Irradiated Human Dermal Fibroblasts Are as Efficient as Mouse Fibroblasts as a Feeder Layer to Improve Human Epidermal Cell Culture Lifespan



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
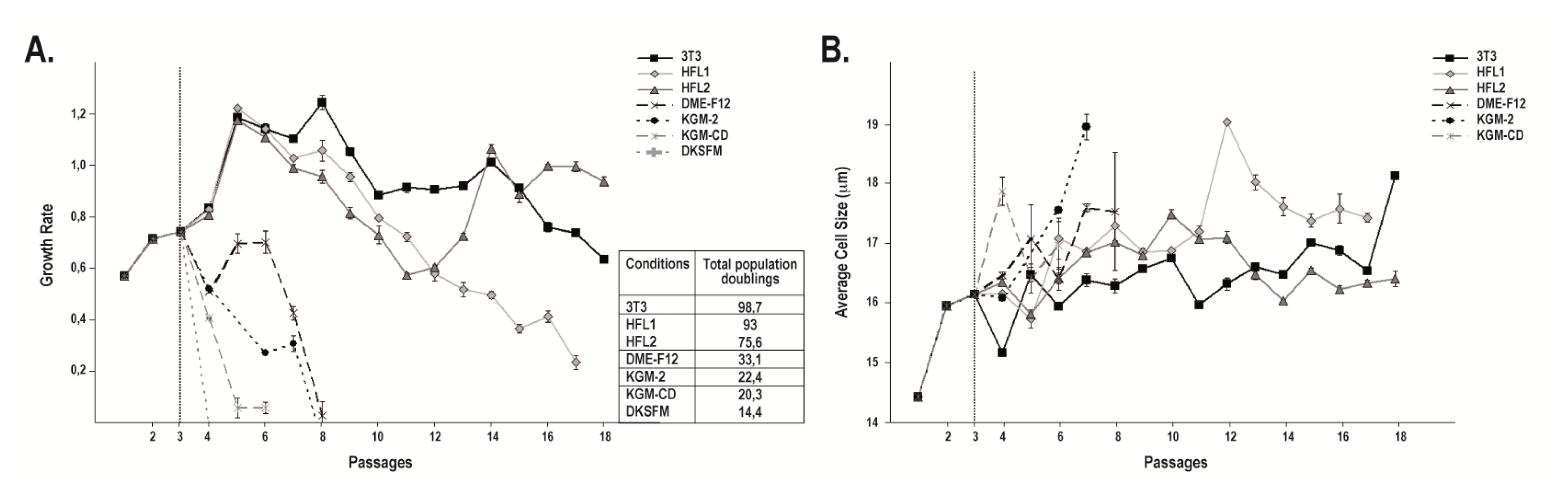
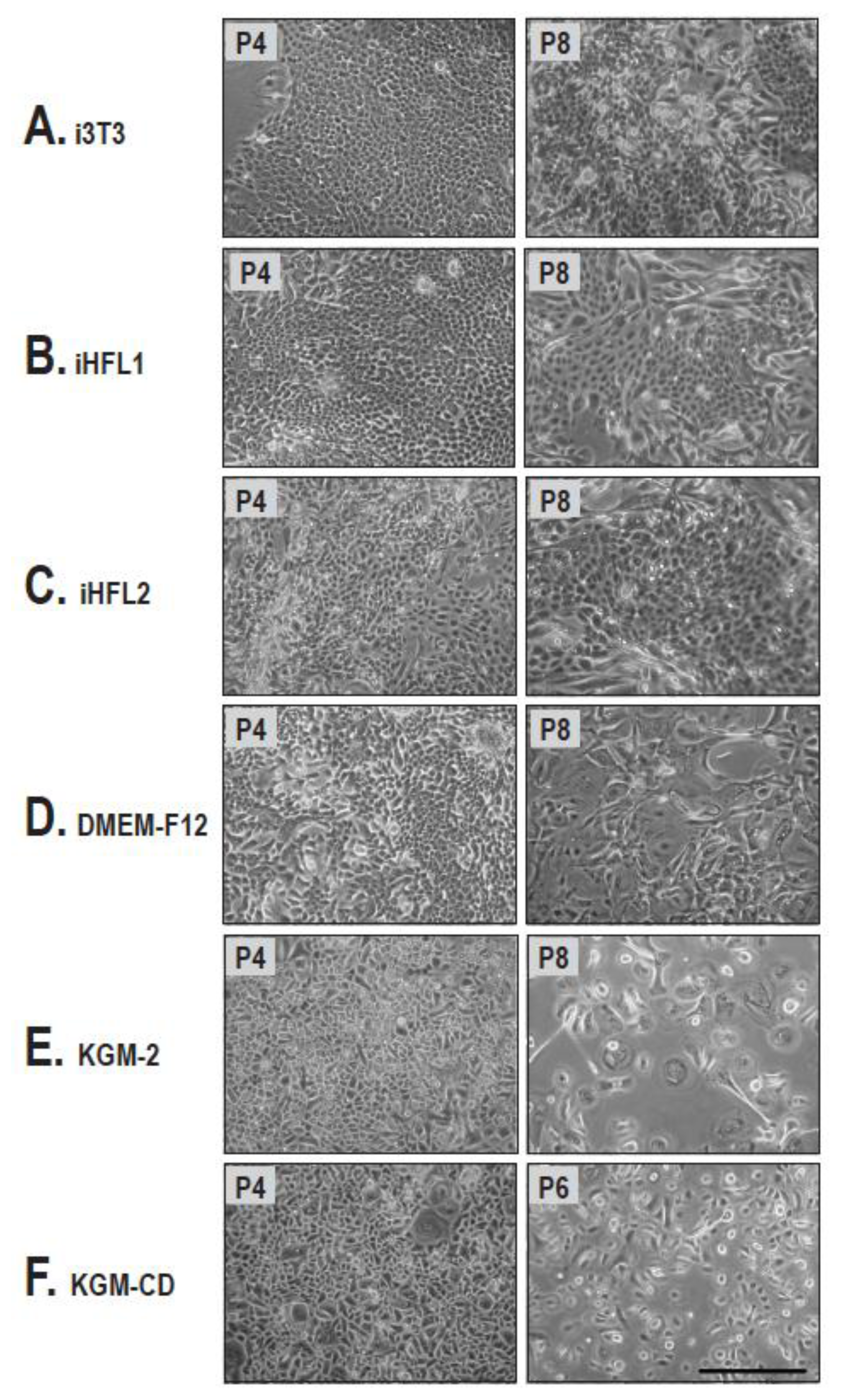
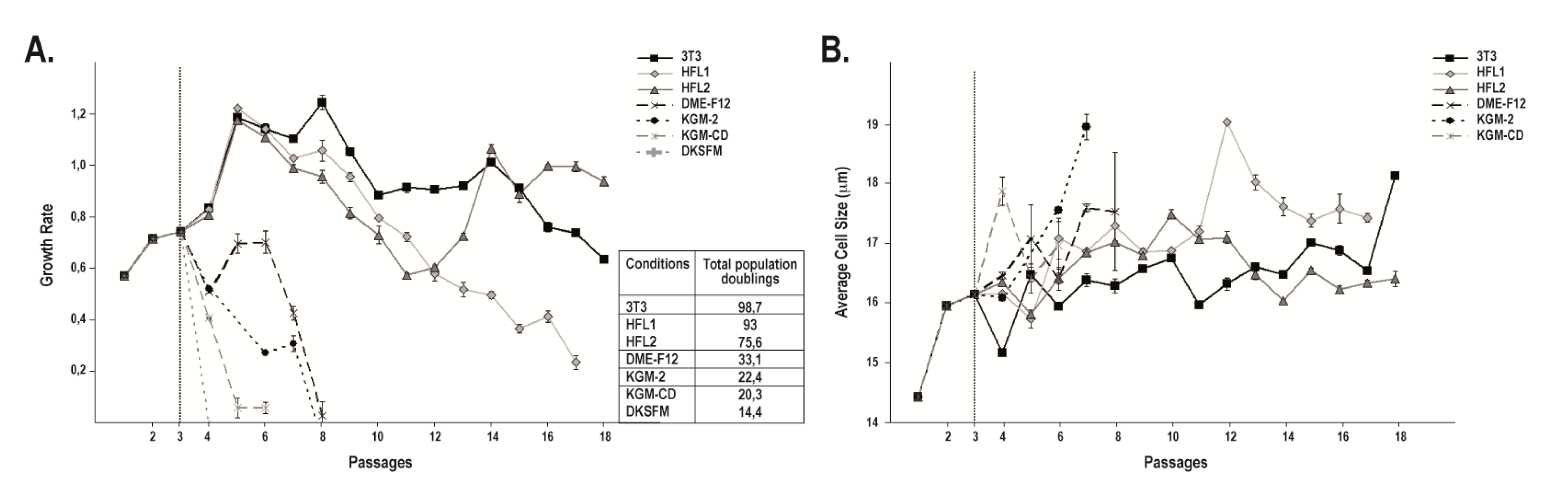
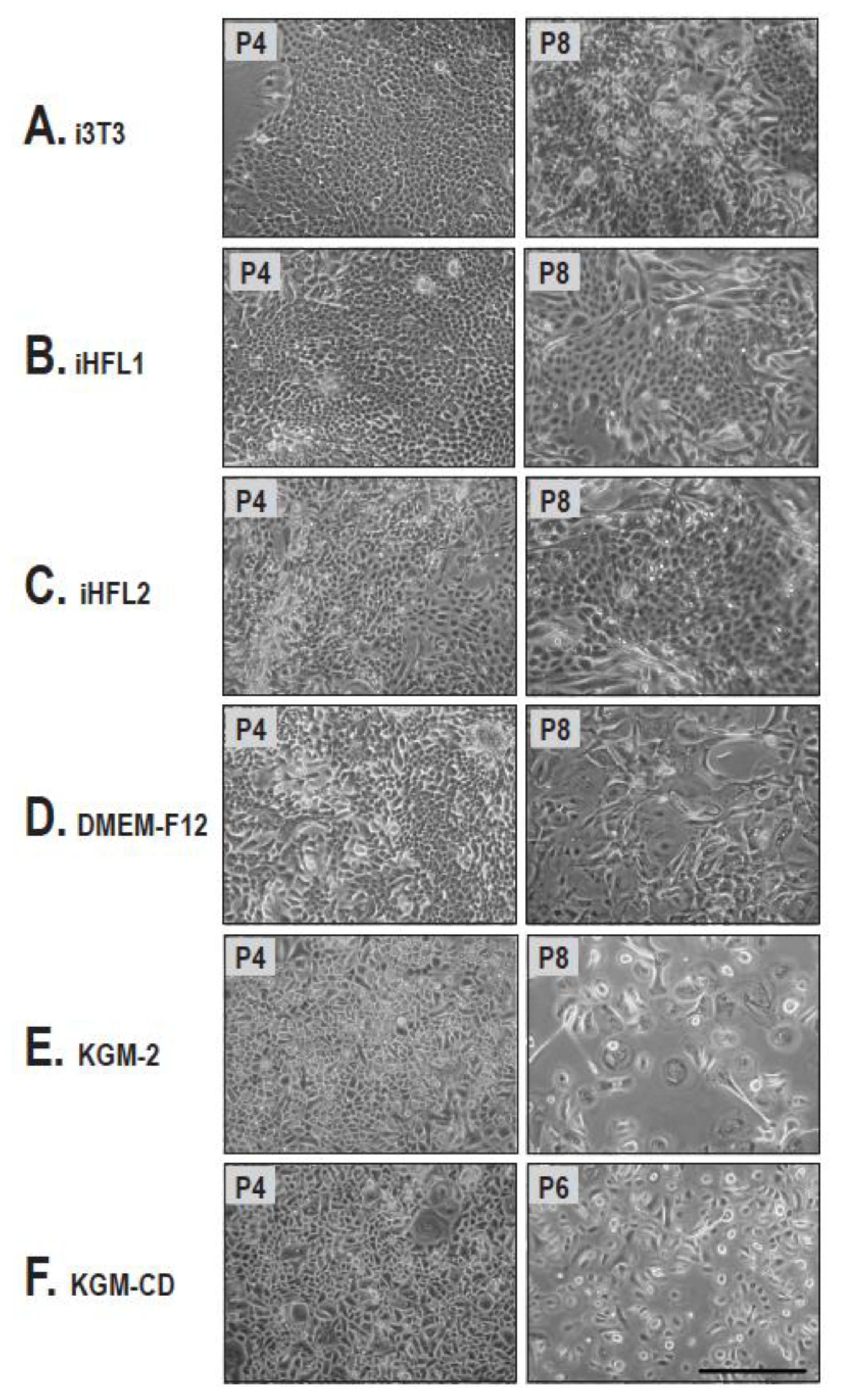
2.1. Cell Morphology and Growth Rate of Keratinocytes Cultured on Human or i3T3 Feeder Layers Are Comparable
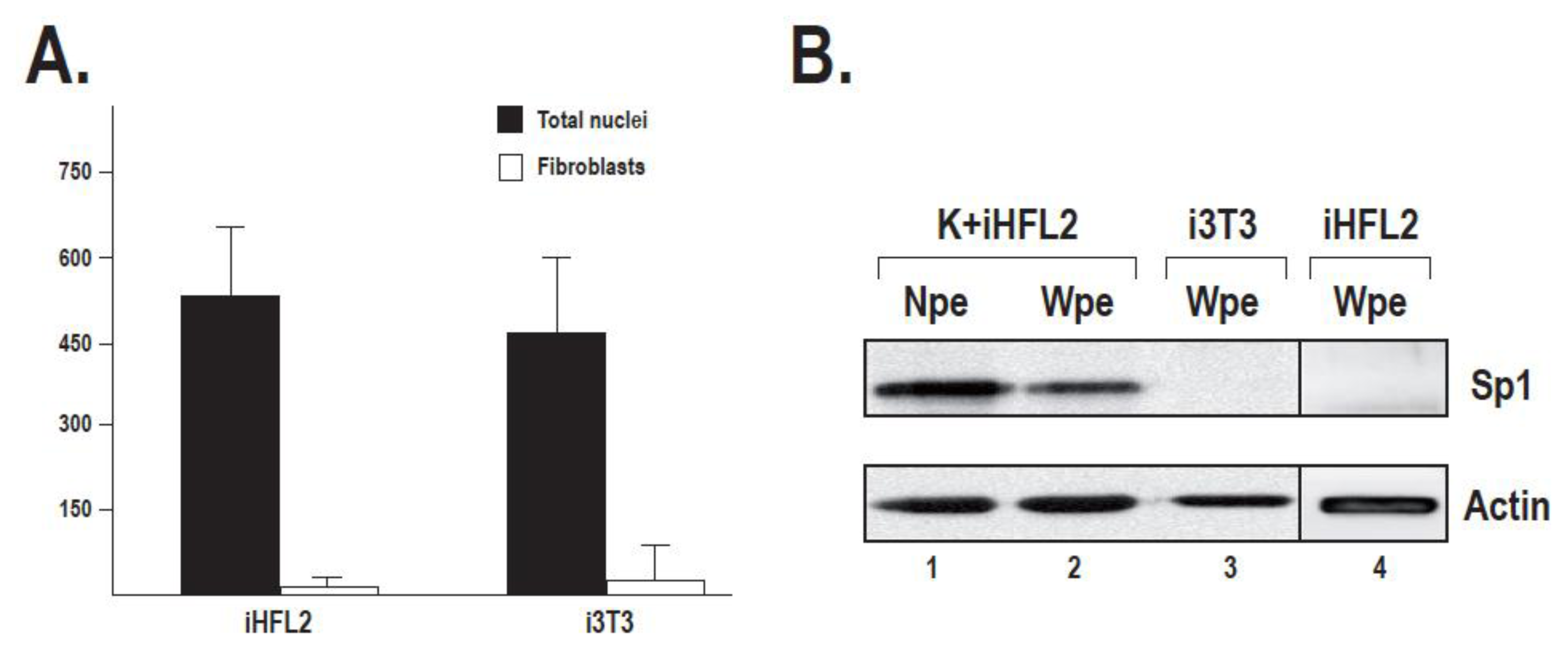
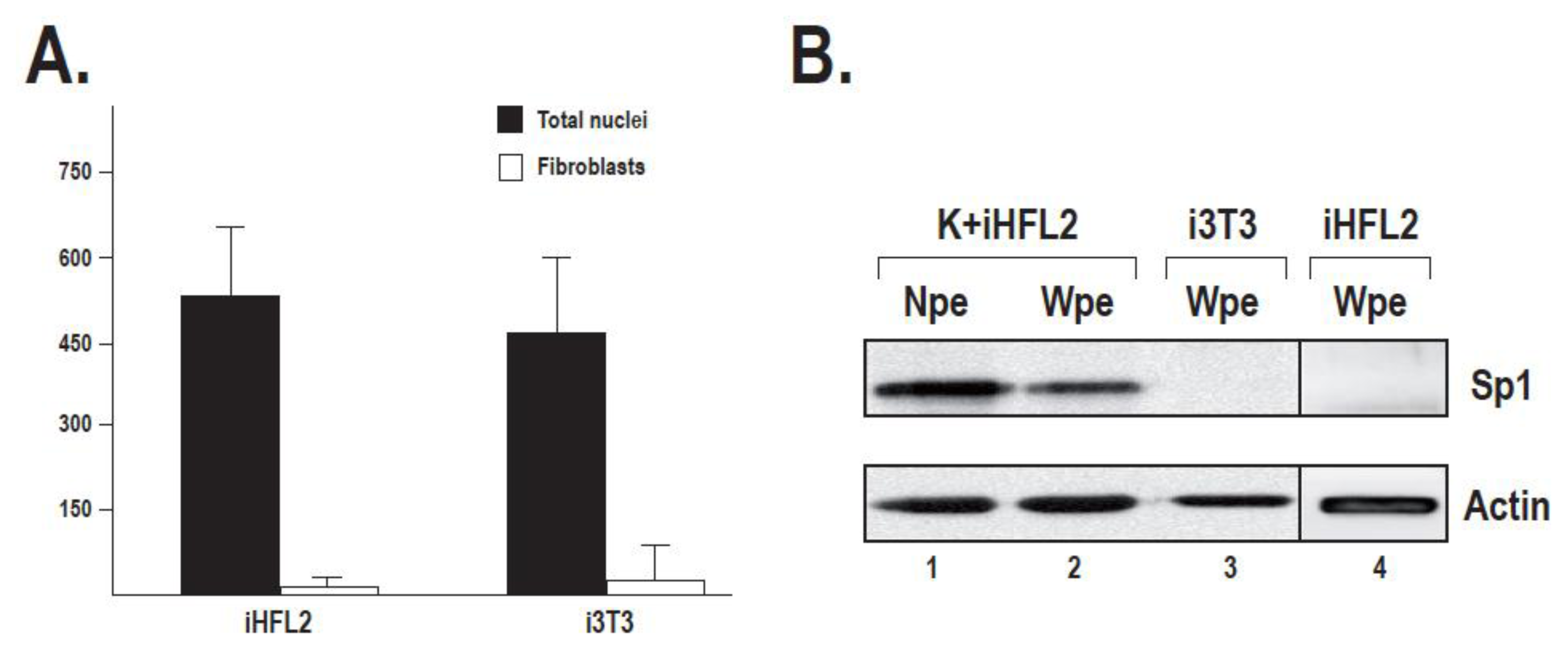
2.2. Sp1 Expression in Fibroblasts Used as Feeder Layers Is Negligible
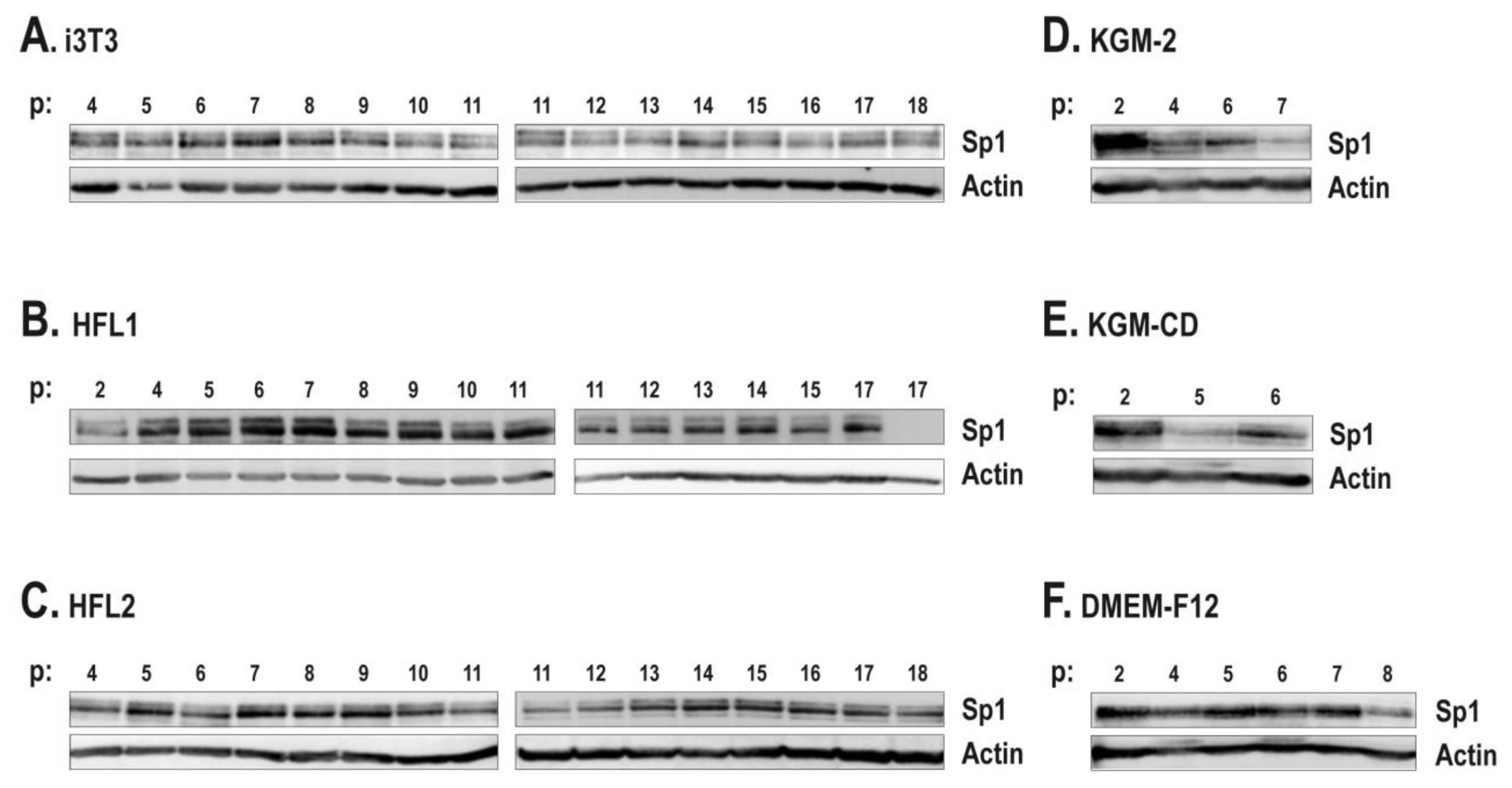
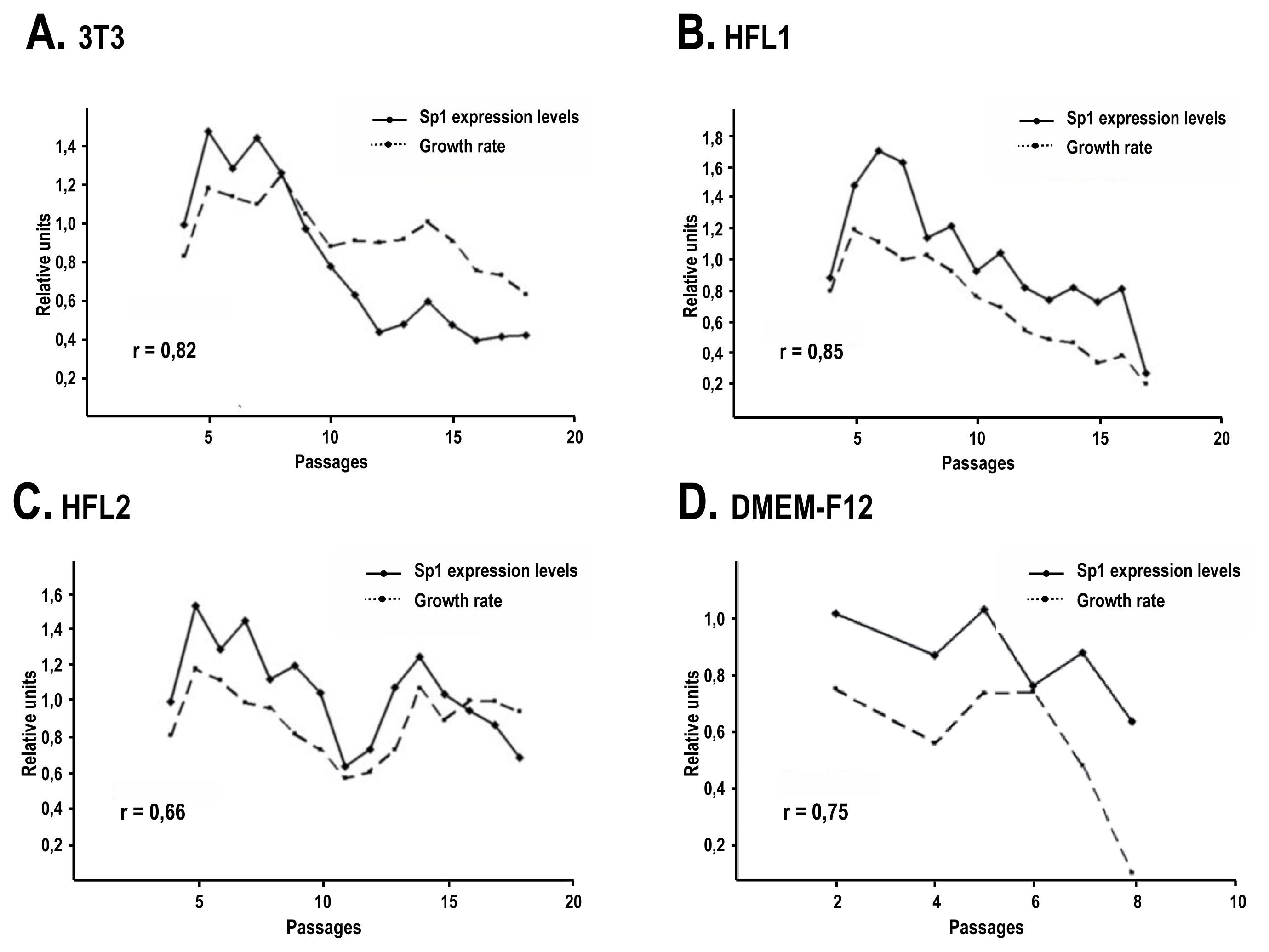
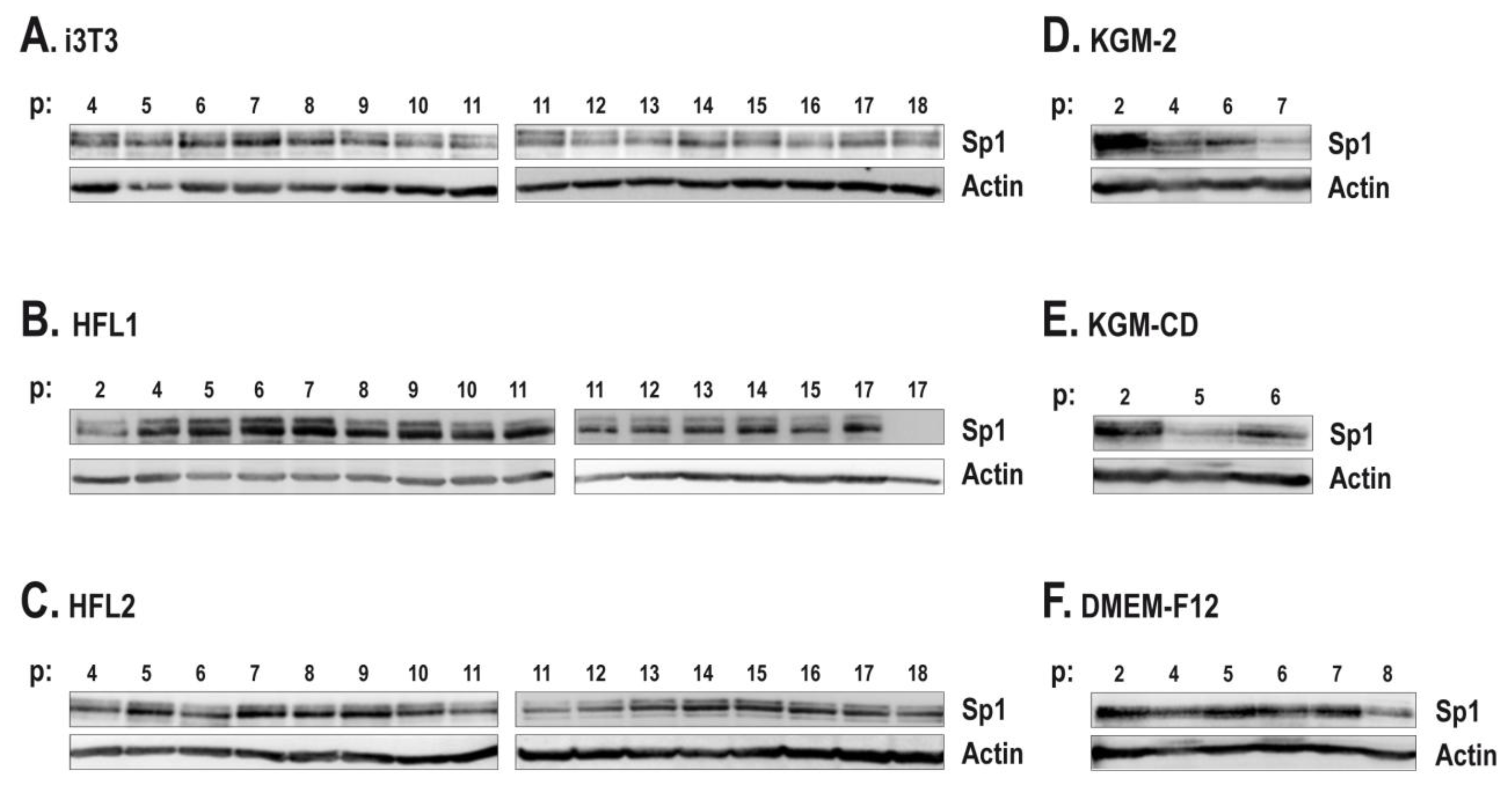
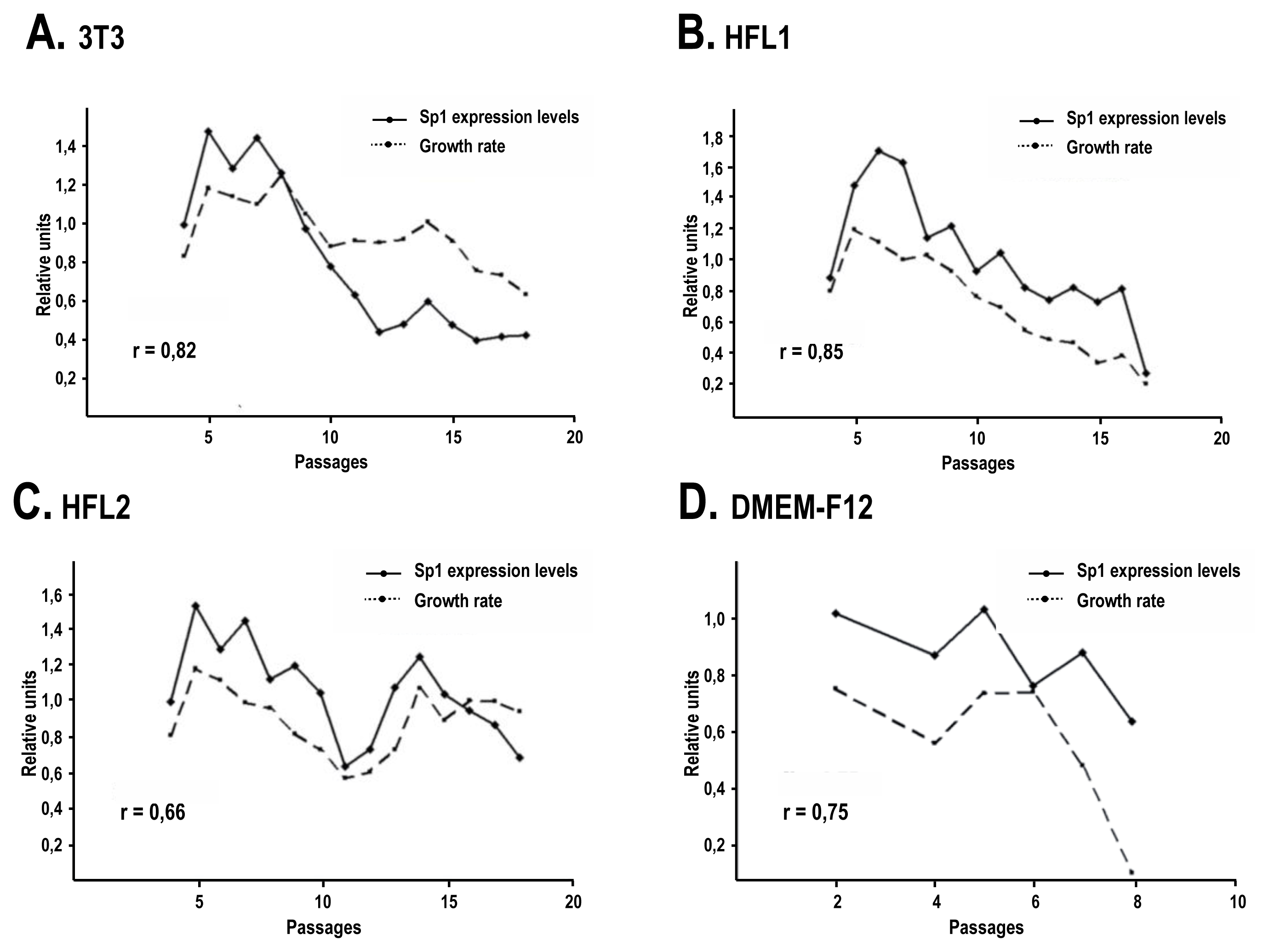
2.3. The Human Feeder Layer Preserves Sp1 Expression in Keratinocytes
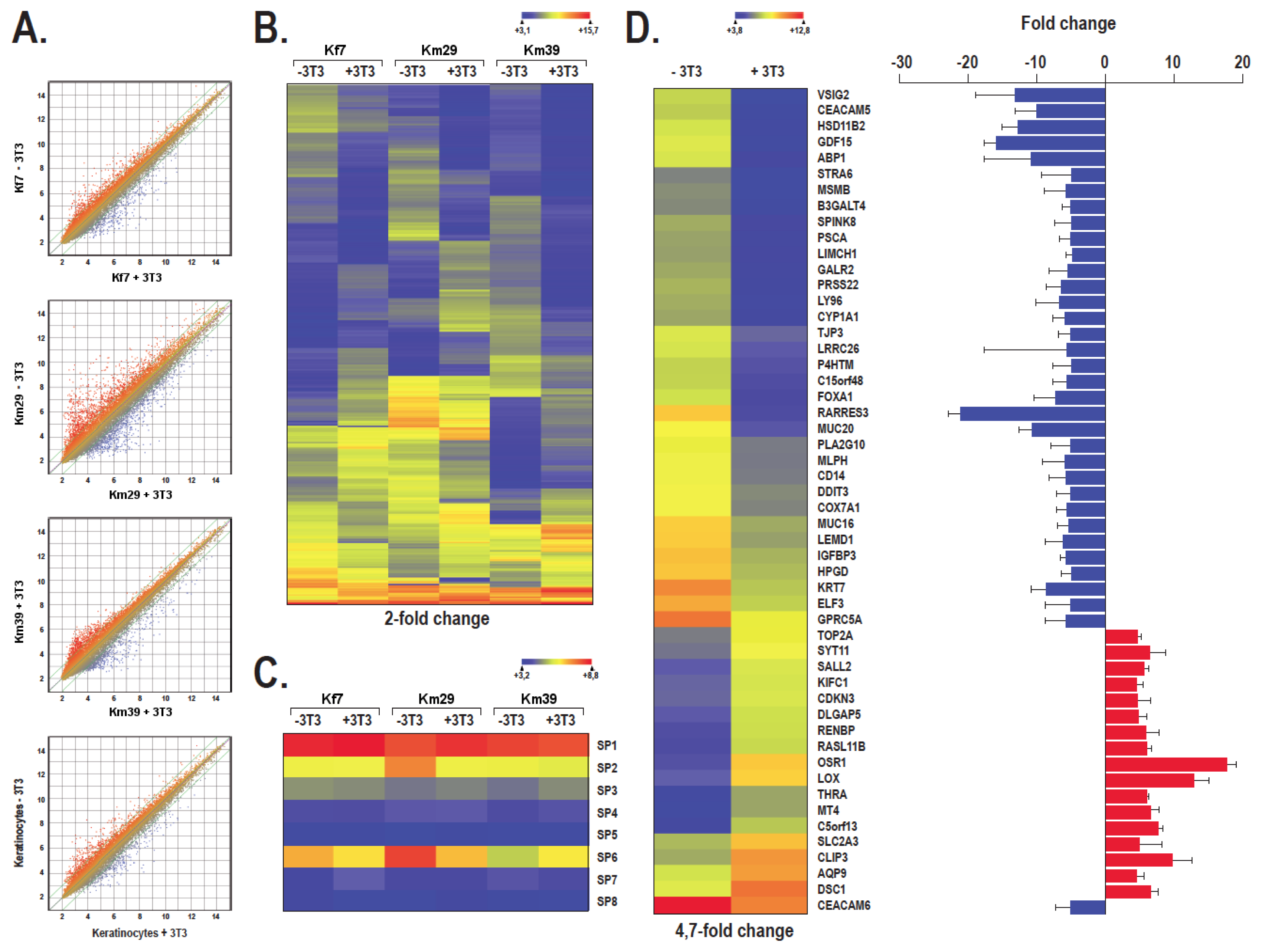
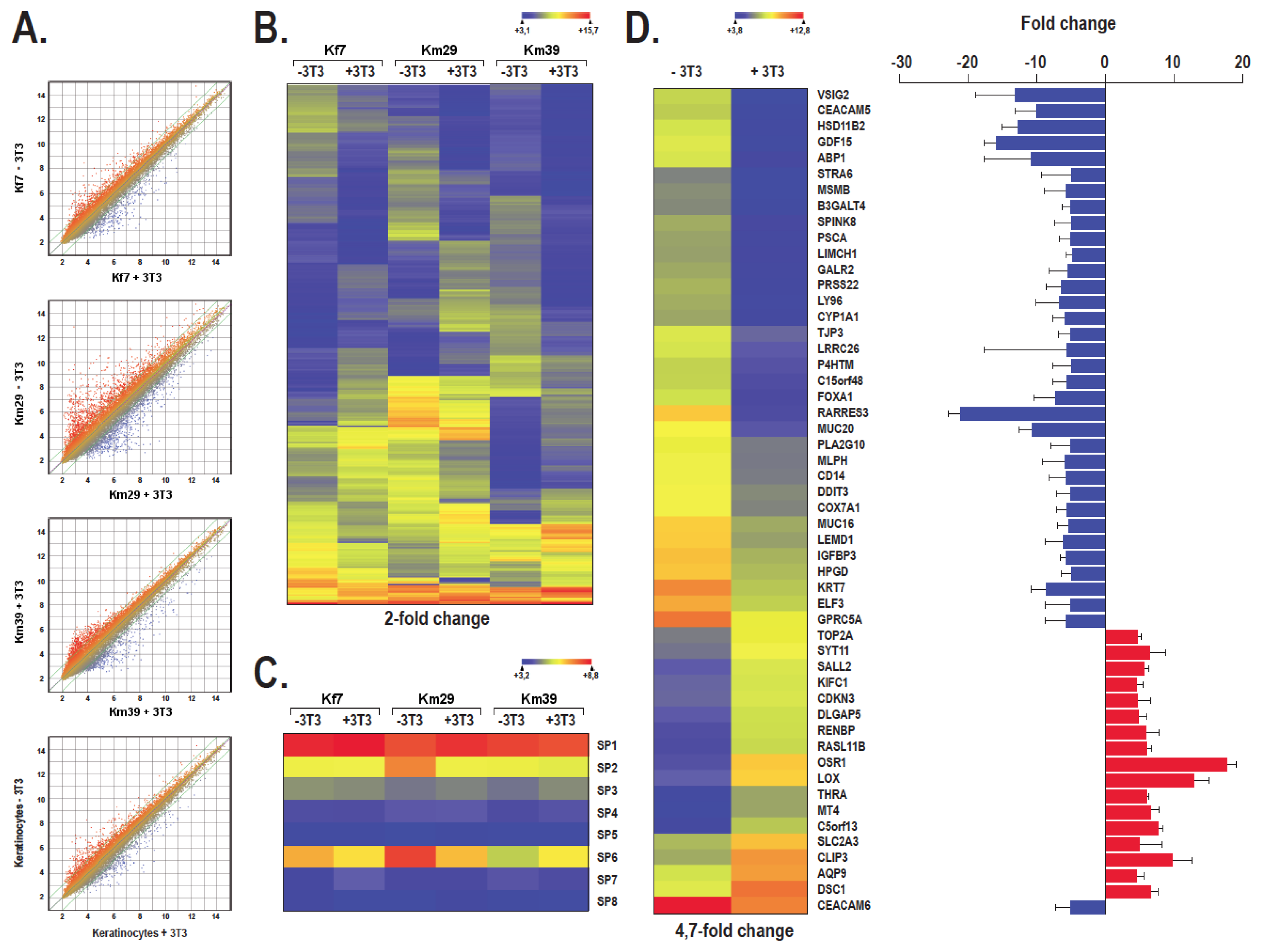
2.4. The Pattern of Genes Expressed by Human Skin Keratinocytes Differs in the Absence or Presence of a Feeder Layer
3. Discussion
4. Experimental Section
4.1. Cell Culture
4.2. Immunofluorescence Analysis
4.3. Western Blot
4.4. Gene Expression Profiling
5. Conclusions
Supplementary Information
ijms-14-04684-s001.docxAcknowledgments
Conflict of Interest
References
- Germain, L.; Goulet, F.; Moulin, V.; Berthod, F.; Auger, F.A. Engineering human tissues for in vivo applications. Ann. N. Y. Acad. Sci 2002, 961, 268–270. [Google Scholar]
- Paquet, C.; Larouche, D.; Bisson, F.; Proulx, S.; Simard-Bisson, C.; Gaudreault, M.; Robitaille, H.; Carrier, P.; Martel, I.; Duranceau, L.; et al. Tissue engineering of skin and cornea: Development of new models for in vitro studies. Ann. N. Y. Acad. Sci 2010, 1197, 166–177. [Google Scholar]
- Supp, D.M.; Boyce, S.T. Engineered skin substitutes: Practices and potentials. Clin. Dermatol 2005, 23, 403–412. [Google Scholar]
