1. Introduction
The regulation of the protein activity plays an important role in modulating cellular processes such as signal transduction, DNA replication, and metabolism [
1,
2]. Protein dysfunction is related to human diseases and disorders [
3–
7], and our ability to regulate enzyme functions and protein-protein interactions provides a promising strategy for therapy. Nanoparticles (NPs) have some advantages over small organic molecules. First, NPs have large specific surface areas for adequate protein binding and biological interactions [
1,
8]. Second, NP can enter cells easily [
10], in contrast to some small molecules and biological molecules. Third, there has been considerable progress in the synthesis of NPs with well controlled dimensions, geometry, and surface properties [
9], to complement the structural complexity of proteins [
11,
12]. Recent developments in nano materials offer a new pathway for controlling protein behavior through surface interactions.
In the past few years, NPs with different dimensions, composition and surface modification were investigated to understand their interactions with proteins. The hydrophobic interaction [
13–
15], π-π stacking [
16–
18] and electrostatic interaction [
19–
21] have been attributed to be major mechanisms of NP-protein interactions. These mechanisms often coexist.
This review, by summarizing recent research on nanoparticle/protein interactions, intends to emphasize the importance of such interactions in biological systems that may cause nanotoxicity issues, and the potential of such molecular recognition events in biomedical applications such as the diagnosis and treatment of human diseases.
2. Effects of Mixed Monolayer Protected Nano Clusters (MMPCs)
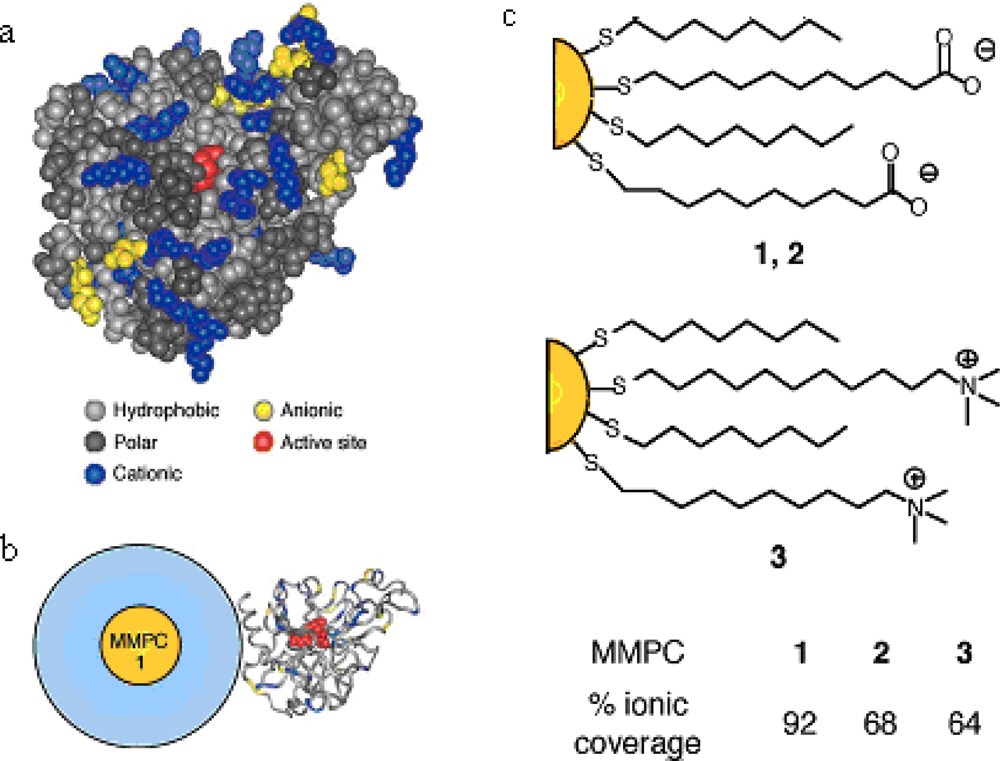
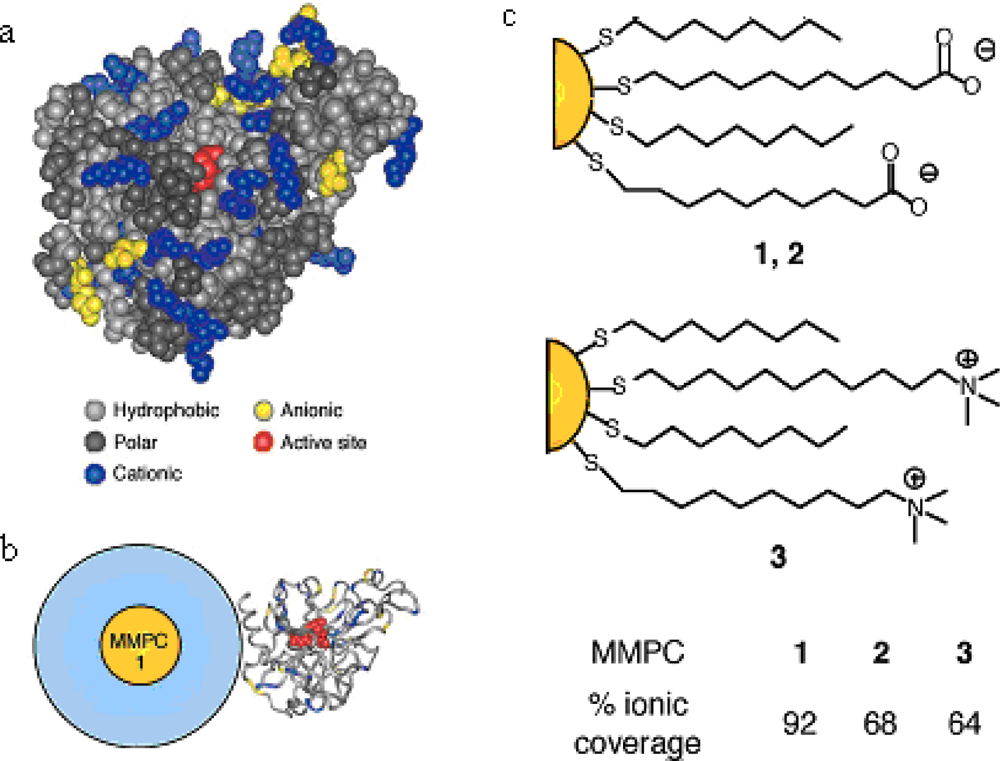
Mixed monolayer protected gold clusters (MMPCs) provide an effective scaffold for biomolecular binding. MMPCs were synthesized using the Brust reduction and Murray place-displacement reaction [
24] to fabricate additional functionalized thiols ended with carboxylate groups and amino groups, respectively [MMPCs
1,
2, and
3 in
Figure 1(c) featuring a 2-nm gold core, with an overall diameter of 6 nm] [
25]. These particles exhibited different effects on chymotrypsin (ChT) activity. The cationic MMPC
3 had no inhibition, while the anionic MMPC
1 and
2 were effective inhibitors of ChT because of the electrostatic complementarity between the carboxylate end groups and the hole of cationic residues located around the periphery of the active site, as shown in
Figure 1(b). Complete inhibition was observed at a 1:5 nanoparticle to ChT ratio.
