An Imprinted Cross-Linked Enzyme Aggregate (iCLEA) of Sucrose Phosphorylase: Combining Improved Stability with Altered Specificity


Abstract
:
1. Introduction
2. Results and Discussion
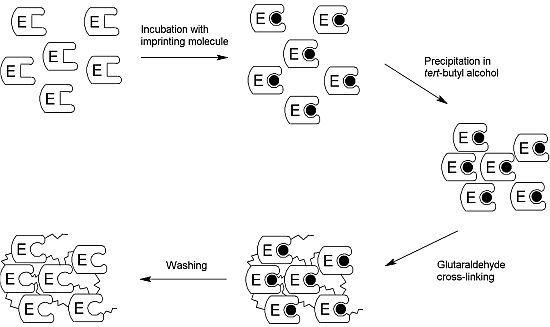
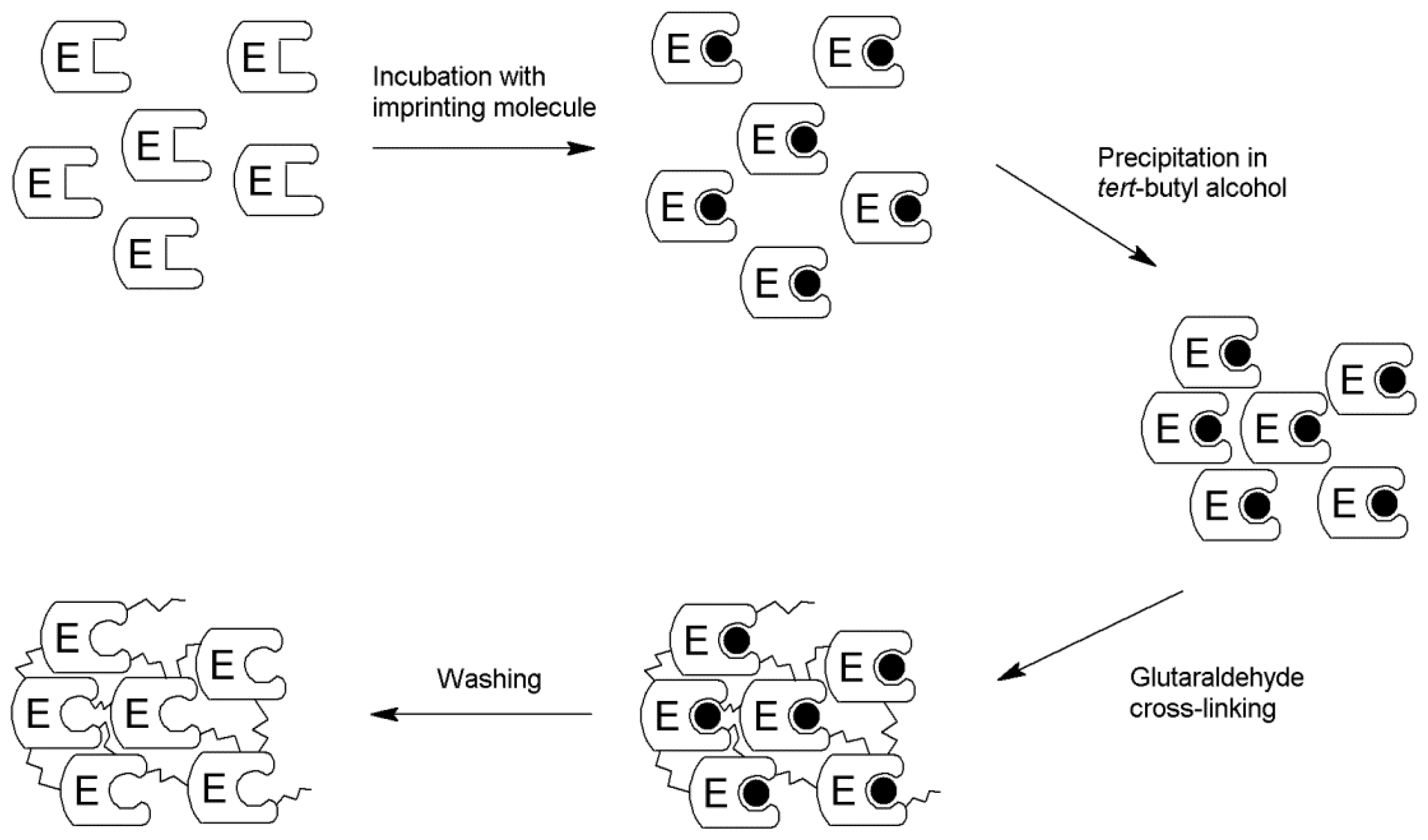
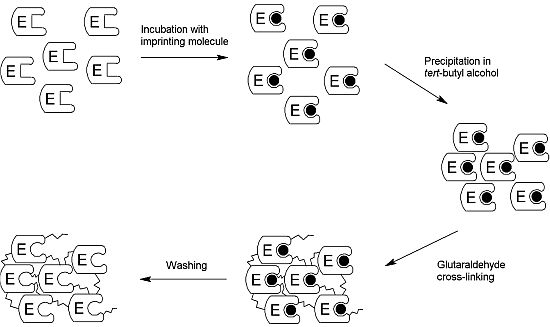
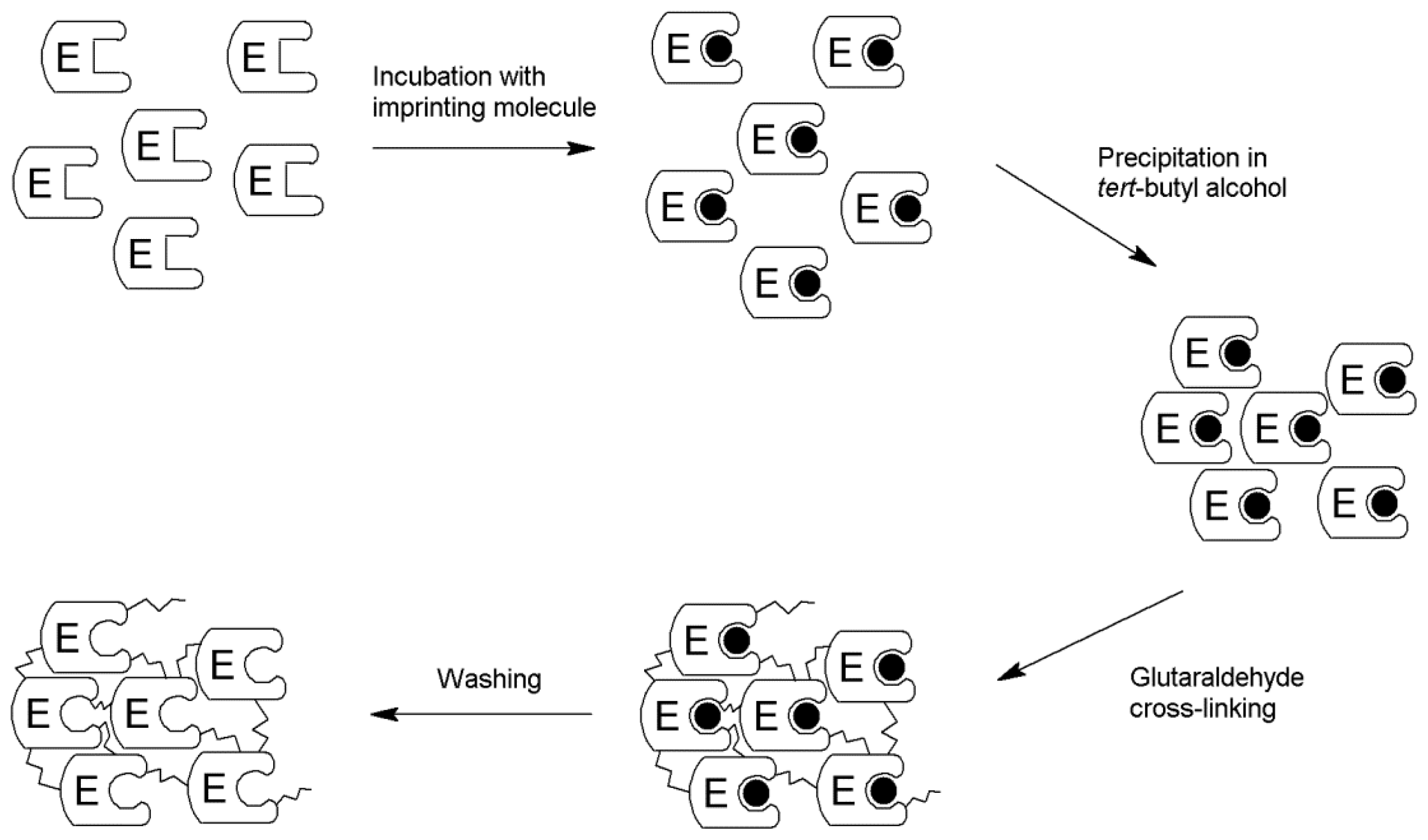
2.1. Production of the iCLEA
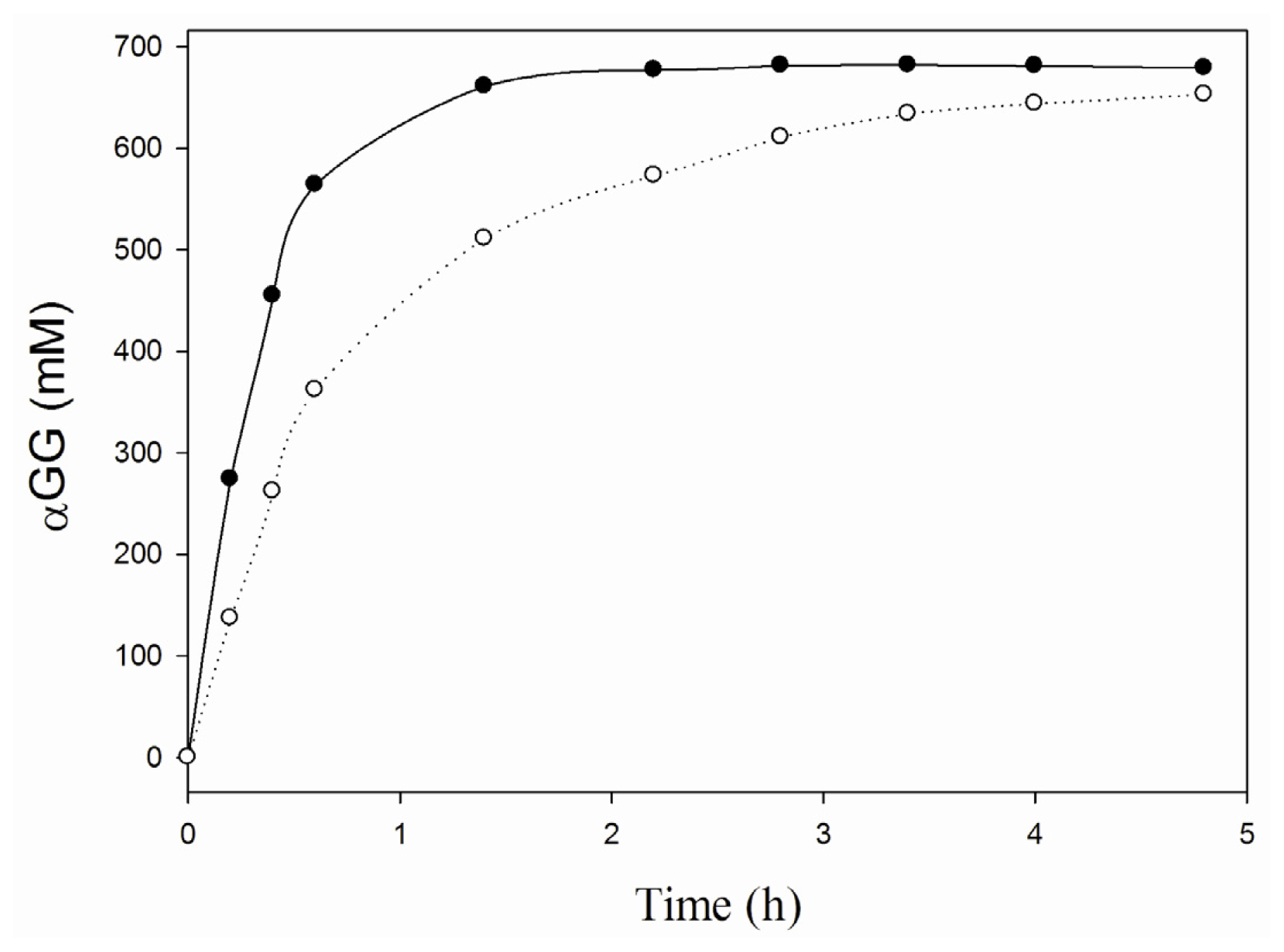
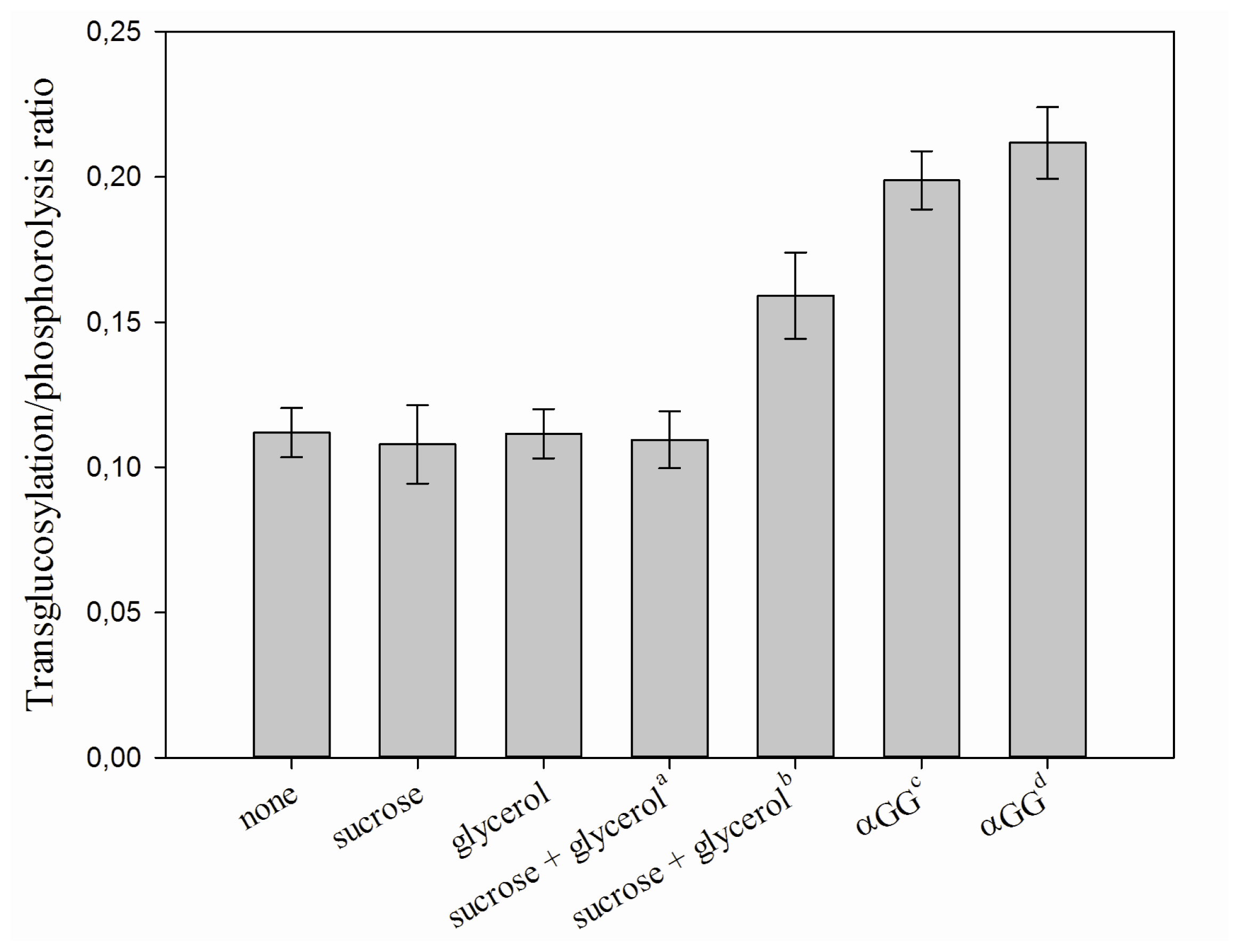
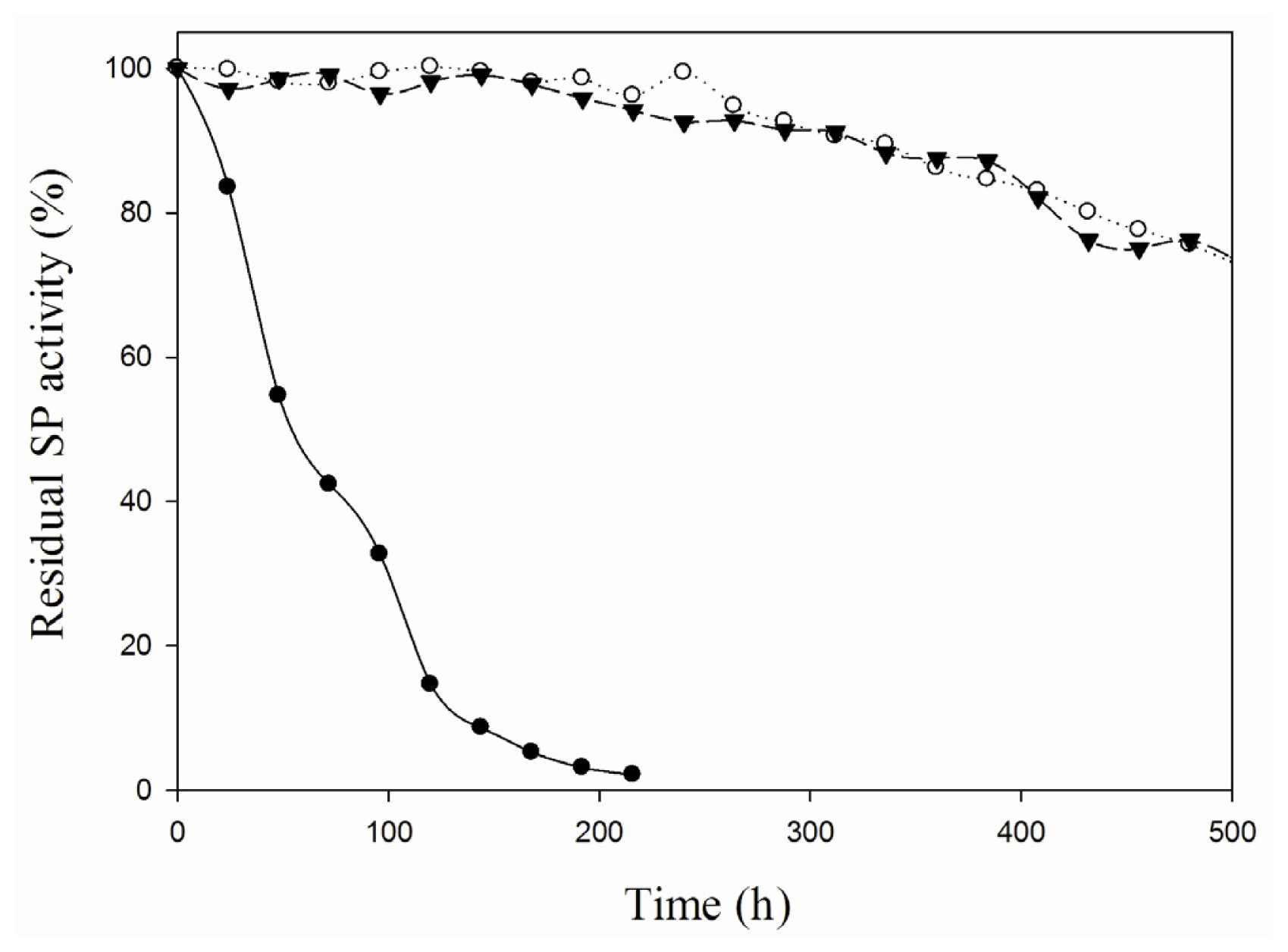
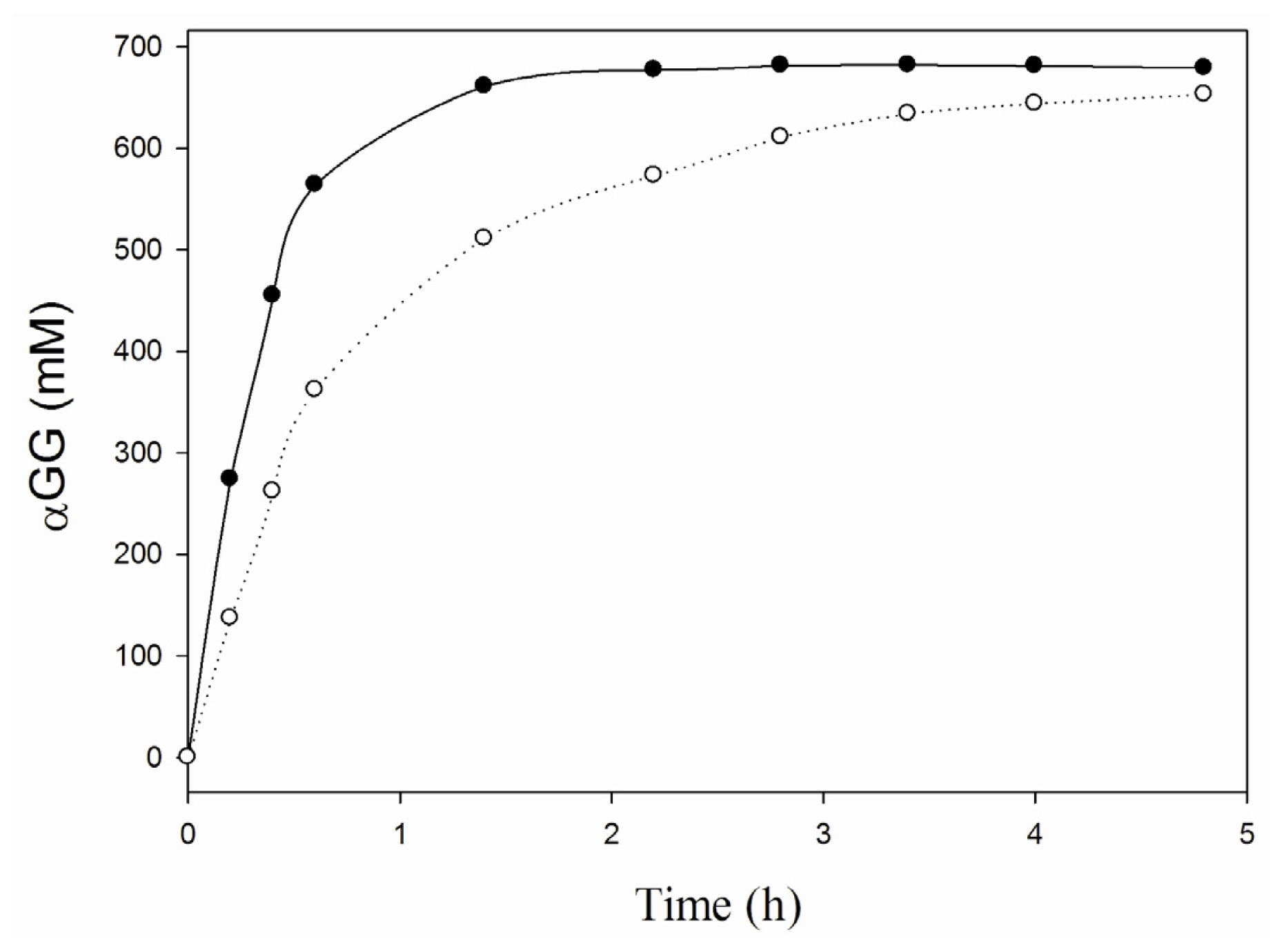
2.2. Characterization of the iCLEA
3. Experimental Section
3.1. Materials
3.2. Enzyme Production and Purification
3.3. Preparation of the iCLEA
3.4. Activity Assays
3.5. Stability Assays
4. Conclusions
Acknowledgments
References
- Henrissat, B. A classification of glycosyl hydrolases based on amino-acid sequence similarities. Biochem. J 1991, 280, 309–316. [Google Scholar]
