Berberine Activates Aryl Hydrocarbon Receptor but Suppresses CYP1A1 Induction through miR-21-3p Stimulation in MCF-7 Breast Cancer Cells

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
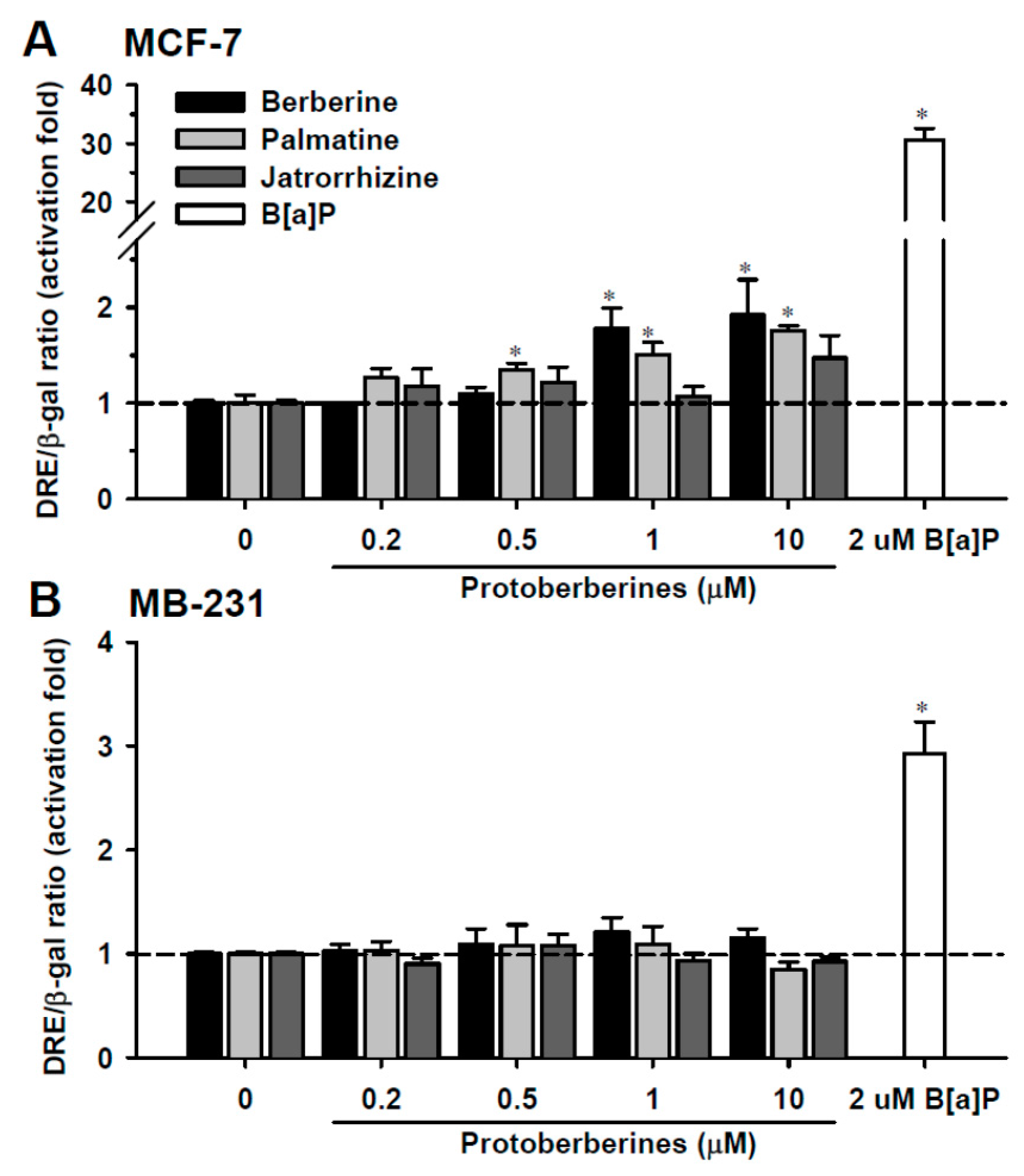
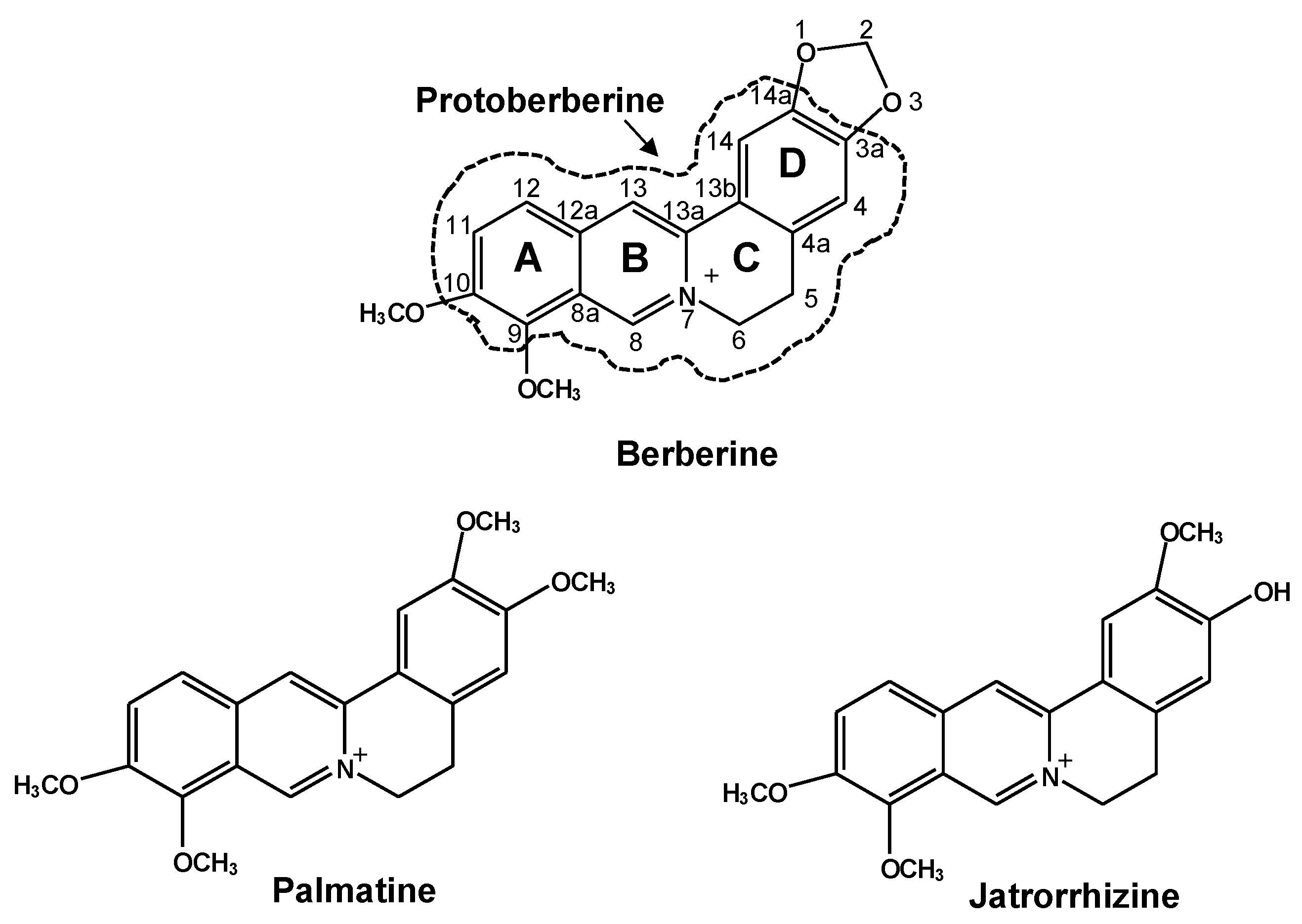
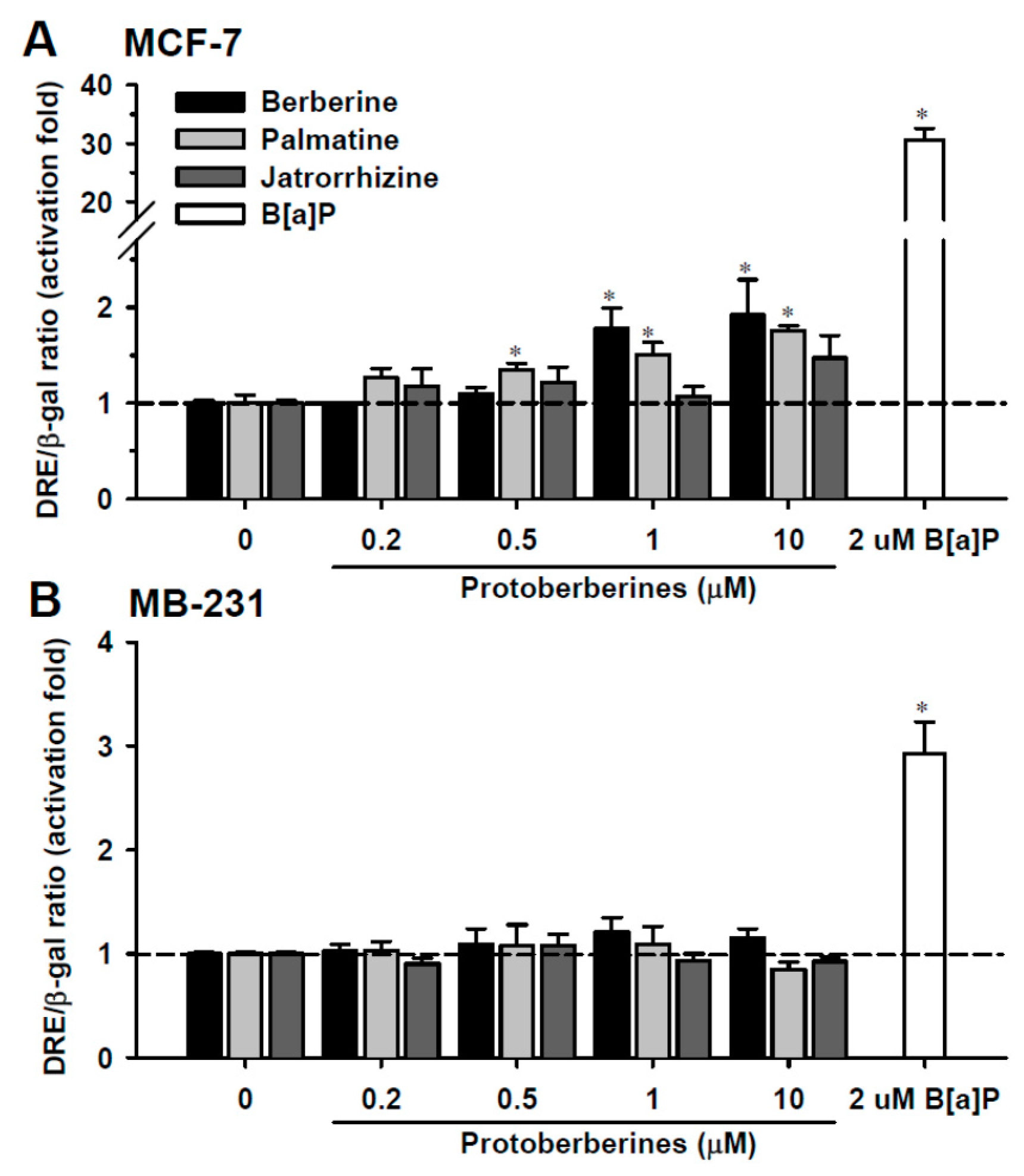
2.1. DRE Activation by Berberine, Palmatine and Jatrorrhizine
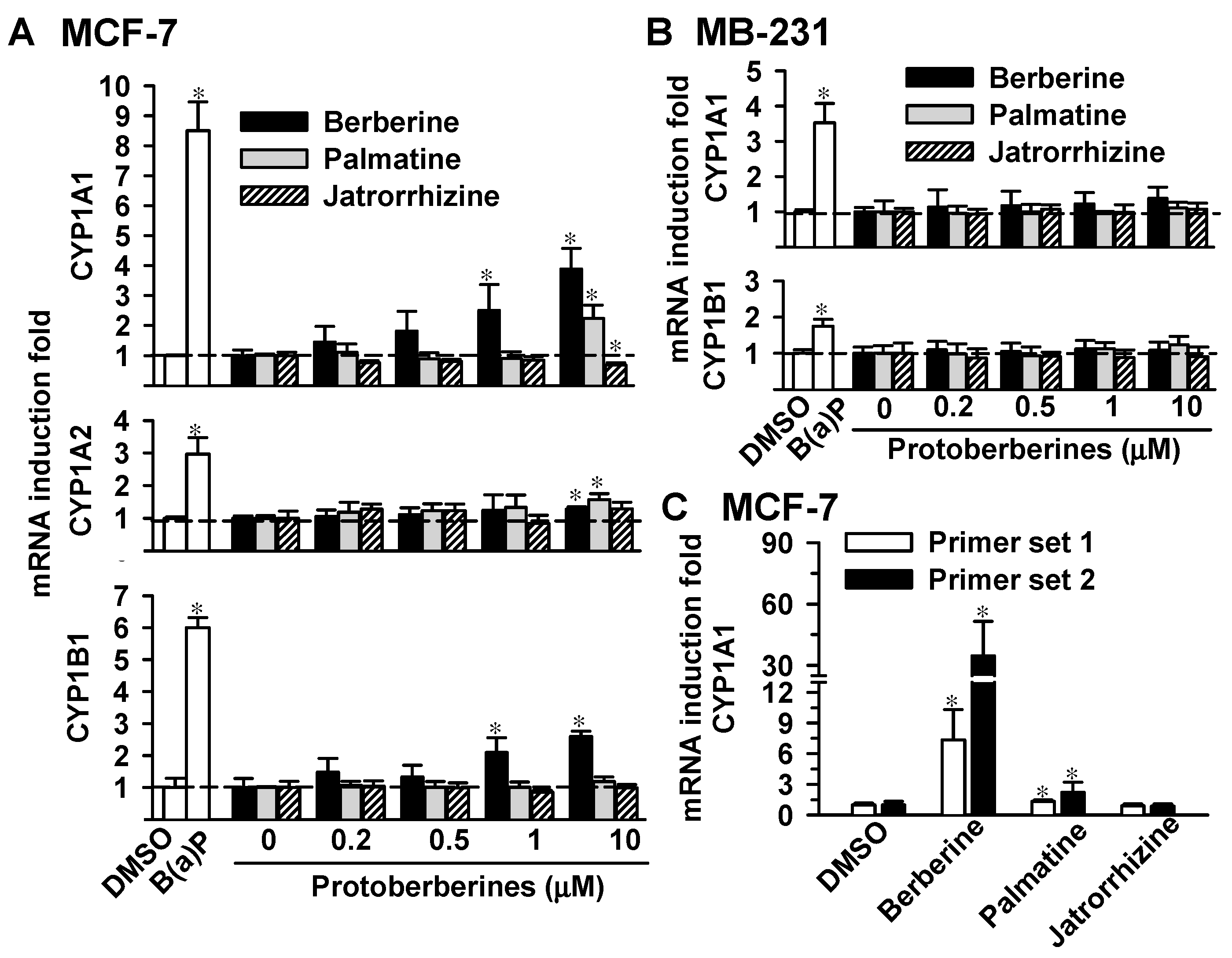
2.2. Effects of Berberine, Palmatine and Jatrorrhizine on mRNA Levels of CYP1s
2.3. Effects of Berberine on EROD Activity and the Protein Levels of CYP1A1 and CYP1B1
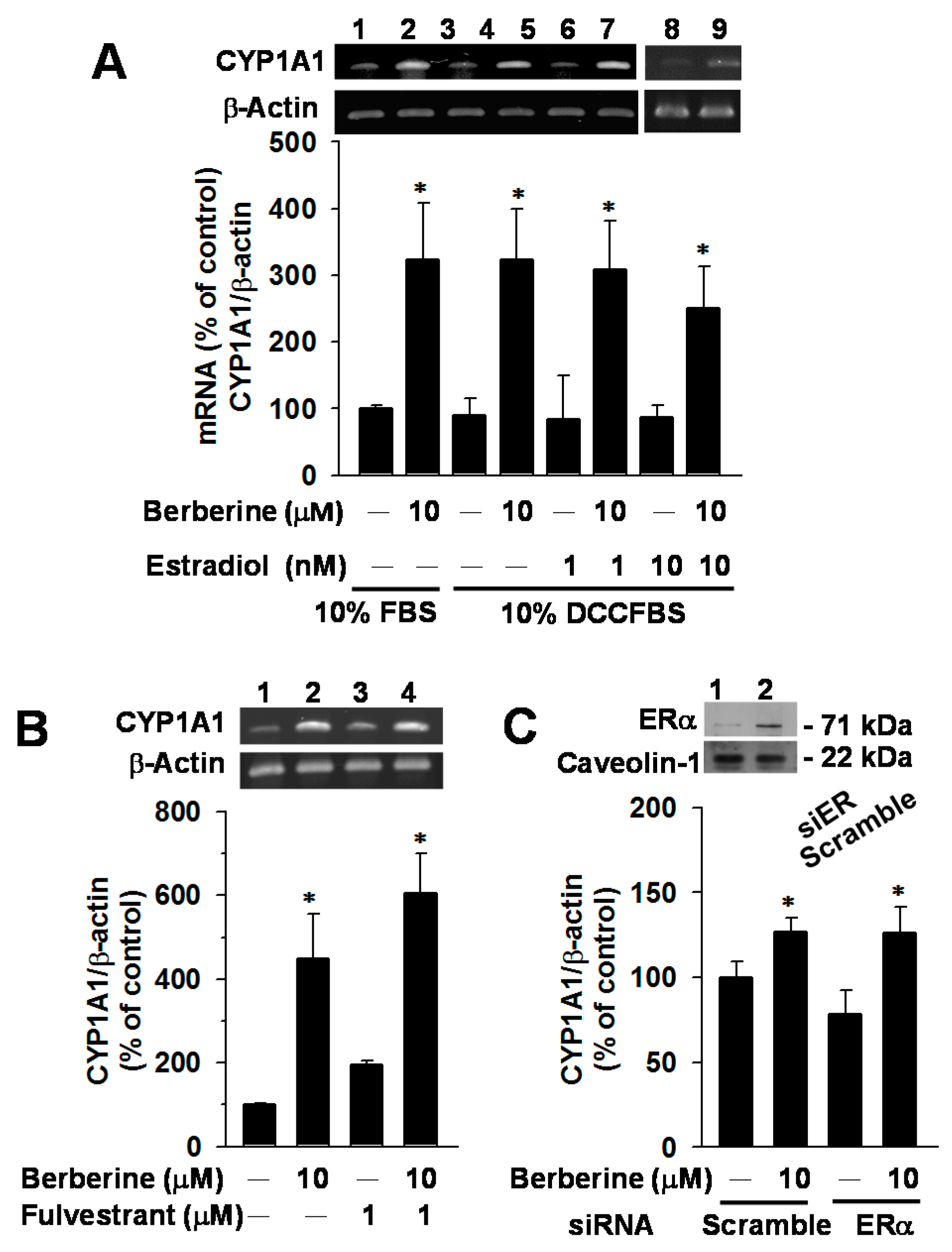
2.4. Effects of E2, Fulvestrant and ERα Knockdown on CYP1A1 mRNA Elevation by Berberine in MCF-7 Cells
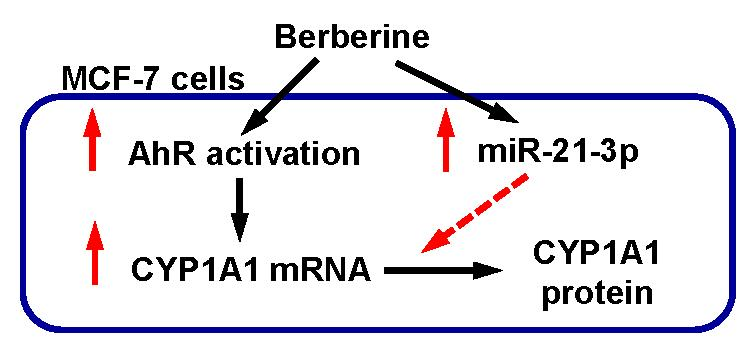
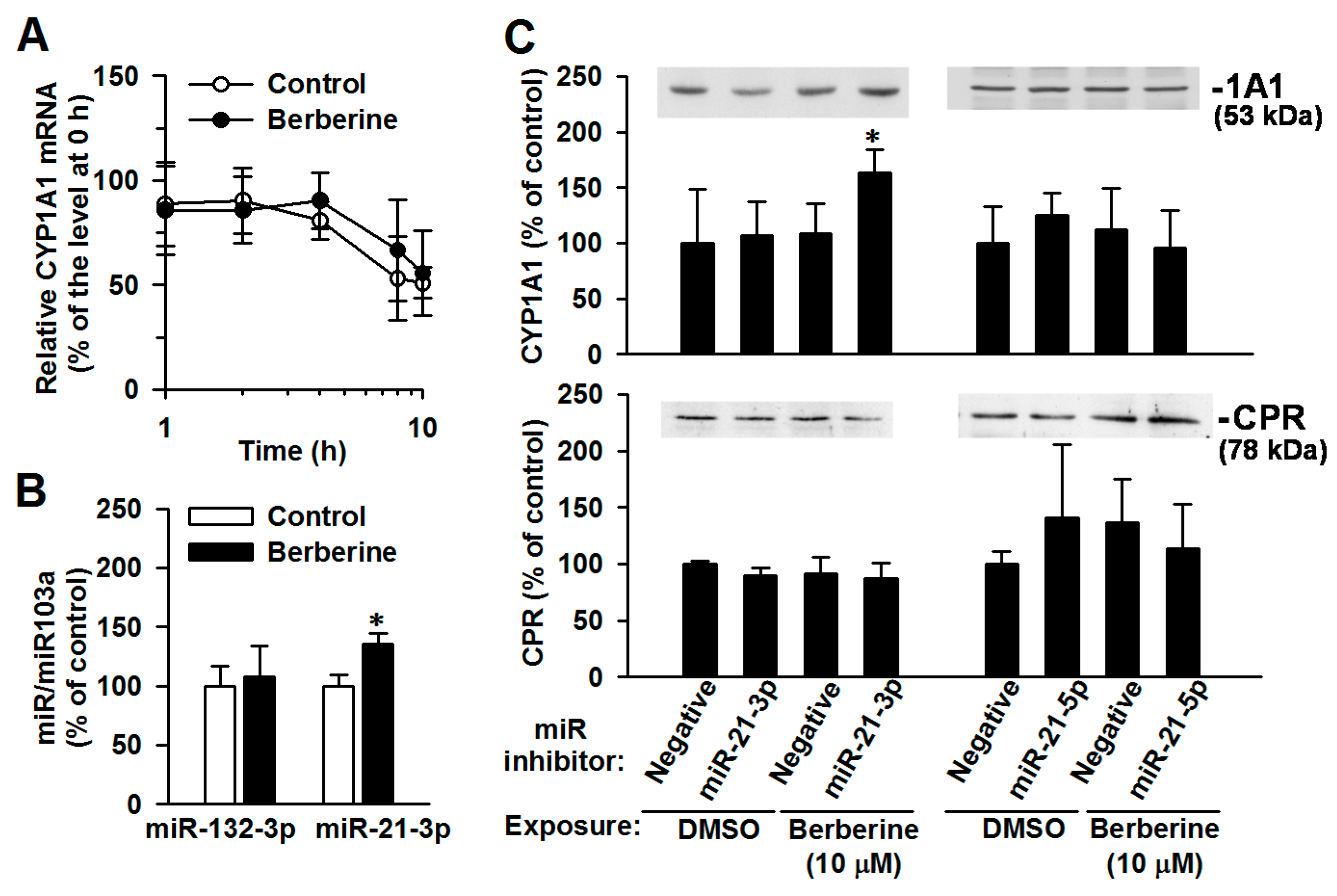
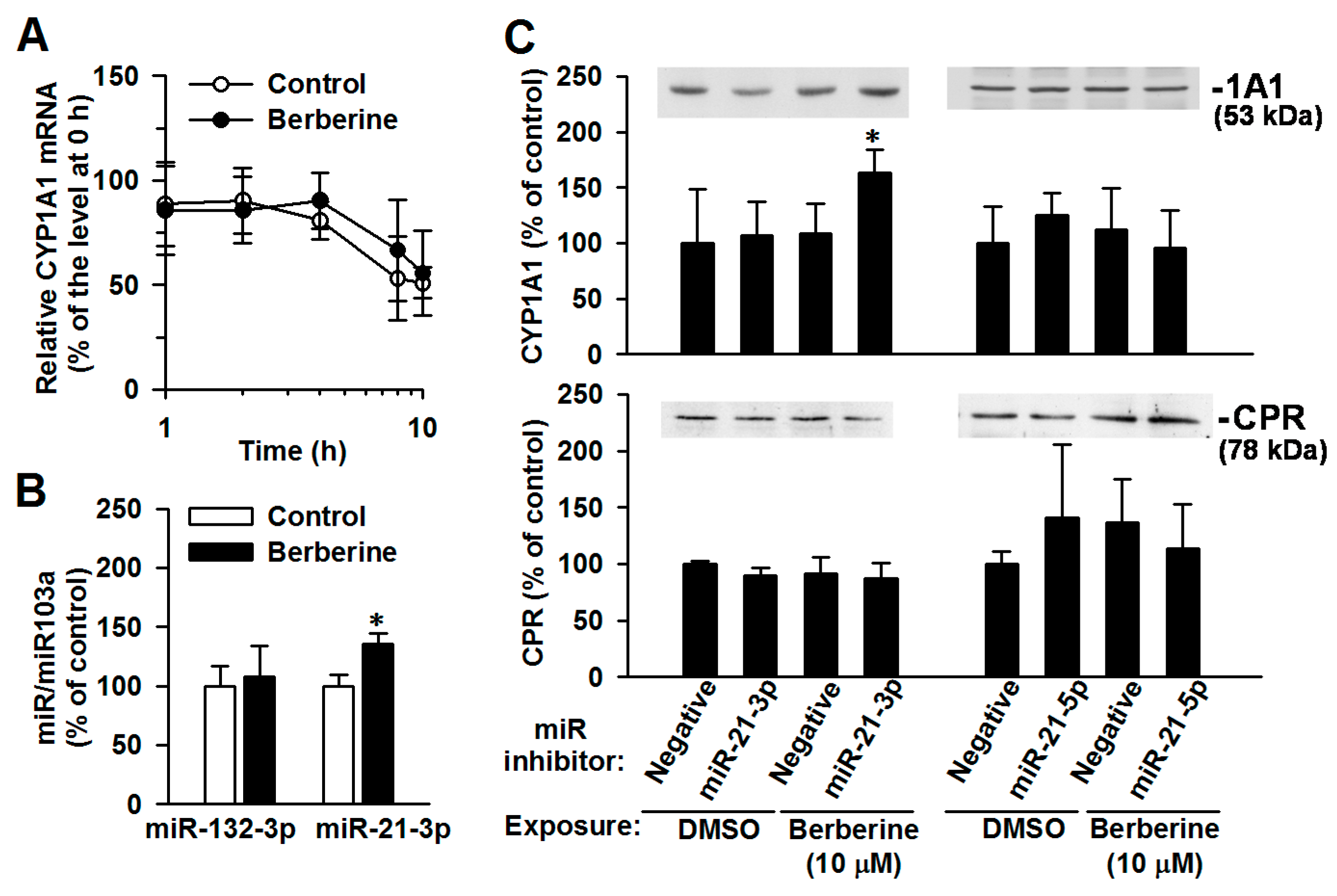
2.5. The Involvement of miR-21-3p in the Post-Transcriptional Regulation of CYP1A1 by Berberine
3. Discussion
4. Materials and Methods
4.1. Chemicals, Enzymes and Antibodies
4.2. Cell Culture and Exposure
4.3. DRE Reporter Assay
4.4. Immunoblot Analysis
4.5. Determination of Cellular EROD Activity
4.6. Stability and Reverse Transcription (RT)-Polymerase Chain Reaction (PCR) Analysis of mRNA
4.7. Transfection of Estrogen Receptor siRNA
4.8. Determination of miRs and Transfection of the miR Inhibitors
4.9. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Nebert, D.W.; Dalton, T.P. The role of cytochrome P450 enzymes in endogenous signaling pathways and environmental carcinogenesis. Nat. Rev. Cancer 2006, 6, 947–960. [Google Scholar] [CrossRef] [PubMed]
