Melatonin Anticancer Effects: Review

{kind=link}
{kind=link}
Abstract
:1. Introduction: General Considerations on the Anticancer Effect of Melatonin
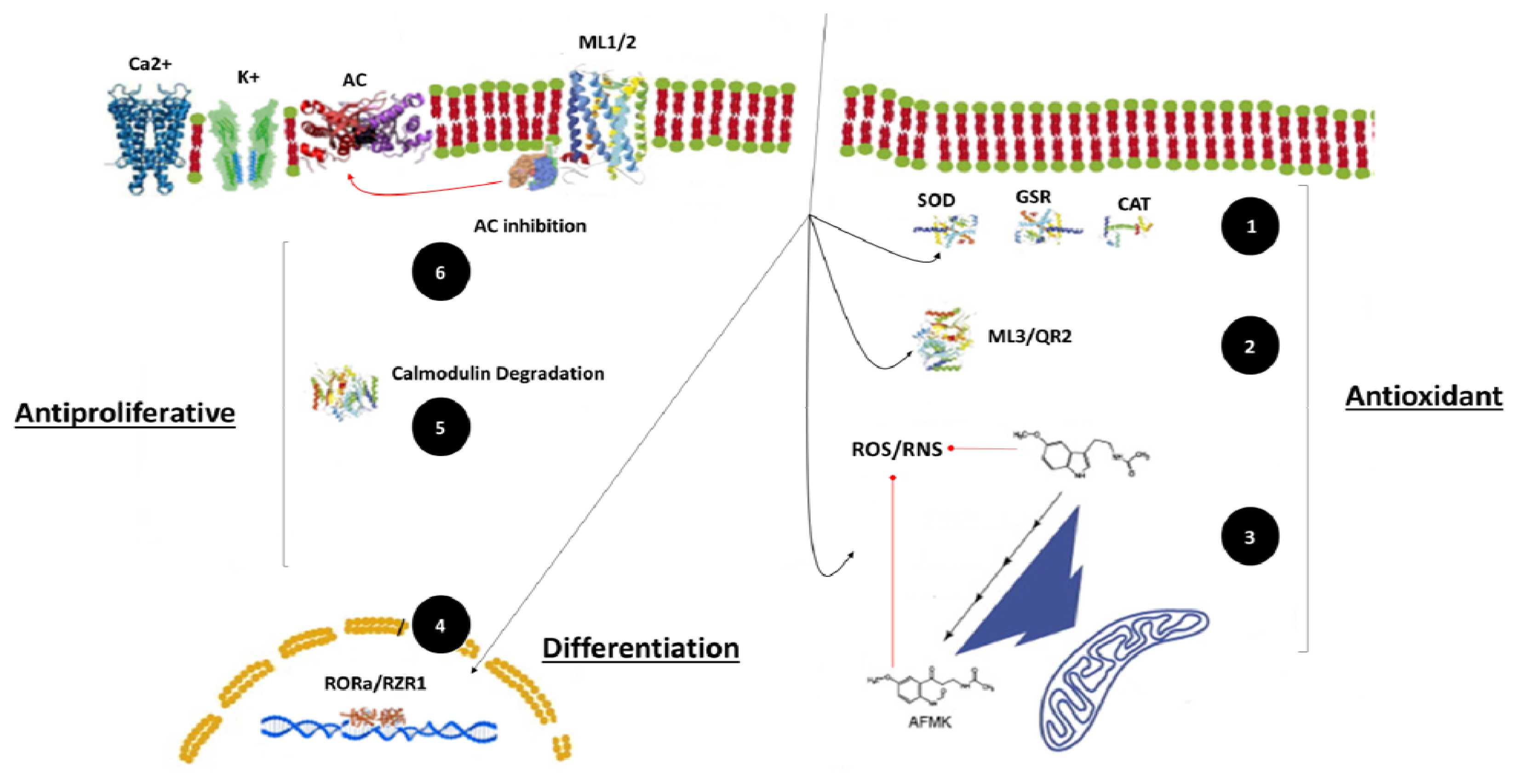
2. The Main Direct Anticancer Mechanisms of Melatonin
2.1. Pro-Apoptotic
2.2. Antiproliferative
2.3. Differentiating
2.4. Anti-Angiogenetic
3. The Main Indirect Anticancer Mechanisms of Melatonin
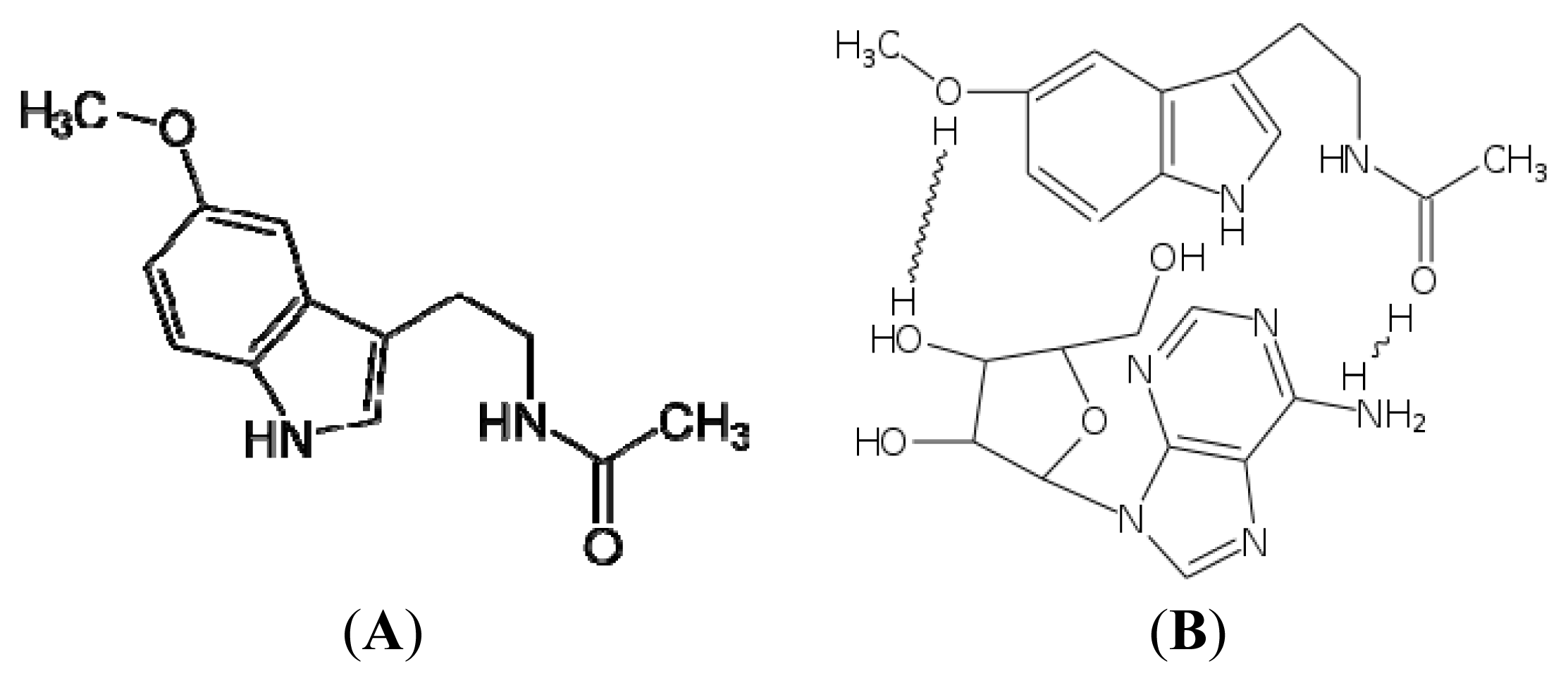
3.1. Free Radical Scavenger Action
3.2. Myeloprotective/Myelostimulant Action
3.3. Melatonin’s Action in Regulating the Immune System
4. Mechanisms of Action and Physiology of Melatonin in Tumors
4.1. The Receptorial System
4.2. Other Mechanisms
4.3. The Amine Precursor Uptake and Decarboxylation System (APUD)
4.4. Platelets and the APUD System
4.5. Melatonin’s Action on Microtubules
5. Melatonin and Tumor Treatment
5.1. Clinical Significance and Therapeutic Application
5.2. Future Prospects after 30 Years of Research
5.3. Indications Regarding the Proposed Dosage of Melatonin in Prevention
5.4. Indications Regarding the Proposed Dosage of Melatonin in the Treatment of Tumors
6. Conclusions
Conflict of Interest
References
- Macchi, M.M.; Bruce, J.N. Human pineal physiology and functional significance of melatonin. Front. Neuroendocrinol 2004, 25, 177–195. [Google Scholar]
- Skwarlo-Sonta, K. Melatonin in immunity: Comparative aspects. Neuro Endocrinol. Lett 2002, 2, 61–66. [Google Scholar]
- Reiter, R.J.; Korkmaz, A. Clinical aspects of melatonin. Saudi Med. J 2008, 29, 1537–1547. [Google Scholar]
- Suwanjang, W.; Phansuwan-Pujito, P.; Govitrapong, P.; Chetsawang, B. The protective effect of melatonin on methamphetamine-induced calpain-dependent death pathway in human neuroblastoma SH-SY5Y cultured cells. J. Pineal Res 2010, 48, 94–101. [Google Scholar]
- Proietti, S.; Cucina, A.; Reiter, R.J.; Bizzarri, M. Molecular mechanisms of melatonin’s inhibitory actions on breast cancers. Cell. Mol. Life Sci 2012. [Google Scholar] [CrossRef]
- Rodriguez-Garcia, A.; Mayo, J.C.; Hevia, D.; Quiros-Gonzalez, I.; Navarro, M.; Sainz, R.M. Phenotypicchanges caused by melatonin increased sensitivity of prostate cancer cells to cytokine-induced apoptosis. J. Pineal Res 2012. [Google Scholar] [CrossRef]
- Zhang, S.; Zuo, L.; Gui, S.; Zhou, Q.; Wei, W.; Wang, Y. Induction of cell differentiation and promotion of endocan gene expression in stomach cancer bymelatonin. Mol. Biol. Rep 2012, 39, 2843–2849. [Google Scholar]
- Gamaleĭ, I.A.; Kirpichnikova, K.M.; Filatova, N.A. Effect of melatonin on the functional properties of transformed cells. Vopr. Onkol 2011, 57, 481–485. [Google Scholar]
- Blask, D.E.; Sauer, L.A.; Dauchy, R.T. Melatonin as a chronobiotic/anticancer agent: Cellular, biochemical, and molecular mechanisms of action and their implications for circadian-based cancer therapy. Curr. Top Med Chem 2002, 2, 113–132. [Google Scholar]
- Sánchez-Barceló, E.J.; Cos, S.; Mediavilla, D.; Martínez-Campa, C.; González, A.; Alonso-González, C. Melatonin-estrogen interactions in breast cancer. J. Pineal Res 2005, 38, 217–222. [Google Scholar]
- Bartsch, H.; Buchberger, A.; Franz, H.; Bartsch, C.; Maidonis, I.; Mecke, D.; Bayer, E. Effect of melatonin and pineal extracts on human ovarian and mammary tumor cells in a chemosensitivity assay. Life Sci 2000, 67, 2953–2960. [Google Scholar]