- Goedl, C.; Schwarz, A.; Mueller, M.; Brecker, L.; Nidetzky, B. Mechanistic differences among retaining disaccharide phosphorylases: Insights from kinetic analysis of active site mutants of sucrose phosphorylase and alpha, alpha-trehalose phosphorylase. Carbohydr. Res 2008, 343, 2032–2040. [Google Scholar]
- Sprogoe, D.; van den Broek, L.A.M.; Mirza, O.; Kastrup, J.S.; Voragen, A.G.J.; Gajhede, M.; Skov, L.K. Crystal structure of sucrose phosphorylase from bifidobacterium adolescentis. Biochemistry 2004, 43, 1156–1162. [Google Scholar]
- Goedl, C.; Sawangwan, T.; Wildberger, P.; Nidetzky, B. Sucrose phosphorylase: A powerful transglucosylation catalyst for the synthesis of alpha-d-glucosides as industrial fine chemicals. Biocatal. Biotransform 2010, 28, 10–21. [Google Scholar]
- Goedl, C.; Sawangwan, T.; Mueller, M.; Schwarz, A.; Nidetzky, B. A high-yielding biocatalytic process for the production of 2-O-(α-d-glucopyranosyl)-sn-glycerol, a natural osmolyte and useful moisturizing ingredient. Angew. Chem. Int. Ed. Engl 2008, 47, 10086–10089. [Google Scholar]
- Sawangwan, T.; Goedl, C.; Nidetzky, B. Single-step enzymatic synthesis of (R)-2-O-α-d-glucopyranosyl glycerate, a compatible solute from micro-organisms that functions as a protein stabiliser. Org. Biomol. Chem 2009, 7, 4267–4270. [Google Scholar]