- Fusenig, N.E.; Worst, P.K. Mouse epidermal cell cultures. I. Isolation and cultivation of epidermal cells from adult mouse skin. J. Invest. Dermatol 1974, 63, 187–193. [Google Scholar]
- Rheinwald, J.G.; Green, H. Serial cultivation of strains of human epidermal keratinocytes: The formation of keratinizing colonies from single cells. Cell 1975, 6, 331–343. [Google Scholar]
- Gallico, G.G., III; O’Connor, N.E.; Compton, C.C.; Kehinde, O.; Green, H. Permanent coverage of large burn wounds with autologous cultured human epithelium. N. Engl. J. Med. 1984, 311, 448–451. [Google Scholar]
- O’Connor, N.E.; Mulliken, J.B.; Banks-Schlegel, S.; Kehinde, O.; Green, H. Grafting of bums with cultured epithelium prepared from autologous epidermal cells. Lancet 1981, 1, 75–78. [Google Scholar]
- Martin, M.J.; Muotri, A.; Gage, F.; Varki, A. Human embryonic stem cells express an immunogenic nonhuman sialic acid. Nat. Med 2005, 11, 228–232. [Google Scholar]
- Auxenfans, C.; Thepot, A.; Justin, V.; Hautefeuille, A.; Shahabeddin, L.; Damour, O.; Hainaut, P. Characterisation of human fibroblasts as keratinocyte feeder layer using p63 isoforms status. Biomed. Mater. Eng 2009, 19, 365–372. [Google Scholar]
- Bullock, A.J.; Higham, M.C.; MacNeil, S. Use of human fibroblasts in the development of a xenobiotic-free culture and delivery system for human keratinocytes. Tissue Eng 2006, 12, 245–255. [Google Scholar]
- Jubin, K.; Martin, Y.; Lawrence-Watt, D.J.; Sharpe, J.R. A fully autologous co-culture system utilising non-irradiated autologous fibroblasts to support the expansion of human keratinocytes for clinical use. Cytotechnology 2011, 63, 655–662. [Google Scholar]
- Sun, T.; Higham, M.; Layton, C.; Haycock, J.; Short, R.; MacNeil, S. Developments in xenobiotic-free culture of human keratinocytes for clinical use. Wound Repair Regen 2004, 12, 626–634. [Google Scholar]
- Mujaj, S.; Manton, K.; Upton, Z.; Richards, S. Serum-free primary human fibroblast and keratinocyte coculture. Tissue Eng 2010, 16, 1407–1420. [Google Scholar]