- Watanabe, M.; Kobayashi, Y.; Takahashi, N.; Kiguchi, K.; Ishizuka, B. Expression of melatonin receptor (MT1) and interaction between melatonin and estrogen in endometrial cancer cell line. J. Obstet. Gynaecol. Res 2008, 34, 567–573. [Google Scholar]
- Heldin, C.H.; Westermark, B. Platelet-derived growth factor and autocrine mechanisms of oncogenic processes. Crit. Rev. Oncog 1991, 2, 109–124. [Google Scholar]
- Lüscher, T.F.; Boulanger, C.M.; Dohi, Y.; Yang, Z.H. Endothelium-derived contracting factors. Hypertension 1992, 19, 117–130. [Google Scholar]
- Comoglio, P.M. Structure, biosynthesis and biochemical properties of the HGF receptor in normal and malignant cells. EXS 1993, 65, 131–165. [Google Scholar]
- Cos, S.; Blask, D.E. Melatonin modulates growth factor activity in MCF-7 human breast cancer cells. J. Pineal Res 1994, 17, 25–32. [Google Scholar]
- Boonstra, J.; Rijken, P.; Humbel, B.; Cremers, F.; Verkleij, A.; van Bergen en Henegouwen, P. The epidermal growth factor. Cell Biol. Int 1995, 19, 413–430. [Google Scholar]
- Ornitz, D.M.; Itoh, N. Fibroblast growth factors. Genome Biol 2001, 2, 1–12. [Google Scholar]
- Ferrara, N.; Gerber, H.P. The role of vascular endothelial growth factor in angiogenesis. Acta Haematol 2002, 106, 148–156. [Google Scholar]
- Trejo, J.L.; Carro, E.; Garcia-Galloway, E.; Torres-Aleman, I. Role of insulin-like growth factor I signaling in neurodegenerative diseases. J. Mol. Med 2004, 82, 156–162. [Google Scholar]
- Matt, P.; Schoenhoff, F.; Habashi, J.; Holm, T.; Van Erp, C.; Loch, D.; Carlson, O.D.; Griswold, B.F.; Fu, Q.; De Backer, J.; et al. Circulating transforming growth factor-β in Marfan syndrome. Circulation 2009, 120, 526–532. [Google Scholar]
- Fischer, T.W.; Zmijewski, M.A.; Wortsman, J.; Slominski, A. Melatonin maintains mitochondrial membrane potential and attenuates activation of initiator (casp-9) and effector caspases (casp-3/ casp-7) and PARP in UVR-exposed HaCaT keratinocytes. J. Pineal Res 2008, 44, 397–407. [Google Scholar]
- Ferreira Cda, S.; Maganhin, C.C.; Simões Rdos, S.; Girão, M.J.; Baracat, E.C.; Soares-Jr, J.M. Melatonin: Cell death modulator. Rev. Assoc. Med. Bras 2010, 56, 715–718. [Google Scholar]
- Sánchez-Hidalgo, M.; Guerrero, J.M.; Villegas, I.; Packham, G.; de la Lastra, C.A. Melatonin, a natural programmed cell death inducer in cancer. Curr. Med. Chem 2012, 19, 3805–3821. [Google Scholar]
- Lissoni, P.; Bolis, S.; Brivio, F.; Fumagalli, L. A phase II study of neuroimmunotherapy with subcutaneous low-dose IL-2 plus the pineal hormone melatonin in untreatable advanced hematologic malignancies. Anticancer Res 2000, 20, 2103–2105. [Google Scholar]
- Trubiani, O.; Recchioni, R.; Moroni, F.; Pizzicannella, J.; Caputi, S.; Di Primio, R. Melatonin provokes cell death in human B-lymphoma cells by mitochondrial-dependent apoptotic pathway activation. J. Pineal Res 2005, 39, 425–431. [Google Scholar]
- Bejarano, I.; Redondo, P.C.; Espino, J.; Rosado, J.A.; Paredes, S.D.; Barriga, C.; Reiter, R.J.; Pariente, J.A.; Rodríguez, A.B. Melatonin induces mitochondrial-mediated apoptosis in human myeloid HL-60 cells. J. Pineal Res 2009, 46, 392–400. [Google Scholar]