The activity assay indicated that the inhibition of ChT by MMPCs was controlled by a two-stage mechanism featuring a fast reversible inhibition, followed by a slower irreversible process. Circular dichroism measurements of the complex demonstrated an almost complete denaturation of the enzyme over time. Dynamic light scattering studies confirmed that inhibition proceeded without substantial MMPC aggregation. The electrostatic nature of the engineered interactions provides a level of selectivity: little or no inhibition of functions of elastase, β-galactosidase, or cellular retinoic acid binding protein by MMPC was observed.
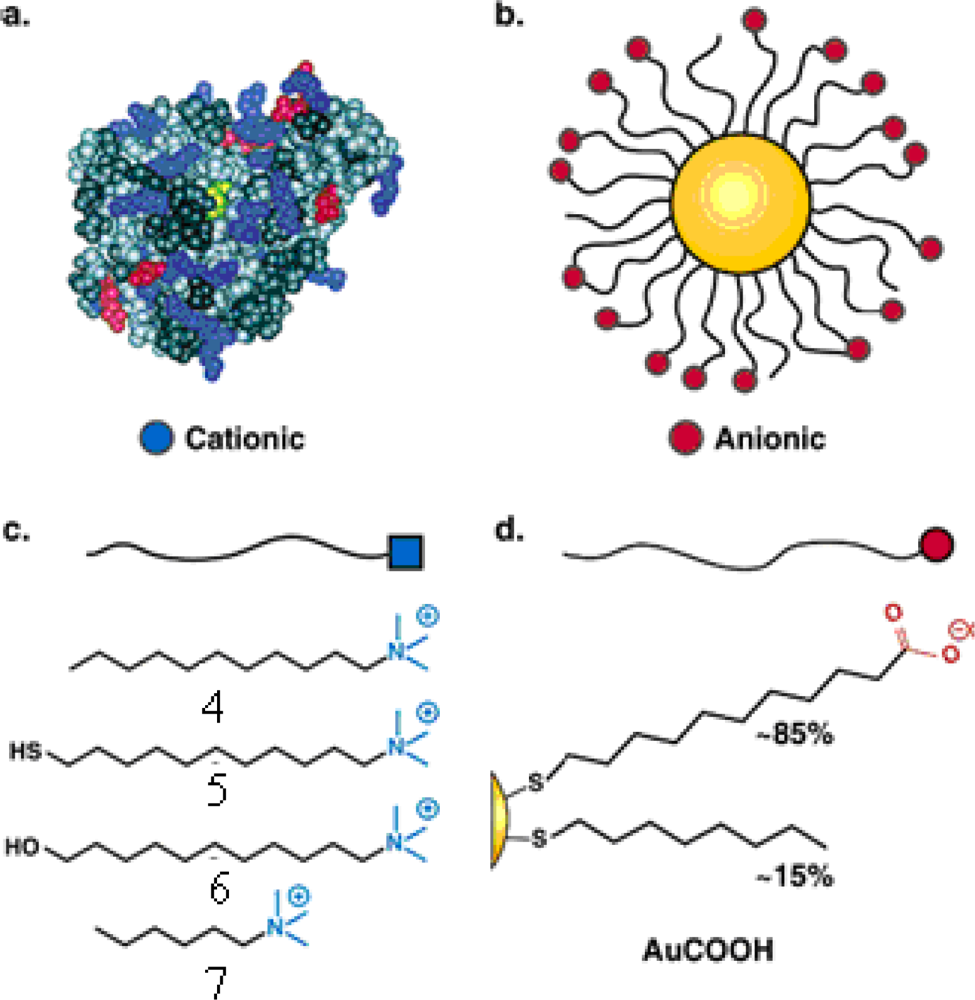
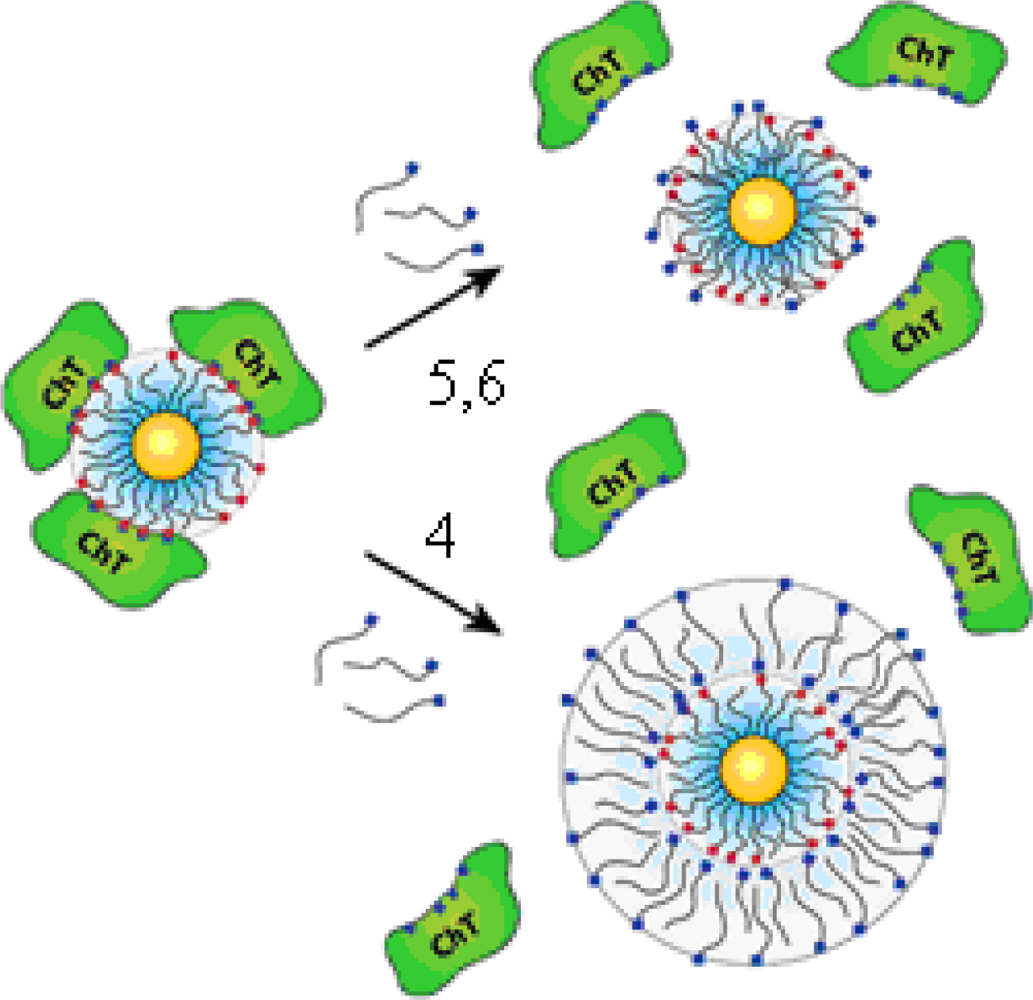
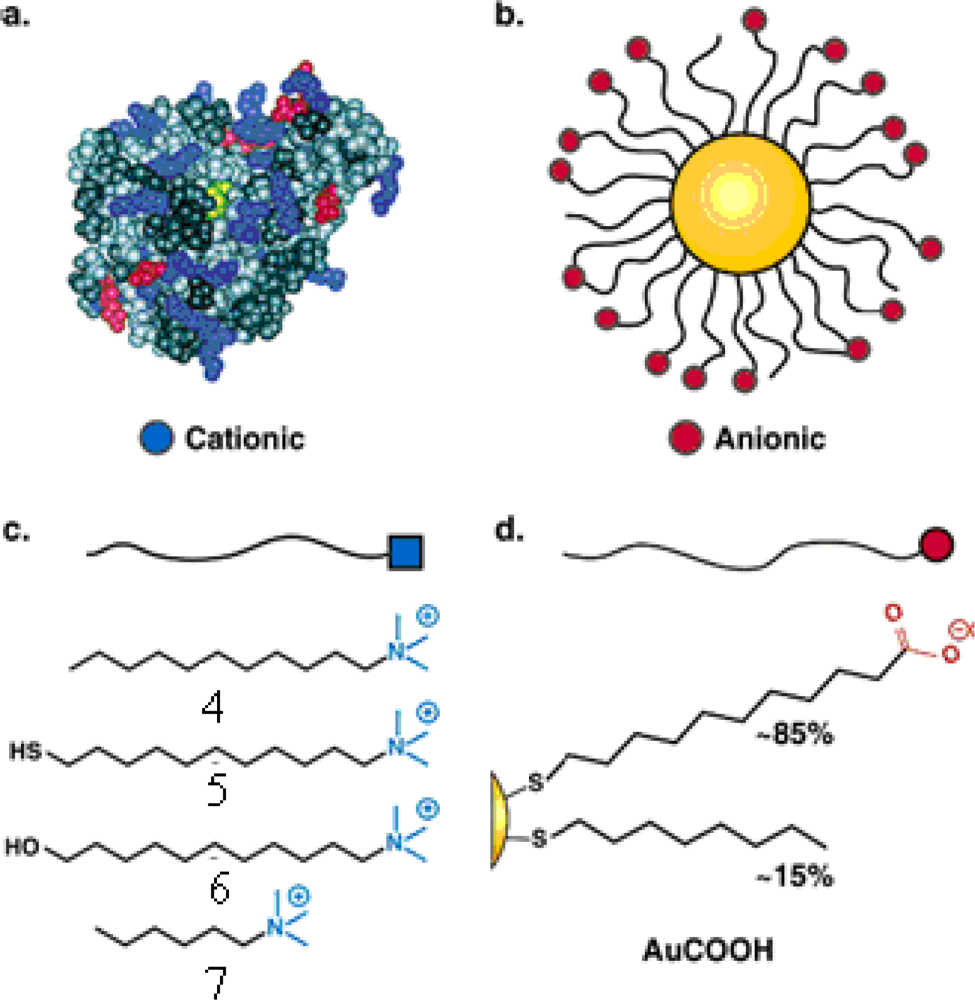
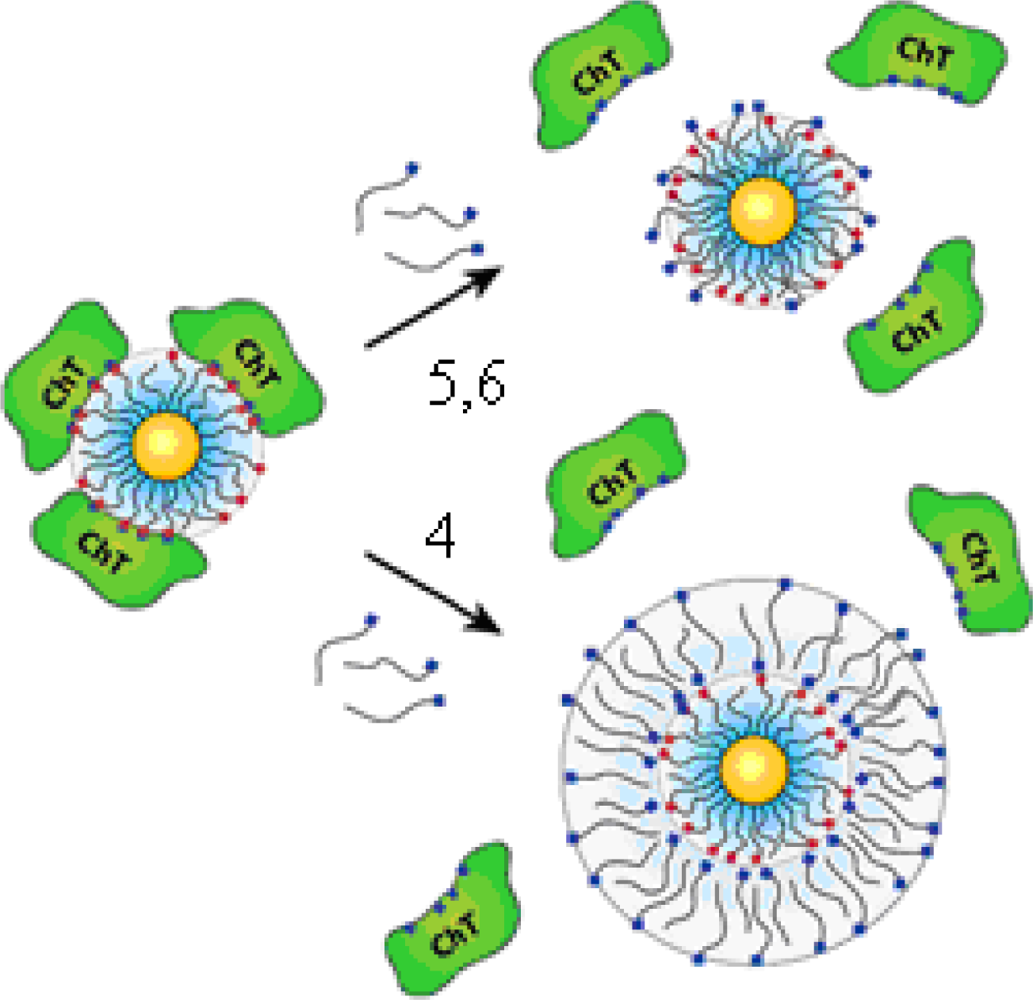
The “irreversible” inhibition of ChT can be reversed through modification of the anionic MMPC surface by the addition of cationic surfactants [
26]. Four derivatives of trimethylamine-functionalized surfactants were used to modifiy the gold nanoparticles (GNPs) with carboxylate endgroups (
4,
5,
6 and
7 in
Figure 2c, featuring a 2-nm gold core, with an overall diameter of 6 nm). Up to 50% of original ChT activity was rescued upon long-chain surfactant addition. Dynamic light scattering studies demonstrated that ChT released from the nanoparticle surface and the conformation characterization of the rescued ChT by fluorescence and fluorescence anisotropy indicated that ChT regained a high degree of native structure upon surfactant addition. The proposed mechanism of the ChT release: the MMPC
