Establishment of Metabolism and Transport Pathways in the Rodent and Human Fetal Liver

Abstract
:1. Introduction
2. Phase I Metabolism
2.1. Cytochrome P450 Enzymes
2.1.1. Mouse Cyp Regulation
2.1.2. Rat Cyp Regulation
2.1.3. Human CYP Regulation
2.1.4. Bile Acid-Related Cyps
2.2. Carboxylesterase Enzymes
2.3. Absolute Expression of Phase I Enzymes in Fetal and Neonatal Mouse Liver
2.4. Potential Mechanisms for the Regulation of Phase I Metabolism during Ontogeny
3. Phase II Metabolism
3.1. Sulfotransferase Enzymes
3.1.1. Mouse Sult Regulation
3.1.2. Human SULT Regulation
3.2. Glutathione S-Transferase Enzymes
3.2.1. Mouse Gst Regulation
3.2.2. Rat Gst Regulation
3.2.3. Human GST Regulation
3.3. UDP-Glucuronosyltransferase Enzymes
3.4. N-Acetyltransferase Enzymes
3.5. Absolute Phase II Enzyme Expression in Fetal and Neonatal Mouse Liver
3.6. Potential Mechanisms for the Regulation of Phase II Metabolism during Ontogeny
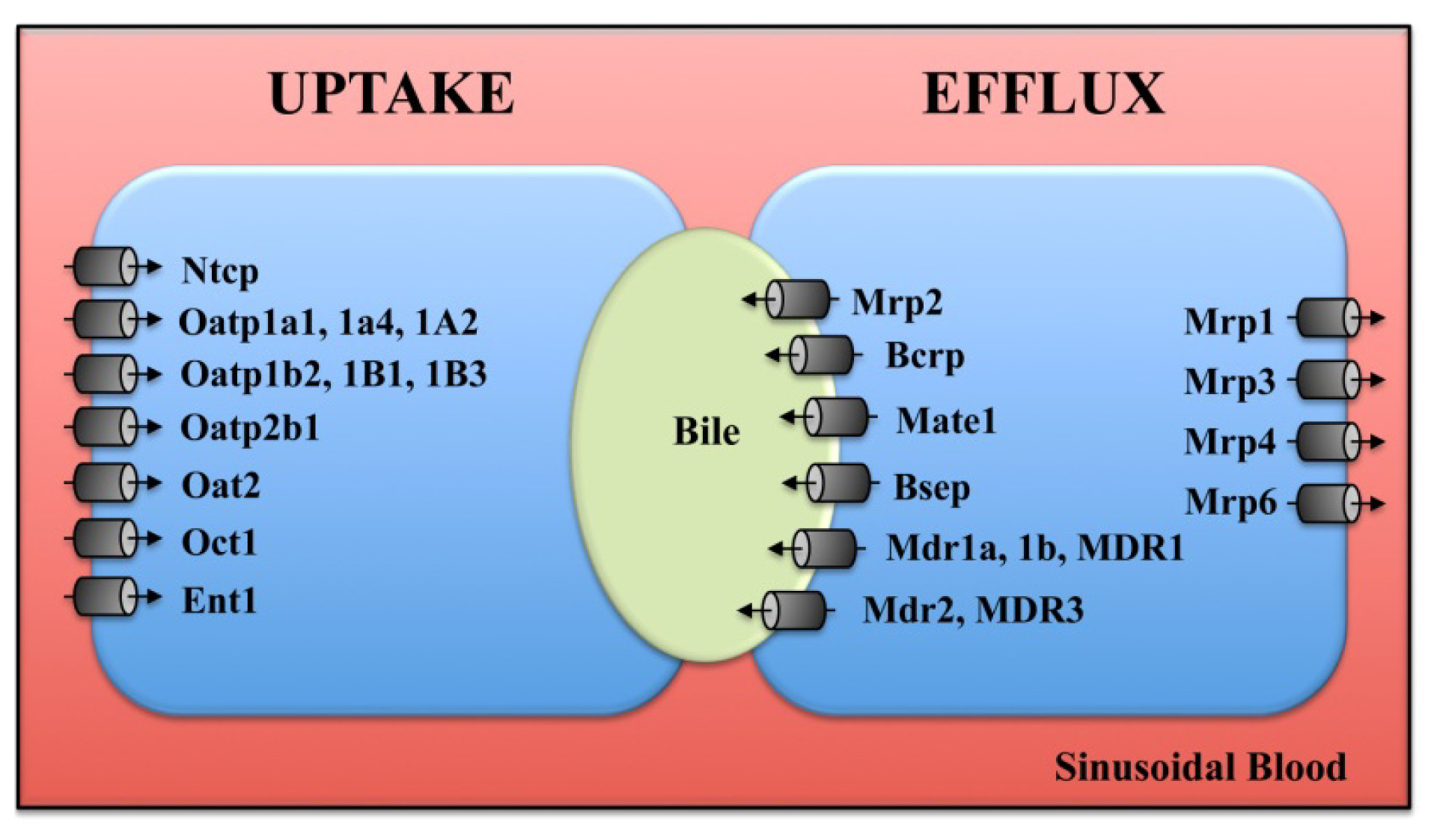
4. Transporters
4.1. Uptake Transporters
4.1.1. Mouse and Rat Uptake Transporter Regulation
4.2. Efflux Transporters
4.2.1. Rodent and Human Efflux Transporter Regulation
4.3. Bile Acid Transporters
4.3.1. Rodent and Human Bile Acid Transporter Regulation
4.4. Absolute Transporter Expression in Fetal and Neonatal Livers
4.5. Potential Ontogenic Mechanisms for Bile Acid Transporter Regulation
5. Conclusions and Future Directions
Acknowledgments
Abbreviations
| Abc | ATP-binding cassette |
| Adh | alcohol dehydrogenase |
| Akr | aldo-keto reductase |
| Aldh | aldehyde dehydrogenase |
| Bcrp | breast cancer resistance protein |
| Bsep | bile salt export pump |
| Car | constitutive androstane receptor |
| Ces | carboxylesterase |
| Cyp | cytochrome P450 |
| Ent | equilibrative nucleoside transporter |
| Fgf | fibroblast growth factor |
| Fmo | flavin-containing monooxygenase |
| Fxr | farnesoid X receptor |
| Gst | glutathione S-transferase |
| Hnf | hepatic nuclear factor |
| Lxr | liver X receptor |
| Mate | multidrug and toxin extrusion |
| Mdr | multidrug resistance |
| Mrp | multidrug resistance-associated protein |
| Nat | N-acetyl transferase |
| Ntcp | sodium taurocholate co-transporting polypeptide |
| Oat | organic anion transporter |
| Oatp | organic anion transporting polypeptide |
| Oct | organic cation transporter |
| Ost | organic solute transporter |
| Ppar | peroxisome proliferator-activated receptor |
| Pxr | pregnane X receptor |
| Slc | solute carrier |
| Sult | sulfotransferase |
| Ugt | UDP-glucuronosyl transferase |
Conflicts of Interest
References
- Fouts, J.R.; Adamson, R.H. Drug metabolism in the newborn rabbit. Science 1959, 129, 897–898. [Google Scholar]
- Alnouti, Y.; Klaassen, C.D. Tissue distribution, ontogeny, and regulation of aldehyde dehydrogenase (Aldh) enzymes mRNA by prototypical microsomal enzyme inducers in mice. Toxicol. Sci 2008, 101, 51–64. [Google Scholar]
- Peng, L.; Cui, J.Y.; Yoo, B.; Gunewardena, S.S.; Lu, H.; Klaassen, C.D.; Zhong, X.B. RNA-sequencing quantification of hepatic ontogeny of phase-I enzymes in mice. Drug Metab. Dispos 2013, 41, 2175–86. [Google Scholar]
- Cherrington, N.J.; Cao, Y.; Cherrington, J.W.; Rose, R.L.; Hodgson, E. Physiological factors affecting protein expression of flavin-containing monooxygenases 1, 3 and 5. Xenobiotica 1998, 28, 673–682. [Google Scholar]
- Dolphin, C.T.; Cullingford, T.E.; Shephard, E.A.; Smith, R.L.; Phillips, I.R. Differential developmental and tissue-specific regulation of expression of the genes encoding three members of the flavin-containing monooxygenase family of man, FMO1, FMO3 and FM04. Eur. J. Biochem 1996, 235, 683–689. [Google Scholar]
- Yeung, C.K.; Lang, D.H.; Thummel, K.E.; Rettie, A.E. Immunoquantitation of FMO1 in human liver, kidney, and intestine. Drug Metab. Dispos 2000, 28, 1107–1111. [Google Scholar]
- De Zwart, L.; Scholten, M.; Monbaliu, J.G.; Annaert, P.P.; van Houdt, J.M.; van den Wyngaert, I.; de Schaepdrijver, L.M.; Bailey, G.P.; Coogan, T.P.; Coussement, W.C.; et al. The ontogeny of drug metabolizing enzymes and transporters in the rat. Reprod. Toxicol 2008, 26, 220–230. [Google Scholar]
- Yang, D.; Pearce, R.E.; Wang, X.; Gaedigk, R.; Wan, Y.J.; Yan, B. Human carboxylesterases HCE1 and HCE2: Ontogenic expression, inter-individual variability and differential hydrolysis of oseltamivir, aspirin, deltamethrin and permethrin. Biochem. Pharmacol 2009, 77, 238–247. [Google Scholar]