- Sánchez-Hidalgo, M.; Lee, M.; de la Lastra, C.A.; Guerrero, J.M.; Packham, G. Melatonin inhibits cell proliferation and induces caspase activation and apoptosis in human malignant lymphoid cell lines. J. Pineal Res 2012, 53, 366–373. [Google Scholar]
- Eck-Enriquez, K.; Kiefer, T.L.; Spriggs, L.L.; Hill, S.M. Pathways through which a regimen of melatonin and retinoic acid induces apoptosis in MCF-7 human breast cancer cells. Breast Cancer Res. Treat 2000, 61, 229–239. [Google Scholar]
- Czeczuga-Semeniuk, E.; Wołczyński, S.; Anchim, T.; Dziecioł, J.; Dabrowska, M.; Pietruczuk, M. Effect of melatonin and all-trans retinoic acid on the proliferation and induction of the apoptotic pathway in the culture of human breast cancer cell line MCF-7. Pol. J. Pathol 2002, 53, 59–65. [Google Scholar]
- Dong, C.; Yuan, L.; Dai, J.; Lai, L.; Mao, L.; Xiang, S.; Rowan, B.; Hill, S.M. Melatonin inhibits mitogenic cross-talk between retinoic acid-related orphan receptor alpha (RORalpha) and ERalpha in MCF-7 human breast cancer cells. Steroids 2010, 75, 944–951. [Google Scholar]
- Margheri, M.; Pacini, N.; Tani, A.; Nosi, D.; Squecco, R.; Dama, A.; Masala, E.; Francini, F.; Zecchi-Orlandini, S.; Formigli, L. Combined effects of melatonin and all-trans retinoic acid and somatostatin on breast cancer cell proliferation and death: Molecular basis for the anticancer effect of these molecules. Eur. J. Pharmacol 2012, 681, 34–43. [Google Scholar]
- Lissoni, P.; Rovelli, F.; Frassineti, A.; Fumagalli, L.; Malysheva, O.; Conti, A.; Maestroni, G. Oncostatic activity of pineal neuroendocrine treatment with the pineal indoles melatonin and 5-methoxytryptamine in untreatable metastatic cancer patients progressing on melatonin alone. Neuro Endocrinol. Lett 2000, 21, 319–323. [Google Scholar]
- Tam, C.W.; Chan, K.W.; Liu, V.W.; Pang, B.; Yao, K.M.; Shiu, S.Y. Melatonin as a negative mitogenic hormonal regulator of human prostate epithelial cell growth: Potential mechanisms and clinical significance. J. Pineal Res 2008, 45, 403–412. [Google Scholar]
- Sánchez-Sánchez, A.M.; Martín, V.; García-Santos, G.; Rodríguez-Blanco, J.; Casado-Zapico, S.; Suarez-Garnacho, S.; Antolín, I.; Rodriguez, C. Intracellular redox state as determinant for melatonin antiproliferative vs. cytotoxic effects in cancer cells. Free Radic. Res 2011, 45, 1333–1341. [Google Scholar]
- Cabrera, J.; Negrín, G.; Estévez, F.; Loro, J.; Reiter, R.J.; Quintana, J. Melatonin decreases cell proliferation and induces melanogenesis in human melanoma SK-MEL-1 cells. J. Pineal Res 2010, 49, 45–54. [Google Scholar]
- Pizarro, J.G.; Yeste-Velasco, M.; Esparza, J.L.; Verdaguer, E.; Pallàs, M.; Camins, A.; Folch, J. The antiproliferative activity of melatonin in B65 rat dopaminergic neuroblastoma cells is related to the downregulation of cell cycle-related genes. J. Pineal Res 2008, 45, 8–16. [Google Scholar]
- Martín, V.; Herrera, F.; Carrera-Gonzalez, P.; García-Santos, G.; Antolín, I.; Rodriguez-Blanco, J.; Rodriguez, C. Intracellular signaling pathways involved in the cell growth inhibition of glioma cells by melatonin. Cancer Res 2006, 66, 1081–1088. [Google Scholar]
- Di Bella, L. Melatonin: An Essential Factor for the Treatment and Recovery from Leukemia and Cancer. Proceedings of International Symposium on Melatonin, Bremen, Germany, September 1980; pp. 161–162.