- Bornscheuer, U.T. Immobilizing enzymes: How to create more suitable biocatalysts. Angew. Chem.-Int. Ed 2003, 42, 3336–3337. [Google Scholar]
- Aerts, D.; Verhaeghe, T.F.; Roman, B.I.; Stevens, C.V.; Desmet, T.; Soetaert, W. Transglucosylation potential of six sucrose phosphorylases toward different classes of acceptors. Carbohydr. Res 2011, 346, 1860–1867. [Google Scholar]
- Cerdobbel, A.; Desmet, T.; de Winter, K.; Maertens, J.; Soetaert, W. Increasing the thermostability of sucrose phosphorylase by multipoint covalent immobilization. J. Biotechnol 2010a, 150, 125–130. [Google Scholar]
- Cerdobbel, A.; de Winter, K.; Desmet, T.; Soetaert, W. Sucrose phosphorylase as cross-linked enzyme aggregate: Improved thermal stability for industrial applications. Biotechnol. J 2010b, 5, 1192–1197. [Google Scholar]
- Sheldon, R.A. Cross-linked enzyme aggregates as industrial biocatalysts. Org. Process Res. Dev 2011, 15, 213–223. [Google Scholar]
- Cerdobbel, A.; de Winter, K.; Aerts, D.; Kuipers, R.; Joosten, H.-J.; Soetaert, W.; Desmet, T. Increasing the thermostability of sucrose phosphorylase by a combination of sequence- and structure-based mutagenesis. Protein Eng. Des. Sel 2011, 24, 829–834. [Google Scholar]
- Wildberger, P.; Luley-Goedl, C.; Nidetzky, B. Aromatic interactions at the catalytic subsite of sucrose phosphorylase: Their roles in enzymatic glucosyl transfer probed with Phe(52)→Ala and Phe(52)→Asn mutants. FEBS Lett 2011, 585, 499–504. [Google Scholar]
- Wiesbauer, J.; Goedl, C.; Schwarz, A.; Brecker, L.; Nidetzky, B. Substitution of the catalytic acid-base glu237 by gln suppresses hydrolysis during glucosylation of phenolic acceptors catalyzed by leuconostoc mesenteroides sucrose phosphorylase. J. Mol. Catal. B Enzym 2009, 65, 24–29. [Google Scholar]
- Stahl, M.; Jeppssonwistrand, U.; Mansson, M.O.; Mosbach, K. Induced stereo- and substrate selectivity of bioimprinted alpha-chymotrypsin in anhydrous organic media. J. Am. Chem. Soc 1991, 113, 9366–9368. [Google Scholar]