5 and MMPC
6 can elicit ChT release by interaction and/or partial displacement of the anionic monolayer. The MMPC
4, a bilayer-type structure, can release ChT when it envelops the MMPC surface, as shown schematically in
Figure 3. The noncovalent nature of the irreversible inhibition of ChT shows that the attenuation of the interactions the MMPC and protein could provide a means of rescuing enzyme activity.
3. Influence of the Size of NP on the Activity of Adsorbed Enzymes
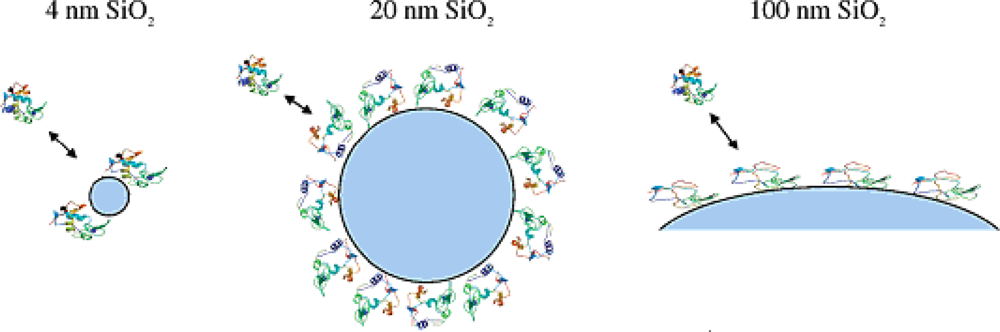
The size of NPs is one of the key parameters that influence the interaction between protein and NPs. Silica NPs with different diameters (4, 20 and 100 nm) were used to investigate the size influence on the structure and enzymatic activity of adsorbed lysozyme, whose dimensions are comparable in size to the 4 nm NPs [
27]. Both adsorption patterns and protein structure and function are strongly dependent on the size of the NPs. The formation of molecular complexes is observed for adsorption onto 4-nm silica. The CD results proved that the loss in α-helix content is strongly dependent on the size of the NPs. The great loss of α helicity was observed for the lysozyme adsorbed onto larger NPs. The activity of lysozyme adsorbed onto silica NPs is lower than that of the free protein, and the fraction of activity lost correlates well with the decrease in α-helix content. These results indicate that the size of the NP, perhaps because of the contributions of surface curvature, influences adsorbed protein structure and function (
