Strategy for Adapting Wine Yeasts for Bioethanol Production

Abstract
:1. Introduction
2. Experimental Section
2.1. Yeast Strains and Growth Media
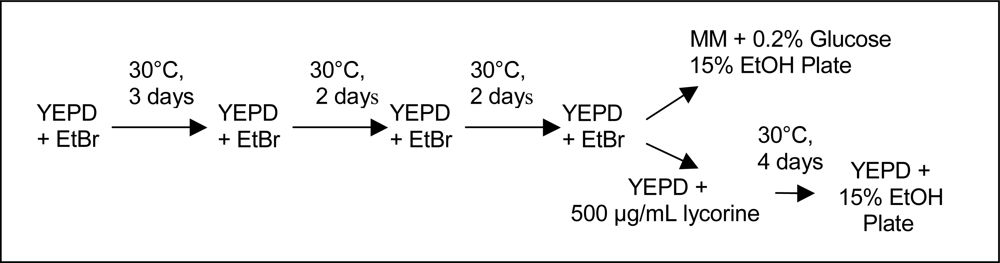
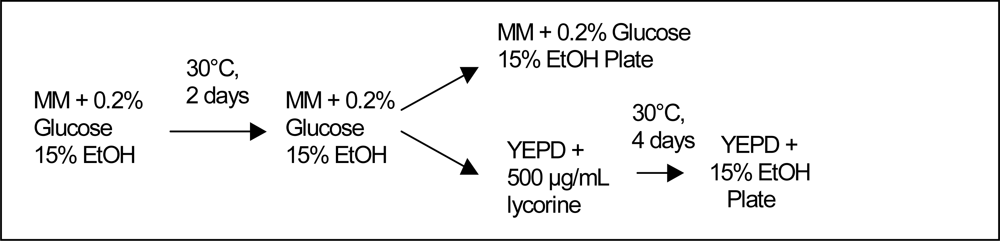
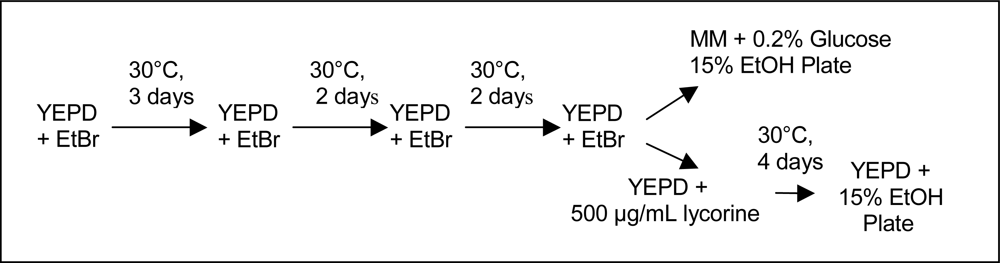
2.2. Culture Methods
2.3. Fermentation Method
2.4. Analysis of Ethanol and Acetic Acid
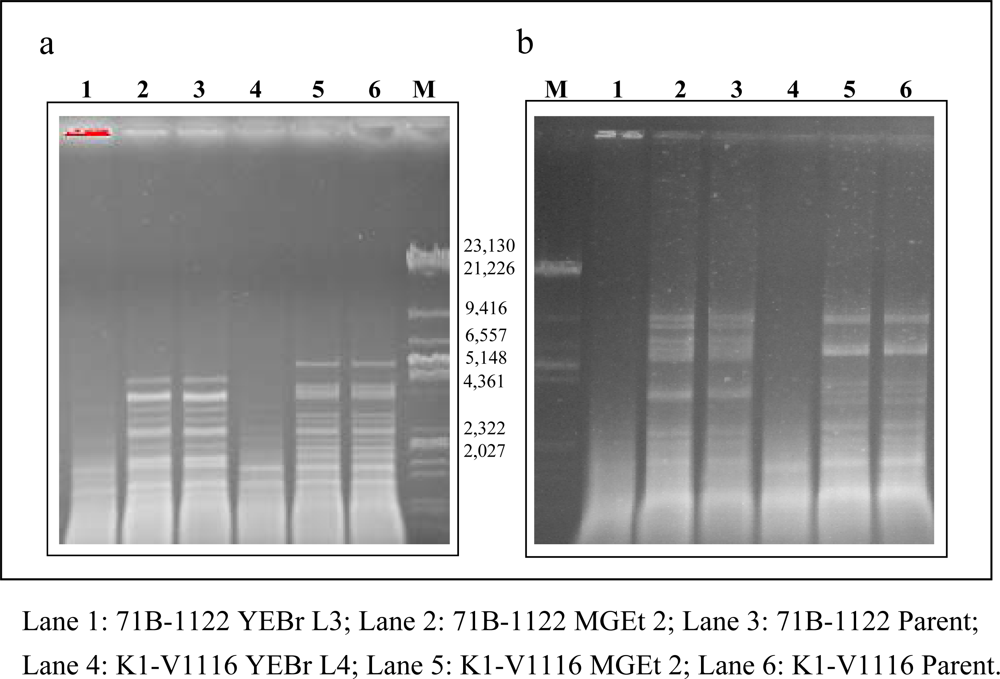
2.5. Analysis of mtDNA by Gel Electrophoresis
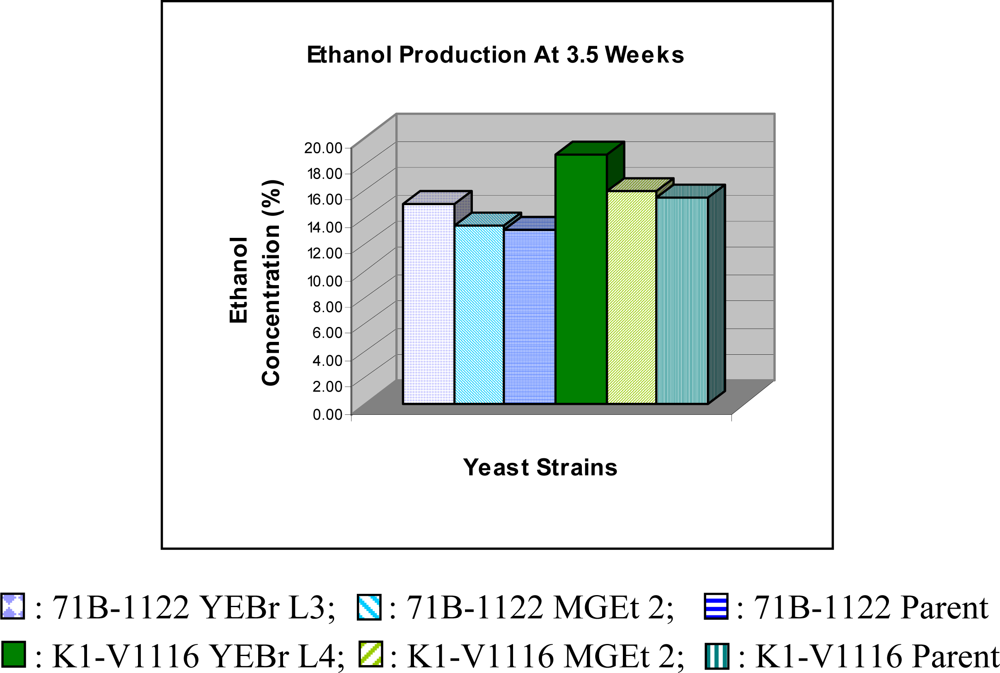
3. Results and Discussion
4. Conclusions
Acknowledgments
References
- Economic Impact of the Ethanol Industry Fact Sheet ( http://www.drivingethanol.org/userdocs/facts/2007/2007EconomicImpact-Special.pdf), accessed November 2008.
- Louime, C; Uckelmann, H. Cellulosic ethanol: Securing the planet future energy needs. Int. J. Mol. Sci 2008, 9, 838–841. [Google Scholar]
- Mielenz, JR. Ethanol production from biomass: technology and commercialization status. Curr. Opin. Microbiol 2001, 4, 324–329. [Google Scholar]
- Demirbas, A. Producing and using bioethanol as an automotive fuel. Energy Sources Part B 2007, 2, 391–401. [Google Scholar]
- Ooi, BG; Wanamaker, LE; Markuszewski, BM; Chong, NS. Genetic and enological analysis of selected Saccharomyces cerevisiae strains for wine production. Int. J. Food Sci. Technol 2008, 43, 1111–1120. [Google Scholar]
- Ibeas, JI; Jimenez, J. Mitochondrial DNA loss caused by ethanol in Saccharomyces flor yeasts. Appl. Environ. Microbiol 1997, 63, 7–12. [Google Scholar]
- Goldring, ES; Grossman, LI; Krunpnick, D; Cryer, DR; Marmur, J. The petite mutation in yeast. Loss of mitochondrial deoxyribonucleic acid during induction of petites with ethidium bromide. J. Mol. Biol 1970, 52, 323–335. [Google Scholar]
- Slonimski, PP; Perrodin, G; Croft, JH. Ethidium bromide induced mutation of yeast mitochondria: complete transformation of cells into respiratory deficient non-chromosomal “petites”. Biochem. Biophys. Res. Commun 1968, 30, 232–239. [Google Scholar]
- Bandas, EL; Zakharov, IA. Induction of rho− mutations in yeast Saccharomyces cerevisiae by ethanol. Mutat. Res 1980, 71, 193–199. [Google Scholar]
