Ultrastructural Interactions and Genotoxicity Assay of Cerium Dioxide Nanoparticles on Mouse Oocytes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
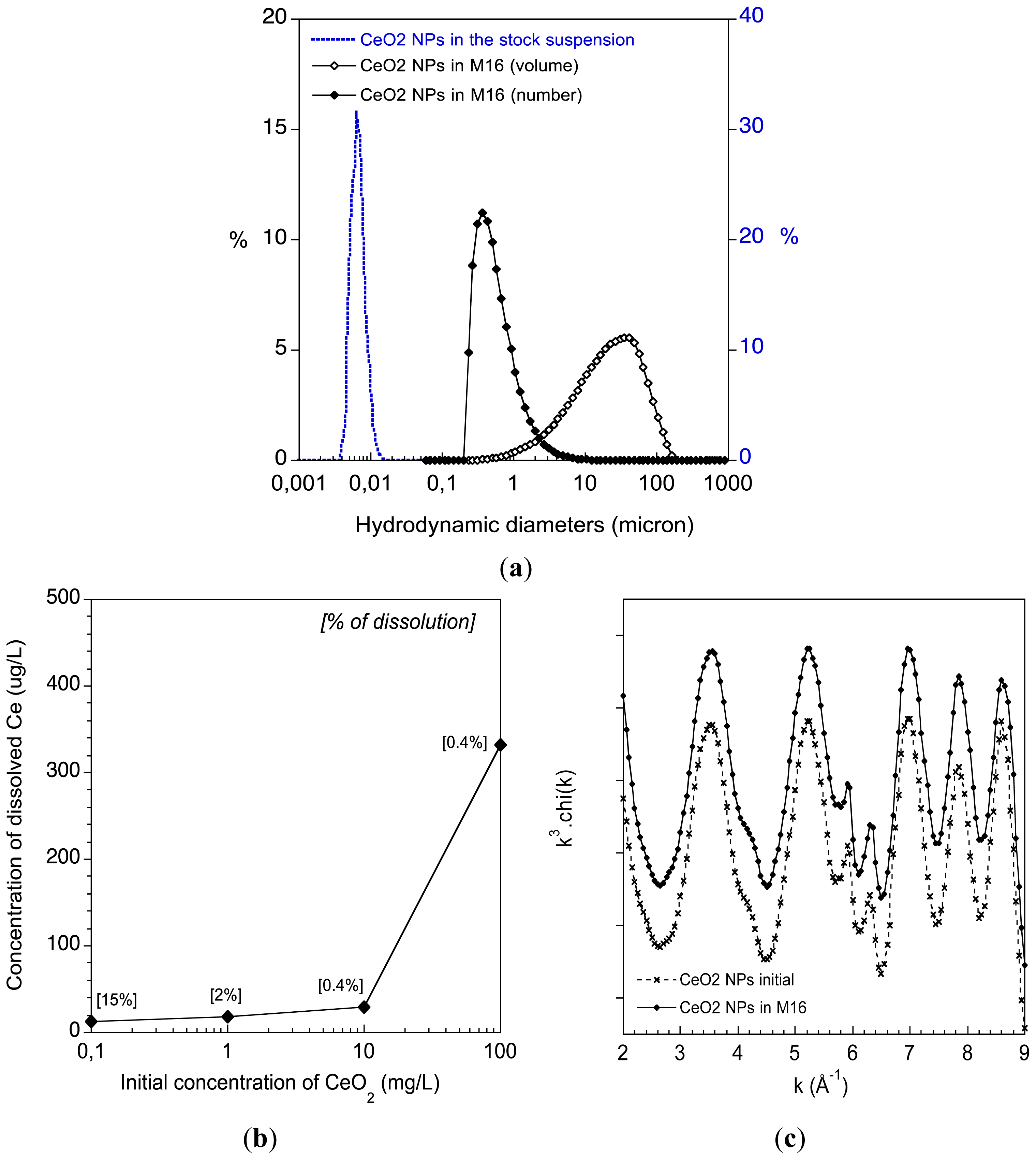
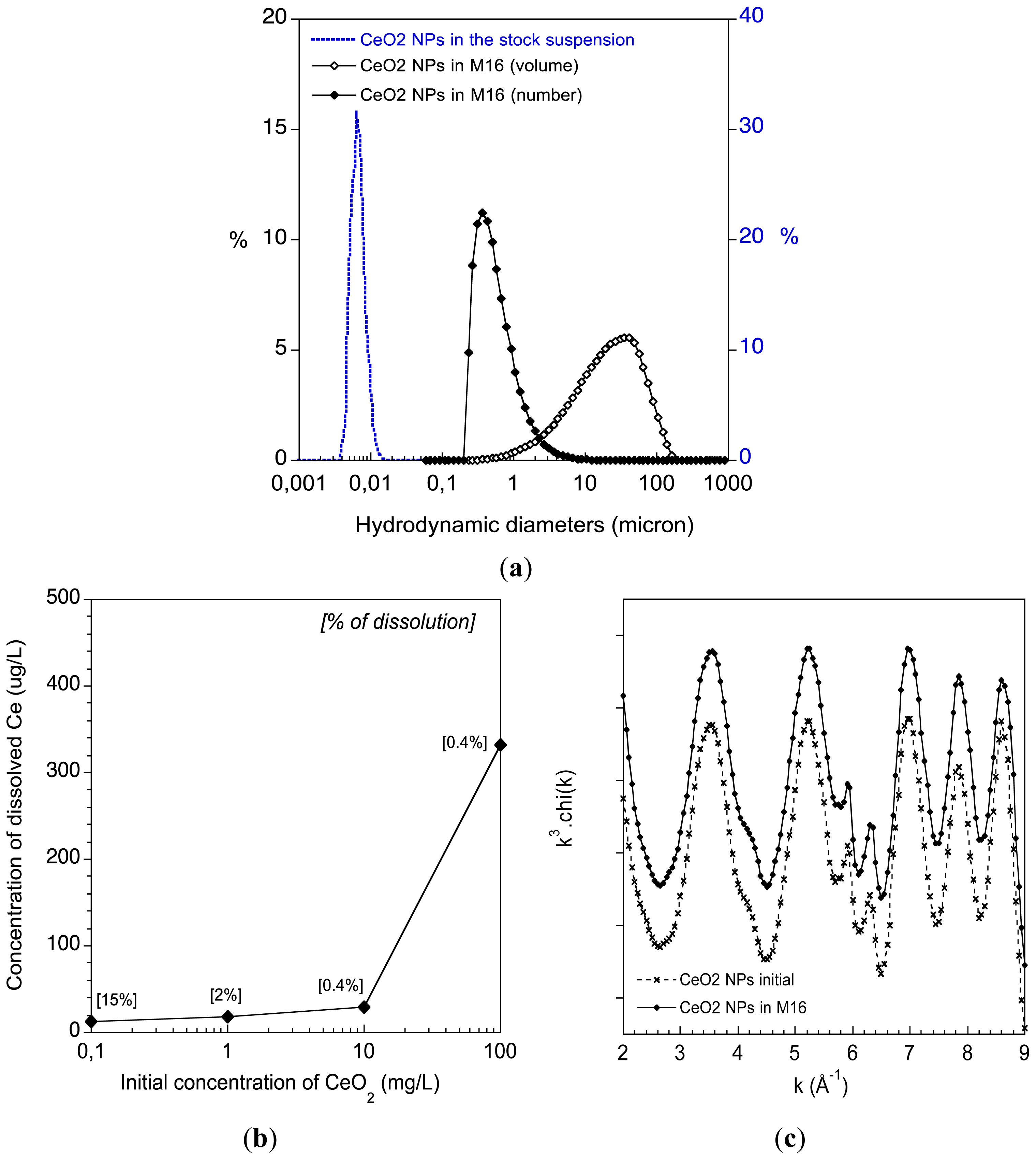
2.1. Physico-Chemical Behaviour of the CeO2 ENPs in the Oocyte Culture Medium M16
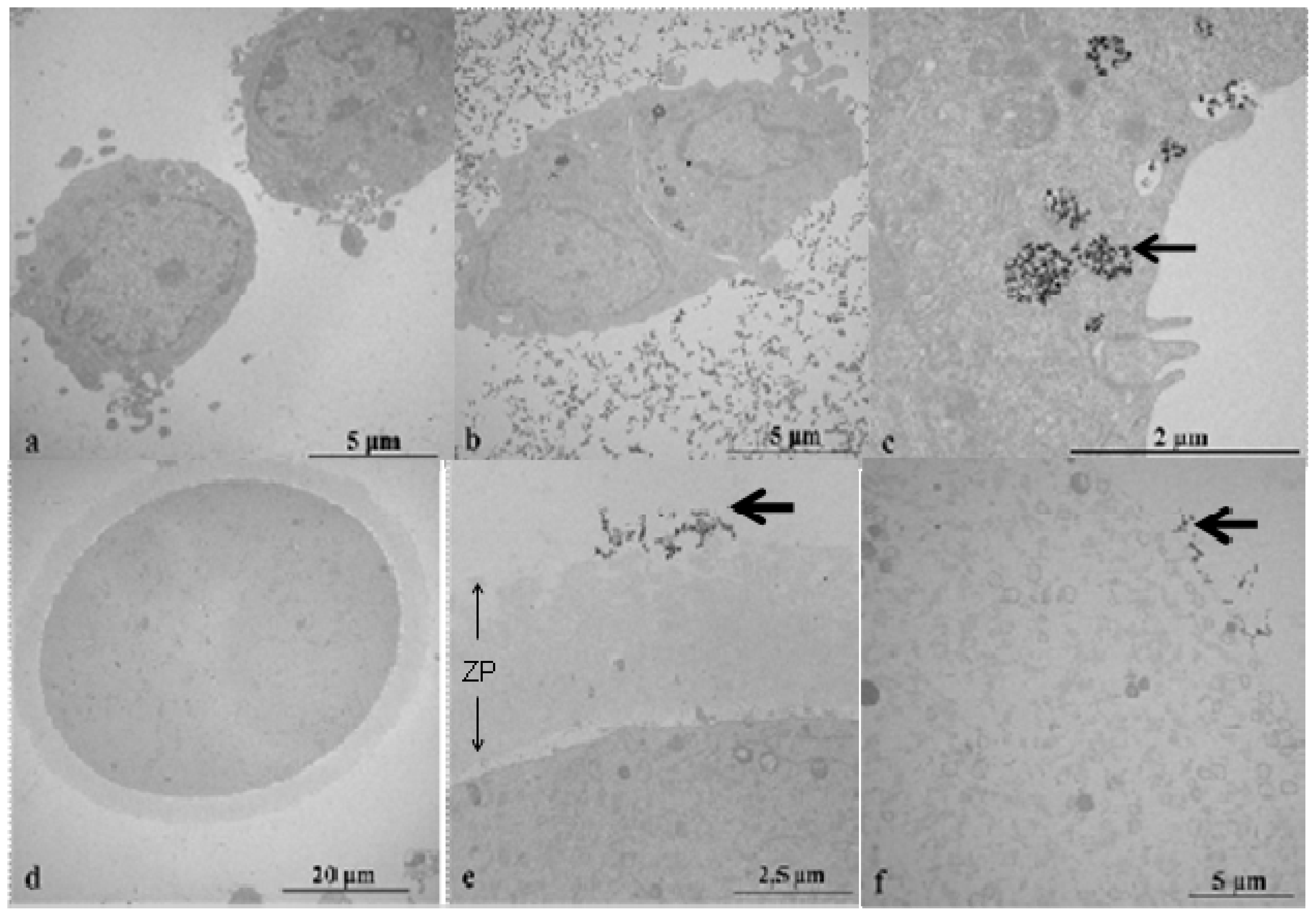
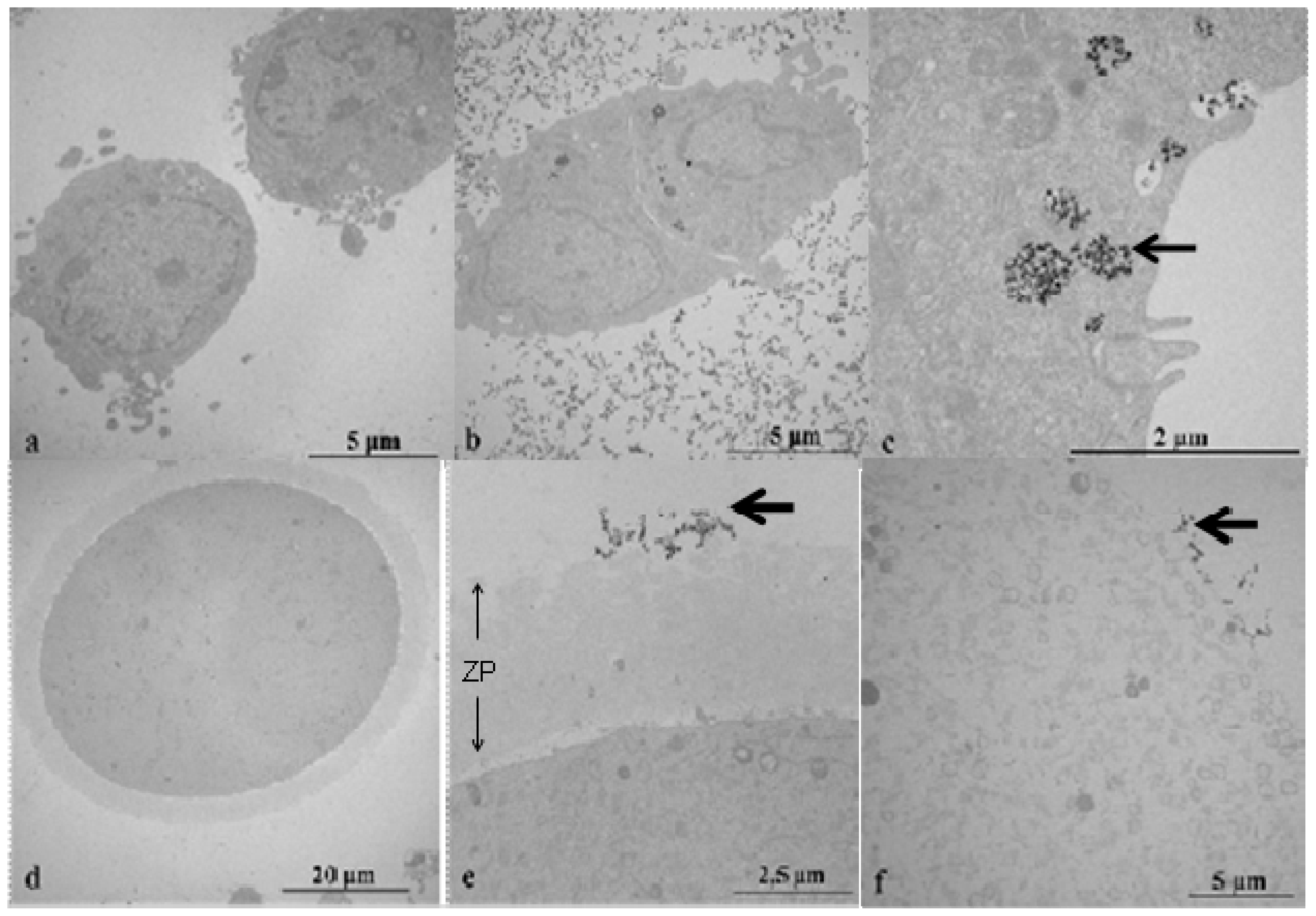
2.2. Transmission Electron Microscopy (TEM) Study of CeO2 ENPs Internalisation
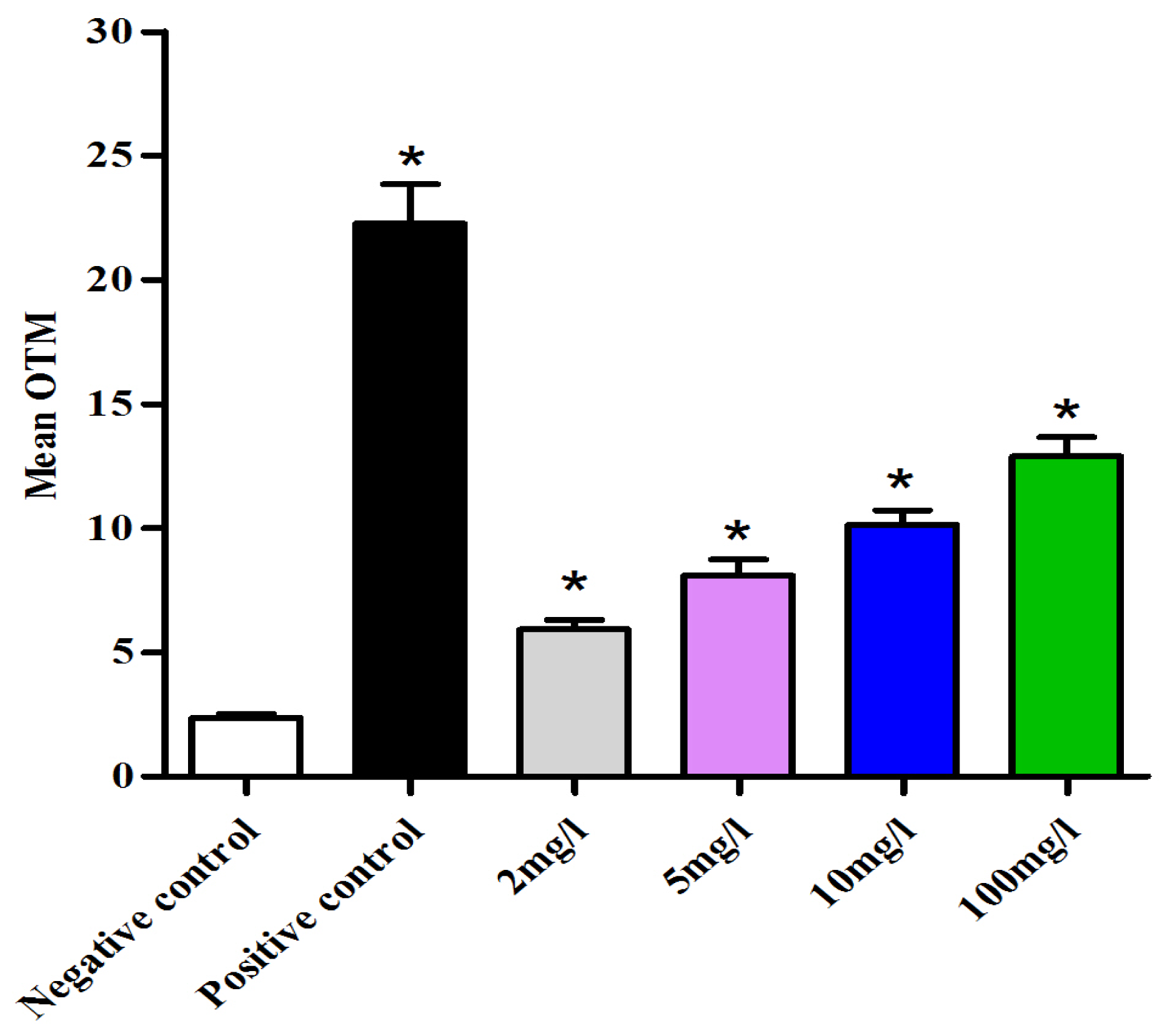