- Lissoni, P.; Barni, S.; Cattaneo, G.; Tancini, G.; Esposti, G.; Esposti, D.; Fraschini, F. Clinical result with the pineal hormone melatonin in advanced cancer resistant to standard antitumor therapies. Oncology 1991, 48, 48–50. [Google Scholar]
- Bubenik, G.A.; Blask, D.E.; Brown, G.M.; Maestroni, G.J.; Pang, S.F.; Reiter, R.J.; Viswanathan, M.; Zisapel, N. Prospects of the clinical utilization of melatonin. Biol. Signals Recept 1998, 7, 195–219. [Google Scholar]
- Di Bella, G. The Di Bella Method (DBM). Neuro. Endocrinol. Lett 2010, 31, 1–42. [Google Scholar]
- El-Missiry, M.A.; Abd El-Aziz, A.F. Influence of melatonin on proliferation and antioxidant system in Ehrlich ascites carcinoma cells. Cancer Lett 2000, 151, 119–125. [Google Scholar]
- Lissoni, P.; Barni, S.; Mandala, M.; Ardizzoia, A.; Paolorossi, F.; Vaghi, M.; Longarini, R.; Malugani, F.; Tancini, G. Decreased toxicity and increased efficacy of cancer chemotherapy using the pineal hormone melatonin in metastatic solid tumor patients with poor clinical status. Eur. J. Cancer 1999, 35, 1688–1692. [Google Scholar]
- Mediavilla, M.D.; Cos, S.; Sanchez-Barcelo, E.J. Melatonin increases p53 and p21WAF1 expression in MCF-7 human breast cancer cells in vitro. Life Sci 1999, 65, 415–420. [Google Scholar]
- Mełen-Mucha, G.; Winczyk, K.; Pawlikowski, M. Somatostatin analogue octreotide and melatonin inhibit bromodeoxyuridine incorporation into cell nuclei and enhance apoptosis in the transplantable murine colon 38 cancer. Anticancer Res 1998, 18, 3615–3619. [Google Scholar]
- Mediavilla, M.D.; Güezmez, A.; Ramos, S.; Kothari, L.; Garijo, F.; Sánchez Barceló, E.J. Effects of melatonin on mammary gland lesions in transgenic mice overexpressing N-ras proto-oncogene. J. Pineal Res 1997, 22, 86–94. [Google Scholar]
- Haus, E.; Dumitriu, L.; Nicolau, G.Y.; Bologa, S.; Sackett-Lundeen, L. Circadian rhythms of basic fibroblast growth factor (bFGF), epidermal growth factor (EGF), insulin-like growth factor-1 (IGF-1), insulin-like growth factor binding protein-3 (IGFBP-3), cortisol, and melatonin in women with breast cancer. Chronobiol. Int 2001, 18, 709–727. [Google Scholar]
- Luchetti, F.; Betti, M.; Canonico, B.; Arcangeletti, M.; Ferri, P.; Galli, F.; Papa, S. ERK MAPK activation mediates the antiapoptotic signaling of melatonin in UVB-stressed U937 cells. Free Radic Biol. Med 2009, 46, 339–351. [Google Scholar]
- Silverman, A.L.; Bronstein, J.C.; Krymgold, S.; Kahlon, D.; Bull, A.W. Decreased levels of 13-hydroxyoctadecadienoic acid (13-HODE) dehydrogenase in neoplastic tissue of human colon biopsies. Cancer Epidemiol. Biomark. Prev 1996, 5, 53–56. [Google Scholar]
- Hill, S.M.; Blask, D.E.; Xiang, S.; Yuan, L.; Mao, L.; Dauchy, R.T.; Dauchy, E.M.; Frasch, T.; Duplesis, T. Melatonin and associated signaling pathways that control normal breast epithelium and breast cancer. J. Mammary Gland Biol. Neoplasia 2011, 16, 235–245. [Google Scholar]
- Lissoni, P.; Rovelli, F.; Malugani, F.; Bucovec, R.; Conti, A.; Maestroni, G.J. Anti-angiogenic activity of melatonin in advanced cancer patients. Neuro Endocrinol. Lett 2001, 22, 45–47. [Google Scholar]
- SoybIr, G.; Topuzlu, C.; OdabaS, O.; Dolay, K.; Bilir, A.; Oksoy, K.F. The effects of melatonin on angiogenesis and wound healing. Surg. Today 2003, 33, 896–901. [Google Scholar]
- Alvarez-García, V.; González, A.; Alonso-González, C.; Martínez-Campa, C.; Cos, S. Regulation of vascular endothelial growth factor by melatonin in human breast cancer cells. J. Pineal Res 2012. [Google Scholar] [CrossRef]
- Park, S.Y.; Jang, W.J.; Yi, E.Y.; Jang, J.Y.; Jung, Y.; Jeong, J.W.; Kim, Y.J. Melatonin suppresses tumor angiogenesis by inhibiting HIF-1alpha stabilization under hypoxia. J. Pineal Res 2010, 48, 178–184. [Google Scholar]
- Kim, K.J.; Choi, J.S.; Kang, I.; Kim, K.W.; Jeong, C.H.; Jeong, J.W. Melatonin suppresses tumor progression by reducing angiogenesis stimulated by HIF-1 in a mouse tumor model. J. Pineal Res. 2012. [Google Scholar] [CrossRef]
- Karbownik, M. Potential anticarcinogenic action of melatonin and other antioxidants mediated by antioxidative mechanisms. Neuro Endocrinol. Lett 2002, 23, 39–44. [Google Scholar]
- Vijayalaxmi; Reiter, R.J.; Tan, D.X.; Herman, T.S.; Thomas, C.R., Jr. Melatonin as a radioprotective agent: A review. Int. J. Radiat. Oncol. Biol. Phys. 2004, 59, 639–653. [Google Scholar]
- Kadoma, Y.; Fujisawa, S. Radical-scavenging activity of melatonin, either alone or in combination with vitamin E, ascorbate or 2-mercaptoethanol as co-antioxidants, using the induction period method. In Vivo 2011, 25, 49–53. [Google Scholar]
- Galano, A. On the direct scavenging activity of melatonin towards hydroxyl and a series of peroxyl radicals. Phys. Chem. Chem. Phys 2011, 13, 7178–7188. [Google Scholar]
- Anwar, M.M.; Mahfouz, H.A.; Sayed, A.S. Potential protective effects of melatonin on bone marrow of rats exposed to cytotoxic drugs. Comp. Biochem. Physiol. A 1998, 119, 493–501. [Google Scholar]
- Rapozzi, V.; Zorzet, S.; Comelli, M.; Mavelli, I.; Perissin, L.; Giraldi, T. Melatonin decreases bone marrow and lymphatic toxicity of adriamycin in mice bearing TLX5 lymphoma. Life Sci 1998, 63, 1701–1713. [Google Scholar]
- Lissoni, P.; Bastone, A.; Sala, R.; Mauri, R.; Rovelli, F.; Viviani, S.; Bajetta, E.; Esposti, D.; Esposti, G.; Di Bella, L.; et al. The clinical significance of melatonin serum determination in oncological patients and its correlations with GH and PRL blood levels. Eur. J. Cancer Clin. Oncol 1987, 23, 949–957. [Google Scholar]
- Di Bella, L.; Rossi, M.T.; Pellegrino, N.; Grimaldi, A.; Santoro, V. Ruolo dei sistema abenulo-epifisario nella regolazione del tasso-piastrinemico. Boll. Soc. It. Biol. Sper 1969, 45, 171. [Google Scholar]
- Rossi, M.T.; Scalera, G.; Di Bella, L. Azione mielotropa della melatonina (MLT). Boll. Soc. It. Biol. Sper 1976, 52, 26. [Google Scholar]
- Rossi, M.T.; Di Bella, L. Melatonin in Thrombocytogenesis. In The Pineal Gland and Cancer. Brain Research Promotion; Gupta, D., Attanasio, A., Reiter, R.J., Eds.; Brain Research Promootion: Tubingen, Germany, 1988; pp. 183–194. [Google Scholar]
- Gualano, L.; Di Bella, L.; Rossi, M.T.; Scalera, G. Effetti della melatonina sui megacariociti viventi di midollo di ratto. Boll. Soc. It. Biol. Sper 1977, 53, 44. [Google Scholar]
- Cardinali, D.P.; Del Zar, M.M.; Vacas, M.I. The effects of melatonin in human platelets. Acta Physiol. Pharmacol. Ther. Latinoam 1993, 43, 1–13. [Google Scholar]
- Di Bella, L.; Bucciarelli, M.; Pagnoni, U.M.; Scalera, G.; Rossi, M.T. Formazione di complessi tra melatonina (mlt) e basi puriniche e pirimidiniche. Boll. Soc. It. Biol. Sper 1976, 52, 157. [Google Scholar]
- Rossi, M.T.; Di Bella, L.; Scalera, G.; Gualano, L. Platelet Turnover as Influenced by Melatonin. Presented at International Symposium on Melatonin, Bremen, Germany, 28–30 September 1980.