- Ristow, H; Seyfarth, A; Lochmann, ER. Chromosomal damages by ethanol and acetaldehyde in Saccharomyces cerevisiae as studied by pulsed field gel electrophoresis. Mutat. Res 1995, 326, 165–170. [Google Scholar]
- Del Giudice, L; Massardo, DR; Manna, F; Evidente, A; Randazzo, G; Wolf, K. Differential effect of alkaloid on rho+, mit−, rho−, and rho0 strains of Saccharomyces cerevisiae. Curr. Genet 1984, 8, 493–498. [Google Scholar]
- Farrant, TJ. One way analysis of variance. In Practical Statistics for the Analytical Scientist-A Bench Guide; Thomas Graham House: Cambridge, 1997; pp. 21–23. [Google Scholar]
- Querol, A; Barrio, E; Huerta, T; Ramon, D. Molecular monitoring of wine fermentations conducted by active dry yeast strains. Appl. Environ. Microbiol 1992, 58, 2948–2953. [Google Scholar]
- Del Giudice, L; Massardo, DR; Pontieri, P; Wolf, K. Interaction between yeast mitochondrial and nuclear genomes: Null alleles of RTG genes affect resistance to the alkaloid lycorine in rho0 petites of Saccharomyces cerevisiae. Gene 2005, 354, 9–14. [Google Scholar]
- Del Giudice, L; Massardo, DR; Manna, F; Wolf, K. Isolation and genetical and biochemical characterization of mutants resistant to the alkaloid lycorine. Curr. Genet 1986, 11, 247–249. [Google Scholar]
- Butow, RA. Cellular responses to mitochondrial dysfunction: It’s not always downhill. Cell Death Differ 2002, 9, 1043–1045. [Google Scholar]
- Massardo, DR; Zweifel, SG; Gunge, N; Miyakawa, I; Sando, N; Del Giudice, A; Wolf, K; Del Giudice, L. Use of lycorine and DAPI staining in Saccharomyces cerevisiae to differentiate between rho0 and rho− cells in a cce1/Δcce1 nuclear background. Can. J. Microbiol 2000, 46, 1058–1065. [Google Scholar]
- Ferguson, RF; von Borstel, RC. Induction of the cytoplasmic ‘petite’ mutation by chemical and physical agents in Saccharomyces cerevisiae. Mutat. Res 1992, 265, 103–148. [Google Scholar]
- Martinez, P; Codon, AC; Perez, L; Benitez, T. Physiological and molecular characterization of flor yeasts: polymorphism of flor yeast populations. Yeast 1995, 11, 1399–1411. [Google Scholar]