- Pillai, S.; Bikle, D.D.; Mancianti, M.L.; Cline, P.; Hincenbergs, M. Calcium regulation of growth and differentiation of normal human keratinocytes: Modulation of differentiation competence by stages of growth and extracellular calcium. J. Cell. Physiol 1990, 143, 294–302. [Google Scholar]
- Scuderi, N.; Anniboletti, T.; Carlesimo, B.; Onesti, M.G. Clinical application of autologous three-cellular cultured skin substitutes based on esterified hyaluronic acid scaffold: Our experience. In Vivo 2009, 23, 991–1003. [Google Scholar]
- Duval, C.; Gaudreault, M.; Vigneault, F.; Touzel-Deschênes, L.; Rochette, P.; Masson-Gadais, B.; Germain, L.; Guerin, S.L. Rescue of the transcription factors Sp1 and NFI in human skin keratinocytes through a feeder-layer-dependent suppression of the proteasome activity. J. Mol. Biol. 2012. in revision. [Google Scholar]
- Masson-Gadais, B.; Fugere, C.; Paquet, C.; Leclerc, S.; Lefort, N.R.; Germain, L.; Guerin, S.L. The feeder layer-mediated extended lifetime of cultured human skin keratinocytes is associated with altered levels of the transcription factors Sp1 and Sp3. J. Cell. Physiol 2006, 206, 831–842. [Google Scholar]
- Wierstra, I. Sp1: Emerging roles—Beyond constitutive activation of TATA-less housekeeping genes. Biochem. Biophys. Res. Commun 2008, 372, 1–13. [Google Scholar]
- Li, L.; Davie, J.R. The role of Sp1 and Sp3 in normal and cancer cell biology. Ann. Anat 2010, 192, 275–283. [Google Scholar]
- Gaudreault, M.; Carrier, P.; Larouche, K.; Leclerc, S.; Giasson, M.; Germain, L.; Guerin, S.L. Influence of sp1/sp3 expression on corneal epithelial cells proliferation and differentiation properties in reconstructed tissues. Invest. Ophthalmol. Vis. Sci 2003, 44, 1447–1457. [Google Scholar]
