Inhibitory Effects of Total Triterpenoid Saponins Isolated from the Seeds of the Tea Plant (Camellia sinensis) on Human Ovarian Cancer Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
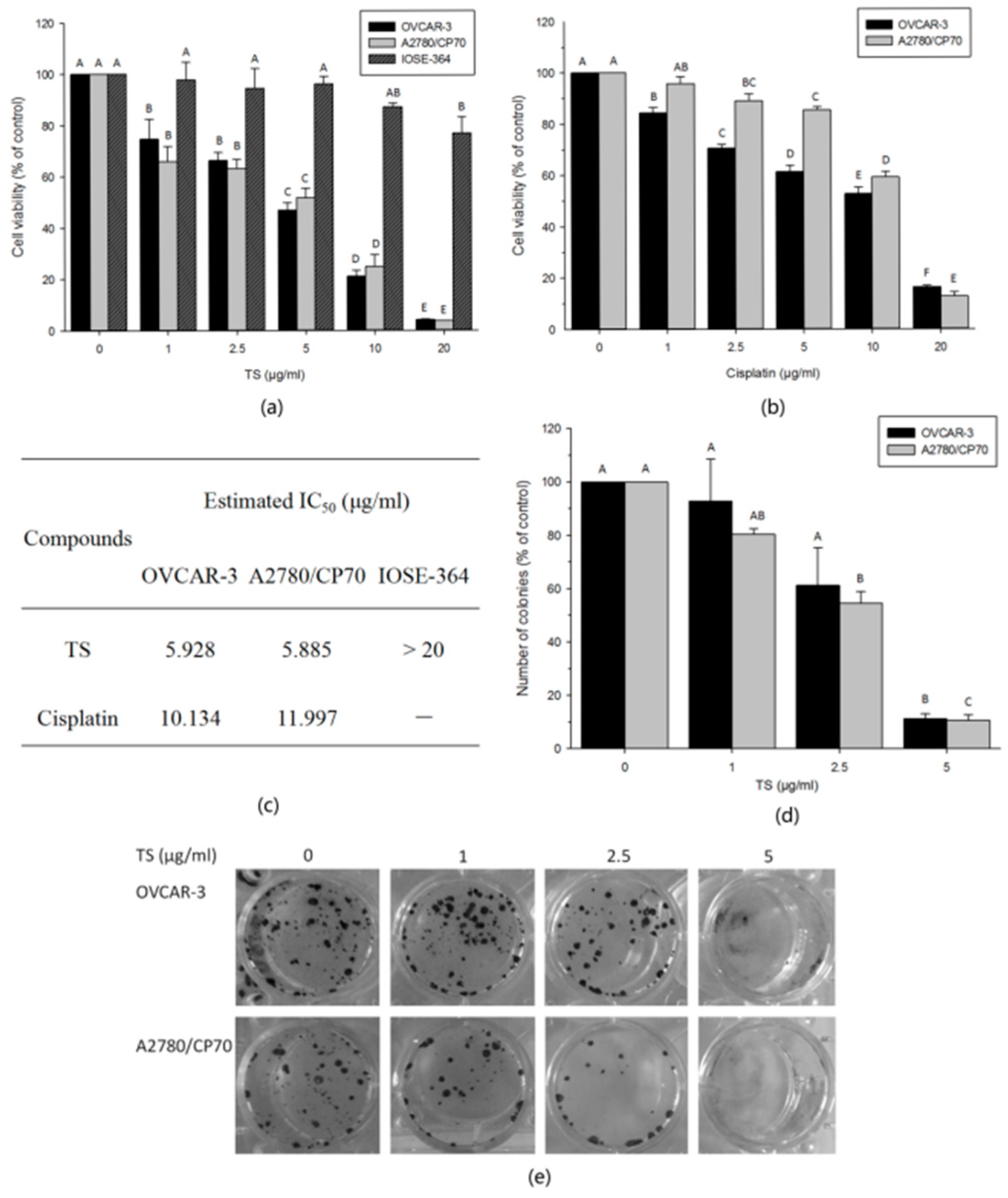