Figure 4).
The same phenomenon was also observed when ribnuclease A [
28] and human carbonic anhydrase [
29] were absorbed onto silica NPs surfaces with different sizes. Urea denaturation analyses showed that the thermodynamic stability of ribnuclease A was decreased upon adsorption on to the NPs, with greater decrease on larger NPs. Therefore, the larger NPs tend to cause unfolding of adsorbed proteins [
28].
5. Regulation of Enzyme Function by Surface-Modified NPs
Compared with traditional materials, NPs have a larger surface area that can be modified with organic molecules through covalent or non-covalent modifications to form functionalized NPs. The suitably functionalized NPs possess the abilities that prinstine NPs do not have, such as preventing non-specific bindings and recognizing specific biomacromolecules. In this way, enzyme activities can be specifically regulated when bound to surface modified NPs.
Three silicon nanowires (SiNW) [
33], unmodified silicon nanowires (SiNW-SiO
2), SiNWs functionalized with carboxylic groups (SiNW-COOH) and highly reactive hydrogen modified SiNW (SiNW-H) were synthesized to investigate their affect on restriction endonucleases and Taq DNA polymerase. The PCR results showed that the inhibition of enzyme activity was determined by the functional groups on SiNWs in the order SiNW-H > SiNW-COOH > SiNW-SiO
2.
