A Comparison of B16 Melanoma Cells and 3T3 Fibroblasts Concerning Cell Viability and ROS Production in the Presence of Melatonin, Tested Over a Wide Range of Concentrations

 , ,
, ,
Abstract
:1. Introduction
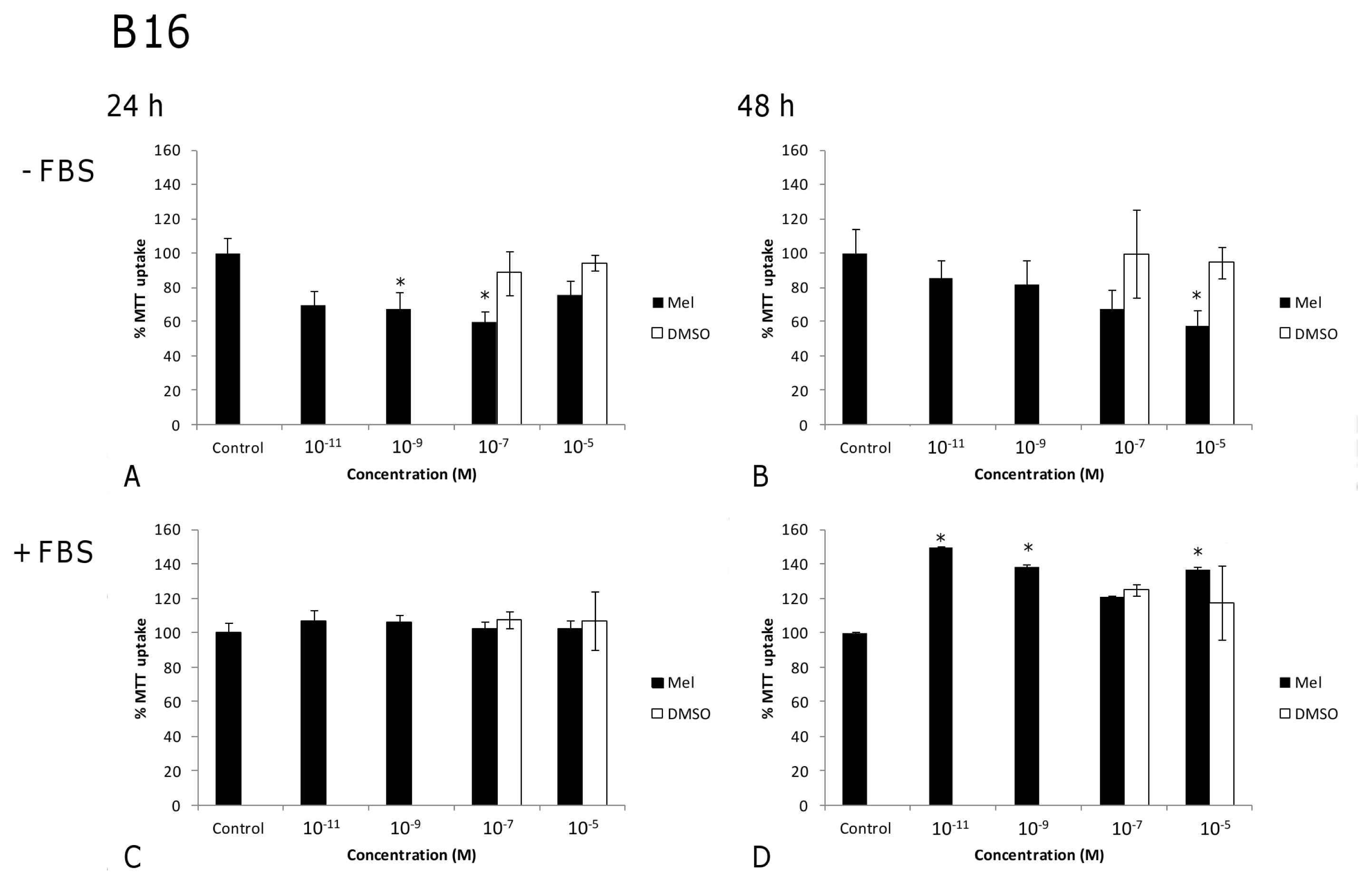
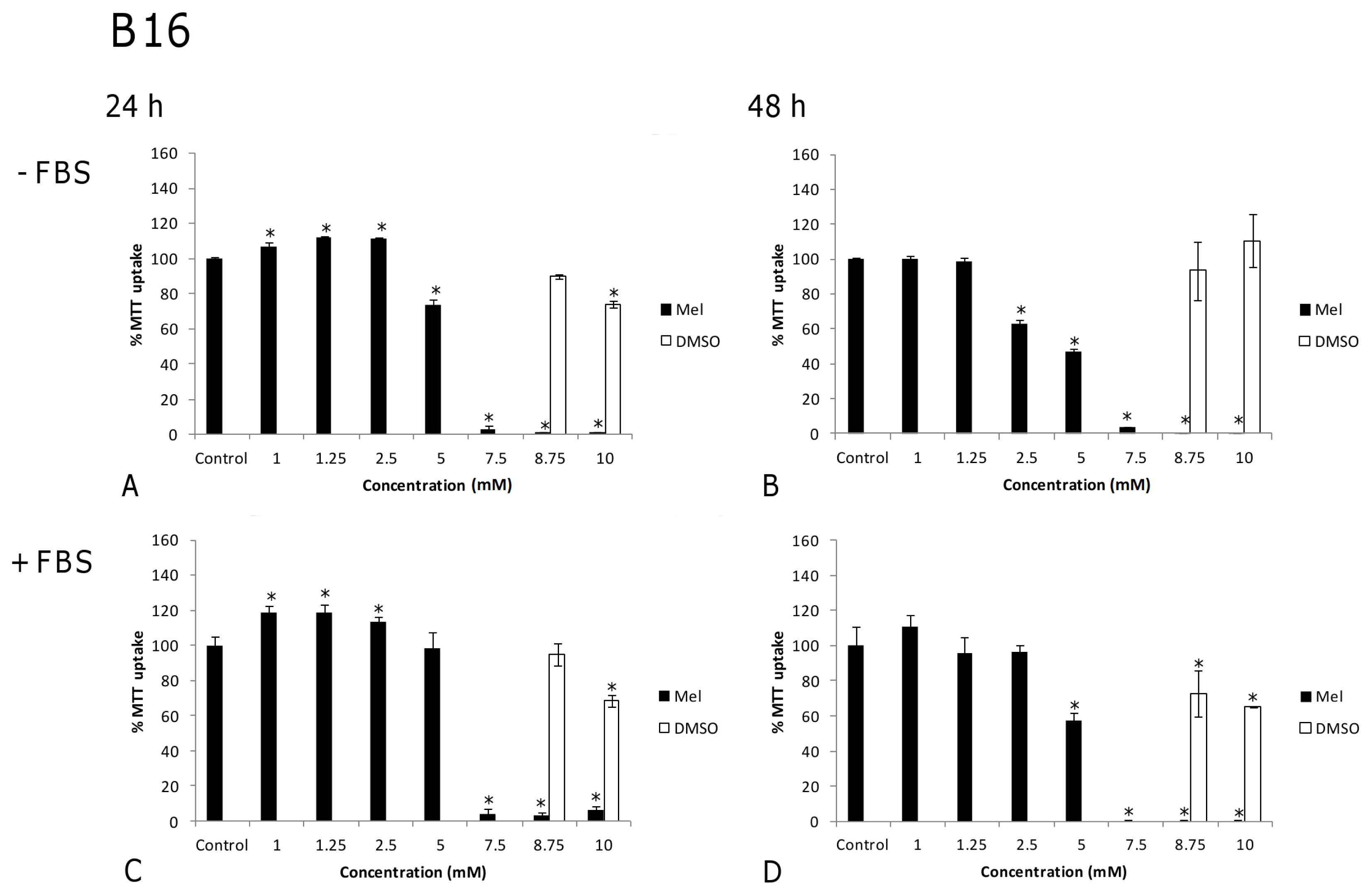
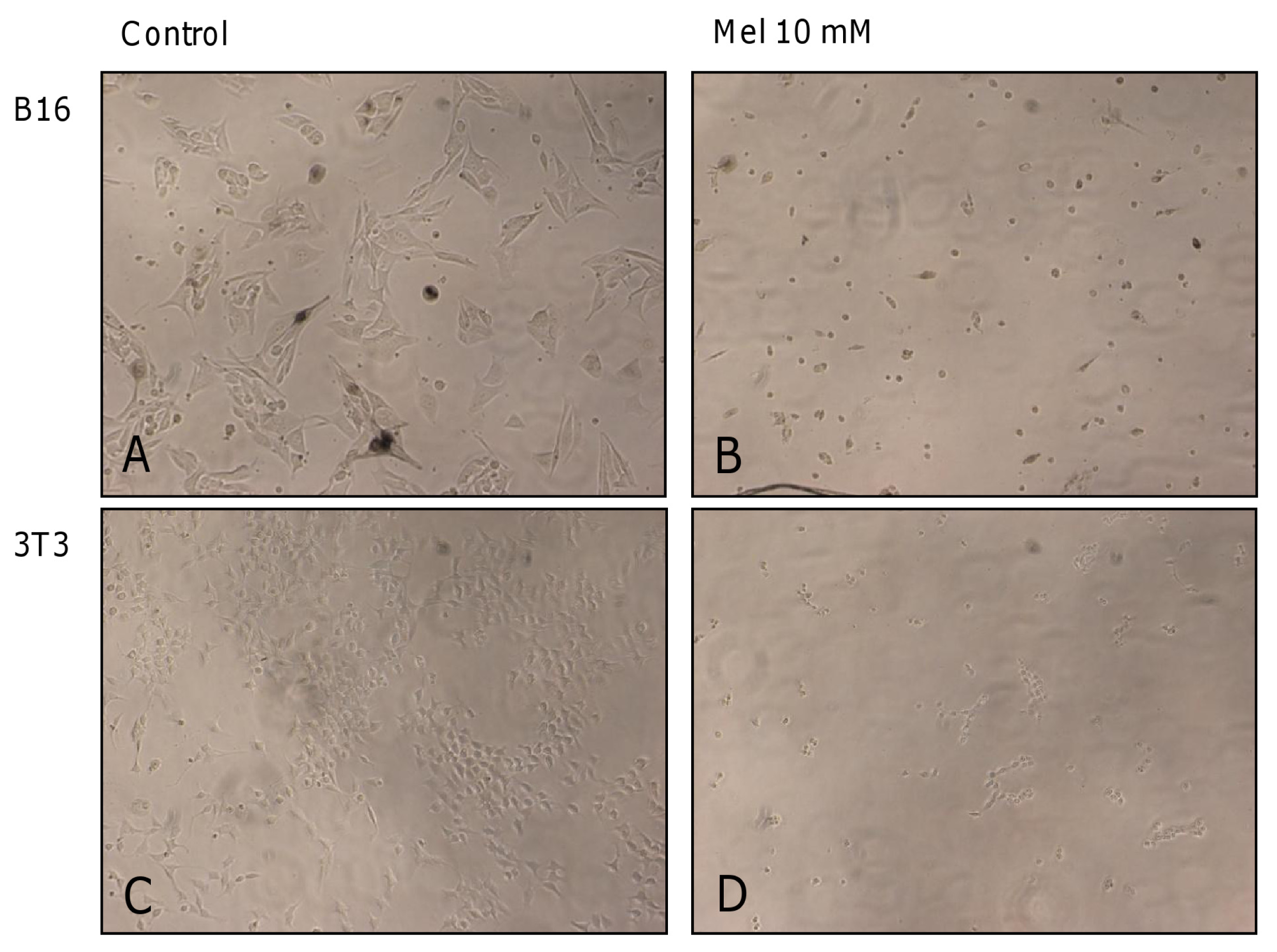
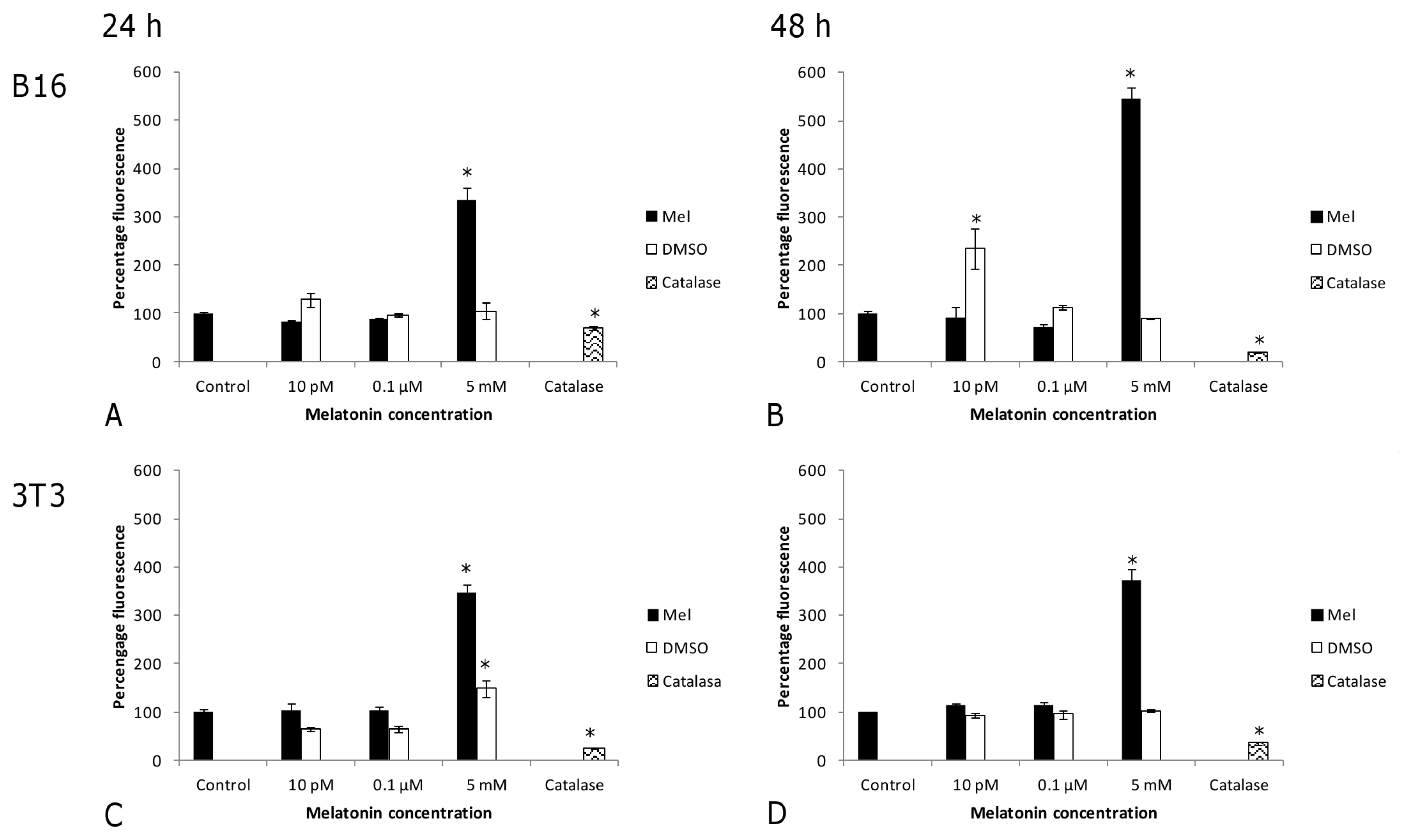
2. Results and Discussion
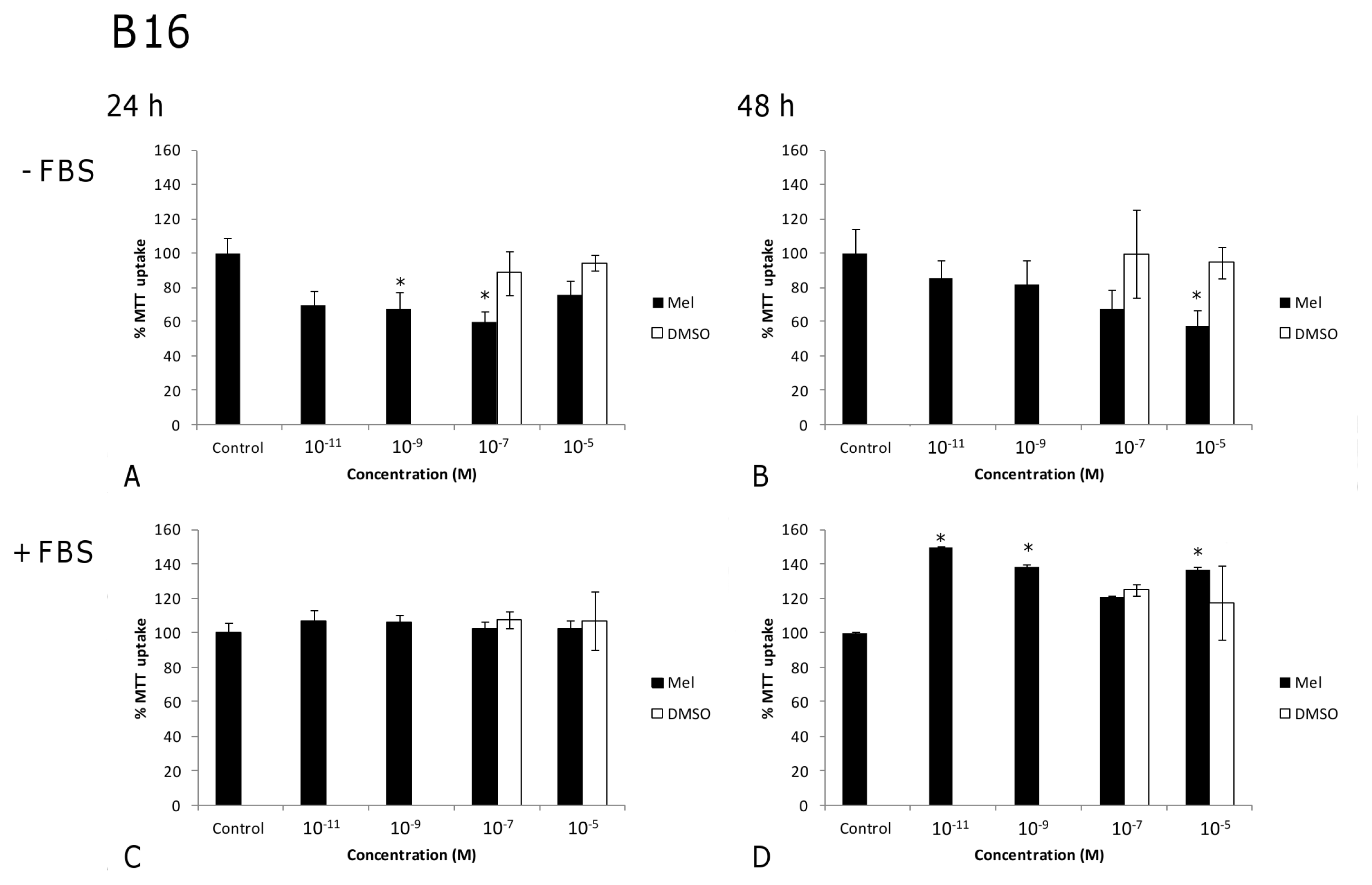
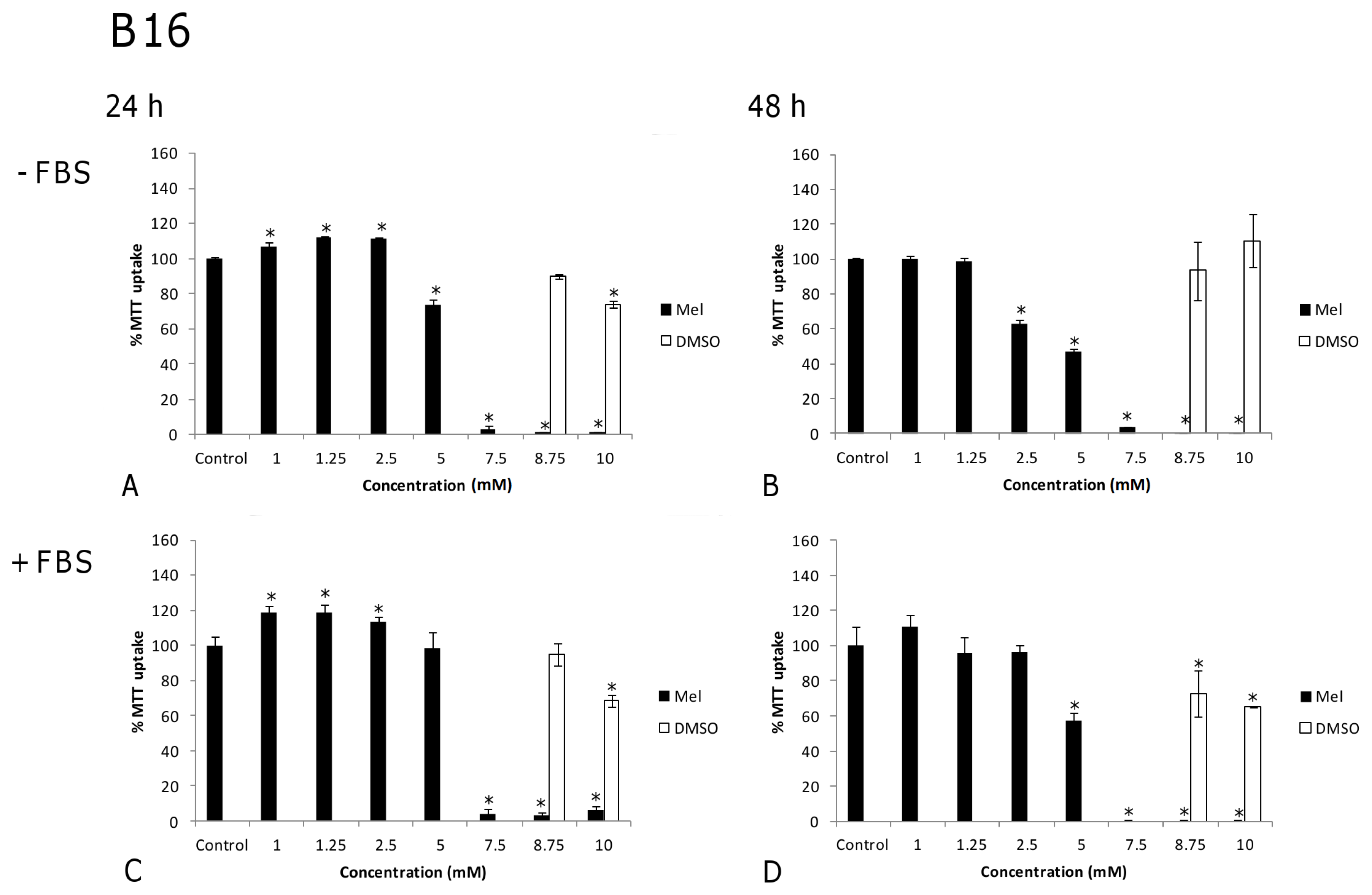
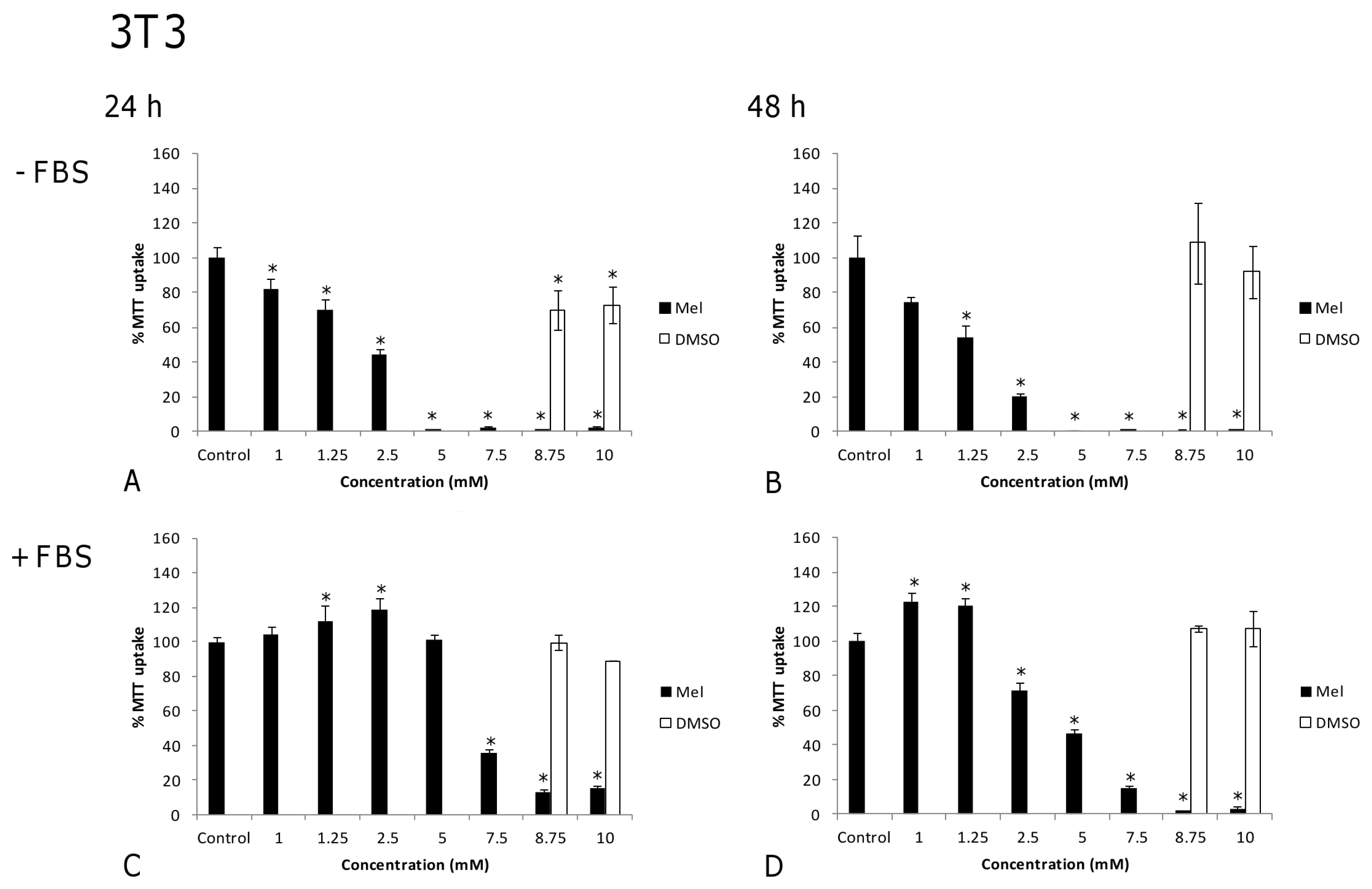
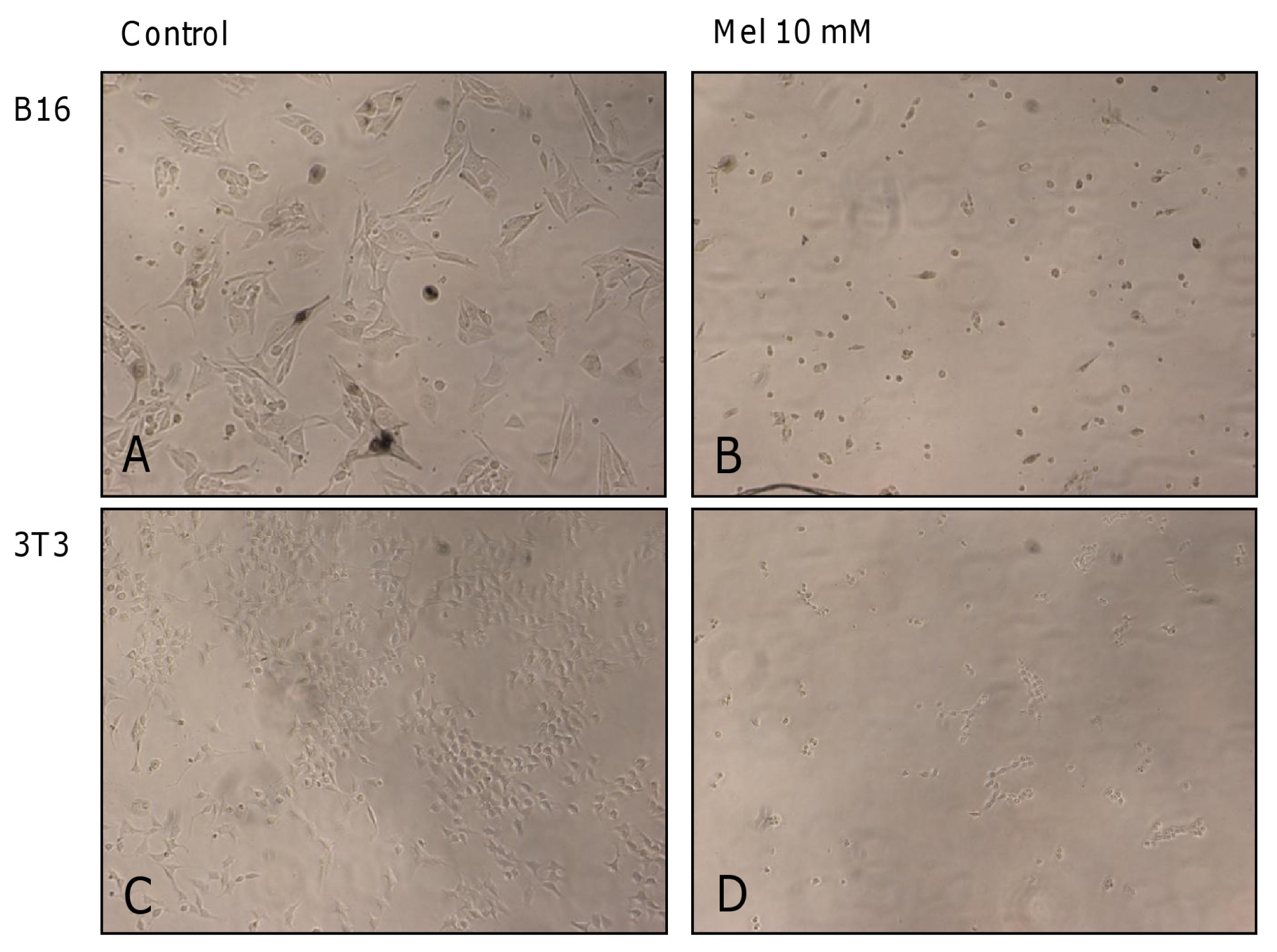
2.1. Results
2.2. Discussion
3. Experimental Section
3.1. Cell Culture
3.2. Cell Viability Assay
3.3. ROS Production Assay
3.4. Microphotographs
3.5. Statistical Analysis
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Hardeland, R. Melatonin: Signaling mechanisms of a pleiotropic agent. Biofactors 2009, 35, 183–192. [Google Scholar]
- Hardeland, R.; Cardinali, D.P.; Srinivasan, V.; Spence, D.W.; Brown, G.M.; Pandi-Perumal, S.R. Melatonin—A pleiotropic, orchestrating regulator molecule. Prog. Neurobiol 2011, 93, 350–384. [Google Scholar]
- Sanchez-Hidalgo, M.; Alarcon de la Lastra, C.; Carrascosa-Salmoral, M.P.; Naranjo, M.C.; Gomez-Corvera, A.; Caballero, B.; Guerrero, J.M. Age-related changes in melatonin synthesis in rat extrapineal tissues. Exp. Gerontol 2009, 44, 328–334. [Google Scholar]
- Koch, B.C.; Nagtegaal, J.E.; Kerkhof, G.A.; ter Wee, P.M. Circadian sleep-wake rhythm disturbances in end-stage renal disease. Nat. Rev. Nephrol 2009, 5, 407–416. [Google Scholar]
- Hardeland, R.; Madrid, J.A.; Tan, D.X.; Reiter, R.J. Melatonin, the circadian multioscillator system and health: The need for detailed analyses of peripheral melatonin signaling. J. Pineal. Res 2012, 52, 139–166. [Google Scholar]
- Carrillo-Vico, A.; Guerrero, J.M.; Lardone, P.J.; Reiter, R.J. A review of the multiple actions of melatonin on the immune system. Endocrine 2005, 27, 189–200. [Google Scholar]
- Reiter, R.J.; Tan, D.X.; Burkhardt, S. Reactive oxygen and nitrogen species and cellular and organismal decline: Amelioration with melatonin. Mech. Ageing Dev 2002, 123, 1007–1019. [Google Scholar]
- Otalora, B.B.; Madrid, J.A.; Alvarez, N.; Vicente, V.; Rol, M.A. Effects of exogenous melatonin and circadian synchronization on tumor progression in melanoma-bearing C57BL6 mice. J. Pineal. Res 2008, 44, 307–315. [Google Scholar]