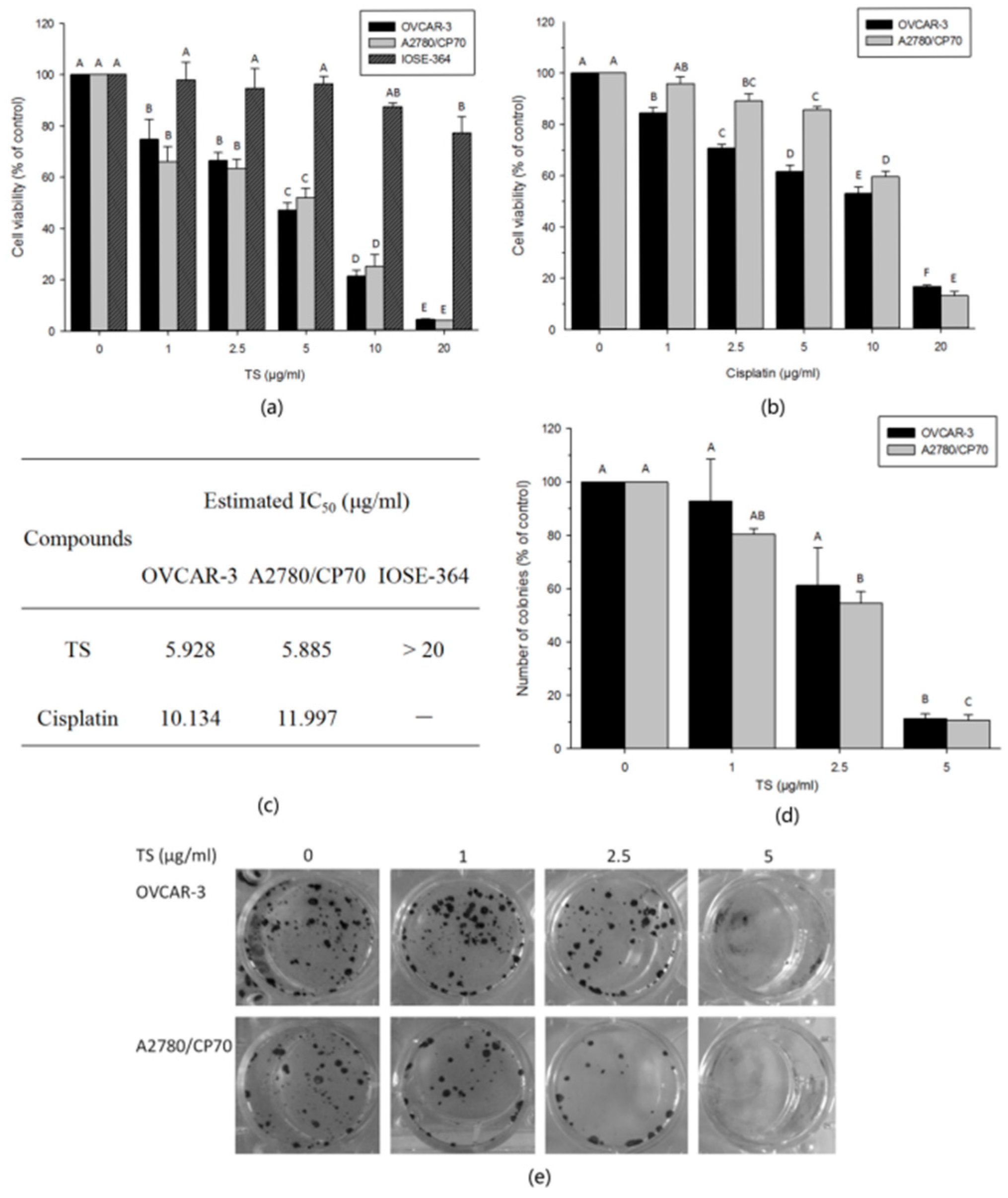
2.1. TS Inhibits Cell Growth and