2.3. CeO2 ENPs Induced DNA Damage in Follicular Cells
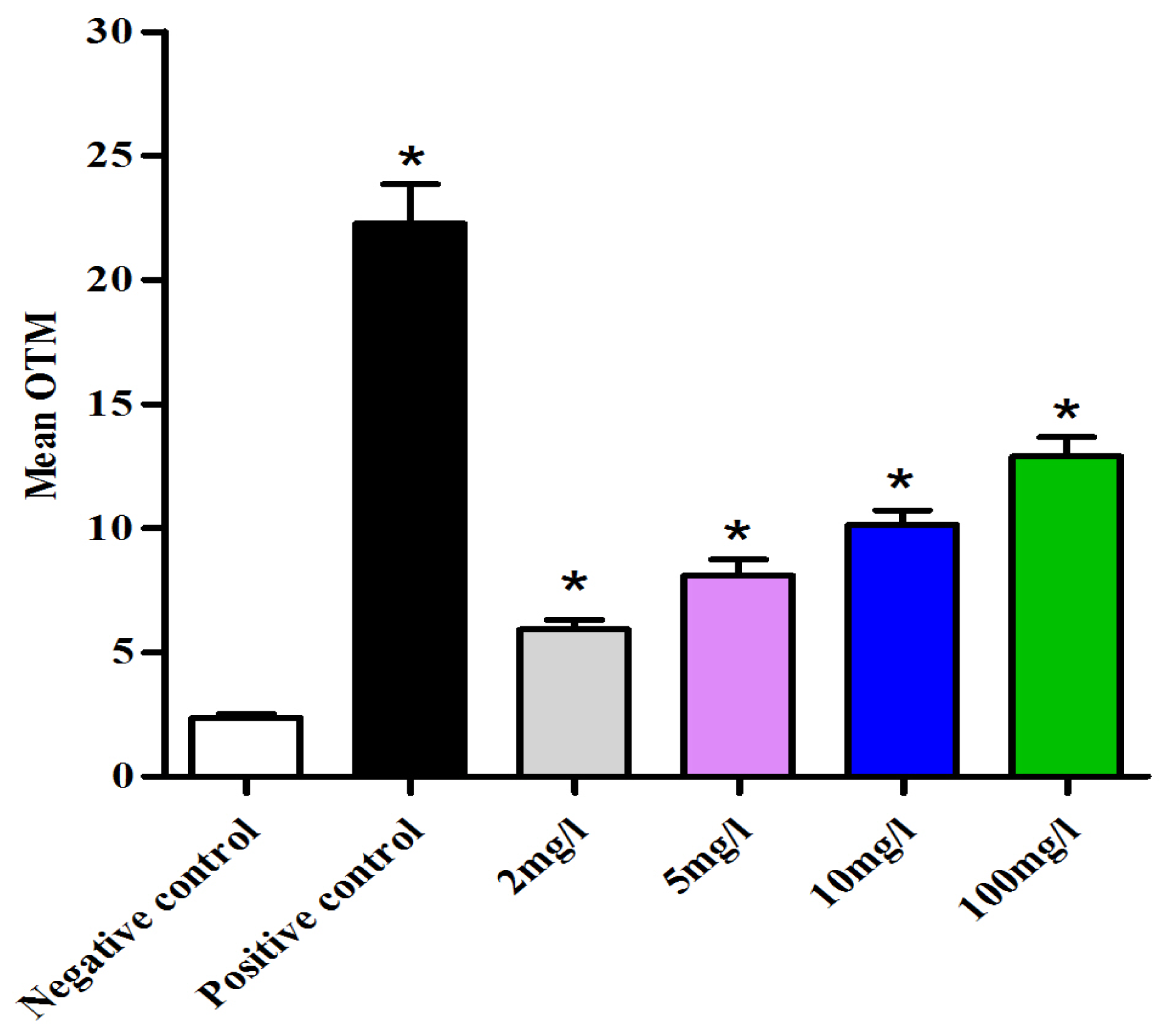
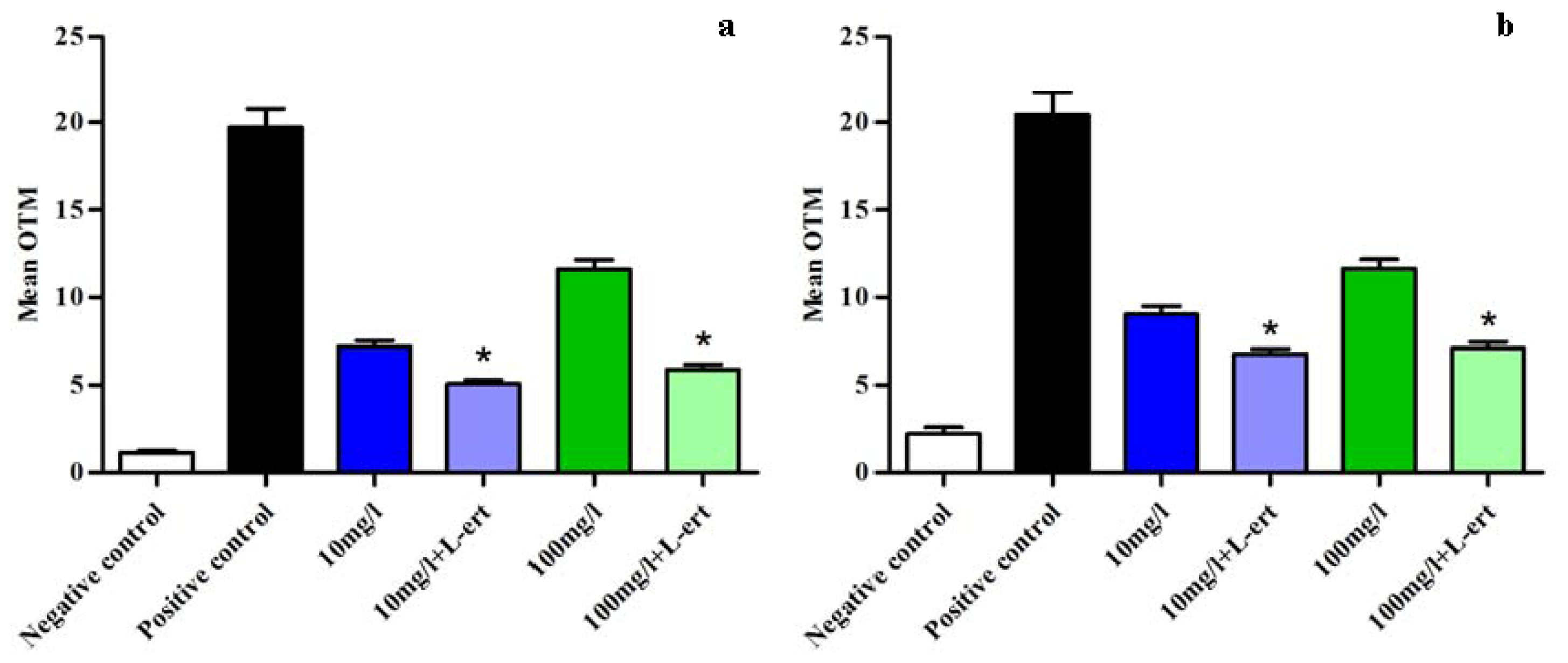
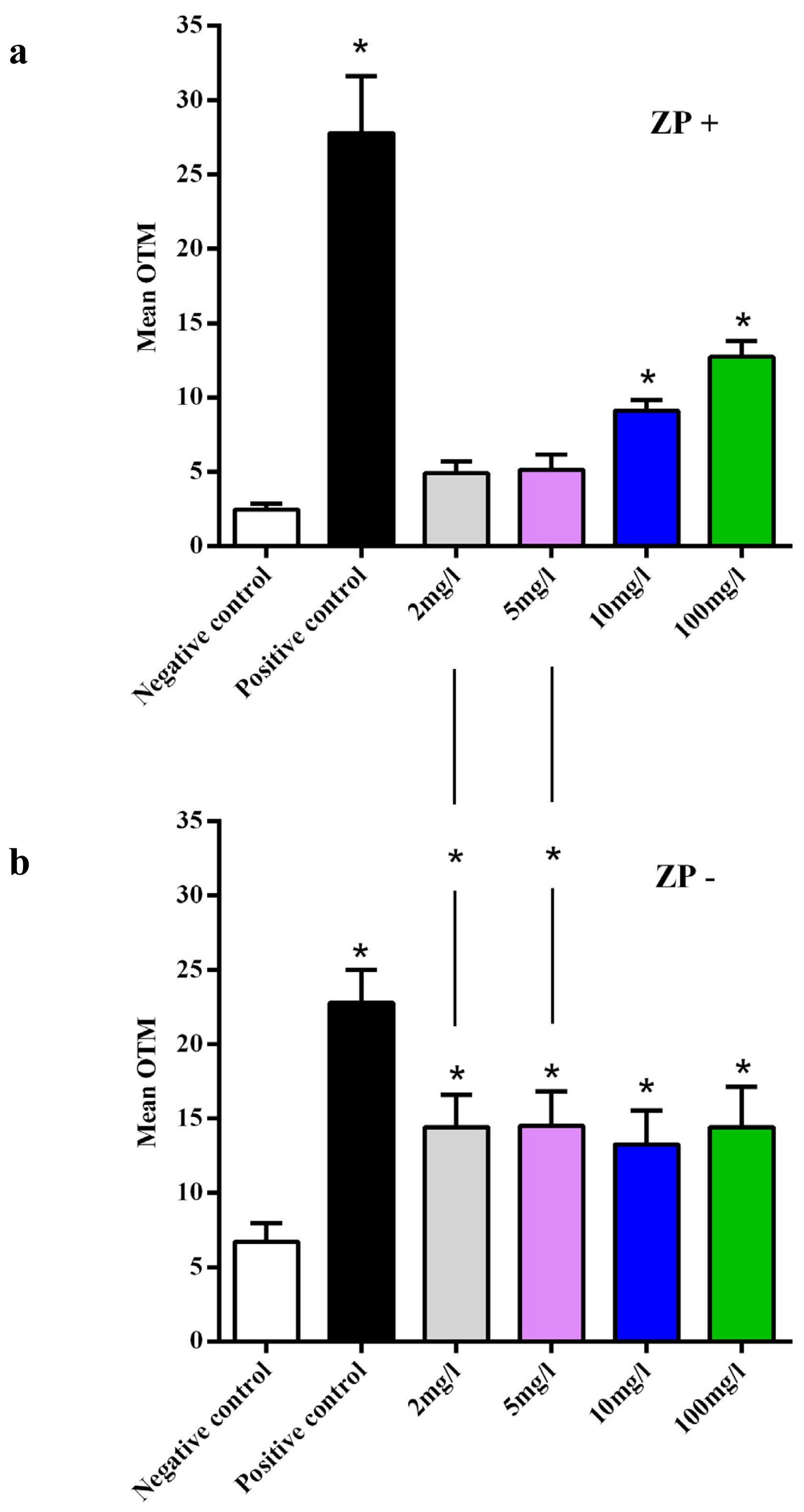
2.4. DNA Damage Assay in Oocytes
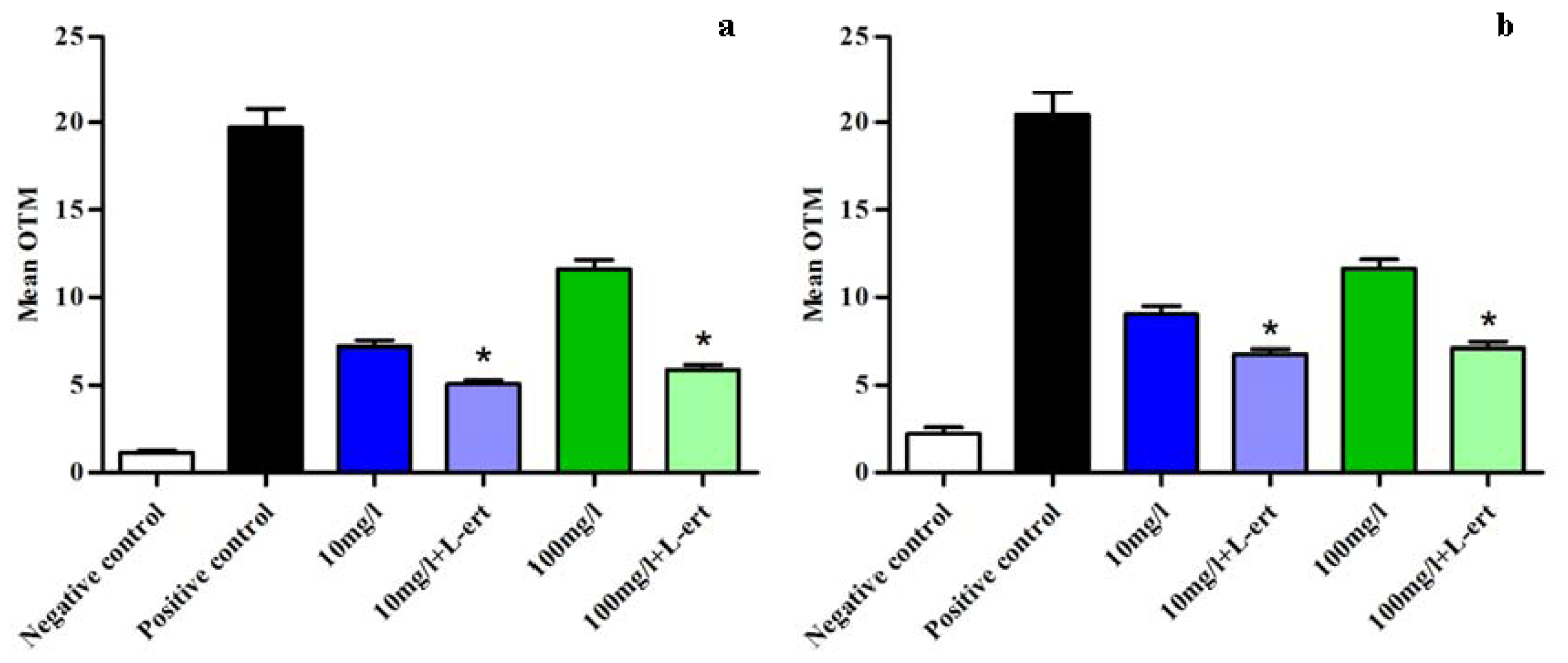
2.5. CeO2 ENPs and Anti-Oxidant
2.6. Discussion
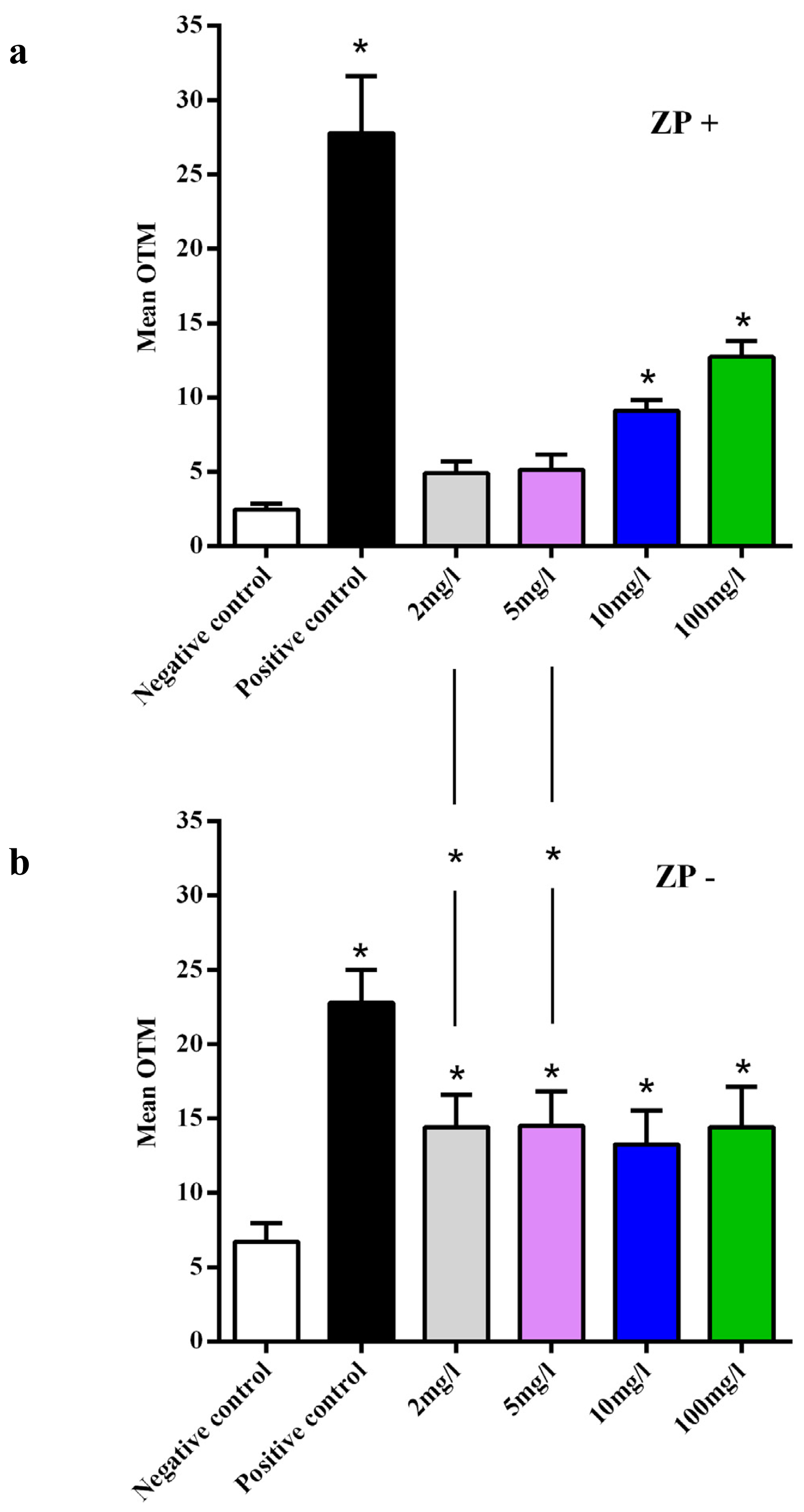
2.6.1. Are Oocytes Protected from CeO2 ENP Induced Oxidative Stress and DNA Damage by Follicular Cell Endocytosis and Zona Pellucida Trapping?
2.6.2. Mechanisms of Oxidative Stress Induced by CeO2 ENPs
3. Experimental Section