- Choudhary, D.; Jansson, I.; Schenkman, J.B.; Sarfarazi, M.; Stoilov, I. Comparative expression profiling of 40 mouse cytochrome P450 genes in embryonic and adult tissues. Arch. Biochem. Biophys 2003, 414, 91–100. [Google Scholar]
- Nelson, D.R.; Zeldin, D.C.; Hoffman, S.M.; Maltais, L.J.; Wain, H.M.; Nebert, D.W. Comparison of cytochrome P450 (CYP) genes from the mouse and human genomes, including nomenclature recommendations for genes, pseudogenes and alternative-splice variants. Pharmacogenetics 2004, 14, 1–18. [Google Scholar]
- Hart, S.N.; Cui, Y.; Klaassen, C.D.; Zhong, X.B. Three patterns of cytochrome P450 gene expression during liver maturation in mice. Drug Metab. Dispos 2009, 37, 116–121. [Google Scholar]
- Sonnier, M.; Cresteil, T. Delayed ontogenesis of CYP1A2 in the human liver. Eur. J. Biochem 1998, 251, 893–898. [Google Scholar]
- Choudhary, D.; Jansson, I.; Stoilov, I.; Sarfarazi, M.; Schenkman, J.B. Expression patterns of mouse and human CYP orthologs (families 1–4 during development and in different adult tissues. Arch. Biochem. Biophys 2005, 436, 50–61. [Google Scholar]
- Peng, L.; Yoo, B.; Gunewardena, S.S.; Lu, H.; Klaassen, C.D.; Zhong, X.B. RNA sequencing reveals dynamic changes of mRNA abundance of cytochromes P450 and their alternative transcripts during mouse liver development. Drug Metab. Dispos 2012, 40, 1198–1209. [Google Scholar]
- Lee, J.S.; Ward, W.O.; Knapp, G.; Ren, H.; Vallanat, B.; Abbott, B.; Ho, K.; Karp, S.J.; Corton, J.C. Transcriptional ontogeny of the developing liver. BMC Genomics 2012, 13, 33. [Google Scholar]
- Omiecinski, C.J.; Hassett, C.; Costa, P. Developmental expression and in situ localization of the phenobarbital-inducible rat hepatic mRNAs for cytochromes CYP2B1, CYP2B2, CYP2C6, and CYP3A1. Mol. Pharmacol 1990, 38, 462–470. [Google Scholar]
- Borlakoglu, J.T.; Scott, A.; Henderson, C.J.; Wolf, C.R. Expression of P450 isoenzymes during rat liver organogenesis. Int J. Biochem 1993, 25, 1659–1668. [Google Scholar]
- Koukouritaki, S.B.; Manro, J.R.; Marsh, S.A.; Stevens, J.C.; Rettie, A.E.; McCarver, D.G.; Hines, R.N. Developmental expression of human hepatic CYP2C9 and CYP2C19. J. Pharmacol. Exp. Ther 2004, 308, 965–974. [Google Scholar]
- De Wildt, S.N.; Kearns, G.L.; Leeder, J.S.; van den Anker, J.N. Cytochrome P450 3A: Ontogeny and drug disposition. Clin. Pharmacokinet 1999, 37, 485–505. [Google Scholar]
- Lacroix, D.; Sonnier, M.; Moncion, A.; Cheron, G.; Cresteil, T. Expression of CYP3A in the human liver—Evidence that the shift between CYP3A7 and CYP3A4 occurs immediately after birth. Eur. J. Biochem 1997, 247, 625–634. [Google Scholar]
- Stevens, J.C.; Hines, R.N.; Gu, C.; Koukouritaki, S.B.; Manro, J.R.; Tandler, P.J.; Zaya, M.J. Developmental expression of the major human hepatic CYP3A enzymes. J. Pharmacol. Exp. Ther 2003, 307, 573–582. [Google Scholar]
- Cui, J.Y.; Aleksunes, L.M.; Tanaka, Y.; Fu, Z.D.; Guo, Y.; Guo, G.L.; Lu, H.; Zhong, X.B.; Klaassen, C.D. Bile acids via FXR initiate the expression of major transporters involved in the enterohepatic circulation of bile acids in newborn mice. Am. J. Physiol. Gastrointest Liver Physiol 2012, 302, G979–996. [Google Scholar]
- Cuesta de Juan, S.; Monte, M.J.; Macias, R.I.; Wauthier, V.; Calderon, P.B.; Marin, J.J. Ontogenic development-associated changes in the expression of genes involved in rat bile acid homeostasis. J. Lipid Res 2007, 48, 1362–1370. [Google Scholar]
- Li, X.Y.; Zhang, C.; Wang, H.; Ji, Y.L.; Wang, S.F.; Zhao, L.; Chen, X.; Xu, D.X. Tumor necrosis factor alpha partially contributes to lipopolysaccharide-induced downregulation of CYP3A in fetal liver: Its repression by a low dose LPS pretreatment. Toxicol. Lett 2008, 179, 71–77. [Google Scholar]
- Blanco, J.; Mulero, M.; Domingo, J.L.; Sanchez, D.J. Gestational exposure to BDE-99 produces toxicity through upregulation of CYP isoforms and ROS production in the fetal rat liver. Toxicol. Sci 2012, 127, 296–302. [Google Scholar]
- Cresteil, T.; Beaune, P.; Kremers, P.; Celier, C.; Guengerich, F.P.; Leroux, J.P. Immunoquantification of epoxide hydrolase and cytochrome P-450 isozymes in fetal and adult human liver microsomes. Eur. J. Biochem 1985, 151, 345–350. [Google Scholar]