- Shimada, T.; Guengerich, F.P. Inhibition of human cytochrome P450 1A1-, 1A2-, and 1B1-mediated activation of procarcinogens to genotoxic metabolites by polycyclic aromatic hydrocarbons. Chem. Res. Toxicol. 2006, 19, 288–294. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Fasco, M.J.; Figge, H.L.; Keyomarsi, K.; Kaminsky, L.S. Expression of cytochromes P450 in human breast tissue and tumors. Drug Metab. Dispos. 1996, 24, 899–905. [Google Scholar] [PubMed]
- Bai, X.; Xie, J.; Sun, S.; Zhang, X.; Jiang, Y.; Pang, D. The associations of genetic polymorphisms in CYP1A2 and CYP3A4 with clinical outcomes of breast cancer patients in northern China. Oncotarget 2017, 8, 38367–38377. [Google Scholar] [CrossRef] [PubMed]
- Denison, M.S.; Nagy, S.R. Activation of the aryl hydrocarbon receptor by structurally diverse exogenous and endogenous chemicals. Annu. Rev. Pharmacol. Toxicol. 2003, 43, 309–334. [Google Scholar] [CrossRef] [PubMed]
- Döhr, O.; Vogel, C.; Abel, J. Different response of 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD)-sensitive genes in human breast cancer MCF-7 and MDA-MB 231 cells. Arch. Biochem. Biophys. 1995, 321, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Wilson, C.L.; Thomsen, J.; Hoivik, D.J.; Wormke, M.T.; Stanker, L.; Holtzapple, C.; Safe, S.H. Aryl hydrocarbon (Ah) nonresponsiveness in estrogen receptor-negative MDA-MB-231 cells is associated with expression of a variant arnt protein. Arch. Biochem. Biophys. 