- Jeffrey, G.A.; Saenger, W. Hydrogen Bonding in Biological Structures; Springer-Verlag: Berlin, Germany, 1991; p. 569. [Google Scholar]
- Carrillo-Vico, A.; Calvo, J.R.; Abreu, P.; Lardone, P.J.; García-Mauriño, S.; Reiter, R.J.; Guerrero, J.M. Evidence of melatonin synthesis by human lymphocytes and its physiological significance: Possible role as intracrine, autocrine, and/or paracrine substance. FASEB J 2004, 18, 537–539. [Google Scholar]
- Hill, S.M.; Spriggs, L.L.; Simon, M.A.; Muraoka, H.; Blask, D.E. The growth inhibitory action of melatonin on human breast cancer cells is linked to the estrogen response system. Cancer Lett 1992, 64, 249–256. [Google Scholar]
- Gonzalez-Haba, M.G.; Garcia-Mauriño, S.; Calvo, J.R.; Goberna, R.; Guerrero, J.M. High-affinity binding of melatonin by human circulating T lymphocytes (CD4+). FASEB J 1995, 9, 1331–1335. [Google Scholar]
- García-Pergañeda, A.; Pozo, D.; Guerrero, J.M.; Calvo, J.R. Signal transduction for melatonin in human lymphocytes: Involvement of a pertussis toxin-sensitive G protein. J. Immunol 1997, 159, 3774–3781. [Google Scholar]
- Konakchieva, R.; Kyurkchiev, S.; Kehayov, I.; Taushanova, P.; Kanchev, L. Selective effect of methoxyindoles on the lymphocyte proliferation and melatonin binding to activated human lymphoid cells. J. Neuroimmunol 1995, 63, 125–132. [Google Scholar]
- Garcia-Mauriño, S.; Gonzalez-Haba, M.G.; Calvo, J.R.; Rafii-El-Idrissi, M.; Sanchez-Margalet, V.; Goberna, R.; Guerrero, J.M. Melatonin enhances IL-2, IL-6, and IFN-gamma production by human circulating CD4+ cells: A possible nuclear receptor-mediated mechanism involving T helper type 1 lymphocytes and monocytes. J. Immunol 1997, 159, 574–581. [Google Scholar]
- Lissoni, P.; Rovelli, F.; Brivio, F.; Brivio, O.; Fumagalli, L. Circadian secretions of IL-2, IL-12, IL-6 and IL-10 in relation to the light/dark rhythm of the pineal hormone melatonin in healthy humans. Nat. Immun 1998, 16, 1–5. [Google Scholar]
- García-Mauriño, S.; Pozo, D.; Carrillo-Vico, A.; Calvo, J.R.; Guerrero, J.M. Melatonin activates Th1 lymphocytes by increasing IL-12 production. Life Sci 1999, 65, 2143–2150. [Google Scholar]
- Benítez-King, G.; Antón-Tay, F. Calmodulin mediates melatonin cytoskeletal effects. Experientia 1993, 49, 635–641. [Google Scholar]
- Soto-Vega, E.; Meza, I.; Ramírez-Rodríguez, G.; Benitez-King, G. Melatonin stimulates calmodulin phosphorylation by protein kinase C. J. Pineal Res 2004, 37, 98–106. [Google Scholar]
- Tan, D.X.; Reiter, R.J.; Manchester, L.C.; Yan, M.T.; El-Sawi, M.; Sainz, R.M.; Mayo, J.C.; Kohen, R.; Allegra, M.; Hardeland, R. Chemical and physical properties and potential mechanisms: Melatonin as a broad spectrum antioxidant and free radical scavenger. Curr. Top. Med. Chem 2002, 2, 181–197. [Google Scholar]
- Naji, L.; Carrillo-Vico, A.; Guerrero, J.M.; Calvo, J.R. Expression of membrane and nuclear melatonin receptors in mouse peripheral organs. Life Sci 2004, 74, 2227–2236. [Google Scholar]
- Girgert, R.; Hanf, V.; Emons, G.; Gründker, C. Membrane-bound melatonin receptor MT1 down-regulates estrogen responsive genes in breast cancer cells. J. Pineal Res 2009, 47, 23–31. [Google Scholar]
- Lopez-Gonzalez, M.A.; Calvo, J.R.; Osuna, C.; Rubio, A.; Guerrero, J.M. Synergistic action of melatonin and vasoactive intestinal peptide in stimulating cyclic AMP production in human lymphocytes. J. Pineal Res 1992, 12, 174–180. [Google Scholar]
- Vacas, M.I.; Del Zar, M.M.; Martinuzzo, M.; Cardinali, D.P. Binding sites for [3H]-melatonin in human platelets. J. Pineal Res 1992, 13, 60–65. [Google Scholar]
- Calvo, J.R.; Rafii-el-Idrissi, M.; Pozo, D.; Guerrero, J.M. Immunomodulatory role of melatonin: Specific binding sites in human and rodent lymphoid cells. J. Pineal Res 1995, 18, 119–126. [Google Scholar]
- Mangelsdorf, D.J.; Evans, R.M. The RXR heterodimers and orphan receptors. Cell 1995, 83, 841–850. [Google Scholar]
- Wiesenberg, I.; Missbach, M.; Kahlen, J.P.; Schräder, M.; Carlberg, C. Transcriptional activation of the nuclear receptor RZR alpha by the pineal gland hormone melatonin and identification od CGP 52608 as a synthetic ligand. Nucleic Acids Res 1995, 23, 327–233. [Google Scholar]
- Renaud, J.P.; Rochel, N.; Ruff, M.; Vivat, V.; Chambon, P.; Gronemeyer, H.; Moras, D. Crystal structure of the RAR-gamma ligand-binding domain bound to all-trans retinoic acid. Nature 1995, 378, 681–689. [Google Scholar]
- Yu, X.P.; Mocharla, H.; Hustmyer, F.G.; Manolagas, S.C. Vitamin D receptor expression in human lymphocytes. Signal requirements and characterization by western blots and DNA sequencing. J. Biol. Chem 1991, 266, 7588–7595. [Google Scholar]