- Shimada, T.; Yamazaki, H.; Mimura, M.; Wakamiya, N.; Ueng, Y.F.; Guengerich, F.P.; Inui, Y. Characterization of microsomal cytochrome P450 enzymes involved in the oxidation of xenobiotic chemicals in human fetal liver and adult lungs. Drug Metab. Dispos 1996, 24, 515–522. [Google Scholar]
- Treluyer, J.M.; Cheron, G.; Sonnier, M.; Cresteil, T. Cytochrome P-450 expression in sudden infant death syndrome. Biochem. Pharmacol 1996, 52, 497–504. [Google Scholar]
- Hines, R.N. Ontogeny of human hepatic cytochromes P450. J. Biochem. Mol. Toxicol 2007, 21, 169–175. [Google Scholar]
- Johnsrud, E.K.; Koukouritaki, S.B.; Divakaran, K.; Brunengraber, L.L.; Hines, R.N.; McCarver, D.G. Human hepatic CYP2E1 expression during development. J. Pharmacol. Exp. Ther 2003, 307, 402–407. [Google Scholar]
- Treluyer, J.M.; Jacqz-Aigrain, E.; Alvarez, F.; Cresteil, T. Expression of CYP2D6 in developing human liver. Eur. J. Biochem 1991, 202, 583–588. [Google Scholar]
- Cui, J.Y.; Renaud, H.J.; Klaassen, C.D. Ontogeny of novel cytochrome P450 gene isoforms during postnatal liver maturation in mice. Drug Metab. Dispos 2012, 40, 1226–1237. [Google Scholar]
- Li, Y.; Cui, Y.; Hart, S.N.; Klaassen, C.D.; Zhong, X.B. Dynamic patterns of histone methylation are associated with ontogenic expression of the Cyp3a genes during mouse liver maturation. Mol. Pharmacol 2009, 75, 1171–1179. [Google Scholar]
- Atterberry, T.T.; Burnett, W.T.; Chambers, J.E. Age-related differences in parathion and chlorpyrifos toxicity in male rats: Target and nontarget esterase sensitivity and cytochrome P450-mediated metabolism. Toxicol. Appl. Pharmacol 1997, 147, 411–418. [Google Scholar]
- Timchalk, C.; Poet, T.S.; Kousba, A.A. Age-dependent pharmacokinetic and pharmacodynamic response in preweanling rats following oral exposure to the organophosphorus insecticide chlorpyrifos. Toxicology 2006, 220, 13–25. [Google Scholar]
- Xiao, D.; Chen, Y.T.; Yang, D.; Yan, B. Age-related inducibility of carboxylesterases by the antiepileptic agent phenobarbital and implications in drug metabolism and lipid accumulation. Biochem. Pharmacol 2012, 84, 232–239. [Google Scholar]
- Zhu, H.J.; Appel, D.I.; Jiang, Y.; Markowitz, J.S. Age- and sex-related expression and activity of carboxylesterase 1 and 2 in mouse and human liver. Drug Metab. Dispos 2009, 37, 1819–1825. [Google Scholar]
- Nahar, M.S.; Kim, J.H.; Sartor, M.A.; Dolinoy, D.C. Bisphenol A-associated alterations in the expression and epigenetic regulation of genes encoding xenobiotic metabolizing enzymes in human fetal liver. Environ. Mol. Mutagen 2013, in press. [Google Scholar]
- Stoilov, I.; Akarsu, A.N.; Sarfarazi, M. Identification of three different truncating mutations in cytochrome P4501B1 (CYP1B1) as the principal cause of primary congenital glaucoma (Buphthalmos) in families linked to the GLC3A locus on chromosome 2p21. Hum. Mol. Genet 1997, 6, 641–647. [Google Scholar]
- Balasubramaniyan, N.; Shahid, M.; Suchy, F.J.; Ananthanarayanan, M. Multiple mechanisms of ontogenic regulation of nuclear receptors during rat liver development. Am. J. Physiol. Gastrointest Liver Physiol 2005, 288, G251–G260. [Google Scholar]
- Kamiya, A.; Inoue, Y.; Gonzalez, F.J. Role of the hepatocyte nuclear factor 4alpha in control of the pregnane X receptor during fetal liver development. Hepatology 2003, 37, 1375–1384. [Google Scholar]
- Klaassen, C.D.; Lu, H.; Cui, J.Y. Epigenetic regulation of drug processing genes. Toxicol. Mech. Methods 2011, 21, 312–324. [Google Scholar]
- Jin, B.; Park, D.W.; Nam, K.W.; Oh, G.T.; Lee, Y.S.; Ryu, D.Y. CpG methylation of the mouse CYP1A2 promoter. Toxicol. Lett 2004, 152, 11–18. [Google Scholar]
- Lu, H.; Gunewardena, S.; Cui, J.Y.; Yoo, B.; Zhong, X.B.; Klaassen, C.D. RNA-sequencing quantification of hepatic ontogeny and tissue distribution of mRNAs of phase II enzymes in mice. Drug Metab. Dispos 2013, 41, 844–857. [Google Scholar]
- Alnouti, Y.; Klaassen, C.D. Tissue distribution and ontogeny of sulfotransferase enzymes in mice. Toxicol. Sci 2006, 93, 242–255. [Google Scholar]
- Richard, K.; Hume, R.; Kaptein, E.; Stanley, E.L.; Visser, T.J.; Coughtrie, M.W. Sulfation of thyroid hormone and dopamine during human development: Ontogeny of phenol sulfotransferases and arylsulfatase in liver, lung, and brain. J. Clin. Endocrinol. Metab 2001, 86, 2734–2742. [Google Scholar]