- Cutando, A.; López-Valverde, A.; Arias-Santiago, S.; de Vicente, J.; de Diego, R.G. Role of melatonin in cancer treatment. Anticancer Res 2012, 32, 2747–2753. [Google Scholar]
- Ghislin, S.; Deshayes, F.; Lauriol, J.; Middendorp, S.; Martins, I.; Al-Daccak, R.; Alcaide-Loridan, C. Plasticity of melanoma cells induced by neural cell crest conditions and three-dimensional growth. Melanoma Res 2012, 22, 184–194. [Google Scholar]
- Blask, D.E.; Sauer, L.A.; Dauchy, R.; Holowachuk, E.W.; Ruhoff, M.S. New actions of melatonin on tumor metabolism and growth. Biol. Signals Recept 1999, 8, 49–55. [Google Scholar]
- Cabrera, J.; Negrín, G.; Estévez, F.; Loro, J.; Reiter, R.J.; Quintana, J. Melatonin decreases cell proliferation and induces melanogenesis in human melanoma SK-MEL-1 cells. J. Pineal. Res 2010, 49, 45–54. [Google Scholar]
- Hill, S.M.; Blask, D.E.; Xiang, S.; Yuan, L.; Mao, L.; Dauchy, R.T; Dauchy, E.M.; Frasch, T.; Duplesis, T. Melatonin and associated signaling pathways that control normal breast epithelium and breast cancer. J. Mammary Gland Biol. Neoplasia 2011, 16, 235–245. [Google Scholar]
- Margheri, M.; Pacini, N.; Tani, A.; Nosi, D.; Squecco, R.; Dama, A.; Masala, E.; Francini, F.; Zecchi-Orlandini, S.; Formigli, L. Combined effects of melatonin and all-trans retinoic acid and somatostatin on breast cancer cell proliferation and death: Molecular basis for the anticancer effect of these molecules. Eur. J. Pharmacol 2012, 681, 34–43. [Google Scholar]
- Shiu, S.Y.; Leung, W.Y.; Tam, C.W.; Liu, V.W.; Yao, K.M. Melatonin MT1 receptor-induced transcriptional up-regulation of p27(Kip1) in prostate cancer antiproliferation is mediated via inhibition of constitutively active nuclear factor kappa B (NF-κB): Potential implications on prostate cancer chemoprevention and therapy. J. Pineal. Res. 2012. [Google Scholar] [CrossRef]
- Martín-Renedo, J.; Mauriz, J.L.; Jorquera, F.; Ruiz-Andrés, O.; González, P.; González-Gallego, J. Melatonin induces cell cycle arrest and apoptosis in hepatocarcinoma HepG2 cell line. J. Pineal. Res. 2008, 45, 532–540. [Google Scholar]
- Carbajo-Pescador, S.; Martín-Renedo, J.; García-Palomo, A.; Tuñón, M.J.; Mauriz, J.L.; González-Gallego, J. Changes in the expression of melatonin receptors induced by melatonin treatment in hepatocarcinoma HepG2 cells. J. Pineal. Res 2009, 47, 330–338. [Google Scholar]