- Verheyen, E.; Schillemans, J.P.; van Wijk, M.; Demeniex, M.A.; Hennink, W.E.; van Nostrum, C.F. Challenges for the effective molecular imprinting of proteins. Biomaterials 2011, 32, 3008–3020. [Google Scholar]
- Vaidya, A.A.; Lele, B.S.; Kulkarni, M.G.; Mashelkar, R.A. Creating a macromolecular receptor by affinity imprinting. J. Appl. Polym. Sci 2001, 81, 1075–1083. [Google Scholar]
- Klein, J.U.; Whitcombe, M.J.; Mulholland, F.; Vulfson, E.N. Template-mediated synthesis of a polymeric receptor specific to amino acid sequences. Angew. Chem.-Int. Ed 1999, 38, 2057–2060. [Google Scholar]
- Peissker, F.; Fischer, L. Crosslinking of imprinted proteases to maintain a tailor-made substrate selectivity in aqueous solutions. Bioorgan. Med. Chem 1999, 7, 2231–2237. [Google Scholar]
- Kaulpiboon, J.; Pongsawasdi, P.; Zimmermann, W. Molecular imprinting of cyclodextrin glycosyltransferases from paenibacillus sp a11 and bacillus macerans with γ-cyclodextrin. FEBS J 2007, 274, 1001–1010. [Google Scholar]