3.1. Chemical Agents
3.2. CeO2 ENPs Physico-Chemical Characterisation in Culture Medium
3.3. Animals
3.4. Oocytes Isolation
3.5. Transmission Electron Microscopy (TEM)
3.6. Exposure Conditions for Genotoxicity Assay
3.7. Incubation of Oocytes with an Anti-Oxidant
3.8. Main Outcome Measures and Statistical Analysis of the Comet Assay
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Bottero, J.Y.; Rose, J.; Wiesner, M.R. Nanotechnologies: Tools for sustainability in a new wave of water treatment processes. Integr. Environ. Assess. Manag 2006, 2, 391–395. [Google Scholar]
- Ko, S.H.; Park, I.; Pan, H.; Grigoropoulos, C.P.; Pisano, A.P.; Luscombe, C.K.; Frechet, J.M.J. Direct nanoimprinting of metal nanoparticles for nanoscale electronics fabrication. Nano Lett 2007, 7, 1869–1877. [Google Scholar]
- Som, C.; Wick, P.; Krug, H.; Nowack, B. Environmental and health effects of nanomaterials in nanotextiles and facade coatings. Environ. Int 2011, 37, 1131–1142. [Google Scholar]
- Botta, C.; Labille, J.; Auffan, M.; Borschneck, D.; Miche, H.; Cabie, M.; Masion, A.; Rose, J.; Bottero, J.Y. TiO2-based nanoparticles released in water from commercialized sunscreens in a life-cycle perspective: structures and quantities. Environ. Pollut 2011, 159, 1543–1550. [Google Scholar]
- Gupta, A.K.; Gupta, M. Synthesis and surface engineering of iron oxide nanoparticles for biomedical applications. Biomaterials 2005, 26, 3995–4021. [Google Scholar]
- Lammers, T.; Hennink, W.E.; Storm, G. Tumour-targeted nanomedicines: Principles and practice. Br. J. Cancer 2008, 99, 392–397. [Google Scholar]
- Auffan, M.; Bottero, J.Y.; Chaneac, C.; Rose, J. Inorganic manufactured nanoparticles: How their physicochemical properties influence their biological effects in aqueous environments. Nanomedicine 2010, 5, 999–1007. [Google Scholar]
- Avis de l’Afsset; Rapport d’Expertise Collective. Évaluation des risques liés aux nanomatériaux pour la population générale et pour l’environnement. La Doc. Fr. 2010, 3, pp. 1–207. Available online: http://www.ladocumentationfrancaise.fr/var/storage/rapports-publics/104000168/0000.pdf (accessed on 25 October 2013).