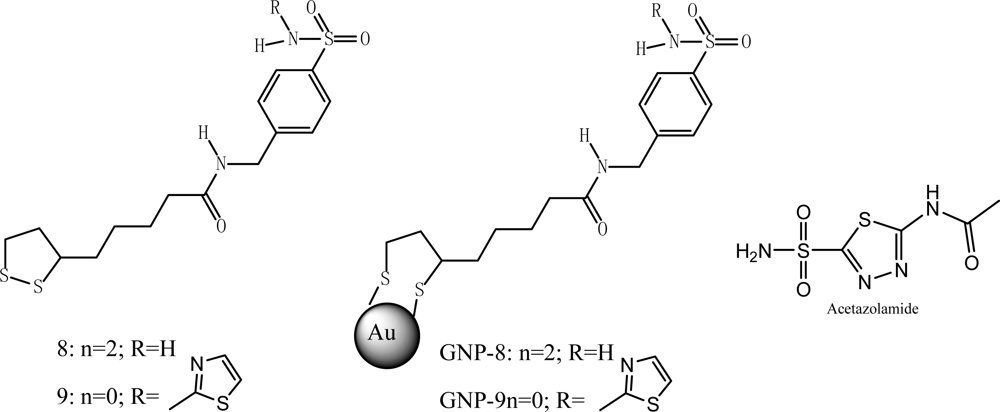
Specific NP/protein binding provides a better opportunity for regulating enzyme functions. Carbonic anhydrase inhibitor
8 linked to gold NPs (GNP
-8, with an average particle size of 3.3 nm, which corresponds to 720–724 Au atoms) showed excellent CA IX inhibition of the tumer-associated isoform and selectivity over hCA I and II (
Table 1,
Figure 6) [
34] due to the fact that their binding pocket contains different residues.
Table 1 listed the inhibition of CA I, II and IX
a by acetazolamide (AZA), sulfonamides
8,
9,
GNP-8 and
GNP-9. It has been also shown that molecules such as antibodies, which can recognize protein specifically, can be linked to NPs to recognize proteins through specific antibody-antigen interactions [
35,
36].
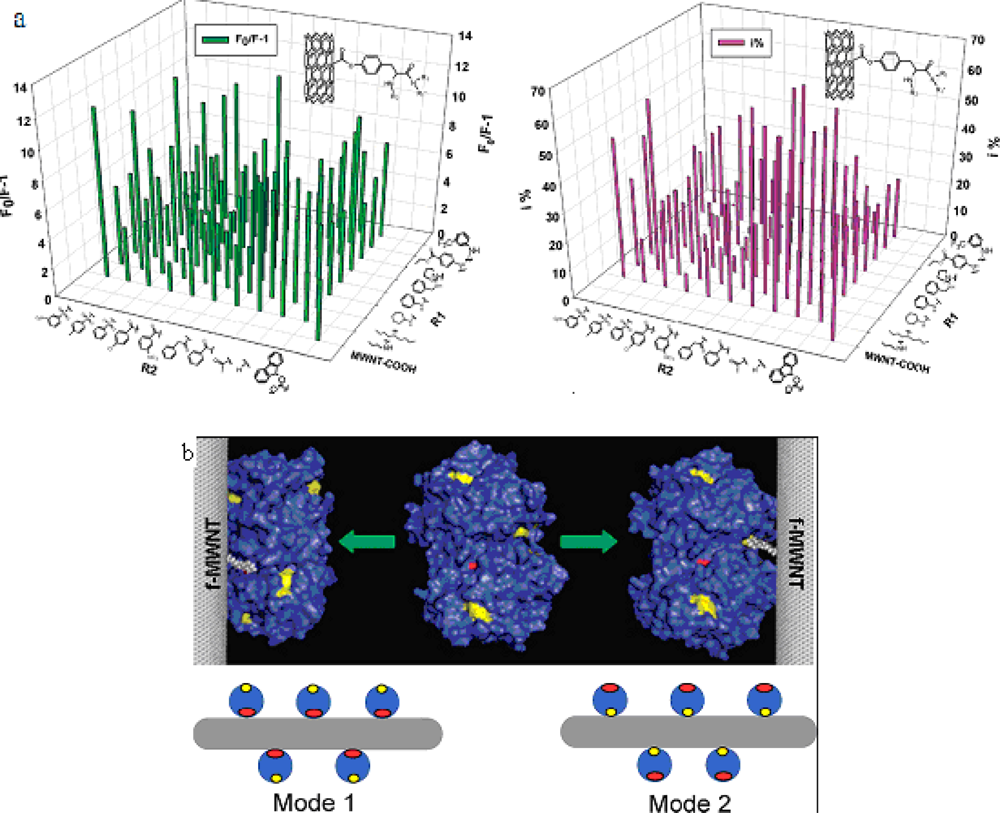
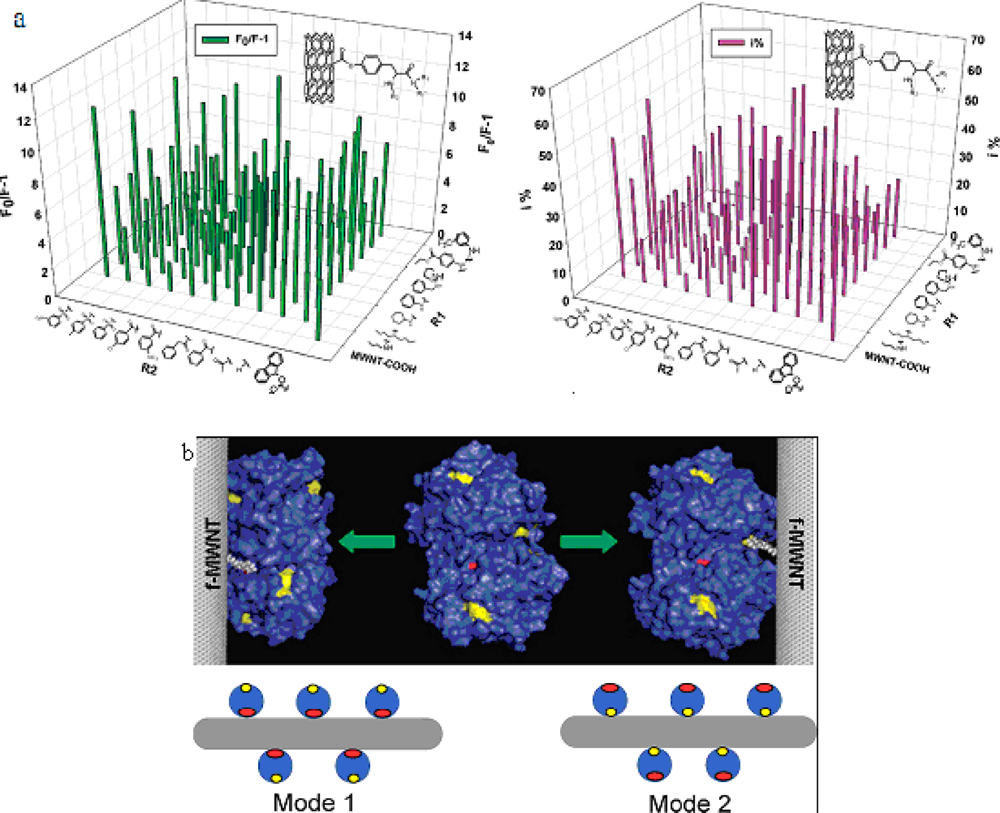
We have synthesized a novel surface-modified multi-wall carbon nanotube combinatorial library containing 80 members by using
in silico design and combinatorial synthesis [
37]. By screening the library we discovered MWNTs with reduced protein binding, reduced cytotoxicity, and immune response. The ChT enzymatic activity and fluorescent quenching were measured by incubating ChT with functionalized multiwalled carbon nanotube (f-MWNTs, were synthesized by chemical vapor deposition. The purity was 95% and the catalyst residue was less than 0.2%. They are 40 nm in diameter and the length is 500–2,000 nm) (