- Oie, Y.; Hayashi, R.; Takagi, R.; Yamato, M.; Takayanagi, H.; Tano, Y.; Nishida, K. A novel method of culturing human oral mucosal epithelial cell sheet using post-mitotic human dermal fibroblast feeder cells and modified keratinocyte culture medium for ocular surface reconstruction. Br. J. Ophthalmol 2010, 94, 1244–1250. [Google Scholar]
- Limat, A.; Hunziker, T.; Boillat, C.; Bayreuther, K.; Noser, F. Post-mitotic human dermal fibroblasts efficiently support the growth of human follicular keratinocytes. J. Invest. Dermatol 1989, 92, 758–762. [Google Scholar]
- Sun, T.T.; Green, H. Differentiation of the epidermal keratinocyte in cell culture: Formation of the cornified envelope. Cell 1976, 9, 511–521. [Google Scholar]
- Rowden, G. Ultrastructural studies of keratinized epithelia of the mouse. III. Determination of the volumes of nuclei and cytoplasm of cells in murine epidermis. J. Invest. Dermatol 1975, 64, 1–3. [Google Scholar]
- Barrandon, Y.; Green, H. Three clonal types of keratinocyte with different capacities for multiplication. Proc. Natl. Acad. Sci. USA 1987, 84, 2302–2306. [Google Scholar]
- Hennings, H.; Michael, D.; Cheng, C.; Steinert, P.; Holbrook, K.; Yuspa, S.H. Calcium regulation of growth and differentiation of mouse epidermal cells in culture. Cell 1980, 19, 245–254. [Google Scholar]
- Black, A.F.; Bouez, C.; Perrier, E.; Schlotmann, K.; Chapuis, F.; Damour, O. Optimization and characterization of an engineered human skin equivalent. Tissue Eng 2005, 11, 723–733. [Google Scholar]
- Germain, L.; Rouabhia, M.; Guignard, R.; Carrier, L.; Bouvard, V.; Auger, F.A. Improvement of human keratinocyte isolation and culture using thermolysin. Burns 1993, 19, 99–104. [Google Scholar]
- Green, H.; Kehinde, O.; Thomas, J. Growth of cultured human epidermal cells into multiple epithelia suitable for grafting. Proc. Natl. Acad. Sci. USA 1979, 76, 5665–5668. [Google Scholar]