- Stanley, E.L.; Hume, R.; Coughtrie, M.W. Expression profiling of human fetal cytosolic sulfotransferases involved in steroid and thyroid hormone metabolism and in detoxification. Mol. Cell. Endocrinol 2005, 240, 32–42. [Google Scholar]
- Duanmu, Z.; Weckle, A.; Koukouritaki, S.B.; Hines, R.N.; Falany, J.L.; Falany, C.N.; Kocarek, T.A.; Runge-Morris, M. Developmental expression of aryl, estrogen, and hydroxysteroid sulfotransferases in pre- and postnatal human liver. J. Pharmacol. Exp. Ther 2006, 316, 1310–1317. [Google Scholar]
- Cappiello, M.; Giuliani, L.; Rane, A.; Pacifici, G.M. Dopamine sulphotransferase is better developed than P-nitrophenol sulphotransferase in the human fetus. Dev. Pharmacol. Ther 1991, 16, 83–88. [Google Scholar]
- Pacifici, G.M.; Kubrich, M.; Giuliani, L.; de Vries, M.; Rane, A. Sulphation and glucuronidation of ritodrine in human foetal and adult tissues. Eur. J. Clin. Pharmacol 1993, 44, 259–264. [Google Scholar]
- Miki, Y.; Nakata, T.; Suzuki, T.; Darnel, A.D.; Moriya, T.; Kaneko, C.; Hidaka, K.; Shiotsu, Y.; Kusaka, H.; Sasano, H. Systemic distribution of steroid sulfatase and estrogen sulfotransferase in human adult and fetal tissues. J. Clin. Endocrinol. Metab 2002, 87, 5760–5768. [Google Scholar]
- Parker, C.R., Jr.; Falany, C.N.; Stockard, C.R.; Stankovic, A.K.; Grizzle, W.E. Immunohistochemical localization of dehydroepiandrosterone sulfotransferase in human fetal tissues. J. Clin. Endocrinol. Metab 1994, 78, 234–236. [Google Scholar]
- Barker, E.V.; Hume, R.; Hallas, A.; Coughtrie, W.H. Dehydroepiandrosterone sulfotransferase in the developing human fetus: Quantitative biochemical and immunological characterization of the hepatic, renal, and adrenal enzymes. Endocrinology 1994, 134, 982–989. [Google Scholar]
- Cui, J.Y.; Choudhuri, S.; Knight, T.R.; Klaassen, C.D. Genetic and epigenetic regulation and expression signatures of glutathione S-transferases in developing mouse liver. Toxicol. Sci 2010, 116, 32–43. [Google Scholar]
- Tee, L.B.; Gilmore, K.S.; Meyer, D.J.; Ketterer, B.; Vandenberghe, Y.; Yeoh, G.C. Expression of glutathione S-transferase during rat liver development. Biochem. J 1992, 282, 209–218. [Google Scholar]
- Raijmakers, M.T.; Steegers, E.A.; Peters, W.H. Glutathione S-transferases and thiol concentrations in embryonic and early fetal tissues. Hum. Reprod 2001, 16, 2445–2450. [Google Scholar]
- Strange, R.C.; Davis, B.A.; Faulder, C.G.; Cotton, W.; Bain, A.D.; Hopkinson, D.A.; Hume, R. The human glutathione S-transferases: Developmental aspects of the GST1, GST2, and GST3 loci. Biochem Genet 1985, 23, 1011–1028. [Google Scholar]
- Kishi, M.; Emi, Y.; Sakaguchi, M.; Ikushiro, S.; Iyanagi, T. Ontogenic isoform switching of UDP-glucuronosyltransferase family 1 in rat liver. Biochem. Biophys. Res. Commun 2008, 377, 815–819. [Google Scholar]
- Burchell, B.; Coughtrie, M.; Jackson, M.; Harding, D.; Fournel-Gigleux, S.; Leakey, J.; Hume, R. Development of human liver UDP-glucuronosyltransferases. Dev. Pharmacol. Ther 1989, 13, 70–77. [Google Scholar]
- Rollins, D.E.; von Bahr, C.; Glaumann, H.; Moldeus, P.; Rane, A. Acetaminophen: Potentially toxic metabolite formed by human fetal and adult liver microsomes and isolated fetal liver cells. Science 1979, 205, 1414–1416. [Google Scholar]
- Ekstrom, L.; Johansson, M.; Rane, A. Tissue distribution and relative gene expression of UDP-glucuronosyltransferases (2B7, 2B15, 2B17) in the human fetus. Drug Metab. Dispos 2013, 41, 291–295. [Google Scholar]
- Pacifici, G.M.; Sawe, J.; Kager, L.; Rane, A. Morphine glucuronidation in human fetal and adult liver. Eur. J. Clin. Pharmacol 1982, 22, 553–558. [Google Scholar]
- Leakey, J.E.; Hume, R.; Burchell, B. Development of multiple activities of UDP-glucuronyltransferase in human liver. Biochem. J 1987, 243, 859–861. [Google Scholar]
- McQueen, C.A.; Mitchell, M.K.; Dang, L.N.; Chau, B.; Tjalkens, R.B.; Philbert, M.A. Prenatal expression of N-acetyltransferases in C57Bl/6 mice. Chem. Biol. Interact 2003, 145, 77–87. [Google Scholar]
- Pacifici, G.M.; Bencini, C.; Rane, A. Acetyltransferase in humans: Development and tissue distribution. Pharmacology 1986, 32, 283–291. [Google Scholar]
- Coughtrie, M.W. Sulfation through the looking glass—Recent advances in sulfotransferase research for the curious. Pharmacogenomics J 2002, 2, 297–308. [Google Scholar]