Figure 7A). By screening the library [
38], we discovered four f-MWNTs that can bind to the catalytic site of ChT site-specifically and inhibit its enzymatic activity competitively (
Figure 7B). Our results demonstrated that specific recognition of ChT and regulation of its functions by surface-modified MWNT.
6. Fine Tuning of NP/Protein Interactions
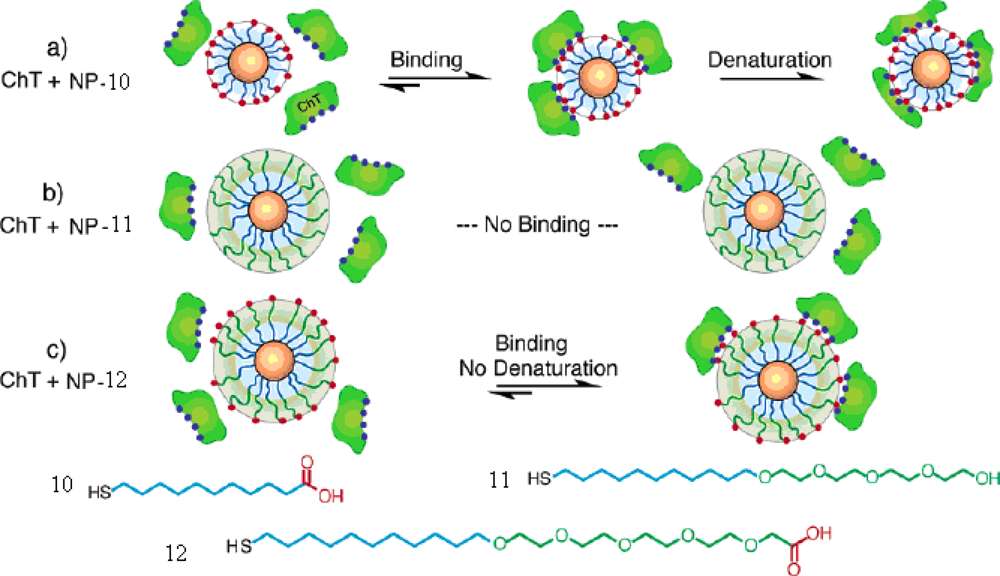
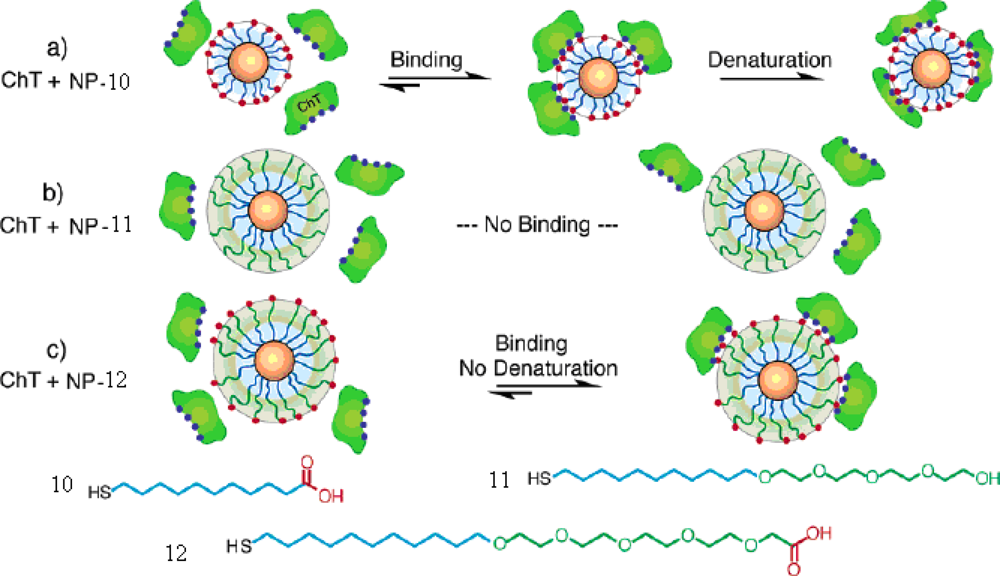
CdSe particles modified with thioalkylated oligo(ethylene glycol) and chain-end carboxylate were synthesized [
39]. The study of the interactions between CdSe NPs and chymotrypsin revealed three levels of inhibition: (1) protein inhibition and denaturation with
NP-10, (2) no protein binding with
NP-11, and (3) inhibition and retention of the protein structure with
NP-12. The
NP-10 with alkanethiol-carboxylate functionalized can bind, inhibit, and denaturate of ChT because of the hydrophobic interactions. The
NP-11, lacking of the carboxylate recognition element, was inert in protein binding. The
NP-12, modified with tetraethylene glycol spacer between the alkyl chain and recognition element, can inhibit ChT reversibly because of the electrostatic interaction between the particle and protein, but prevents hydrophobic interactions caused by the interior alkyl chain (
Figure 8).