1997, 346, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Wihlen, B.; Ahmed, S.; Inzunza, J.; Matthews, J. Estrogen receptor subtype- and promoter-specific modulation of aryl hydrocarbon receptor-dependent transcription. Mol. Cancer Res. 2009, 7, 977–986. [Google Scholar] [CrossRef] [PubMed]
- Marques, M.; Laflamme, L.; Gaudreau, L. Estrogen receptor α can selectively repress dioxin receptor-mediated gene expression by targeting DNA methylation. Nucleic Acids Res. 2013, 41, 8094–8106. [Google Scholar] [CrossRef] [PubMed]
- Douglas, J.A.; Follett, J.M.; Parmenter, G.A.; Sansom, C.E.; Perry, N.B.; Littler, R.A. Seasonal variation of biomass and bioactive alkaloid content of goldenseal, Hydrastis canadensis. Fitoterapia 2010, 81, 925–928. [Google Scholar] [CrossRef] [PubMed]
- Jabbarzadeh Kaboli, P.; Rahmat, A.; Ismail, P.; Ling, K.H. Targets and mechanisms of berberine, a natural drug with potential to treat cancer with special focus on breast cancer. Eur. J. Pharmacol. 2014, 740, 584–595. [Google Scholar] [CrossRef] [PubMed]
- Lo, S.N.; Chang, Y.P.; Tsai, K.C.; Chang, C.Y.; Wu, T.S.; Ueng, Y.F. Inhibition of CYP1 by berberine, palmatine, and jatrorrhizine: Selectivity, kinetic characterization, and molecular modeling. Toxicol. Appl. Pharmacol. 2013, 272, 671–680. [Google Scholar] [CrossRef] [PubMed]
- Wincent, E.; Bengtsson, J.; Mohammadi Bardbori, A.; Alsberg, T.; Luecke, S.; Rannug, U.; Rannug, A. Inhibition of cytochrome P4501-dependent clearance of the endogenous agonist FICZ as a mechanism for activation of the aryl hydrocarbon receptor. Proc. Natl. Acad. Sci. USA 2012, 109, 4479–4484. [Google Scholar] [CrossRef] [PubMed]