- Her, C.; Kaur, G.P.; Athwal, R.S.; Weinshilboum, R.M. Human sulfotransferase SULT1C1: cDNA cloning, tissue-specific expression, and chromosomal localization. Genomics 1997, 41, 467–470. [Google Scholar]
- Hobkirk, R. Steroid sulfotransferases and steroid sulfate sulfatases: Characteristics and biological roles. Can. J. Biochem Cell. Biol 1985, 63, 1127–1144. [Google Scholar]
- Knight, T.R.; Choudhuri, S.; Klaassen, C.D. Constitutive mRNA expression of various glutathione S-transferase isoforms in different tissues of mice. Toxicol. Sci 2007, 100, 513–524. [Google Scholar]
- Thomson, R.E.; Bigley, A.L.; Foster, J.R.; Jowsey, I.R.; Elcombe, C.R.; Orton, T.C.; Hayes, J.D. Tissue-specific expression and subcellular distribution of murine glutathione S-transferase class kappa. J. Histochem. Cytochem 2004, 52, 653–662. [Google Scholar]
- Li, W.; Gu, Y.; James, M.O.; Hines, R.N.; Simpson, P.; Langaee, T.; Stacpoole, P.W. Prenatal and postnatal expression of glutathione transferase zeta 1 in human liver and the roles of haplotype and subject age in determining activity with dichloroacetate. Drug Metab. Dispos 2012, 40, 232–239. [Google Scholar]
- Evans, W.E.; Relling, M.V. Pharmacogenomics: Translating functional genomics into rational therapeutics. Science 1999, 286, 487–491. [Google Scholar]
- Mitchell, M.K.; Futscher, B.W.; McQueen, C.A. Developmental expression of N-acetyltransferases in C57BI/6 mice. Drug Metab. Dispos 1999, 27, 261–264. [Google Scholar]
- Araki, Y.; Sakakibara, Y.; Boggaram, V.; Katafuchi, J.; Suiko, M.; Nakajima, H.; Liu, M.C. Tissue-specific and developmental stage-dependent expression of a novel rat Dopa/tyrosine sulfotransferase. Int. J. Biochem. Cell. Biol 1997, 29, 801–806. [Google Scholar]
- Liu, L.; Klaassen, C.D. Ontogeny and hormonal basis of female-dominant rat hepatic sulfotransferases. J. Pharmacol. Exp. Ther 1996, 279, 386–391. [Google Scholar]
- Liu, L.; Klaassen, C.D. Ontogeny and hormonal basis of male-dominant rat hepatic sulfotransferases. Mol. Pharmacol 1996, 50, 565–572. [Google Scholar]
- Sonoda, J.; Xie, W.; Rosenfeld, J.M.; Barwick, J.L.; Guzelian, P.S.; Evans, R.M. Regulation of a xenobiotic sulfonation cascade by nuclear pregnane X receptor (Pxr). Proc. Natl. Acad. Sci. USA 2002, 99, 13801–13806. [Google Scholar]
- Fang, H.L.; Strom, S.C.; Cai, H.; Falany, C.N.; Kocarek, T.A.; Runge-Morris, M. Regulation of human hepatic hydroxysteroid sulfotransferase gene expression by the peroxisome proliferator-activated receptor alpha transcription factor. Mol. Pharmacol 2005, 67, 1257–1267. [Google Scholar]
- Hempel, N.; Wang, H.; LeCluyse, E.L.; McManus, M.E.; Negishi, M. The human sulfotransferase SULT1A1 gene is regulated in a synergistic manner by Sp1 and GA binding protein. Mol. Pharmacol 2004, 66, 1690–1701. [Google Scholar]
- Cui, J.Y.; Gunewardena, S.S.; Yoo, B.; Liu, J.; Renaud, H.J.; Lu, H.; Zhong, X.B.; Klaassen, C.D. RNA-Seq reveals different mRNA abundance of transporters and their alternative transcript isoforms during liver development. Toxicol. Sci 2012, 127, 592–608. [Google Scholar]
- Gao, B.; St Pierre, M.V.; Stieger, B.; Meier, P.J. Differential expression of bile salt and organic anion transporters in developing rat liver. J. Hepatol 2004, 41, 201–208. [Google Scholar]
- Chen, H.L.; Liu, Y.J.; Feng, C.H.; Wu, C.Y.; Shyu, M.K.; Yuan, R.H.; Chang, M.H. Developmental expression of canalicular transporter genes in human liver. J. Hepatol 2005, 43, 472–477. [Google Scholar]
- Sharma, S.; Ellis, E.C.; Gramignoli, R.; Dorko, K.; Tahan, V.; Hansel, M.; Mattison, D.R.; Caritis, S.N.; Hines, R.N.; Venkataramanan, R.; et al. Hepatobiliary disposition of 17-OHPC and taurocholate in fetal human hepatocytes: A comparison with adult human hepatocytes. Drug Metab. Dispos 2013, 41, 296–304. [Google Scholar]
- Hardikar, W.; Ananthanarayanan, M.; Suchy, F.J. Differential ontogenic regulation of basolateral and canalicular bile acid transport proteins in rat liver. J. Biol. Chem 1995, 270, 20841–20846. [Google Scholar]
- St-Pierre, M.V.; Stallmach, T.; Freimoser Grundschober, A.; Dufour, J.F.; Serrano, M.A.; Marin, J.J.; Sugiyama, Y.; Meier, P.J. Temporal expression profiles of organic anion transport proteins in placenta and fetal liver of the rat. Am. J. Physiol. Regul. Integr. Comp. Physiol 2004, 287, R1505–R1516. [Google Scholar]
- Alnouti, Y.; Petrick, J.S.; Klaassen, C.D. Tissue distribution and ontogeny of organic cation transporters in mice. Drug Metab. Dispos 2006, 34, 477–482. [Google Scholar]