In order to explore how the linkages between recognition elements and NP core affect the NP/enzyme interacion. A series of
l-amino acid functionalized GNPs with oligo (ethylene glycol) tethers of varying length were studied. It has been shown that amino acid side chains can maintain the ChT structure while the alkyl chains denature the protein as a result of nonspecific hydrophobic interactions [
40].
Structure diversity can also be generated by introducing amino acids in the surface modification. The hydrophobic interaction and the complementary electrostatic interactions between Au nanoparticles with
l-amino acids as endgroups and ChT also plays an important role in regulating ChT activity [
41].
Protein-protein recognition is a key aspect of the complex cellular functions, such as apoptosis and angiogenesis. Control over interprotein recognition holds the potential in therapeutic applications. Thiolates with biocompatible PEG linker and trimethyl-amine end group were used to functionalize Au nanoparticles. And it can inhibit interactions between cytochrome c and cytochrome c perxidase in the low nM concentration range [
42].
7. Conclusions
Enzyme dysfunction is related to human diseases. It is desirable to be able to regulate enzyme conformation and function. Nano sensors incorporating enzymes also require that protein conformation is not altered, further emphasizing the importance of protein regulation. NPs can be selected to specifically bind enzymes and control their functions after surface modifications. Such NP/protein interactions can be fine-tuned to maintain protein structure or alter it on purpose.
Acknowledgments
This work was supported by Shandong University, the American Lebanese Syrian Associated Charities (ALSAC) and St. Jude Children’s Research Hospital.
References
- Gadek, TR; Nicholas, JB. Small molecule antagonists of proteins. Biochem. Parmacol 2003, 65, 1–8. [Google Scholar]
- Toogood, PL. Inhibition of protein-protein association by small molecules: Approaches and progress. J. Med. Chem 2002, 45, 1543–1558. [Google Scholar]
- Armbruster, BN; Banik, SSR; Guo, CH; Smith, AC; Counter, C. N-Terminal domains of the human telomerase catalytic subunit required for enzyme activity in vivo. Mol. Cell. Biol 2001, 21, 7775–7786. [Google Scholar]
- Shultz, MD; Ham, Y-W; Lee, S-G; Davis, DA; Brown, C; Chemielewski, J. Small-molecule dimerization inhibitors of wild-type and mutant HIV protease: A focused library approach. J. Am. Chem. Soc 2004, 126, 9886–9887. [Google Scholar]
- Longthorne, VL; Williams, GT. Caspase activity is required for commitment to fas-mediated apoptosis. EMBO J 1997, 16, 3805–3812. [Google Scholar]
- Singh, I. Biochemistry of peroxisomes in health and disease. Mol. Cell. Biochem 1997, 167, 1–29. [Google Scholar]
- D’Armiento, J. Matrix metalloproteinase disruption of the extracellular matrix and cardiac dysfunction. Trends Cardiovasc. Med 2002, 12, 97–101. [Google Scholar]
- Erkizan, HV; Kong, YL; Merchant, M; Schlottmann, S; Barber-Rotenberg, JS; Yuan, LS; Abaan, OD; Chou, T-H; Dakshanamurthy, S; Brown, ML; Ueren, A; Toretsky, JA. A small molecule blocking oncogenic protein EWS-FLI1 interaction with RNA helicase A inhibits growth of Ewing’s sarcoma. Nature Med 2009, 15, 750–757. [Google Scholar]
- Lo Conte, L; Chothia, C; Janin, J. The atomic structure of protein-protein recognition sites. Mol. Bio 1999, 285, 2177–2198. [Google Scholar]
- Porter, AE; Gass, M; Muller, K; Skepper, JN; Midgley, PA; Welland, M. Direct imaging of single-walled carbon nanotubes in cells. Nat. Nanotechnol 2007, 2, 713–717. [Google Scholar]
- Mu, QX; Liu, W; Xing, YH; Zhou, HY; Li, ZW; Zhang, Y; Ji, LH; Wang, F; Si, ZK; Zhang, B; Yan, B. Protein binding by functionalized multiwalled carbon nanotubes is governed by the surface chemistry of both parties and the nanotube diameter. J. Phys. Chem. C 2008, 112, 3300–3307. [Google Scholar]
- Peng, XG; Manna, L; Yang, WD; Wickham, J; Scher, E; Kadavanich, A; Alivisatos, AP. Shape control of CdSe nanocrystals. Nature 2000, 404, 59–61. [Google Scholar]
- Strano, MS; Dyke, CA; Usrey, ML; Barone, PW; Allen, MJ; Shan, HW; Kittrell, C; Hauge, RH; Tour, JM; Smalley, RE. Electronic structure control of single-walled carbon nanotube functionalization. Science 2003, 301, 1519–1522. [Google Scholar]
- You, CC; Arvizo, RR; Rotello, VM. Regulation of α-chymotrypsin activity on the surface of substrate-functionalized gold nanoparticles. Chem Comm 2006, 2905–2907. [Google Scholar]
- Erlanger, BF; Chen, BX; Zhu, M; Brus, L. Binding of an anti-fullerene IgG monoclonal antibody to single wall carbon nanotubes. Nano Lett 2001, 1, 465–467. [Google Scholar]
- Lin, Y; Allard, LF; Sun, Y-P. Protein-affinity of single-walled carbon nanotubes in water. J. Phy. Chem. B 2004, 108, 3760–3764. [Google Scholar]
- Stina, L; Lynch, I; Eva, T. Systematic investigation of the thermodynamics of HSA adsorption to N-iso-propylacrylamine/N-tert-butylacrylamide copolymer nanoparticles. Effects of particle size and hydrophobicity. Nano Lett 2007, 7, 914–920. [Google Scholar]