- Vrzal, R.; Zdarilova, A.; Ulrichova, J.; Blaha, L.; Giesy, J.P.; Dvorak, Z. Activation of the aryl hydrocarbon receptor by berberine in HepG2 and H4IIE cells: Biphasic effect on CYP1A1. Biochem. Pharmacol. 2005, 70, 925–936. [Google Scholar] [CrossRef] [PubMed]
- Vrba, J.; Havlikova, M.; Gerhardova, D.; Ulrichova, J. Palmatine activates AhR and upregulates CYP1A activity in HepG2 cells but not in human hepatocytes. Toxicol. In Vitro 2014, 28, 693–699. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Pope, C.; Cheng, X.; Zhou, H.; Klaassen, C.D. Dose-response of berberine on hepatic cytochromes P450 mRNA expression and activities in mice. J. Ethnopharmacol. 2011, 138, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Rieger, J.K.; Klein, K.; Winter, S.; Zanger, U.M. Expression variability of absorption, distribution, metabolism, excretion-related microRNAs in human liver: Influence of nongenetic factors and association with gene expression. Drug Metab. Dispos. 2013, 41, 1752–1762. [Google Scholar] [CrossRef] [PubMed]
- Lo, T.F.; Tsai, W.C.; Chen, S.T. MicroRNA-21-3p, a berberine-induced miRNA, directly down-regulates human methionine adenosyltransferases 2A and 2B and inhibits hepatoma cell growth. PLoS ONE 2013, 8, e75628. [Google Scholar] [CrossRef] [PubMed]