- Cheng, X.; Maher, J.; Chen, C.; Klaassen, C.D. Tissue distribution and ontogeny of mouse organic anion transporting polypeptides (Oatps). Drug Metab. Dispos 2005, 33, 1062–1073. [Google Scholar]
- Van Kalken, C.K.; Giaccone, G.; van der Valk, P.; Kuiper, C.M.; Hadisaputro, M.M.; Bosma, S.A.; Scheper, R.J.; Meijer, C.J.; Pinedo, H.M. Multidrug resistance gene (P-glycoprotein) expression in the human fetus. Am. J. Pathol 1992, 141, 1063–1072. [Google Scholar]
- Rosati, A.; Maniori, S.; Decorti, G.; Candussio, L.; Giraldi, T.; Bartoli, F. Physiological regulation of P-glycoprotein, MRP1, MRP2 and cytochrome P450 3A2 during rat ontogeny. Dev. Growth Differ 2003, 45, 377–387. [Google Scholar]
- Cui, Y.J.; Cheng, X.; Weaver, Y.M.; Klaassen, C.D. Tissue distribution, gender-divergent expression, ontogeny, and chemical induction of multidrug resistance transporter genes (Mdr1a, Mdr1b, Mdr2) in mice. Drug Metab. Dispos 2009, 37, 203–210. [Google Scholar]
- Zinchuk, V.S.; Okada, T.; Akimaru, K.; Seguchi, H. Asynchronous expression and colocalization of Bsep and Mrp2 during development of rat liver. Am. J. Physiol. Gastrointest Liver Physiol 2002, 282, G540–G548. [Google Scholar]
- Maher, J.M.; Slitt, A.L.; Cherrington, N.J.; Cheng, X.; Klaassen, C.D. Tissue distribution and hepatic and renal ontogeny of the multidrug resistance-associated protein (Mrp) family in mice. Drug Metab. Dispos 2005, 33, 947–955. [Google Scholar]
- Dubuisson, C.; Cresteil, D.; Desrochers, M.; Decimo, D.; Hadchouel, M.; Jacquemin, E. Ontogenic expression of the Na(+)-independent organic anion transporting polypeptide (oatp) in rat liver and kidney. J. Hepatol 1996, 25, 932–940. [Google Scholar]
- Lee, J.S.; Ward, W.O.; Liu, J.; Ren, H.; Vallanat, B.; Delker, D.; Corton, J.C. Hepatic xenobiotic metabolizing enzyme and transporter gene expression through the life stages of the mouse. PLoS One 2011, 6, e24381. [Google Scholar]
- Fakhoury, M.; de Beaumais, T.; Guimiot, F.; Azougagh, S.; Elie, V.; Medard, Y.; Delezoide, A.L.; Jacqz-Aigrain, E. mRNA expression of MDR1 and major metabolising enzymes in human fetal tissues. Drug Metab. Pharmacokinet 2009, 24, 529–536. [Google Scholar]
- Klaassen, C.D.; Aleksunes, L.M. Xenobiotic, bile acid, and cholesterol transporters: Function and regulation. Pharmacol. Rev 2010, 62, 1–96. [Google Scholar]
- Tomer, G.; Ananthanarayanan, M.; Weymann, A.; Balasubramanian, N.; Suchy, F.J. Differential developmental regulation of rat liver canalicular membrane transporters Bsep and Mrp2. Pediatr. Res 2003, 53, 288–294. [Google Scholar]
- Zhou, S.; Zong, Y.; Ney, P.A.; Nair, G.; Stewart, C.F.; Sorrentino, B.P. Increased expression of the Abcg2 transporter during erythroid maturation plays a role in decreasing cellular protoporphyrin IX levels. Blood 2005, 105, 2571–2576. [Google Scholar]
- Boyer, J.L.; Hagenbuch, B.; Ananthanarayanan, M.; Suchy, F.; Stieger, B.; Meier, P.J. Phylogenic and ontogenic expression of hepatocellular bile acid transport. Proc. Natl. Acad. Sci. USA 1993, 90, 435–438. [Google Scholar]
- Suchy, F.J.; Bucuvalas, J.C.; Goodrich, A.L.; Moyer, M.S.; Blitzer, B.L. Taurocholate transport and Na+-K+-ATPase activity in fetal and neonatal rat liver plasma membrane vesicles. Am. J. Physiol 1986, 251, G665–G673. [Google Scholar]
- Landry, C.; Clotman, F.; Hioki, T.; Oda, H.; Picard, J.J.; Lemaigre, F.P.; Rousseau, G.G. HNF-6 is expressed in endoderm derivatives and nervous system of the mouse embryo and participates to the cross-regulatory network of liver-enriched transcription factors. Dev. Biol 1997, 192, 247–257. [Google Scholar]



{kind=link}
{kind=link}
| Enzyme | Mouse | Rat | Human | References |
|---|---|---|---|---|
| Alcohol Dehydrogenases | ||||
| Adh1 | ↓(M) | [3] | ||
| Adh4 | ↓(M) | [3] | ||
| Adh5 | ↓(M) | [3] | ||
| Adh6-ps1 | ↓(M) | [3] | ||
| Adh7 | ↓(M) | [3] | ||
| Aldo-Keto Reductases | ||||
| Akr1 | ↓(M) | [3] | ||
| Akr7 | ↓(M) | [3] | ||
| Aldehyde Dehydrogenases | ||||
| Aldh1 | ↓(M) | [2,3] | ||
| Aldh2 | ↓(M) | [2,3] | ||
| Aldh3a2 | ↓(M) | [2,3] | ||
| Aldh4a1 | ↓(M) | [2,3] | ||
| Aldh6a1 | ↓(M) | [2,3] | ||
| Aldh7a1 | ↓(M) | [2,3] | ||
| Carboxylesterases | ||||
| Ces | ↓(A) | [7] | ||
| Ces1/CES1 | ↓(M) | ↓(M,P,A) | [3,8] | |
| Ces2/CES2 | ↓(M) | ↓(M) | [3,8] | |
| Ces3 | ↓(M) | [3] | ||