- Thepot, A.; Desanlis, A.; Venet, E.; Thivillier, L.; Justin, V.; Morel, A.P.; Defraipont, F.; Till, M.; Krutovskikh, V.; Tommasino, M.; et al. Assessment of transformed properties in vitro and of tumorigenicity in vivo in primary keratinocytes cultured for epidermal sheet transplantation. J. Skin Cancer 2011, 2011, 936546. [Google Scholar]
- Pellegrini, G.; Golisano, O.; Paterna, P.; Lambiase, A.; Bonini, S.; Rama, P.; De Luca, M. Location and clonal analysis of stem cells and their differentiated progeny in the human ocular surface. J. Cell. Biol 1999, 145, 769–782. [Google Scholar]
- Sharma, S.M.; Fuchsluger, T.; Ahmad, S.; Katikireddy, K.R.; Armant, M.; Dana, R.; Jurkunas, U.V. Comparative analysis of human-derived feeder layers with 3T3 fibroblasts for the ex vivo expansion of human limbal and oral epithelium. Stem Cell Rev 2011, 8, 696–705. [Google Scholar]
- Goulet, F.; Poitras, A.; Rouabhia, M.; Cusson, D.; Germain, L.; Auger, F.A. Stimulation of human keratinocyte proliferation through growth factor exchanges with dermal fibroblasts in vitro. Burns 1996, 22, 107–112. [Google Scholar]
- Jean, J.; Lapointe, M.; Soucy, J.; Pouliot, R. Development of an in vitro psoriatic skin model by tissue engineering. J. Dermatol. Sci 2009, 53, 19–25. [Google Scholar]
- Rao, J.; Zhang, F.; Donnelly, R.J.; Spector, N.L.; Studzinski, G.P. Truncation of Sp1 transcription factor by myeloblastin in undifferentiated HL60 cells. J. Cell. Phys 1998, 175, 121–128. [Google Scholar]
- Ye, X.; Liu, S.F. Lipopolysaccharide causes Sp1 protein degradation by inducing a unique trypsin-like serine protease in rat lungs. Biochim. Biophys. Acta 2007, 1773, 243–253. [Google Scholar]
- Yasuda, S.; Morokawa, N.; Wong, G.W.; Rossi, A.; Madhusudhan, M.S.; Sali, A.; Askew, Y.S.; Adachi, R.; Silverman, G.A.; Krilis, S.A.; et al. Urokinase-type plasminogen activator is a preferred substrate of the human epithelium serine protease tryptase epsilon/PRSS22. Blood 2005, 105, 3893–3901. [Google Scholar]