- Yde, C.W.; Issinger, O.G. Enhancing cisplatin sensitivity in MCF-7 human breast cancer cells by down-regulation of Bcl-2 and cyclin D1. Int. J. Oncol. 2006, 29, 1397–1404. [Google Scholar] [CrossRef] [PubMed]
- Tomankova, K.; Polakova, K.; Pizova, K.; Binder, S.; Havrdova, M.; Kolarova, M.; Kriegova, E.; Zepletalova, J.; Malina, L.; Horakova, J.; et al. In vitro cytotoxicity analysis of doxorubicin-loaded/superparamagnetic iron oxide colloidal nanoassemblies on MCF7 and NIH3T3 cell lines. J. Nanomed. 2015, 10, 949–961. [Google Scholar] [CrossRef] [PubMed]
- Linnenbaum, M.; Busker, M.; Kraehling, J.R.; Behrends, S. Heme oxygenase isoforms differ in their subcellular trafficking during hypoxia and are differentially modulated by cytochrome P450 reductase. PLoS ONE 2012, 7, e35483. [Google Scholar] [CrossRef] [PubMed]
- Poljaková, J.; Eckschlager, T.; Činátl, J.; Kizek, R.; Frei, E.; Stiborová, M. Electrochemical determination of enzymes metabolizing ellipticine in thyroid cancer cells—A tool to explain the mechanism of ellipticine toxicity to these cells. Int. J. Electrochem. Sci. 2013, 8, 1573–1585. [Google Scholar]
- Xie, J.; Huang, X.; Park, M.S.; Pham, H.M.; Chan, W.K. Differential suppression of the aryl hydrocarbon receptor nuclear translocator-dependent function by an aryl hydrocarbon receptor PAS-A-derived inhibitory molecule. Biochem. Pharmacol. 2014, 88, 253–265. [Google Scholar] [CrossRef] [PubMed]
- Chanyshev, M.D.; Kosorotikov, N.I.; Titov, S.E.; Kolesnikov, N.N.; Gulyaeva, L.F. Expression of microRNAs, CYP1A1 and CYP2B1 in the livers and ovaries of female rats treated with DDT and PAHs. Life Sci. 2014, 103, 95–100. [Google Scholar] [CrossRef] [PubMed]