- Radogna, F.; Paternoster, L.; Albertini, M.C.; Accorsi, A.; Cerella, C.; D’Alessio, M.; de Nicola, M.; Nuccitelli, S.; Magrini, A.; Bergamaschi, A.; et al. Melatonin as an apoptosis antagonist. Ann. N. Y. Acad. Sci 2006, 1090, 226–233. [Google Scholar]
- Cos, S.; Mediavilla, M.D.; Fernández, R.; González-Lamuño, D.; Sánchez-Barceló, E.J. Does melatonin induce apoptosis in MCF-7 human breast cancer cells in vitro? J. Pineal. Res 2002, 32, 90–96. [Google Scholar]
- Cucina, A.; Proietti, S.; D’Anselmi, F.; Coluccia, P.; Dinicola, S.; Frati, L.; Bizzarri, M. Evidence for a biphasic apoptotic pathway induced by melatonin in MCF-7 breast cancer cells. J. Pineal. Res 2009, 46, 172–180. [Google Scholar]
- Mao, L.; Cheng, Q.; Guardiola-Lemaître, B.; Schuster-Klein, C.; Dong, C.; Lai, L.; Hill, S.M. In vitro and in vivo antitumor activity of melatonin receptor agonists. J. Pineal. Res 2010, 49, 210–221. [Google Scholar]
- Yang, Q.H.; Xu, J.-N.; Xu, R.K.; Pang, S.F. Antiproliferative effects of melatonin on the growth of rat pituitary prolactin-secreting tumor cells in vitro. J. Pineal. Res 2007, 42, 172–179. [Google Scholar]
- Esposito, E.; Iacono, A.; Muià, C.; Crisafulli, C.; Raso, G.M.; Bramanti, P.; Meli, R.; Cuzzocrea, S. Signal transduction pathways involved in protective effects of melatonin in C6 glioma cells. J. Pineal. Res 2008, 44, 78–87. [Google Scholar]
- Roth, J.A.; Rabin, R.; Agnello, K. Melatonin suppression of PC12 cell growth and death. Brain Res 1997, 768, 63–70. [Google Scholar]
- Ying, S.W.; Niles, L.P.; Crocker, C. Human malignant melanoma cells express high-affinity receptors for melatonin: Antiproliferative effects of melatonin and 6-chloromelatonin. Eur. J. Pharmacol 1993, 246, 89–96. [Google Scholar]
- Hu, D.N.; Roberts, J.E. Melatonin inhibits growth of cultured human uveal melanoma cells. Melanoma Res 1997, 7, 27–31. [Google Scholar]
- Hu, D.N.; McCormick, S.A.; Roberts, J.E. Effects of melatonin, its precursors and derivatives on the growth of cultured human uveal melanoma cells. Melanoma Res 1998, 8, 205–210. [Google Scholar]
- Helton, R.A.; Harrison, W.A.; Kelley, K.; Kane, M.A. Melatonin interactions with cultured murine B16 melanoma cells. Melanoma Res 1993, 3, 403–413. [Google Scholar]