- Wong, G.W.; Yasuda, S.; Madhusudhan, M.S.; Li, L.; Yang, Y.; Krilis, S.A.; Sali, A.; Stevens, R.L. Human tryptase epsilon (PRSS22), a new member of the chromosome 16p13.3 family of human serine proteases expressed in airway epithelial cells. J. Biol. Chem 2001, 276, 49169–49182. [Google Scholar]
- Li, W.; Danilenko, D.M.; Bunting, S.; Ganesan, R.; Sa, S.; Ferrando, R.; Wu, T.D.; Kolumam, G.A.; Ouyang, W.; Kirchhofer, D. The serine protease marapsin is expressed in stratified squamous epithelia and is up-regulated in the hyperproliferative epidermis of psoriasis and regenerating wounds. J. Biol. Chem 2009, 284, 218–228. [Google Scholar]
- Lavoie, A.; Fugere, C.; Beauparlant, A.; Goyer, B.; Larouche, D.; Paquet, C.; Desgagne, M.; Sauve, S.; Robitaille, H.; Dunnwald, M.; et al. Human epithelial stem cells persist within tissue-engineered skin produced by the self-assembly approach. Tissue Eng. 2012. [Google Scholar] [CrossRef]
- DiSepio, D.; Ghosn, C.; Eckert, R.L.; Deucher, A.; Robinson, N.; Duvic, M.; Chandraratna, R.A.; Nagpal, S. Identification and characterization of a retinoid-induced class II tumor suppressor/growth regulatory gene. Proc. Natl. Acad. Sci. USA 1998, 95, 14811–14815. [Google Scholar]
- Jans, R.; Sturniolo, M.T.; Eckert, R.L. Localization of the TIG3 transglutaminase interaction domain and demonstration that the amino-terminal region is required for TIG3 function as a keratinocyte differentiation regulator. J. Invest. Dermatol 2008, 128, 517–529. [Google Scholar]
- Sturniolo, M.T.; Dashti, S.R.; Deucher, A.; Rorke, E.A.; Broome, A.M.; Chandraratna, R.A.; Keepers, T.; Eckert, R.L. A novel tumor suppressor protein promotes keratinocyte terminal differentiation via activation of type I transglutaminase. J. Biol. Chem 2003, 278, 48066–48073. [Google Scholar]
- Acquafreda, T.; Soprano, K.J.; Soprano, D.R. GPRC5A: A potential tumor suppressor and oncogene. Cancer Biol. Ther 2009, 8, 963–965. [Google Scholar]