- Vaidya, A.; Borck, A.; Manns, A.; Fischer, L. Altering glucose oxidase to oxidize d-galactose through crosslinking of imprinted protein. Chembiochem 2004, 5, 132–135. [Google Scholar]
- Wang, M.; Qi, W.; Jia, C.; Ren, Y.; Su, R.; He, Z. Enhancement of activity of cross-linked enzyme aggregates by a sugar-assisted precipitation strategy: Technical development and molecular mechanism. J. Biotechnol 2011, 156, 30–38. [Google Scholar]
- Desmet, T.; Soetaert, W. Enzymatic glycosyl transfer: Mechanisms and applications. Biocatal. Biotransform 2011, 29. [Google Scholar] [CrossRef]
- Kitao, S.; Yoshida, S.; Horiuchi, T.; Sekine, H.; Kusakabe, I. Formation of kojibiose and nigerose by sucrose phosphorylase. Biosci. Biotechnol. Biochem 1994, 58, 790–791. [Google Scholar]
- Aerts, D.; Verhaeghe, T.; de Mey, M.; Desmet, T.; Soetaert, W. A constitutive expression system for high-throughput screening. Eng. Life Sci 2011, 11, 10–19. [Google Scholar]
- Silverstein, R.; Voet, J.; Reed, D.; Abeles, R.H. Purification and mechanism of action of sucrose phosphorylase. J. Biol. Chem 1967, 242, 1338–1346. [Google Scholar]