| Flavin-Containing Monooxygenases | ||||
| Fmo1/FMO1 | ↓(M)↔(P) | ↑(M,P) | [3–6] | |
| Fmo2 | ↓(M) | [3] | ||
| Fmo3/FMO3 | ↓(M,P) | ↓(M) | [3–5] | |
| Fmo4/FMO4 | ↔(M) | ↔(M) | [3,5] | |
| Fmo5 | ↓(M,P) | [3,4] | ||
| Enzyme | Mouse | Rat | Human | References |
|---|---|---|---|---|
| Cytochrome P450s | ||||
| Cyp1a1/CYP1A1 | ↑(M)↓(A) | ↓(M,P,A) | [7,12,13] | |
| Cyp1a2/CYP1A2 | ↓(M) | ↓(M,A) | ↓(M,P,A) | [7,11–15] |
| Cyp1b1/CYP1B1 | ↓(M) | ↑(M) | [11,13] | |
| Cyp2a4 | ↓(M) | [11,14,15] | ||
| Cyp2a5 | ↓(M) | [14,15] | ||
| Cyp2a12 | ↓(M) | [14,15] | ||
| Cyp2a22 | ↓(M) | [14] | ||
| Cyp2b1 | ↓(M,A) | [7,16,17] | ||
| Cyp2b2 | ↓(M,A) | [7,16] | ||
| Cyp2b9 | ↓(M) | [14,15] | ||
| Cyp2b10 | ↓(M) | [11,14,15] | ||
| Cyp2b13 | ↓(M) | [14,15] | ||
| Cyp2b23 | ↓(M) | [14] | ||
| Cyp2c6 | ↓(M) | [16] | ||
| Cyp2c7 | ↓(M) | [17] | ||
| CYP2C9 | ↓(P,A) | [18] | ||
| CYP2C19 | ↓(P,A) | [18] | ||
| Cyp2c29 | ↓(M) | [14,15] | ||
| Cyp2c37 | ↓(M) | [14] | ||
| Cyp2c38 | ↓↔(M) | [14,15] | ||
| Cyp2c39 | ↓↔(M) | [14,15] | ||
| Cyp2c40 | ↓(M) | [14] | ||
| Cyp2c44 | ↓(M) | [14,15] | ||
| Cyp2c50 | ↓(M) | [14,15] | ||
| Cyp2c54 | ↓(M) | [14,15] | ||
| Cyp2c55 | ↓(M) | [14] | ||
| Cyp2c66 | ↓(M) | [11] | ||
| Cyp2c67 | ↓(M) | [14] | ||
| Cyp2c68 | ↓(M) | [14,15] | ||
| Cyp2c69 | ↓(M) | [14] | ||
| Cyp2c70 | ↓(M) | [14,15] | ||
| Cyp2d9 | ↓(M) | [14,15] | ||
| Cyp2d10 | ↓(M) | [14,15] | ||
| Cyp2d11 | ↓(M) | [14] | ||
| Cyp2d12 | ↓(M) | [14] | ||
| Cyp2d13 | ↓↔(M) | [14,15] | ||
| Cyp2d22 | ↓(M) | [11,14,15] | ||
| Cyp2d26 | ↓(M) | [14,15] | ||
| Cyp2d34 | ↓(M) | [14] | ||
| Cyp2d37-ps | ↓(M) | [14] | ||
| Cyp2d40 | ↓(M) | [14] | ||
| Cyp2e1/CYP2E1 | ↓(M) | ↓(M,A) | ↓(M) | [7,11,13–15] |
| Cyp2f2 | ↓(M) | [11,14,15] | ||
| Cyp2g1 | ↓↔(M) | [14,15] | ||
| Cyp2j5 | ↓(M) | [14,15] | ||
| Cyp2j6 | ↓(M) | [11,14] | ||
| Cyp2r1/CYP2R1 | ↓(M) | ↓(M) | [13,14] | |
| Cyp2s1/CYP2S1 | ↓(M) | [13] | ||
| Cyp2u1/CYP2U1 | ↓(M) | ↓(M) | [13,14] | |
| Cyp2w1/CYP2W1 | ↑(M) | [13] | ||
| Cyp3a1 | ↓(M,A) | [7,16,17] | ||
| Cyp3a2 | ↓(♂M,A)↑(♀M) | [7] | ||
| Cyp3a11/CYP3A4 | ↓(M) | ↓(M) | ↓(M,P,A) | [11,14,15,19–24] |
| CYP3A5 | ↔(M,P,A) | [19,21] | ||
| Cyp3a13 | ↓(M) | [11,14,15] | ||
| Cyp3a16/CYP3A7 | ↑↔(M) | ↑(M,P,A) | [11,14,15,19–21] | |
| Cyp3a25 | ↓(M) | [11,14,15] | ||
| Cyp3a41a/b | ↑(♂M) ↓(♀M) | [11,14,15] | ||
| Cyp3a44 | ↓(M) | [14,15] | ||
| Cyp3a59 | ↓(M) | [14] | ||
| Cyp4a1 | ↔(M)↓(♂A)↑(♀A) | [7] | ||
| Cyp4a10 | ↓(M) | [11,14,15] | ||
| Cyp4a12a/b | ↓(M) | [14] | ||
| Cyp4a14 | ↓↔(M) | [14,15] | ||
| Cyp4a31 | ↓(M) | [14,15] | ||
| Cyp4a32 | ↓(M) | [14] | ||
| Cyp4b1/CYP4B1 | ↓(M) | ↓(M) | [13,14] | |
| Cyp4f13 | ↓↔(M) | [14,15] | ||
| Cyp4f14 | ↓(M) | [14,15] | ||
| Cyp4f15 | ↓(M) | [14,15] | ||
| Cyp4f16 | ↓(M) | [14] | ||
| Cyp4f18 | ↓↔(M) | [11,14] | ||
| Cyp4f39 | ↓(M) | [14] | ||
| Cyp4v3 | ↓(M) | [14,15] | ||
| Cyp4x1/CYP4X1 | ↓(M) | ↓(M) | [13] | |
| Cyp7a1 | ↓↔(M) | ↓(M) | [14,15,22,23] | |
| Cyp7b1 | ↓↔(M) | [14,15,22] | ||
| Cyp8b1 | ↓(M) | ↓(M) | [14,15,22,23] | |
| Cyp17a1 | ↓(M) | [14] | ||
| Cyp20a1 | ↓(M) | [14] | ||
| Cyp26a1 | ↓(M) | [14,15] | ||
| Cyp26b1 | ↓(M) | [14] | ||
| Cyp27 | ↓(M) | [23] | ||
| Cyp27a1 | ↓(M) | [14,15,22] | ||
| Cyp39a1 | ↓(M) | [14] | ||
| Cyp51 | ↓↔(M) | [14,15] | ||
| Enzyme | Mouse | Rat | Human | References |
|---|---|---|---|---|
| Sulfotransferases | ||||
| Sult1a1/SULT1A1 | ↓↔(M) | ↔↓(P)↓(A) | [15,44–49] | |
| SULT1A3 | ↑(P,A) | [46,47,49,50] | ||
| Sult1b1 | ↓(♂M) | [44] | ||
| Sult1c1/SULT1C1 | ↑(M) | [45] | ||
| Sult1c2/SULT1C2 | ↓(M) | ↑(P) | [44,45,47] | |
| Sult1d1 | ↓↔(M) | [15,44,45] | ||
| Sult1e1/SULT1E1 | ↔(♂M) | ↑(M,P)↔(A) | [44,47,48,51] | |
| Sult2a1/SULT2A1 | ↓(M) | ↓(P,A) | [44,45,47,48,52,53] | |
| Sult3a1 | ↓(♀M)↔(♂M) | [15,45] | ||
| Papss2 | ↓(♀M)↔(♂M) | [45] | ||
| GlutathioneS-transferases | ||||
| Gsta1/2/GSTA1/2 | ↓(M) | ↓(P) | ↓(P) | [15,44,54–57] |
| Gsta3 | ↓(M) | [15,44,54] | ||
| Gsta4 | ↓(M) | [44,54] | ||
| Gsta8 | ↓(P) | [55] | ||
| Gsta10 | ↑(P) | [55] | ||
| Gstk1 | ↓(M) | [15,54] | ||
| Gstm1/GSTM1 | ↓↔(M) | ↓(P) | ↑(P) | [15,44,54–56] |
| Gstm2 | ↓(M) | [15,44,54] | ||
| Gstm3 | ↓↔(M) | ↓(P) | [15,44,54,55] | |
| Gstm4 | ↓(M) | ↓(P) | [15,44,54,55] | |
| Gstm5 | ↑(M) | [15,44,54] | ||
| Gstm6 | ↓↔(M) | [15,44,54] | ||
| Gstm7 | ↓(♂M) | [44] | ||
| Gsto1 | ↓(M) | [44,54] | ||
| Gstp1/2/GSTP1 | ↓(M) | ↑(P) | [15,44,54,56,57] | |