Colony Formation In Vitro
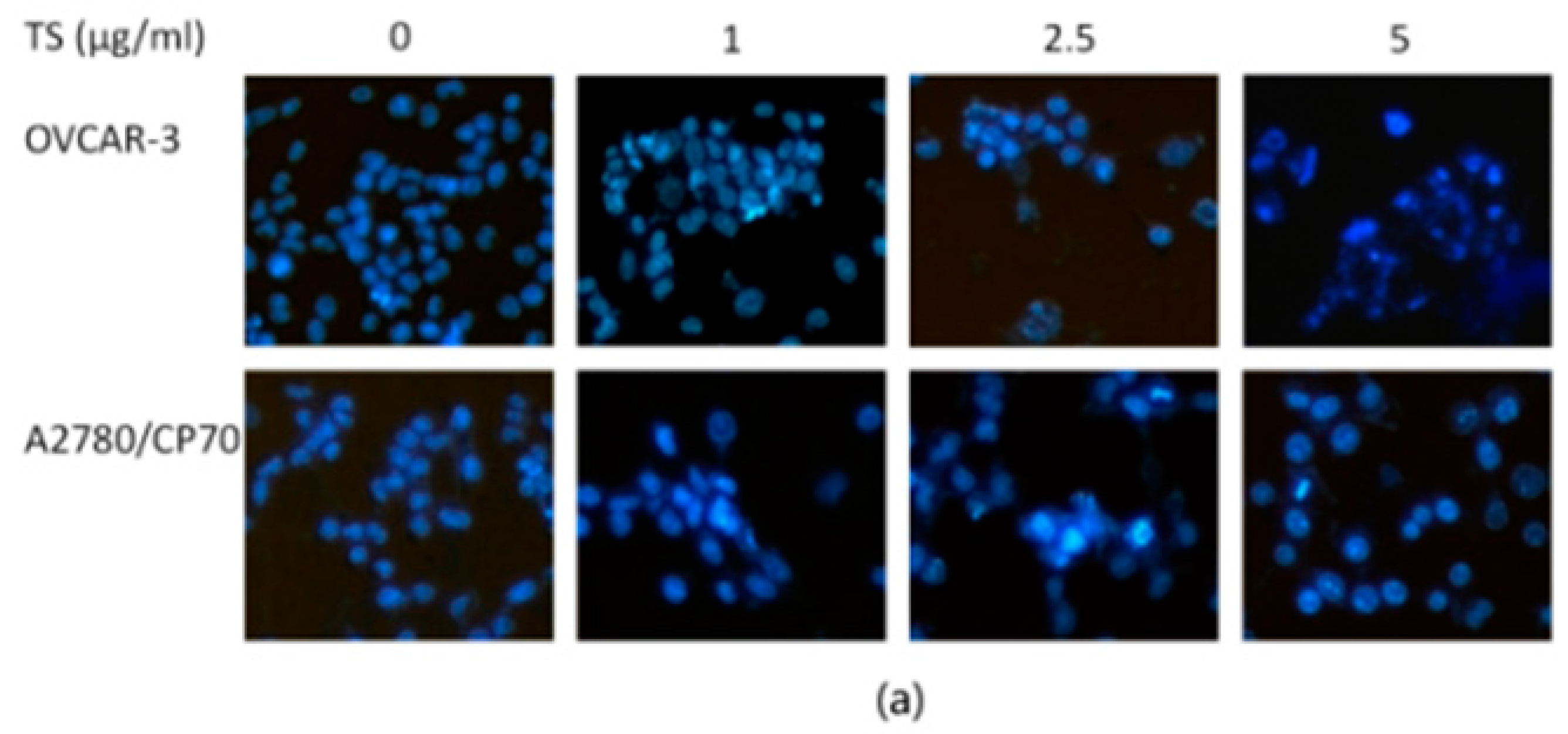