- Waffenschmidt, S.; Jaenicke, L. Assay of reducing sugars in the nanomole range with 2,2′-bicinchoninate. Anal. Biochem 1987, 165, 337–340. [Google Scholar]
- Werner, W.; Rey, H.G.; Wielinge, H. Properties of a new chromogen for determination of glucose in blood according to god/pod-method. Z. Anal. Chem 1970, 252, 224–228. [Google Scholar]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the foling phenol reagent. J. Biol. Chem 1951, 193, 265–275. [Google Scholar]
- Cabirol, F.L.; Tan, P.L.; Tay, B.; Cheng, S.; Hanefeld, U.; Sheldon, R.A. Linum usitatissimum hydroxynitrile lyase cross-linked enzyme aggregates: A recyclable enantioselective catalyst. Adv. Synth. Catal 2008, 350, 2329–2338. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Imprinting molecule | [C] a(M) | Precipitation (%) | SPresidual b (%) |
|---|---|---|---|
| None | / | 97.8 | 1.2 |
| Sucrose | 0.2 | 96.4 | 1.6 |
| Sucrose | 0.5 | 52.1 | 38.5 |
| Sucrose | 1 | 40.6 | 56.8 |
| Glycerol | 0.5 | 98.2 | 1.1 |
| Glycerol | 1 | 96.6 | 1.3 |