- Aguilera, A; Benitez, T. Role of mitochondria in ethanol tolerance of Saccharomyces cerevisiae. Arch. Microbiol 1985, 142, 389–392. [Google Scholar]
- Jimenez, J; Benitez, T. Yeast cell viability under conditions of high temperature and ethanol concentrations depends on the mitochondrial genome. Curr. Genet 1988, 13, 461–469. [Google Scholar]
- van Uden, N. Ethanol toxicity and ethanol tolerance in yeasts. Annu. Rep. Ferment. Proc 1985, 8, 11–58. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Yeast Strains | 71B-1122 YEBr L3 | 71B-1122 MGEt 2 | 71B-1122 MGEt L3 | 71B-1122 MGEt L4 | 71B-1122 Parent | K1-V1116 YEBr L4 | K1-V1116 MGEt 2 | K1-V1116 Parent |
|---|---|---|---|---|---|---|---|---|
| Ethanol % (v/v) | 12.3 ± 1.0 | 11.6 ± 0.7 | 11.2 ± 0.6 | 11.7 ± 1.4 | 12.1 ± 0.9 | 12.5 ± 1.5 | 12.3 ± 1.8 | 12.4 ± 0.6 |
| Acetic Acid % (v/v) | 0.09 ± 0.02 | 0.10 ± 0.01 | 0.09 ± 0.01 | 0.10 ± 0.02 | 0.11 ± 0.02 | 0.07 ± 0.00 | 0.11 ± 0.01 | 0.11 ± 0.01 |
| Yeast Strains | 71B-1122 YEBr L3 | 71B-1122 MGEt 2 | 71B-1122 Parent | K1-V1116 YEBr L4 | K1-V1116 MGEt 2 | K1-V1116 Parent | ANOVA F valuea |
|---|---|---|---|---|---|---|---|
| Ethanol % (v/v) (3.5 weeks) | 15.0 ± 0.3 | 13.4 ± 0.8 | 13.1 ± 0.7 | 18.8 ± 0.8 | 16.0 ± 0.6 | 15.5 ± 0.5 | 33.53 |
| Ethanol % (v/v) (4 weeks) | 15.4 ± 0.4 | 12.2 ± 1.5 | 11.8 ± 1.2 | 19.4 ± 0.5 | 16.6 ± 1.0 | 15.2 ± 0.4 | 27.02 |
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/). This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ooi, B.G.; Lankford, K.R. Strategy for Adapting Wine Yeasts for Bioethanol Production. Int. J. Mol. Sci. 2009, 10, 385-394. https://doi.org/10.3390/ijms10010385
Ooi BG, Lankford KR. Strategy for Adapting Wine Yeasts for Bioethanol Production. International Journal of Molecular Sciences. 2009; 10(1):385-394. https://doi.org/10.3390/ijms10010385
Chicago/Turabian StyleOoi, Beng Guat, and Kevin R. Lankford. 2009. "Strategy for Adapting Wine Yeasts for Bioethanol Production" International Journal of Molecular Sciences 10, no. 1: 385-394. https://doi.org/10.3390/ijms10010385