- Wen, C.J.; Wu, L.X.; Fu, L.J.; Shen, D.Y.; Zhang, X.; Zhang, Y.W.; Yu, J.; Zhou, H.H. Preferential induction of CYP1A1 over CYP1B1 in human breast cancer MCF-7 cells after exposure to berberine. Asian Pac. J. Cancer Prev. 2014, 15, 495–499. [Google Scholar] [CrossRef] [PubMed]
- Romay, M.C.; Che, N.; Becker, S.N.; Pouldar, D.; Hagopian, R.; Xiao, X.; Lusis, A.J.; Berliner, J.A.; Civelek, M. Regulation of NF-κB signaling by oxidized glycerophospholipid and IL-1β induced miRs-21–3p and -27a-5p in human aortic endothelial cells. J. Lipid Res. 2015, 56, 38–50. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.L.; Ma, Y.M.; Shi, R.; Wang, T.M.; Zhang, N.; Wang, C.H.; Yang, Y. Identification of the toxic constituents in Rhizoma Coptidis. J. Ethnopharmacol. 2010, 128, 357–364. [Google Scholar] [CrossRef] [PubMed]
- Hanieh, H.; Alzahrani, A. MicroRNA-132 suppresses autoimmune encephalomyelitis by inducing cholinergic anti-inflammation: A new Ahr-based exploration. Eur. J. Immunol. 2013, 43, 2771–2782. [Google Scholar] [PubMed]
- Wang, C.W.; Huang, C.C.; Chou, P.H.; Chang, Y.P.; Wei, S.; Guengerich, F.P.; Chou, Y.C.; Wang, S.F.; Lai, P.S.; Pavel, S.; et al. 7-Ketocholesterol and 27-hydroxycholesterol decreased doxorubicin sensitivity in breast cancer cells: Estrogenic activity and mTOR pathway. Oncotarget 2017, 8, 66033–66050. [Google Scholar] [CrossRef] [PubMed]