- Tao, Q.; Fujimoto, J.; Men, T.; Ye, X.; Deng, J.; Lacroix, L.; Clifford, J.L.; Mao, L.; van Pelt, C.S.; Lee, J.J.; Lotan, D.; Lotan, R. Identification of the retinoic acid-inducible Gprc5a as a new lung tumor suppressor gene. J. Natl. Cancer Inst 2007, 99, 1668–1682. [Google Scholar]
- Deng, J.; Fujimoto, J.; Ye, X.F.; Men, T.Y.; van Pelt, C.S.; Chen, Y.L.; Lin, X.F.; Kadara, H.; Tao, Q.; Lotan, D.; et al. Knockout of the tumor suppressor gene Gprc5a in mice leads to NF-kappaB activation in airway epithelium and promotes lung inflammation and tumorigenesis. Cancer Prev. Res. (Phila) 2010, 3, 424–437. [Google Scholar]
- Fuchs, E.; Green, H. Regulation of terminal differentiation of cultured human keratinocytes by vitamin A. Cell 1981, 25, 617–625. [Google Scholar]
- King, I.A.; O'Brien, T.J.; Buxton, R.S. Expression of the “skin-type” desmosomal cadherin DSC1 is closely linked to the keratinization of epithelial tissues during mouse development. J. Invest. Dermatol 1996, 107, 531–538. [Google Scholar]
- Chidgey, M.; Brakebusch, C.; Gustafsson, E.; Cruchley, A.; Hail, C.; Kirk, S.; Merritt, A.; North, A.; Tselepis, C.; Hewitt, J.; et al. Mice lacking desmocollin 1 show epidermal fragility accompanied by barrier defects and abnormal differentiation. J. Cell Biol 2001, 155, 821–832. [Google Scholar]
- Nuber, U.A.; Schafer, S.; Stehr, S.; Rackwitz, H.R.; Franke, W.W. Patterns of desmocollin synthesis in human epithelia: Immunolocalization of desmocollins 1 and 3 in special epithelia and in cultured cells. Eur. J. Cell Biol 1996, 71, 1–13. [Google Scholar]
- Pellegrini, G.; Rama, P.; de Luca, M. Vision from the right stem. Trends Mol. Med 2010, 17, 1–7. [Google Scholar]
- Rochon, M.H.; Fradette, J.; Fortin, V.; Tomasetig, F.; Roberge, C.J.; Baker, K.; Berthod, F.; Auger, F.A.; Germain, L. Normal human epithelial cells regulate the size and morphology of tissue-engineered capillaries. Tissue Eng 2010, 16, 1457–1468. [Google Scholar]