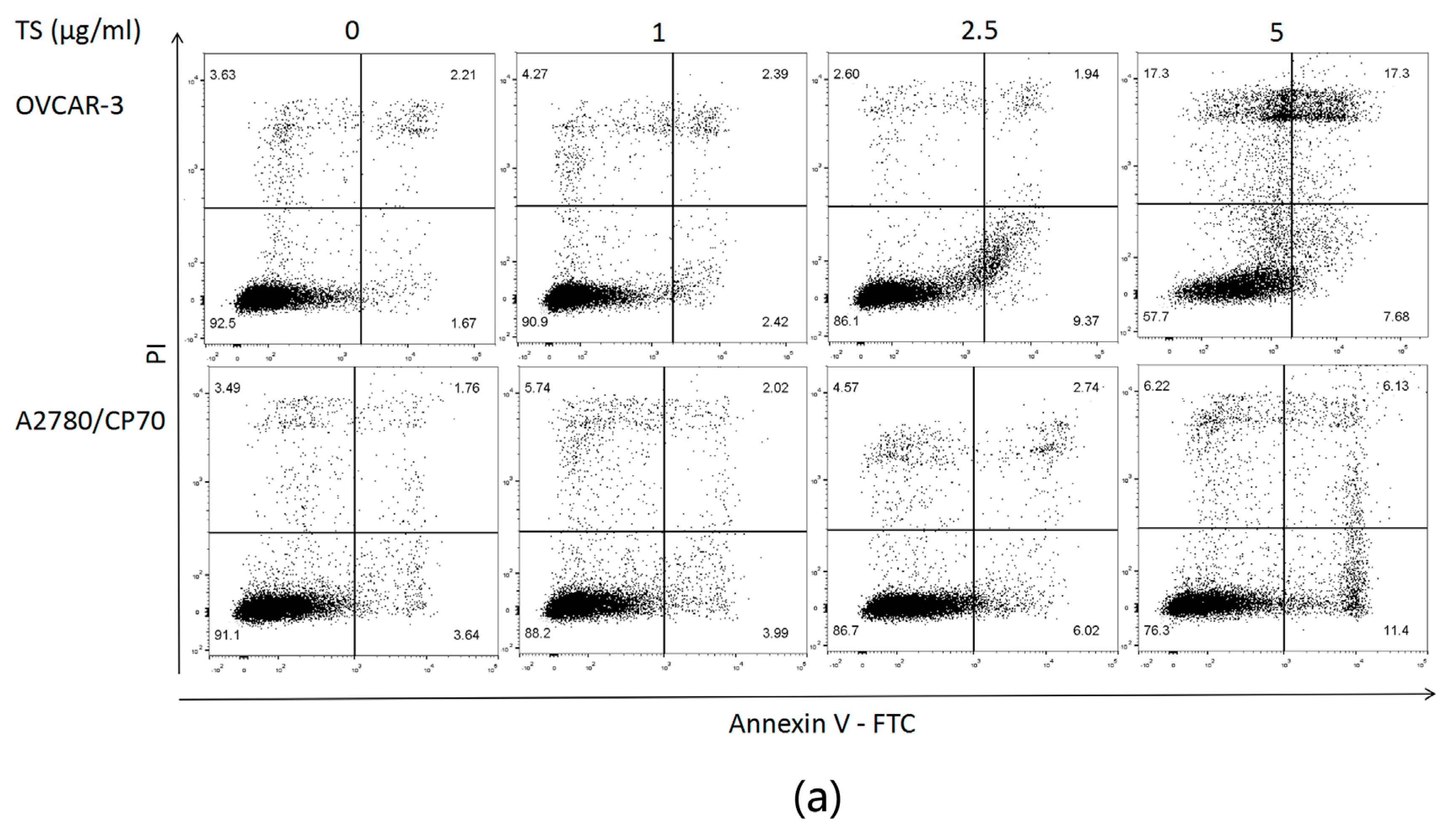
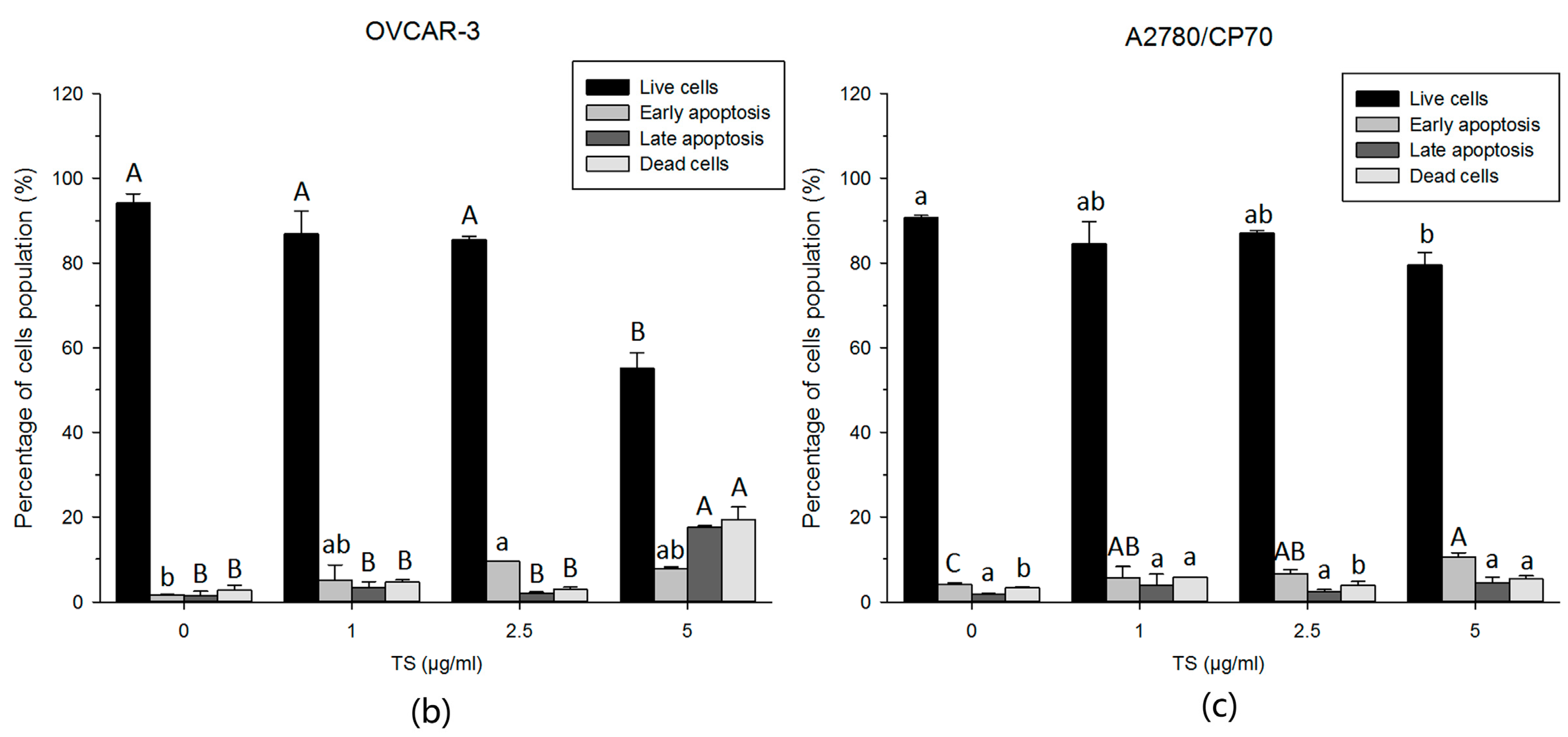