- Cos, S.; Garcia-Bolado, A.; Sánchez-Barceló, E.J. Direct antiproliferative effects of melatonin on two metastatic cell sublines of mouse melanoma (B16BL6 and PG19). Melanoma Res 2001, 11, 197–201. [Google Scholar]
- Fischer, T.W.; Zmijewski, M.A.; Zbytek, B.; Sweatman, T.W.; Slominski, R.M.; Wortsman, J.; Slominski, A. Oncostatic effects of the indole melatonin and expression of its cytosolic and nuclear receptors in cultured human melanoma cell lines. Int. J. Oncol 2006, 29, 665–672. [Google Scholar]
- Panzer, A.; Lottering, M.L.; Bianchi, P.; Glencross, D.K.; Stark, J.H.; Seegers, J.C. Melatonin has no effect on the growth, morphology or cell cycle of human breast cancer (MCF-7), cervical cancer (HeLa), osteosarcoma (MG-63) or lymphoblastoid (TK6) cells. Cancer Lett 1998, 122, 17–23. [Google Scholar]
- Yerneni, L.K.; Jayaraman, S. Pharmacological action of high doses of melatonin on B16 murine melanoma cells depends on cell number at time of exposure. Melanoma Res 2003, 13, 113–117. [Google Scholar]
- Mengeaud, V.; Skene, D.; Pévet, P.; Ortonne, J.P. No high affinity melatonin binding sites are detected in murine melanoma cells and in normal human melanocytes cultured in vitro. Melanoma Res 1994, 4, 87–91. [Google Scholar]
- Carbajo-Pescador, S.; García-Palomo, A.; Martín-Renedo, J.; Piva, M.; González-Gallego, J.; Mauriz, J.L. Melatonin modulation of intracellular signaling pathways in hepatocarcinoma HepG2 cell line: Role of the MT1 receptor. J. Pineal. Res 2011, 51, 463–471. [Google Scholar]
- Barlow-Walden, L.R.; Reiter, R.J.; Abe, M.; Pablos, M.; Menendez-Pelaez, A.; Chen, L.D.; Poeggeler, B. Melatonin stimulates brain glutathione peroxidase activity. Neurochem. Int 1995, 26, 497–502. [Google Scholar]
- Rodriguez, C.; Mayo, J.C.; Sainz, R.M.; Antolín, I.; Herrera, F.; Martín, V.; Reiter, R.J. Regulation of antioxidant enzymes: A significant role for melatonin. J. Pineal. Res 2004, 36, 1–9. [Google Scholar]
- Leon, J.; Acuña-Castroviejo, D.; Sainz, R.M.; Mayo, J.C.; Tan, D.-X.; Reiter, R.J. Melatonin and mitochondrial function. Minireview. Life Sci 2004, 75, 765–790. [Google Scholar]
- Morrey, K.M.; McLachlan, J.A.; Serkin, C.D.; Bakouche, O. Activation of human monocytes by the pineal hormone melatonin. J. Immunol 1994, 153, 2671–2680. [Google Scholar]