- Li, XJ; Chen, W; Zhan, QW; Dai, LM. Direct measurements of interactions between polypeptides and carbon nanotubes. J. Phys. Chem. B 2006, 110, 12621–12625. [Google Scholar]
- Irle, S; Zheng, GS; Elstner, M; Morokuma, K. Formation of fullerene molecules from carbon nanotubes: A quantum chemical molecular dynamics study. Nano Lett 2003, 3, 465–470. [Google Scholar]
- Chen, RJ; Zhang, YG; Wang, DW; Dai, HJ. Noncovalent sidewall functionalization of single-walled carbon nanotubes for protein immobilization. J. Am. Chem. Soc 2001, 123, 3838–3839. [Google Scholar]
- Hong, R; Emrick, T; Rotello, VM. Monolayer-controlled substrate selectivity using noncovalent enzyme-nanoparticle conjugates. J. Am. Chem. Soc 2004, 126, 13572–13573. [Google Scholar]
- You, C-C; Arvizo, RR; Rotello, VM. Regulation of α-chymotrypsin activity on the surface of substrate-functionalized gold nanoparticles. Chem Comm 2006, 2905–2907. [Google Scholar]
- Bradley, K; Briman, M; Star, A; Gruner, G. Charge transfer from adsorbed proteins. Nano Lett 2004, 4, 253–256. [Google Scholar]
- Hostetler, MJ; Green, SJ; Stoke, JJ; Murray, RW. Monolayers in three dimensions: Synthesis and electrochemistry of ω-functionalized alkanethiolate-stabilized gold clusters compound. J. Am. Chem. Soc 1996, 118, 4212–4213. [Google Scholar]
- Fischer, NO; Mclntosh, CM; Simard, JM; Rotello, VM. Inhibition of chymotrypsin through surface binding using nanoparticle-based receptors. Proc. Nat. Acad. Sci. USA 2002, 99, 5018–5023. [Google Scholar]
- Fischer, NO; Verma, A; Goodman, CM; Simard, JM; Rotello, VM. Reversible “irreversible” inhibition of chymotrypsin using nanoparticle receptors. J. Am. Chem. Soc 2003, 125, 13387–13391. [Google Scholar]
- Vertegel, AA; Siegel, RW; Dordick, JS. Silica nanoparticle size influences the structure and enzymatic activity of adsorded lysozyme. Langmuir 2004, 20, 6800–6807. [Google Scholar]
- Shang, W; Nuffer, HH; Dordick, JS; Siegel, RW. Unfolding of ribonuclease A on silica nanoparticle surfaces. Nano Lett 2007, 7, 1991–1995. [Google Scholar]
- Lundqvist, M; Sethson, I; Jonsson, BH. Protein adsorption onto silica nanoparticles: Conformational changes depend on the particle’s curvature and the protein stability. Langmuir 2004, 20, 10639–10647. [Google Scholar]
- Asuri, P; Bale, SS; Pangule, RC; Shah, DA; Kane, RS; Dordick, JS. Structure, function, and stability of enzymes covalently attached to single-walled carbon nanotube. Langmuir 2007, 23, 12318–12321. [Google Scholar]
- Karajanagi, SS; Vertegel, AA; Kane, RS; Dordick, JS. Structure and function of enzymes adsorbed onto single-walled carbon nanotubes. Langmuir 2004, 20, 11594–11599. [Google Scholar]
- Lin, Y; Allard, LF; Sun, YP. Protein-affinity of single-walled carbon nanotubes in water. J. Phys. Chem. B 2004, 108, 3760–3764. [Google Scholar]
- Yi, CQ; Fong, C-C; Chen, WW; Qi, SJ; Lee, S-T; Yang, MS. Inhibition of biochemical reactions by silica nanowires through modulating enzyme activities. ChemBioChem 2007, 8, 1225–1229. [Google Scholar]
- Stiti, M; Cecchi, A; Rami, M; Abdaoui, M; Barragan-Montero, V; Scozzafava, A; Guari, Y; Winum, J-Y; Supuran, CT. Carbonic anhydrase inhibitor coated gold nanoparticles selectively inhibit the tumor-associated isoform IX over the cytosolic lsozymes I and II. J. Am. Chem. Soc 2008, 130, 16130–16131. [Google Scholar]
- Lu, FS; Gu, LR; Meziani, MJ; Wang, X; Luo, PJ; Veca, LM; Cao, L; Sun, Y-P. Advances in bioapplications of carbon nanotubes. Adv. Mater 2009, 21, 139–152. [Google Scholar]
- Tara Elkin, T; Jiang, XP; Taylor, S; Lin, Y; Gu, LR; Yang, H; Brown, J; Collins, S; Sun, Y-P. Immuno-carbon nanotubes and recognition of pathogens. ChemBioChem 2005, 6, 640–643. [Google Scholar]
- Zhou, HY; Mu, QX; Gao, NN; Liu, AF; Xing, YH; Gao, SL; Zhang, Q; Qu, GB; Chen, YY; Liu, G; Zhang, B; Yan, B. A nano-combinatorial library strategy for the discovery of nanotubes with reduced protein-binding, cytotoxicity, and immune response. Nano Lett 2008, 8, 859–865. [Google Scholar]
- Zhang, B; Xing, YH; Li, ZW; Zhou, HY; Mu, QX; Yan, B. Functionalized carbon nanotubes specifically bind to chymotrypsin catalytic site and regulate its enzymatic function. Nano Lett 2009, 9, 2280–2284. [Google Scholar]
- Hong, R; Fishcer, NO; Verma, A; Goodman, CM; Emrick, T; Retello, VM. Control of protein structure and function through surface recognition by tailored nanopartcile scaffolds. J. Am. Chem. Soc 2004, 126, 739–743. [Google Scholar]
- You, C-C; De, M; Rotello, VM. Contrasting effect of exterior and interior hydrophobic moieties in the complexation of amino acid functionalized gold clusters with α-chymotrypsin. Org. Lett 2005, 7, 5685–5688. [Google Scholar]
- You, C-C; De, M; Han, G; Rotello, VM. Tunable inhibition and denaturation of α-chymotrypsin with amino acid-functionalized gold nanoparticles. J. Am. Chem. Soc 2005, 127, 12873–12881. [Google Scholar]
- Bayraktar, H; Ghosh, PS; Rotello, VM; Knapp, MJ. Disruption of protein-protein interactions using nanopartciles: Inhibition of cytochrome C peroxidase. Chem Commun 2006, 1390–1392. [Google Scholar]
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}