- Su, C.R.; Ueng, Y.F.; Dung, N.X.; Vijaya Bhaskar Reddy, M.; Wu, T.S. Cytochrome P3A4 inhibitors and other constituents of Fibraurea tinctoria. J. Nat. Prod. 2007, 70, 1930–1933. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.P.; Chang, L.W.; Yao, H.T.; Chang, H.; Tsai, H.T.; Tsai, M.H.; Yeh, T.K.; Lin, P. Involvement of oxidative stress and activation of aryl hydrocarbon receptor in elevation of CYP1A1 expression and activity in lung cells and tissues by arsenic: An in vitro and in vivo study. Toxicol. Sci. 2009, 107, 385–393. [Google Scholar] [CrossRef] [PubMed]
- Pan, W.C.; Chen, R.M.; Shen, Y.C.; Chen, C.C.; Ueng, Y.F. Suppressive effect of tobacco smoke extracts on oral P-glycoprotein function and its impact in smoke-induced insult to oral epidermal cells. Toxicol. Lett. 2009, 185, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Peters, A.K.; van Londen, K.; Bergman, A.; Bohonowych, J.; Denison, M.S.; van den Berg, M.; Sanderson, J.T. Effects of polybrominated diphenyl ethers on basal and TCDD-induced ethoxyresorufin activity and cytochrome P450-1A1 expression in MCF-7, HepG2, and H4IIE cells. Toxicol. Sci. 2004, 82, 488–496. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Antona, C.; Donato, M.T.; Pareja, E.; Gomez-Lechon, M.J.; Castell, J.V. Cytochrome P-450 mRNA expression in human liver and its relationship with enzyme activity. Arch. Biochem. Biophys. 2001, 393, 308–315. [Google Scholar] [CrossRef] [PubMed]