- Radogna, F.; Paternoster, L.; de Nicola, M.; Cerella, C.; Ammendola, S.; Bedini, A.; Tarzia, G.; Aquilano, K.; Ciriolo, M.; Ghibelli, L. Rapid and transient stimulation of intracellular reactive oxygen species by melatonin in normal and tumor leukocytes. Toxicol. Appl. Pharmacol 2009, 239, 37–45. [Google Scholar]
- Cristofanon, S.; Uguccioni, F.; Cerella, C.; Radogna, F.; Dicato, M.; Ghibelli, L.; Diederich, M. Intracellular prooxidant activity of melatonin induces a survival pathway involving NF-κB activation. Ann. N. Y. Acad. Sci 2009, 1171, 472–478. [Google Scholar]
- García-Mauriño, S.; González-Haba, M.G.; Calvo, J.R.; Goberna, R.; Guerrero, J.M. Involvement of nuclear binding sites for melatonin in the regulation of IL-2 and IL-6 production by human blood mononuclear cells. J. Neuroimmunol 1998, 92, 76–84. [Google Scholar]
- Kühlwein, E.; Irwin, M. Melatonin modulation of lymphocyte proliferation and Th1/Th2 cytokine expression. J. Neuroimmunol 2001, 117, 51–57. [Google Scholar]
- Espino, J.; Bejarano, I.; Paredes, S.D.; Barriga, C.; Rodríguez, A.B.; Pariente, J.A. Protective effect of melatonin against human leukocyte apoptosis induced by intracellular calcium overload: Relation with its antioxidant actions. J. Pineal. Res 2011, 51, 195–206. [Google Scholar]
- Bejarano, I.; Redondo, P.C.; Espino, J.; Rosado, J.A.; Paredes, S.D.; Barriga, C.; Reiter, R.J.; Pariente, J.A.; Rodríguez, A.B. Melatonin induces mitochondrial-mediated apoptosis in human myeloid HL-60 cells. J. Pineal. Res 2009, 46, 392–400. [Google Scholar]
- Bejarano, I.; Espino, J.; Barriga, C.; Reiter, R.J.; Pariente, J.A.; Rodríguez, A.B. Pro-oxidant Effect of Melatonin in Tumor Leucocytes: Relation with its Cytotoxic and Pro-Apoptotic Effects. Basic Clin. Pharmacol. Toxicol 2011, 108, 14–20. [Google Scholar]
- Osseni, R.A.; Rat, P.; Bogdan, A.; Warnet, J.M.; Touitou, Y. Evidence of prooxidant and antioxidant action of melatonin on human liver cell line HepG2. Life Sci 2000, 68, 387–399. [Google Scholar]
- Albertini, M.C.; Radogna, F.; Accorsi, A.; Uguccioni, F.; Paternoster, L.; Cerella, C.; de Nicola, M.; D’Alessio, M.; Bergamaschi, A.; Magrini, A.; et al. Intracellular pro-oxidant activity of melatonin deprives U937 cells of reduced glutathione without affecting glutathione peroxidase activity. Ann. N. Y. Acad. Sci 2006, 1091, 10–16. [Google Scholar]