- Larouche, D.; Paquet, C.; Fradette, J.; Carrier, P.; Auger, F.A.; Germain, L. Regeneration of skin and cornea by tissue engineering. Methods Mol. Biol 2009, 482, 233–256. [Google Scholar]
- Roy, R.J.; Gosselin, P.; Guerin, S.L. A short protocol for micro-purification of nuclear proteins from whole animal tissue. Biotechniques 1991, 11, 770–777. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Bisson, F.; Rochefort, É.; Lavoie, A.; Larouche, D.; Zaniolo, K.; Simard-Bisson, C.; Damour, O.; Auger, F.A.; Guérin, S.L.; Germain, L. Irradiated Human Dermal Fibroblasts Are as Efficient as Mouse Fibroblasts as a Feeder Layer to Improve Human Epidermal Cell Culture Lifespan. Int. J. Mol. Sci. 2013, 14, 4684-4704. https://doi.org/10.3390/ijms14034684
Bisson F, Rochefort É, Lavoie A, Larouche D, Zaniolo K, Simard-Bisson C, Damour O, Auger FA, Guérin SL, Germain L. Irradiated Human Dermal Fibroblasts Are as Efficient as Mouse Fibroblasts as a Feeder Layer to Improve Human Epidermal Cell Culture Lifespan. International Journal of Molecular Sciences. 2013; 14(3):4684-4704. https://doi.org/10.3390/ijms14034684
Chicago/Turabian StyleBisson, Francis, Éloise Rochefort, Amélie Lavoie, Danielle Larouche, Karine Zaniolo, Carolyne Simard-Bisson, Odile Damour, François A. Auger, Sylvain L. Guérin, and Lucie Germain. 2013. "Irradiated Human Dermal Fibroblasts Are as Efficient as Mouse Fibroblasts as a Feeder Layer to Improve Human Epidermal Cell Culture Lifespan" International Journal of Molecular Sciences 14, no. 3: 4684-4704. https://doi.org/10.3390/ijms14034684
APA StyleBisson, F., Rochefort, É., Lavoie, A., Larouche, D., Zaniolo, K., Simard-Bisson, C., Damour, O., Auger, F. A., Guérin, S. L., & Germain, L. (2013). Irradiated Human Dermal Fibroblasts Are as Efficient as Mouse Fibroblasts as a Feeder Layer to Improve Human Epidermal Cell Culture Lifespan. International Journal of Molecular Sciences, 14(3), 4684-4704. https://doi.org/10.3390/ijms14034684