| Glycerol | 2 | 97.6 | 1.4 |
| sucrose + glycerol | 0.5 + 2 | 68.9 | 25.6 |
| sucrose + glycerol | 0.2 + 2 | 96.6 | 1.5 |
| αGG | 0.2 | 97.3 | 1.4 |
| αGG | 1 | 96.4 | 1.2 |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
De Winter, K.; Soetaert, W.; Desmet, T. An Imprinted Cross-Linked Enzyme Aggregate (iCLEA) of Sucrose Phosphorylase: Combining Improved Stability with Altered Specificity. Int. J. Mol. Sci. 2012, 13, 11333-11342. https://doi.org/10.3390/ijms130911333
De Winter K, Soetaert W, Desmet T. An Imprinted Cross-Linked Enzyme Aggregate (iCLEA) of Sucrose Phosphorylase: Combining Improved Stability with Altered Specificity. International Journal of Molecular Sciences. 2012; 13(9):11333-11342. https://doi.org/10.3390/ijms130911333
Chicago/Turabian StyleDe Winter, Karel, Wim Soetaert, and Tom Desmet. 2012. "An Imprinted Cross-Linked Enzyme Aggregate (iCLEA) of Sucrose Phosphorylase: Combining Improved Stability with Altered Specificity" International Journal of Molecular Sciences 13, no. 9: 11333-11342. https://doi.org/10.3390/ijms130911333