- Zhang, H.M.; Zhang, Y.; Zhang, B.X. The role of mitochondrial complex III in melatonininduced ROS production in cultured mesangial cells. J. Pineal. Res 2011, 50, 78–82. [Google Scholar]
- Zwirska-Korczala, K.; Jochem, J.; Adamczyk-Sowa, M.; Sowa, P.; Polaniak, R.; Birkner, E.; Latocha, M.; Pilc, K.; Suchanek, R. Influence of melatonin on cell proliferation, antioxidative enzyme activities and lipid peroxidation in 3T3-L1 preadipocytes-an in vitro study. J. Physiol. Pharmacol 2005, 56, 91–99. [Google Scholar]
- Nah, S.S.; Won, H.J.; Park, H.J.; Ha, E.; Chung, J.H.; Cho, H.Y.; Baik, H.H. Melatonin inhibits human fibroblast-like synoviocyte proliferation via extracellular signal-regulated protein kinase/P21(CIP1)/P27(KIP1) pathways. J. Pineal. Res 2009, 47, 70–74. [Google Scholar]
- Carossino, A.M.; Lombardi, A.; Matucci-Cerinic, M.; Pignone, A.; Cagnoni, M. Effect of melatonin on normal and sclerodermic skin fibroblast proliferation. Clin. Exp. Rheumatol 1996, 14, 493–498. [Google Scholar]
- Xie, Y.F.; Zhang, J.C.; Liu, S.J.; Dai, L.B.; Du, G.W. Effect of melatonin on proliferation and apoptosis of fibroblasts in human hypertrophic scar. Zhonghua Shao Shang Za Zhi 2011, 27, 422–426. [Google Scholar]
- Alvarez-García, V.; González, A.; Alonso-González, C.; Martínez-Campa, C.; Cos, S. Regulation of vascular endothelial growth factor by melatonin in human breast cancer cells. J. Pineal. Res. 2012. [Google Scholar] [CrossRef]
- González, A.; Alvarez-García, V.; Martínez-Campa, C.; Alonso-González, C.; Cos, S. Melatonin promotes differentiation of 3T3-L1 fibroblasts. J. Pineal. Res 2012, 52, 12–20. [Google Scholar]
- Izykowska, I.; Cegielski, M.; Gebarowska, E.; Podhorska-Okolow, M.; Piotrowska, A.; Zabel, M.; Dziegiel, P. Effect of melatonin on human keratinocytes and fibroblasts subjected to UVA and UVB radiation in vitro. In Vivo 2009, 23, 739–745. [Google Scholar]
- Capoluongo, E. Insulin-like growth factor system and sporadic malignant melanoma. Am. J. Pathol 2011, 178, 26–31. [Google Scholar]
- Ucar, D.A.; Kurenova, E.; Garrett, T.J.; Cance, W.G.; Nyberg, C.; Cox, A.; Massoll, N.; Ostrov, D.A.; Lawrence, N.; Sebti, S.M.; et al. Disruption of the protein interaction between FAK and IGF-1R inhibits melanoma tumor growth. Cell Cycle 2012, 11, 3250–3259. [Google Scholar]
- Ellerhorst, J.A.; Diwan, A.H.; Dang, S.M.; Uffort, D.G.; Johnson, M.K.; Cooke, C.P.; Grimm, E.A. Promotion of melanoma growth by the metabolic hormone leptin. Oncol. Rep 2010, 23, 901–907. [Google Scholar]