- Adorini, L.; Daniel, K.C.; Penna, G. Vitamin D receptor agonists, cancer and the immune system: An intricate relationship. Curr. Top. Med. Chem 2006, 6, 1297–1301. [Google Scholar]
- Morgan, P.J.; Barrett, P.; Howell, H.E.; Helliwell, R. Melatonin receptors: Localization, molecular pharmacology and physiological significance. Neurochem. Int 1994, 24, 101–146. [Google Scholar]
- Mazzucchelli, C.; Pannacci, M.; Nonno, R.; Lucini, V.; Fraschini, F.; Stankov, B.M. The melatonin receptor in the human brain: cloning experiments and distribution studies. Brain Res. Mol. Brain Res 1996, 39, 117–126. [Google Scholar]
- Weaver, D.R.; Reppert, S.M. The Mel1a melatonin receptor gene is expressed in human suprachiasmatic nuclei. Neuroreport 1996, 8, 109–112. [Google Scholar]
- Al-Ghoul, W.M.; Herman, M.D.; Dubocovich, M.L. Melatonin receptor subtype expression in human cerebellum. Neuroreport 1998, 9, 4063–4068. [Google Scholar]
- Dubocovich, M.L.; Markowska, M. Functional MT1 and MT2 melatonin receptors in mammals. Endocrine 2005, 27, 101–110. [Google Scholar]
- Di Bella, L. Orientamenti fisiologici nella terapia delle emopatie. Bull. Sc. Med 1974, 145, 1–3. [Google Scholar]
- Di Bella, L.; Rossi, M.T.; Scalera, G. Perspectives in pineal function. Prog. Brain Res 1979, 52, 475–478. [Google Scholar]
- Kornblihtt, L.I.; Finocchiaro, L.; Molinas, F.C. Inhibitory effect of melatonin on platelet activation induced by collagen and arachidonic acid. J. Pineal Res 1993, 14, 184–191. [Google Scholar]
- Garcia-Perganeda, A.; Guerriero, J.M.; Rafii-El-Idrissi, M.; Paz Romero, M.; Pozo, D.; Calvo, J.R. Characterization of membrane melatonin receptor in mouse peritoneal macrophages: Inhibition of adenylyl cyclase by a pertussis toxin-sensitive G protein. J. Neuroimmunol 1999, 95, 85–94. [Google Scholar]
- Di Bella, L.; Bruschi, C.; Gualano, L. Melatonin effects on megakaryocyte membrane patch-clamp outward K+ current. Med. Sci. Monit. 2002, 8, BR527–531. [Google Scholar]
- Steffens, F.; Zhou, X.B.; Sausbier, U.; Sailer, C.; Motejlek, K.; Ruth, P.; Olcese, J.; Korth, M.; Wieland, T. Melatonin receptor signaling in pregnant and nonpregnant rat uterine myocytes as probed by large conductance Ca2+-activated K+ channel activity. Mol. Endocrinol 2003, 17, 2103–2115. [Google Scholar]
- Hou, S.W.; Zheng, P.; Sun, F.Y. Melatonin inhibits outward delayed rectifier potassium currents in hippocampal CA1 pyramidal neuron via intracellular indole-related domains. J. Pineal Res 2004, 36, 242–249. [Google Scholar]
- Sampson, S.R.; Lupowitz, Z.; Braiman, L.; Zisapel, N. Role of protein kinase C-alpha inmelatonin signal transduction. Mol. Cell. Endocrinol 2006, 252, 82–87. [Google Scholar]
- Martín, V.; Herrera, F.; García-Santos, G.; Antolín, I.; Rodriguez-Blanco, J.; Medina, M.; Rodriguez, C. Involvement of protein kinase C in melatonin’s oncostatic effect in C6 glioma cells. J. Pineal Res 2007, 43, 239–244. [Google Scholar]
- Quiros, I.; Mayo, J.C.; Garcia-Suarez, O.; Hevia, D.; Martin, V.; Rodríguez, C.; Sainz, R.M. Melatonin prevents glucocorticoid inhibition of cell proliferation and toxicity in hippocampal cells by reducing glucocorticoid receptor nuclear translocation. J. Steroid Biochem. Mol. Biol 2008, 110, 116–124. [Google Scholar]
- Sainz, R.M.; Mayo, J.C.; Tan, D.X.; León, J.; Manchester, L.; Reiter, R.J. Melatonin reduces prostate cancer cell growth leading to neuroendocrine differentiation via a receptor and PKA independent mechanism. Prostate 2005, 63, 29–43. [Google Scholar]
- Kvetnoi, I.M.; Raikhlin, N.T. Clinical pathology of the APUD system (apudopathy). Klin. Med. (Mosk) 1978, 56, 15–22. [Google Scholar]
- Polak, J.M.; Bloom, S.R. The diffuse neuroendocrine system. Studies of this newly discovered controlling system in health. J. Histochem. Cytochem 1979, 27, 1398–1400. [Google Scholar]
- Raikhlin, N.T.; Kvetnoi, I.M. APUD system and neuroendocrine tumors (“apudomas”). Arkh. Patol 1997, 39, 74–80. [Google Scholar]
- Maluf, H.M.; Koerner, F.C. Carcinomas of the breast with endocrine differentiation: A review. Virchows Arch 1994, 425, 449–457. [Google Scholar]
- Bonkhoff, H.; Stein, U.; Remberger, K. Endocrine-paracrine cell types in the prostate and prostatic adenocarcinoma are postmiotic cells. Hum. Pathol 1995, 26, 167–170. [Google Scholar]
- Raikhlin, N.T.; Kvetnoi, I.M. The APUD system (diffuse endocrine system) in normal and patological states. Physiol. Gen. Biol. Rev 1994, 8, 1–44. [Google Scholar]
- Sanchez-Barcelo, E.J.; Mediavilla, M.D.; Tucker, H.A. Influence of melatonin on mammary gland growth: in vivo and in vitro studies. Proc. Soc. Exp. Biol. Med 1990, 194, 103–107. [Google Scholar]