- Golijanin, D.; Tan, J.Y.; Kazior, A.; Cohen, E.G.; Russo, P.; Dalbagni, G.; Auborn, K.J.; Subbaramaiah, K.; Dannenberg, A.J. Cyclooxygenase-2 and microsomal prostaglandin E synthase-1 are overexpressed in squamous cell carcinoma of the penis. Clin. Cancer Res. 2004, 10, 1024–1031. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Not available. |


© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lo, S.-N.; Wang, C.-W.; Chen, Y.-S.; Huang, C.-C.; Wu, T.-S.; Li, L.-A.; Lee, I.-J.; Ueng, Y.-F. Berberine Activates Aryl Hydrocarbon Receptor but Suppresses CYP1A1 Induction through miR-21-3p Stimulation in MCF-7 Breast Cancer Cells. Molecules 2017, 22, 1847. https://doi.org/10.3390/molecules22111847
Lo S-N, Wang C-W, Chen Y-S, Huang C-C, Wu T-S, Li L-A, Lee I-J, Ueng Y-F. Berberine Activates Aryl Hydrocarbon Receptor but Suppresses CYP1A1 Induction through miR-21-3p Stimulation in MCF-7 Breast Cancer Cells. Molecules. 2017; 22(11):1847. https://doi.org/10.3390/molecules22111847
Chicago/Turabian StyleLo, Sheng-Nan, Chun-Wei Wang, Yueh-Shieh Chen, Chiung-Chiao Huang, Tian-Shung Wu, Lih-Ann Li, I-Jung Lee, and Yune-Fang Ueng. 2017. "Berberine Activates Aryl Hydrocarbon Receptor but Suppresses CYP1A1 Induction through miR-21-3p Stimulation in MCF-7 Breast Cancer Cells" Molecules 22, no. 11: 1847. https://doi.org/10.3390/molecules22111847
APA StyleLo, S.-N., Wang, C.-W., Chen, Y.-S., Huang, C.-C., Wu, T.-S., Li, L.-A., Lee, I.-J., & Ueng, Y.-F. (2017). Berberine Activates Aryl Hydrocarbon Receptor but Suppresses CYP1A1 Induction through miR-21-3p Stimulation in MCF-7 Breast Cancer Cells. Molecules, 22(11), 1847. https://doi.org/10.3390/molecules22111847