- Kadekaro, A.L.; Andrade, L.N.; Floeter-Winter, L.M.; Rollag, M.D.; Virador, V.; Vieira, W.; Castrucci, A.M. MT-1 melatonin receptor expression increases the antiproliferative effect of melatonin on S-91 murine melanoma cells. J. Pineal. Res 2004, 36, 204–211. [Google Scholar]
- Slominski, A.; Pruski, D. Melatonin inhibits proliferation and melanogenesis in rodent melanoma cells. Exp. Cell Res 1993, 206, 189–194. [Google Scholar]
- Pickering, D.S.; Niles, L.P. Expression of nanomolar-affinity binding sites for melatonin in Syrian hamster RPMI 1846 melanoma cells. Cell Signal 1992, 4, 201–207. [Google Scholar]
- Eison, A.S.; Mullins, U.L. Melatonin binding sites are functionally coupled to phosphoinositide hydrolysis in Syrian hamster RPMI 1846 melanoma cells. Life Sci. 1993, 53, PL393–398. [Google Scholar]
- Souza, A.V.; Visconti, M.A.; Castrucci, A.M. Melatonin biological activity and binding sites in human melanoma cells. J. Pineal. Res 2003, 34, 242–248. [Google Scholar]
- Danielczyk, K.; Dziegiel, P. The expression of MT1 melatonin receptor and Ki-67 antigen in melanoma malignum. Anticancer Res 2009, 29, 3887–3895. [Google Scholar]
- Bubis, M.; Zisapel, N. A role for NAD+ and cADP-ribose in melatonin signal transduction. Mol. Cell Endocrinol 1998, 137, 59–67. [Google Scholar]
- Bubis, M.; Zisapel, N. Involvement of cGMP in cellular melatonin responses. Biol. Cell 1999, 91, 45–49. [Google Scholar]
- Bubis, M.; Anis, Y.; Zisapel, N. Enhancement by melatonin of GTP exchange and ADP ribosylation reactions. Mol. Cell Endocrinol 1996, 123, 139–148. [Google Scholar]
- Blask, D.E.; Sauer, L.A.; Dauchy, R.T. Melatonin as a chronobiotic/anticancer agent: Cellular, biochemical, and molecular mechanisms action and their implications for circadian-based cancer therapy. Curr. Top Med. Chem 2002, 2, 113–132. [Google Scholar]
- Benítez-King, G.; Soto-Vega, E.; Ramírez-Rodriguez, G. Melatonin modulates microfilament phenotypes in epithelial cells: Implications for adhesion and inhibition of cancer cell migration. Histol. Histopathol 2009, 24, 789–799. [Google Scholar]
- Hardeland, R.; Poeggeler, B.; Pappolla, M.A. Mitochondrial actions of melatonin—An endeavor to identify their adaptive and cytoprotective mechanisms. Proc. Saxon Acad. Sci 2009, 65, 14–31. [Google Scholar]