- Cassee, F.R.; van Balen, E.C.; Singh, C.; Green, D.; Muijser, H.; Weinstein, J.; Dreher, K. Exposure, health and ecological effects review of engineered nanoscale cerium and cerium oxide associated with its use as a fuel additive. Crit. Rev. Toxicol 2011, 41, 213–229. [Google Scholar]
- Blanchard, V.; Blanchet, P. Color stability for wood products during use: Effects of inorganic nanoparticles. BioResources 2011, 6, 1219–1229. [Google Scholar]
- Niu, J.; Azfer, A.; Rogers, L.M.; Wang, X.; Kolattukudy, P.E. Cardioprotective effects of cerium oxide nanoparticles in a transgenic murine model of cardiomyopathy. Cardiovasc. Res 2007, 73, 549–559. [Google Scholar]
- Tarnuzzer, R.W.; Colon, J.; Patil, S.; Seal, S. Vacancy engineered ceria nanostructures for protection from radiation-induced cellular damage. Nano Lett 2005, 5, 2573–2577. [Google Scholar]
- Park, B.; Donaldson, K.; Duffin, R.; Tran, L.; Kelly, F.; Mudway, I.; Morin, J.P.; Guest, R.; Jenkinson, P.; Samaras, Z.; et al. Hazard and risk assessment of a nanoparticulate cerium oxide-based diesel fuel additive—A case study. Inhal. Toxicol 2008, 20, 547–566. [Google Scholar]
- Oberdorster, G. Safety assessment for nanotechnology and nanomedicine: Concepts of nanotoxicology. J. Intern. Med 2010, 267, 89–105. [Google Scholar]
- Hansen, S.F.; Baun, A. European regulation affecting nanomaterials—Review of limitations and future recommendations. Dose Response 2012, 10, 364–383. [Google Scholar]
- Oberdorster, G.; Maynard, A.; Donaldson, K.; Castranova, V.; Fitzpatrick, J.; Ausman, K.; Carter, J.; Karn, B.; Kreyling, W.; Lai, D.; et al. Principles for characterizing the potential human health effects from exposure to nanomaterials: Elements of a screening strategy. Part. Fibre Toxicol 2005, 2, 8. [Google Scholar]
- Warheit, D.B.; Donner, E.M. Rationale of genotoxicity testing of nanomaterials: Regulatory requirements and appropriateness of available OECD test guidelines. Nanotoxicology 2010, 4, 409–413. [Google Scholar]
- Celardo, I.; Pedersen, J.Z.; Traversa, E.; Ghibelli, L. Pharmacological potential of cerium oxide nanoparticles. Nanoscale 2011, 3, 1411–1420. [Google Scholar]
- Heckert, E.G.; Karakoti, A.S.; Seal, S.; Self, W.T. The role of cerium redox state in the SOD mimetic activity of nanoceria. Biomaterials 2008, 29, 2705–2709. [Google Scholar]
- Hirst, S.M.; Karakoti, A.; Singh, S.; Self, W.; Tyler, R.; Seal, S.; Reilly, C.M. Bio-distribution and in vivo antioxidant effects of cerium oxide nanoparticles in mice. Environ. Toxicol 2013, 28, 107–118. [Google Scholar]
- Celardo, I.; Traversa, E.; Ghibelli, L. Cerium oxide nanoparticles: A promise for applications in therapy. J. Exp. Ther. Oncol 2011, 9, 47–51. [Google Scholar]
- Asati, A.; Santra, S.; Kaittanis, C.; Perez, J.M. Surface-charge-dependent cell localization and cytotoxicity of cerium oxide nanoparticles. ACS Nano 2010, 4, 5321–5331. [Google Scholar]
- Rodea-Palomares, I.; Boltes, K.; Fernandez-Pinas, F.; Leganes, F.; Garcia-Calvo, E.; Santiago, J.; Rosal, R. Physicochemical characterization and ecotoxicological assessment of CeO2 nanoparticles using two aquatic microorganisms. Toxicol. Sci 2011, 119, 135–145. [Google Scholar]
- Van Hoecke, K.; de Schamphelaere, K.A.C.; van der Meeren, P.; Smagghe, G.; Janssen, C.R. Aggregation and ecotoxicity of CeO2 nanoparticles in synthetic and natural waters with variable pH, organic matter concentration and ionic strength. Environ. Pollut 2011, 159, 970–976. [Google Scholar]
- Shaw, B.J.; Handy, R.D. Physiological effects of nanoparticles on fish: A comparison of nanometals vs. metal ions. Environ. Int 2011, 37, 1083–1097. [Google Scholar]
- Zhang, X.; Sun, H.; Zhang, Z.; Niu, Q.; Chen, Y.; Crittenden, J.C. Enhanced bioaccumulation of cadmium in carp in the presence of titanium dioxide nanoparticles. Chemosphere 2007, 67, 160–166. [Google Scholar]
- Pelletier, D.A.; Suresh, A.K.; Holton, G.A.; McKeown, C.K.; Wang, W.; Gu, B.; Mortensen, N.P.; Allison, D.P.; Joy, D.C.; Allison, M.R.; et al. Effects of engineered cerium oxide nanoparticles on bacterial growth and viability. Appl. Environ. Microbiol 2010, 76, 7981–7989. [Google Scholar]
- Li, J.J.; Muralikrishnan, S.; Ng, C.T.; Yung, L.Y.; Bay, B.H. Nanoparticle-induced pulmonary toxicity. Exp. Biol. Med 2010, 235, 1025–1033. [Google Scholar]
- Smita, S.; Gupta, S.K.; Bartonova, A.; Dusinska, M.; Gutleb, A.C.; Rahman, Q. Nanoparticles in the environment: Assessment using the causal diagram approach. Environ. Heal 2012, 11, S13. [Google Scholar]
- Auffan, M.; Rose, J.; Orsiere, T.; Demeo, M.; Thill, A.; Zeyons, O.; Proux, O.; Masion, A.; Chaurand, P.; Spalla, O.; et al. CeO2 nanoparticles induce DNA damage towards human dermal fibroblasts in vitro. Nanotoxicology 2009, 3, 161–171. [Google Scholar]
- Priester, J.H.; Ge, Y.; Mielke, R.E.; Horst, A.M.; Moritz, S.C.; Espinosa, K.; Gelb, J.; Walker, S.L.; Nisbet, R.M.; An, Y.J.; et al. Soybean susceptibility to manufactured nanomaterials with evidence for food quality and soil fertility interruption. Proc. Natl. Acad. Sci. USA 2012, 109, E2451–E2456. [Google Scholar]
- Becker, H.; Herzberg, F.; Schulte, A.; Kolossa-Gehring, M. The carcinogenic potential of nanomaterials, their release from products and options for regulating them. Int. J. Hyg. Environ. Health 2011, 214, 231–238. [Google Scholar]
- Al-Saleh, I.; Shinwari, N.; Al-Amodi, M. Accumulation of mercury in ovaries of mice after the application of skin-lightening creams. Biol. Trace Elem. Res 2009, 131, 43–54. [Google Scholar]
- Morishita, Y.; Yoshioka, Y.; Satoh, H.; Nojiri, N.; Nagano, K.; Abe, Y.; Kamada, H.; Tsunoda, S.; Nabeshi, H.; Yoshikawa, T.; et al. Distribution and histologic effects of intravenously administered amorphous nanosilica particles in the testes of mice. Biochem. Biophys. Res. Commun 2012, 420, 297–301. [Google Scholar]
- Klein, J.P.; Boudard, D.; Cadusseau, J.; Palle, S.; Forest, V.; Pourchez, J.; Cottier, M. Testicular biodistribution of 450 nm fluorescent latex particles after intramuscular injection in mice. Biomed. Microdevices 2013, 15, 427–436. [Google Scholar] [Green Version]
- Lim, D.; Roh, J.; Eom, H.; Choi, J.Y.; Hyun, J.; Choi, J. Oxidative stress-related PMK-1 P38 MAPK activation as a mechanism for toxicity of silver nanoparticles to reproduction in the nematode Caenorhabditis elegans. Environ. Toxicol. Chem 2012, 31, 585–592. [Google Scholar]
