing cellular asymmetry: Asymmetric cell division and its implications for stem cells and cancer. Genes. Dev 2009, 23, 2675–2699. [Google Scholar]
- Teckchandani, A.; Mulkearns, E.E.; Randolph, T.W.; Toida, N.; Cooper, J.A. The clathrin adaptor dab2 recruits eh domain scaffold proteins to regulate integrin beta1 endocytosis. Mol. Biol. Cell 2012, 23, 2905–2916. [Google Scholar]
- Powelka, A.M.; Sun, J.; Li, J.; Gao, M.; Shaw, L.M.; Sonnenberg, A.; Hsu, V.W. Stimulation-dependent recycling of integrin beta1 regulated by arf6 and rab11. Traffic 2004, 5, 20–36. [Google Scholar]
- Naslavsky, N.; Rahajeng, J.; Sharma, M.; Jovic, M.; Caplan, S. Interactions between ehd proteins and rab11-fip2: A role for ehd3 in early endosomal transport. Mol. Biol. Cell 2006, 17, 163–177. [Google Scholar]
- Lock, J.G.; Stow, J.L. Rab11 in recycling endosomes regulates the sorting and basolateral transport of e-cadherin. Mol. Biol. Cell 2005, 16, 1744–1755. [Google Scholar]
- Palacios, F.; Price, L.; Schweitzer, J.; Collard, J.G.; D’Souza-Schorey, C. An essential role for arf6-regulated membrane traffic in adherens junction turnover and epithelial cell migration. EMBO J 2001, 20, 4973–4986. [Google Scholar]
- Fujita, Y.; Krause, G.; Scheffner, M.; Zechner, D.; Leddy, H.E.; Behrens, J.; Sommer, T.; Birchmeier, W. Hakai, a c-cbl-like protein, ubiquitinates and induces endocytosis of the e-cadherin complex. Nat. Cell Biol 2002, 4, 222–231. [Google Scholar]
- Malacombe, M.; Ceridono, M.; Calco, V.; Chasserot-Golaz, S.; McPherson, P.S.; Bader, M.F.; Gasman, S. Intersectin-1l nucleotide exchange factor regulates secretory granule exocytosis by activating cdc42. EMBO J 2006, 25, 3494–3503. [Google Scholar]
- Ding, X.; Yang, Z.; Zhou, F.; Hu, X.; Zhou, C.; Luo, C.; He, Z.; Liu, Q.; Li, H.; Yan, F.; et al. Human intersectin 2 (itsn2) binds to eps8 protein and enhances its degradation. BMB Rep 2012, 45, 183–188. [Google Scholar]



{kind=link}
{kind=link}
| Species 1 | # genes 2 | ITSN1 | ITSN2 | GEF domain 3 |
|---|---|---|---|---|
| Homo sapiens | 2 | Chr 21 | Chr 2 | Yes |
| Pan troglodytes | 2 | Chr 21 | Chr 2A | Yes |
| Macaca mulatta | 2 | Chr 19 | Chr 2 | Yes |
| Callithrix jacchus | 2 | Chr 21 | Chr 14 | Yes |
| Nomascus leucogenys | 2 | ND 5 | ND | Yes |
| Pongo abelii | 2 | Chr 21 | Chr 2A | Yes |
| Canis lupus familiaris | 2 | Chr 31 | Chr 17 | Yes |
| Felis catus4 | 2 | ND | ND | Yes |
| Bos taurus | 2 | Chr 1 | Chr 11 | Yes |
| Cavia porcellus | 2 | ND | ND | Yes |
| Mus musculus | 2 | Chr 16 | Chr 12 | Yes |
| Rattus norvegicus | 2 | Chr 11 | Chr 6 | Yes |
| Cricetulus griseus | 2 | ND | ND | Yes |
| Ailuropoda melanoleuca | 2 | ND | ND | Yes |
| Equus caballus | 2 | Chr 15 | Chr 26 | Yes |
| Sus scrofa | 2 | Chr 13 | Chr 3 | Yes |
| Sorex araneus | 2 | ND | ND | Yes |
| Tupaia belangeri4 | 2 | ND | ND | Yes |
| Oryctolagus cuniculus | 2 | ND | ND | Yes |
| Loxodonta africana4 | 2 | ND | ND | Yes |
| Monodelphis domestica | 1 | Chr 4 | - | Yes |
| Erinaceus europaeus4 | 1 | ND | ND | ? 6 |
| Echinops telfairi4 | 2 | ND | ND | Yes |
| Anolis carolinensis | 2 | Chr 3 | Chr 1 | Yes |
| Meleagris gallopavo | 2 | Chr1 | Chr 2 | Yes |
| Gallus gallus | 2 | Chr 1 | Chr 3 | Yes |
| Myotis lucifugus4 | 1 | - | ND | Yes |
| Xenopus laevis | 2 | ND | ND | Yes 9 |
| Xenopus tropicalis | 2 | ND | ND | Yes |
| Ornithorhynchus anatinus | 2 | ND | Chr 2 | Yes |
| Oryzias latipes | 2 | Chr 24 | Chr 3 | Yes |
| Oreochromis niloticus | 2 | ND | ND | Yes |
| Danio rerio7 | 3 | Chr 1 | Chr 20/Chr 17 | Yes |
| Takifugu rubripes7 | 3 | ND | ND/ND | Yes |
| Tetraodon nigroviridis7 | 3 | Chr 17 | Chr 5/Chr 14 | Yes |
| Taeniopygia guttata | 2 | Chr 1B | Chr 3 | Yes |
| Gasterosteus aculeatus7 | 3 | Group VI | Group II & Group XVIII | Yes |
| Ciona savignyi | 1 | ND | - | No |
| Ciona intestinalis | 2 | ND | ND | Yes |
| Nasonia vitripennis | 1 | ND | - | Yes |
| Acyrthosiphon pisum | 1 | - | ND | Yes |
| Anopheles gambiae8 | 1 | 3R | - | No |
| Aedes aegypti8 | 1 | ND | No | |
| Drosophila pseudoobscura8 | 1 | ND | - | No |
| Drosophila melanogaster8 | 1 | 2L | - | No |
| Tribolium castaneum4 | 1 | LG3 | - | ? 6 |
| Apis mellifera8 | 1 | - | ND | Yes |
| Caenorhabditis briggsae | 1 | ND | - | No |
| Caenorhabditis elegans | 1 | Chr 4 | - | No |
| Caenorhabditis remanei | 1 | ND | - | No |
| Hydra magnipapillata | 2 | ND | ND | Yes |
| Strongylocentrotus purpuratus | 2 | ND | ND | ? 6 |
| Cryptococcus neoformans | 1 | ND | - | Yes |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Hunter, M.P.; Russo, A.; O'Bryan, J.P. Emerging Roles for Intersectin (ITSN) in Regulating Signaling and Disease Pathways. Int. J. Mol. Sci. 2013, 14, 7829-7852. https://doi.org/10.3390/ijms14047829
Hunter MP, Russo A, O'Bryan JP. Emerging Roles for Intersectin (ITSN) in Regulating Signaling and Disease Pathways. International Journal of Molecular Sciences. 2013; 14(4):7829-7852. https://doi.org/10.3390/ijms14047829
Chicago/Turabian StyleHunter, Michael P., Angela Russo, and John P. O'Bryan. 2013. "Emerging Roles for Intersectin (ITSN) in Regulating Signaling and Disease Pathways" International Journal of Molecular Sciences 14, no. 4: 7829-7852. https://doi.org/10.3390/ijms14047829