- Hardeland, R. Melatonin, mitochondrial electron flux and leakage: Recent findings and resolution of contradictory results. Adv. Stud. Biol 2009, 1, 207–230. [Google Scholar]
- Hardeland, R. Melatonin and its metabolites as anti-nitrosating and anti-nitrating agents. J. Exp. Integ. Med 2011, 1, 67–81. [Google Scholar]
- Chen, T.R. In situ detection of Mycoplasma contamination in cell cultures by fluorescent Hoechst 33258 stain. Exp. Cell Res 1977, 104, 255–262. [Google Scholar]
- Denizot, F.; Lang, R. Rapid colorimetric assay for cell growth and survival. J. Inmunol. Methods 1986, 89, 271–277. [Google Scholar]
- Osseni, R.A.; Debbasch, C.; Christen, M.-O.; Rat, P.; Warnet, J.-M. Tacrine induced reactive oxygen species in a human liver cell line: The role of athenole dithiolethione as a scavenger. Toxicol. In Vitro 1999, 13, 683–688. [Google Scholar]
- Cole, E.R.; Crank, G. The estimation of melatonin in blood serum. Biochem. Med 1972, 8, 37–43. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Osmolality (mOsm/Kg) |
|---|---|
| Control | 333 |
| 10 mM | 425 |
| 8.75 mM | 382 |
| 7.5 mM | 364 |
| 5 mM | 365 |
| 2.5 mM | 346 |
| 1.25 mM | 341 |
| 1 mM | 342 |
© 2013 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Bonmati-Carrion, M.A.; Álvarez-Sánchez, N.; Hardeland, R.; Madrid, J.A.; Rol, M.A. A Comparison of B16 Melanoma Cells and 3T3 Fibroblasts Concerning Cell Viability and ROS Production in the Presence of Melatonin, Tested Over a Wide Range of Concentrations. Int. J. Mol. Sci. 2013, 14, 3901-3920. https://doi.org/10.3390/ijms14023901
Bonmati-Carrion MA, Álvarez-Sánchez N, Hardeland R, Madrid JA, Rol MA. A Comparison of B16 Melanoma Cells and 3T3 Fibroblasts Concerning Cell Viability and ROS Production in the Presence of Melatonin, Tested Over a Wide Range of Concentrations. International Journal of Molecular Sciences. 2013; 14(2):3901-3920. https://doi.org/10.3390/ijms14023901
Chicago/Turabian StyleBonmati-Carrion, Maria Angeles, Nuria Álvarez-Sánchez, Rüdiger Hardeland, Juan Antonio Madrid, and Maria Angeles Rol. 2013. "A Comparison of B16 Melanoma Cells and 3T3 Fibroblasts Concerning Cell Viability and ROS Production in the Presence of Melatonin, Tested Over a Wide Range of Concentrations" International Journal of Molecular Sciences 14, no. 2: 3901-3920. https://doi.org/10.3390/ijms14023901