- Musee, N.; Oberholster, P.J.; Sikhwivhilu, L.; Botha, A.M. The effects of engineered nanoparticles on survival, reproduction, and behaviour of freshwater snail, Physa acuta (Draparnaud, 1805). Chemosphere 2010, 81, 1196–1203. [Google Scholar]
- Philbrook, N.A.; Winn, L.M.; Afrooz, A.R.; Saleh, N.B.; Walker, V.K. The effect of TiO2 and Ag nanoparticles on reproduction and development of Drosophila melanogaster and CD-1 mice. Toxicol. Appl. Pharmacol 2011, 257, 429–436. [Google Scholar]
- Ramsden, C.S.; Henry, T.B.; Handy, R.D. Sub-lethal effects of titanium dioxide nanoparticles on the physiology and reproduction of zebrafish. Aquat. Toxicol 2013, 126, 404–413. [Google Scholar]
- Schlich, K.; Klawonn, T.; Terytze, K.; Hund-Rinke, K. Effects of silver nanoparticles and silver nitrate in the earthworm reproduction test. Environ. Toxicol. Chem 2013, 32, 181–188. [Google Scholar]
- Wang, J.; Zhu, X.; Zhang, X.; Zhao, Z.; Liu, H.; George, R.; Wilson-Rawls, J.; Chang, Y.; Chen, Y. Disruption of zebrafish (Danio rerio) reproduction upon chronic exposure to TiO2 nanoparticles. Chemosphere 2011, 83, 461–467. [Google Scholar]
- Braydich-Stolle, L.; Hussain, S.; Schlager, J.J.; Hofmann, M.C. In vitro cytotoxicity of nanoparticles in mammalian germline stem cells. Toxicol. Sci 2005, 88, 412–419. [Google Scholar]
- Braydich-Stolle, L.K.; Lucas, B.; Schrand, A.; Murdock, R.C.; Lee, T.; Schlager, J.J.; Hussain, S.M.; Hofmann, M.C. Silver nanoparticles disrupt GDNF/Fyn kinase signaling in spermatogonial stem cells. Toxicol. Sci 2010, 116, 577–589. [Google Scholar]
- Wiwanitkit, V.; Sereemaspun, A.; Rojanathanes, R. Effect of gold nanoparticles on spermatozoa: The first world report. Fertil. Steril 2009, 91, e7–e8. [Google Scholar]
- Chaudhury, K.; Babu, N.K.; Singh, A.K.; Das, S.; Kumar, A.; Seal, S. Mitigation of endometriosis using regenerative cerium oxide nanoparticles. Nanomed. Nanotechnol. Biol. Med 2013, 9, 439–448. [Google Scholar]
- Berthelot-Ricou, A.; Perrin, J.; di Giorgio, C.; de Meo, M.; Botta, A.; Courbiere, B. Comet assay on mouse oocytes: An improved technique to evaluate genotoxic risk on female germ cells. Fertil. Steril 2011, 95, 1452–1457. [Google Scholar]
- Tice, R.R.; Agurell, E.; Anderson, D.; Burlinson, B.; Hartmann, A.; Kobayashi, H.; Miyamae, Y.; Rojas, E.; Ryu, J.C.; Sasaki, Y.F. Single cell gel/comet assay: Guidelines for in vitro and in vivo genetic toxicology testing. Environ. Mol. Mutagen 2000, 35, 206–221. [Google Scholar]
- Singh, N.P.; McCoy, M.T.; Tice, R.R.; Schneider, E.L. A simple technique for quantitation of low levels of DNA damage in individual cells. Exp. Cell Res 1988, 175, 184–191. [Google Scholar]
- Browning, L.M.; Lee, K.J.; Huang, T.; Nallathamby, P.D.; Lowman, J.E.; Xu, X.-H.N. Random walk of single gold nanoparticles in zebrafish embryos leading to stochastic toxic effects on embryonic developments. Nanoscale 2009, 1, 138–152. [Google Scholar]
- Levy, R.; Shaheen, U.; Cesbron, Y.; See, V. Gold nanoparticles delivery in mammalian live cells: A critical review. Nano Rev 2010, 1. [Google Scholar] [CrossRef]
- Rejman, J.; Oberle, V.; Zuhorn, I.S.; Hoekstra, D. Size-dependent internalization of particles via the pathways of clathrin- and caveolae-mediated endocytosis. Biochem. J 2004, 377, 159–169. [Google Scholar]
- Faraji, A.H.; Wipf, P. Nanoparticles in cellular drug delivery. Bioorg. Med. Chem 2009, 17, 2950–2962. [Google Scholar]
- Yoshida, T.; Yoshioka, Y.; Matsuyama, K.; Nakazato, Y.; Tochigi, S.; Hirai, T.; Kondoh, S.; Nagano, K.; Abe, Y.; Kamada, H.; et al. Surface modification of amorphous nanosilica particles suppresses nanosilica-induced cytotoxicity, ROS generation, and DNA damage in various mammalian cells. Biochem. Biophys. Res. Commun 2012, 427, 748–752. [Google Scholar]
- Nel, A.E.; Madler, L.; Velegol, D.; Xia, T.; Hoek, E.M.V.; Somasundaran, P.; Klaessig, F.; Castranova, V.; Thompson, M. Understanding biophysicochemical interactions at the nano-bio interface. Nat. Mater 2009, 8, 543–557. [Google Scholar]
- Zeyons, O.; Thill, A.; Chauvat, F.; Menguy, N.; Cassier-Chauvat, C.; Orear, C.; Daraspe, J.; Auffan, M.; Rose, J.; Spalla, O. Direct and Indirect CeO2 Nanoparticles Toxicity for Escherichia coli and Synechocystis. Available online: http://www.informahealthcare.com/doi/abs/10.3109/17435390903305260 (accessed on 21 August 2013).
- Cicco, A.D.; Aquilanti, G.; Minicucci, M.; Principi, E.; Novello, N.; Cognigni, A.; Olivi, L. Novel XAFS capabilities at ELETTRA synchrotron light source. J. Phys 2009, 190, 012043. [Google Scholar]
- Ravel, B.; Newville, M. ATHENA, ARTEMIS, HEPHAESTUS: Data analysis for X-ray absorption spectroscopy using IFEFFIT. J. Synchrotron Radiat 2005, 12, 537–541. [Google Scholar]
- Roustan, A.; Perrin, J.; Berthelot-Ricou, A.; Lopez, E.; Botta, A.; Courbiere, B. Evaluating methods of mouse euthanasia on the oocyte quality: Cervical dislocation vs. isoflurane inhalation. Lab. Anim 2012, 46, 167–169. [Google Scholar]
- Akanmu, D.; Cecchini, R.; Aruoma, O.I.; Halliwell, B. The antioxidant action of ergothioneine. Arch. Biochem. Biophys 1991, 288, 10–16. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Courbiere, B.; Auffan, M.; Rollais, R.; Tassistro, V.; Bonnefoy, A.; Botta, A.; Rose, J.; Orsière, T.; Perrin, J. Ultrastructural Interactions and Genotoxicity Assay of Cerium Dioxide Nanoparticles on Mouse Oocytes. Int. J. Mol. Sci. 2013, 14, 21613-21628. https://doi.org/10.3390/ijms141121613
Courbiere B, Auffan M, Rollais R, Tassistro V, Bonnefoy A, Botta A, Rose J, Orsière T, Perrin J. Ultrastructural Interactions and Genotoxicity Assay of Cerium Dioxide Nanoparticles on Mouse Oocytes. International Journal of Molecular Sciences. 2013; 14(11):21613-21628. https://doi.org/10.3390/ijms141121613
Chicago/Turabian StyleCourbiere, Blandine, Mélanie Auffan, Raphaël Rollais, Virginie Tassistro, Aurélie Bonnefoy, Alain Botta, Jérôme Rose, Thierry Orsière, and Jeanne Perrin. 2013. "Ultrastructural Interactions and Genotoxicity Assay of Cerium Dioxide Nanoparticles on Mouse Oocytes" International Journal of Molecular Sciences 14, no. 11: 21613-21628. https://doi.org/10.3390/ijms141121613