- Maestroni, G.J.; Conti, A. Melatonin in human breast cancer tissue: Association with nuclear grade and estrogen receptor status. Lab. Invest 1996, 75, 557–561. [Google Scholar]
- Del Zar, M.M.; Martinuzzo, M.; Cardinali, D.P.; Carreras, L.O.; Vacas, M.I. Diurnal variation in melatonin effect on adenosine triphosphate and serotonin release by human platelets. Acta Endocrinol. (Copenh) 1990, 123, 453–458. [Google Scholar]
- Champier, J.; Claustrat, B.; Besançon, R.; Eymin, C.; Killer, C.; Jouvet, A.; Chamba, G.; Fèvre-Montange, M. Evidence for tryptophan hydroxylase and hydroxy-indol-O-methyl-transferase mRNAs in human blood platelets. Life Sci 1997, 60, 2191–2197. [Google Scholar]
- Zucker, M.B.; Borrelli, J. Quantity, assay and release of serotonin in human platelets. J. Appl. Physiol 1955, 7, 425–431. [Google Scholar]
- Marmaras, V.J.; Mimikos, N. Enzymic formation of serotonin in mammalian blood platelets and red cells. Experientia 1971, 27, 196–197. [Google Scholar]
- Martín, F.J.; Atienza, G.; Aldegunde, M.; Míguez, J.M. Melatonin effect on serotonin uptake and release in rat platelets: Diurnal variation in responsiveness. Life Sci 1993, 53, 1079–1087. [Google Scholar]
- Ubeda, A.; Trillo, M.A.; House, D.E.; Blackman, C.F. A 50 Hz magnetic field blocks melatonin-induced enhancement of junctional transfer in normal C3H/10T1/2 cells. Carcinogenesis 1995, 16, 2945–2949. [Google Scholar]
- Kojima, T.; Mochizuki, C.; Mitaka, T.; Mochizuki, Y. Effects of melatonin on proliferation, oxidative stress and Cx32 gap junction protein expression in primary cultures of adult rat hepatocytes. Cell Struct. Funct 1997, 22, 347–356. [Google Scholar]
- Cos, S.; Fernandez, R. Melatonin effects on intercellular junctional communication in MCF-7 human breast cancer cells. J. Pineal Res 2000, 29, 166–171. [Google Scholar]
- Meléndez, J.; Maldonado, V.; Ortega, A. Effect of melatonin on beta-tubulin and MAP2 expression in NIE-115 cells. Neurochem. Res 1996, 21, 653–658. [Google Scholar]
- Benitez-King, G.; Túnez, I.; Bellon, A.; Ortíz, G.G.; Antón-Tay, F. Melatonin prevents cytoskeletalalterations and oxidative stress induced by okadaic acid in N1E-115 cells. Exp. Neurol 2003, 182, 151–159. [Google Scholar]
- Mills, E.; Wu, P.; Seely, D.; Guyatt, G. Melatonin in the treatment of cancer: A systematic review of randomized controlled trials and meta-analysis. J. Pineal Res 2005, 39, 360–366. [Google Scholar]
- Lissoni, P. Biochemotherapy with standard chemotherapies plus the pineal hormone melatonin in the treatment of advanced solid neoplasms. Pathol. Biol. (Paris) 2007, 55, 201–204. [Google Scholar]
- Sánchez-Barceló, E.J.; Mediavilla, M.D.; Tan, D.X.; Reiter, R.J. Clinical uses of melatonin: Evaluation of human trials. Curr. Med. Chem 2010, 17, 2070–2095. [Google Scholar]
- Seely, D.; Wu, P.; Fritz, H.; Kennedy, D.A.; Tsui, T.; Seely, A.J.; Mills, E. Melatonin as adjuvant cancer care with and without chemotherapy: A systematic review and meta-analysis of randomized trials. Integr. Cancer Ther 2012, 11, 293–303. [Google Scholar]
- Wang, Y.M.; Jin, B.Z.; Ai, F.; Duan, C.H.; Lu, Y.Z.; Dong, T.F.; Fu, Q.L. The efficacy and safety of melatonin in concurrent chemotherapy or radiotherapy for solid tumors: A meta-analysis of randomized controlled trials. Cancer Chemother. Pharmacol 2012, 69, 1213–1220. [Google Scholar]
- Lissoni, P.; Fumagalli, L.; Paolorossi, F.; Rovelli, F.; Roselli, M.G.; Maestroni, G.J. Anticancer neuroimmunomodulation by pineal hormones other than melatonin: Preliminary phase II study of the pineal indole 5-methoxytryptophol in association with low-dose IL-2 and melatonin. J. Biol. Regul. Homeost. Agents 1997, 11, 119–122. [Google Scholar]
- Vijayalaxmi, T.C.R., Jr; Reiter, R.J.; Herman, T.S. Melatonin: From basic research to cancer treatment clinics. J. Clin. Oncol. 2002, 20, 2575–2601. [Google Scholar]
- Bartsch, C.; Bartsch, H.; Karasek, M. Melatonin in clinical oncology. Neuro Endocrinol. Lett 2002, 23, 30–38. [Google Scholar]
- Jung, B.; Ahmad, N. Melatonin in cancer management: Progress and promise. Cancer Res 2006, 66, 9789–9793. [Google Scholar]
- Grant, S.G.; Melan, M.A.; Latimer, J.J.; Witt-Enderby, P.A. Melatonin and breast cancer: Cellular mechanisms, clinical studies and future perspectives. Expert Rev. Mol. Med 2009, 11, e5. [Google Scholar]
- Schernhammer, E.S.; Giobbie-Hurder, A.; Gantman, K.; Savoie, J.; Scheib, R.; Parker, L.M.; Chen, W.Y. A randomized controlled trial of oral melatonin supplementation and breast cancer biomarkers. Cancer Causes Control 2012, 23, 609–616. [Google Scholar]
- Di Bella, L. Physiological basis for a rational therapy of bone marrow diseases. Presented at XVIth International Congress of Hematology, Kyoto, Japan, 5–11 September 1976.