| Gstp7 | ↑(P) | [55] | ||
| Gstt1 | ↓↔(M) | [15,44,54] | ||
| Gstt2 | ↔(M) | [15,44,54] | ||
| Gstt3 | ↓↔(M) | [15,44,54] | ||
| Gstz1/GSTZ1 | ↓(M) | ↓(P,A) | [15,44,54,56] | |
| Mgst1 | ↓(M) | [15,44] | ||
| Mgst2 | ↑↔(M) | [44,54] | ||
| Mgst3 | ↓↑(M) | [15,44,54] | ||
| UDP Glucuronosyltransferases | ||||
| Ugt1a1/UGT1A1 | ↓(♂M) | ↓(M,P,A) | ↓(A) | [44,58,59] |
| UGT1A3 | ↓(A) | [59] | ||
| Ugt1a5 | ↓(♂M) | ↓(M,P) | [44,58] | |
| Ugt1a6/UGT1A6 | ↓(♂M) | ↑(M,P) | ↓(A) | [44,58,60] |
| Ugt1a9 | ↓(♂M) | [44] | ||
| Ugt2a3 | ↓(M) | [15,44] | ||
| Ugt2b1 | ↓(M) | [15,44] | ||
| Ugt2b5 | ↓(M) | [15,44] | ||
| UGT2B7 | ↓(M,A) | [61,62] | ||
| UGT2B15 | ↓(M) | [61] | ||
| UGT2B17 | ↓(M,A) | [61,63] | ||
| Ugt2b34 | ↓↔(M) | [15,44] | ||
| Ugt2b35 | ↓(M) | [15,44] | ||
| Ugt2b36 | ↓(M) | [15,44] | ||
| Ugt2b37 | ↓↔(M) | [15,44] | ||
| Ugt2b38 | ↓(♂M) | [44] | ||
| Ugt3a1 | ↓↔(M) | [15,44] | ||
| Ugt3a2 | ↓(M) | [15,44] | ||
| N-Acetyltransferases | ||||
| Nat1/NAT1 | ↓(M,A) | ↔(A) | [44,64,65] | |
| Nat2 | ↓(M,A) | [44,64] | ||
| Nat8 | ↓(♂M) | [44] | ||
| Transporter | Mouse | Rat | Human | References |
|---|---|---|---|---|
| Uptake Transporters | ||||
| Slc1a5 | ↑(M) | [15] | ||
| Slc2a1 | ↑(M) | [15] | ||
| Slc2a3 | ↑(M) | [15] | ||
| Slc3a2 | ↑(M) | [15] | ||
| Slc4a1 | ↑(M) | [15] | ||
| Slc6a9 | ↔(M) | [15] | ||
| Slc7a1 | ↑↔(M) | [15] | ||
| Slc7a5 | ↑(M) | [15] | ||
| Slc10a1/Ntcp/NTCP | ↓(M,P) | ↓(M,P) | ↓(M) | [22,80–85] |
| Slc10a2/Asbt | ↓(M) | [80] | ||
| Slc14a1 | ↑(M) | [15] | ||
| Slc16a1 | ↑(M) | [15] | ||
| Slc17a1/Npt1 | ↓(M) | [80] | ||
| Slc20a1 | ↑(M) | [15] | ||
| Slc22a1/Oct1 | ↓(M) | ↓(M) | [7,22,80,86] | |
| Slc22a2/Oct2 | ↑(♀M)↓(♂M) | [7] | ||
| Slc22a4/Octn1 | ↑(M) | [80] | ||
| Slc22a5/Octn2 | ↔(M) | [80] | ||
| Slc22a6/Oat1 | ↓(M) | [7] | ||
| Slc22a7/Oat2 | ↓(M) | ↓(M) | [7,22,80,85] | |
| Slc22a8/Oat3 | ↑(M) | ↑(M) | [7,80] | |
| Slc25a4 | ↑(M) | [15] | ||
| Slc25a37 | ↑(M) | [15] | ||
| Slc25a38 | ↔(M) | [15] | ||
| Slc29a1/Ent1 | ↔(M) | [22,80] | ||
| Slc29a3/Ent3 | ↔(M) | [80] | ||
| Slc38a1 | ↑(M) | [15] | ||
| Slc38a5 | ↑(M) | [15] | ||
| Slc39a5 | ↑(M) | [15] | ||
| Slc39a8 | ↔(M) | [15] | ||
| Slc43a1 | ↑(M) | [15] | ||
| Slc43a3 | ↑(M) | [15] | ||
| Slc47a1/Mate1 | ↓(M) | [22,80] | ||
| Slc51a/Ostα | ↔(M) | [22] | ||
| Slc51b/Ostβ | ↔(M) | [22,80] | ||
| Slco1a1/Oatp1a1 | ↓(M) | ↓(M,♂P) | [22,80,81,85,87] | |
| Slco1a4/Oatp1a4 | ↓↔(M) | ↑↓(M)↓(♂P) | [7,22,80,81,87] | |
| Slco1a6/Oatp1a6 | ↔(M) | [87] | ||
| SLCO1B1/OATP1B1 | ↓(M) | [83] | ||
| Slco1b2/Oatp1b2 | ↓(M) | ↓(M,♂P) | [22,80,81,85,87] | |
| SLCO1B3/OATP1B3 | ↓(M) | [83] | ||
| Slco4a1/Oatp4a1 | ↓(M) | [85] | ||
| Slco2a1/Oatp2a1 | ↓↔(M) | [80,87] | ||
| Slco2b1/Oatp2b1/2B1 | ↓(M) | ↓(M) | ↓(M) | [22,80,83,85,87] |
| Efflux Transporters | ||||
| Abca1 | ↑↔(M) | [22,80] | ||
| ABCB1/MDR1 | ↓(M,P) | [83,88] | ||
| Abcb1a/Mdr1a | ↓↔(M) | [7,89] | ||
| Abcb1b/Mdr1b | ↔(M) | ↓(M) | [7,89,90] | |
| Abcb4/Mdr2/MDR3 | ↓↔(M) | ↓(M,P) | [22,80,82,83,90] | |
| Abcb10 | ↑(M) | [15] | ||
| Abcb11/Bsep/BSEP | ↓↔(M)↓(P) | ↑↓(M,P) | ↓(M,P) | [7,22,80–83,85,91] |
| Abcc1/Mrp1 | ↑(M) | ↑(M) | [7,80,85,89] | |
| Abcc2/Mrp2/MRP2 | ↓(M) | ↓↔(M,P) | ↓(M,P) | [7,22,80–83,85,89,91,92] |
| Abcc3/Mrp3/MRP3 | ↓(M) | ↑↔(M) | ↓(M) | [7,22,80,83,85,92] |
| Abcc4/Mrp4/MRP4 | ↑↔(M) | ↔(M) | [22,80,83,92] | |
| Abcc5/Mrp5 | ↑(M) | [80] | ||
| Abcc6/Mrp6 | ↓(M) | ↑↓(M)↓(♂P) | [7,22,80,81,85,92] | |
| Abcg2/Bcrp/BCRP | ↑↔(M) | ↑(M) | ↓↔(M) | [7,15,22,80,83] |
| Abcg5 | ↓↔(M) | [22,80] | ||
| Abcg8 | ↔(M) | [22,80] | ||
| Atp7b | ↑(M) | [80] | ||
| Atp8b1 | ↓(M) | [22] | ||
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Moscovitz, J.E.; Aleksunes, L.M. Establishment of Metabolism and Transport Pathways in the Rodent and Human Fetal Liver. Int. J. Mol. Sci. 2013, 14, 23801-23827. https://doi.org/10.3390/ijms141223801
Moscovitz JE, Aleksunes LM. Establishment of Metabolism and Transport Pathways in the Rodent and Human Fetal Liver. International Journal of Molecular Sciences. 2013; 14(12):23801-23827. https://doi.org/10.3390/ijms141223801
Chicago/Turabian StyleMoscovitz, Jamie E., and Lauren M. Aleksunes. 2013. "Establishment of Metabolism and Transport Pathways in the Rodent and Human Fetal Liver" International Journal of Molecular Sciences 14, no. 12: 23801-23827. https://doi.org/10.3390/ijms141223801