- Mediavilla, M.D.; Sanchez-Barcelo, E.J.; Tan, D.X.; Manchester, L.; Reiter, R.J. Basic mechanisms involved in the anti-cancer effects of melatonin. Curr. Med. Chem 2010, 17, 4462–4481. [Google Scholar]
- Di Bella, L.; Gualano, L.; Bruschi, C.; Minuscoli, S.; Tarozzi, G. Cytochalasin B influence on megakaryocyte patch-clamp. Adv. Exp. Med. Biol 1999, 460, 373–376. [Google Scholar]
- Lee, Y.J.; Tsai, C.H.; Hwang, J.J.; Sheu, T.; Keng, P.C. Involvement of a p53-independent and post-ttranscriptional up-regulation for p21WAF/CIP1 following destabilization of the actin cytoskeleton. Int. J. Oncol 2009, 34, 581–589. [Google Scholar]
- Goldberger, R.F.; Epstein, C.J.; Anfinsen, C.B. Purification and properties of a microsomal enzyme system catalyzing the reactivation of reduced ribonuclease and lysozyme. J. Biol. Chem 1964, 239, 1406–1410. [Google Scholar]
- Anfinsen, C.B.; Redfield, R.R. Protein structure in relation to function and biosynthesis. Adv. Protein Chem 1956, 11, 1–100. [Google Scholar]
- Anfinsen, C.B. The tertiary structure of ribonuclease. Brookhaven Symp. Biol 1962, 15, 184–198. [Google Scholar]
- Ellis, R.J. Chaperonins. Curr. Biol 1999, 9, R352. [Google Scholar]
- Ellis, R.J. Protein folding: importance of the Anfinsen cage. Curr. Biol 2003, 13, R881–R883. [Google Scholar]
- Davis, S.; Mirick, D.K. Residential magnetic fields, medication use, and the risk of breast cancer. Epidemiology 2007, 18, 266–269. [Google Scholar]
- Davis, S.; Mirick, D.K.; Chen, C.; Stanczyk, F.Z. Night shift work and hormone levels in women. Cancer Epidemiol. Biomarkers Prev 2012, 21, 609–618. [Google Scholar]
- Di Bella, G. Complete objective response to biological therapy of plurifocal breast carcinoma. Neuro. Endocrinol. Lett 2008, 29, 857–866. [Google Scholar]
- Di Bella, G. The Di Bella Method (DBM) improved survival, objective response and performance status in a retrospective observational clinical study on 122 cases of breast cancer. Neuro Endocrinol. Lett 2011, 32, 751–762. [Google Scholar]
- Di Bella, G.; Biagio, C. The Di Bella Method (DBM) improved survival, objective response and performance status in a retrospective observational clinical study on 23 tumors of the head and neck. Neuro Endocrinol Lett 2012, 33, 249–256. [Google Scholar]
- Di Bella, L. Presented at First National Conference on Melatonin: From Research to Action, Reggio Calabria, Italy, 25 January 1997.
- Di Bella, L. Cancro: Siamo Sulla Strada Giusta? Travel Factory: Roma, Italy, 1997. [Google Scholar]
- Richards, M.A.; Stockton, D.; Babb, P.; Coleman, M.P. How many deaths have been avoided through improvements in cancer survival? BMJ 2000, 320, 895–898. [Google Scholar]
- Morgan, G.; Ward, R.; Barton, M. The contribution of cytotoxic chemotherapy to 5-year survival in adult malignancies. Clin. Oncol 2004, 16, 549–560. [Google Scholar]
- Atra, A.; Gerrard, M.; Hobson, R.; Imeson, J.D.; Ashley, S.; Pinkerton, C.R. Improved cure rate in children with B-cell acute lymphoblastic leukaemia (B-ALL) and stage IV B-cell non-Hodgkin’s lymphoma (B-NHL)—Results of the UKCCSG 9003 protocol. Br. J. Cancer 1998, 77, 2281–2285. [Google Scholar]
- Ghesquières, H.; Ferlay, C.; Sebban, C.; Perol, D.; Bosly, A.; Casasnovas, O.; Reman, O.; Coiffier, B.; Tilly, H.; Morel, P.; et al. Long-term follow-up of an age-adapted C5R protocol followed by radiotherapy in 99 newly diagnosed primary CNS lymphomas: A prospective multicentric phase II study of the Groupe d’Etude des Lymphomes de l’Adulte (GELA). Ann. Oncol 2010, 21, 842–850. [Google Scholar]
- Sun, Y.; Campisi, J.; Higano, C.; Beer, T.M.; Porter, P.; Coleman, I.; True, L.; Nelson, P.S. Treatment-induced damage to the tumor microenvironment promotes prostate cancer therapy resistance through WNT16B. Nat. Med 2012, 18, 1359–1368. [Google Scholar]
- Lagadec, C.; Vlashi, E.; Della Donna, L.; Dekmezian, C.; Pajonk, F. Radiation-induced reprogramming of breast cancer cells. Stem Cells 2012, 30, 833. [Google Scholar]
- Norsa, A.; Martino, V. Somatostatin, retinoids, melatonin, vitamin D, bromocriptine, and cyclophosphamide in chemotherapy-pretreated patients with advanced lung adenocarcinoma and low performance status. Cancer Biother. Radiopharm 2007, 22, 50–55. [Google Scholar]
- Norsa, A.; Martino, V. Somatostatin, retinoids, melatonin, vitamin D, bromocriptine, and cyclophosphamide in advanced non-small-cell lung cancer patients with low performance status. Cancer Biother. Radiopharm 2006, 21, 68–73. [Google Scholar]
- Todisco, M. Chronic lymphocytic leukemia: Long-lasting remission with combination of cyclophosphamide, somatostatin, bromocriptine, retinoids, melatonin, and ACTH. Cancer Biother. Radiopharm 2009, 24, 353–355. [Google Scholar]
- Todisco, M. Low-grade non-Hodgkin lymphoma at advanced stage: a case successfully treated with cyclophosphamide plus somatostatin, bromocriptine, retinoids, and melatonin. Am. J. Ther 2007, 14, 113–115. [Google Scholar]
- Todisco, M. Relapse of high-grade non-Hodgkin’s lymphoma after autologous stem cell transplantation: A case successfully treated with cyclophosphamide plus somatostatin, bromocriptine, melatonin, retinoids, and ACTH. Am. J. Ther 2006, 13, 556–557. [Google Scholar]
- Todisco, M.; Casaccia, P.; Rossi, N. Cyclophosphamide plus somatostatin, bromocriptin, retinoids, melatonin and ACTH in the treatment of low-grade non-Hodgkin’s lymphomas at advanced stage: Results of a phase II trial. Cancer Biother. Radiopharm 2001, 16, 171–177. [Google Scholar]
- Di Bella, G. Il Metodo Di Bella; Mattioli: Parma, Italy, 2005. [Google Scholar]
© 2013 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Di Bella, G.; Mascia, F.; Gualano, L.; Di Bella, L. Melatonin Anticancer Effects: Review. Int. J. Mol. Sci. 2013, 14, 2410-2430. https://doi.org/10.3390/ijms14022410
Di Bella G, Mascia F, Gualano L, Di Bella L. Melatonin Anticancer Effects: Review. International Journal of Molecular Sciences. 2013; 14(2):2410-2430. https://doi.org/10.3390/ijms14022410
Chicago/Turabian StyleDi Bella, Giuseppe, Fabrizio Mascia, Luciano Gualano, and Luigi Di Bella. 2013. "Melatonin Anticancer Effects: Review" International Journal of Molecular Sciences 14, no. 2: 2410-2430. https://doi.org/10.3390/ijms14022410
APA StyleDi Bella, G., Mascia, F., Gualano, L., & Di Bella, L. (2013). Melatonin Anticancer Effects: Review. International Journal of Molecular Sciences, 14(2), 2410-2430. https://doi.org/10.3390/ijms14022410