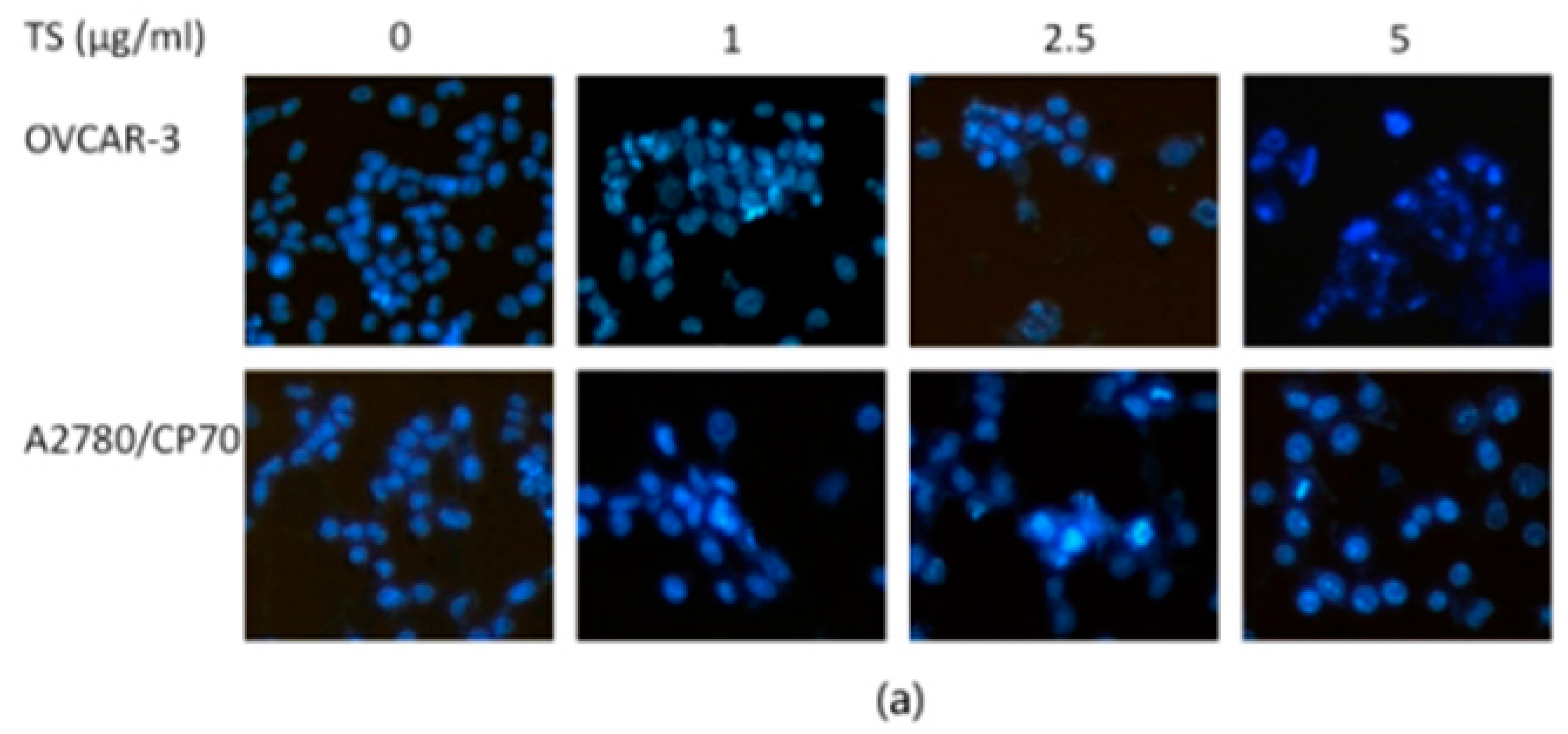
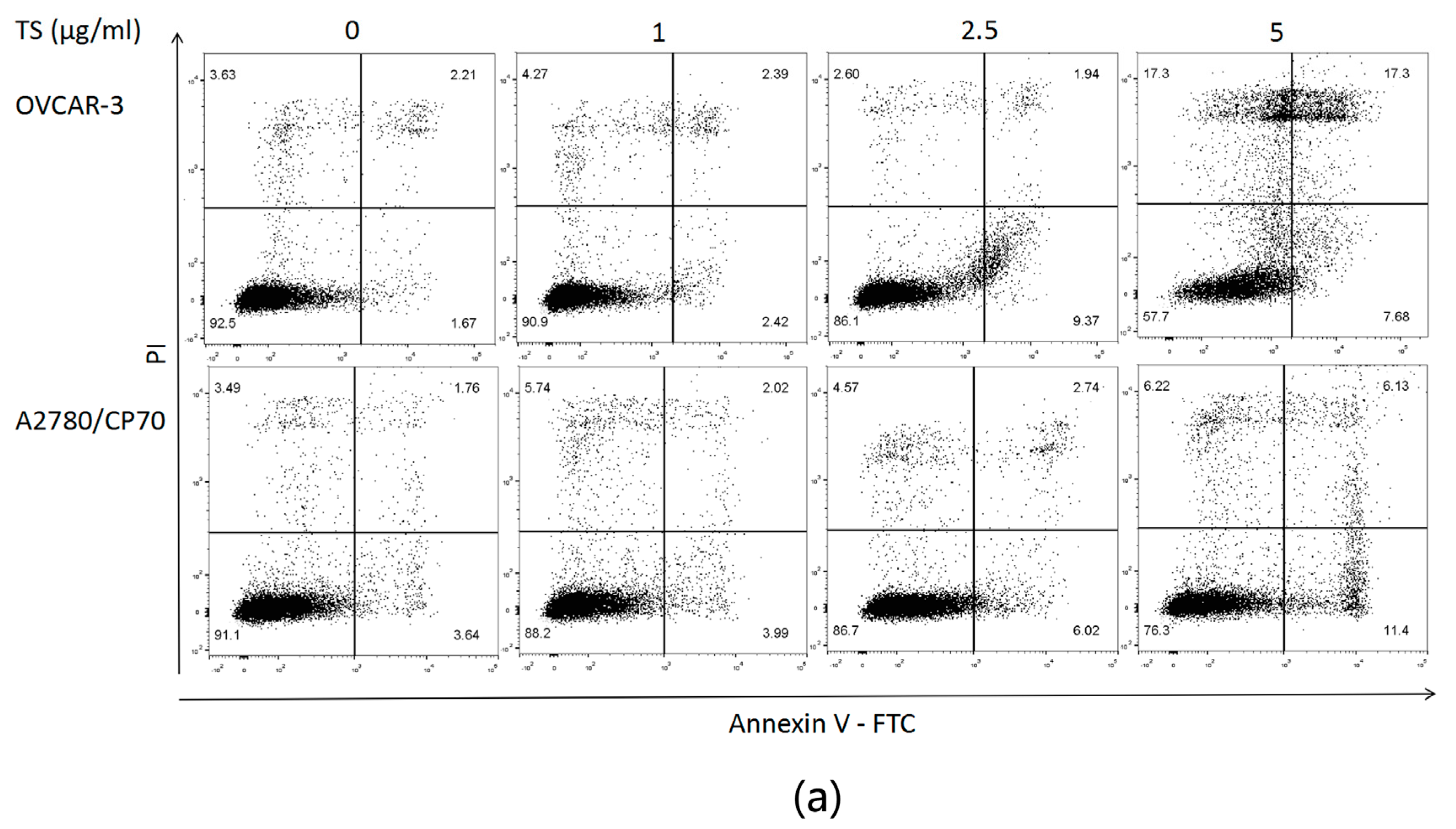
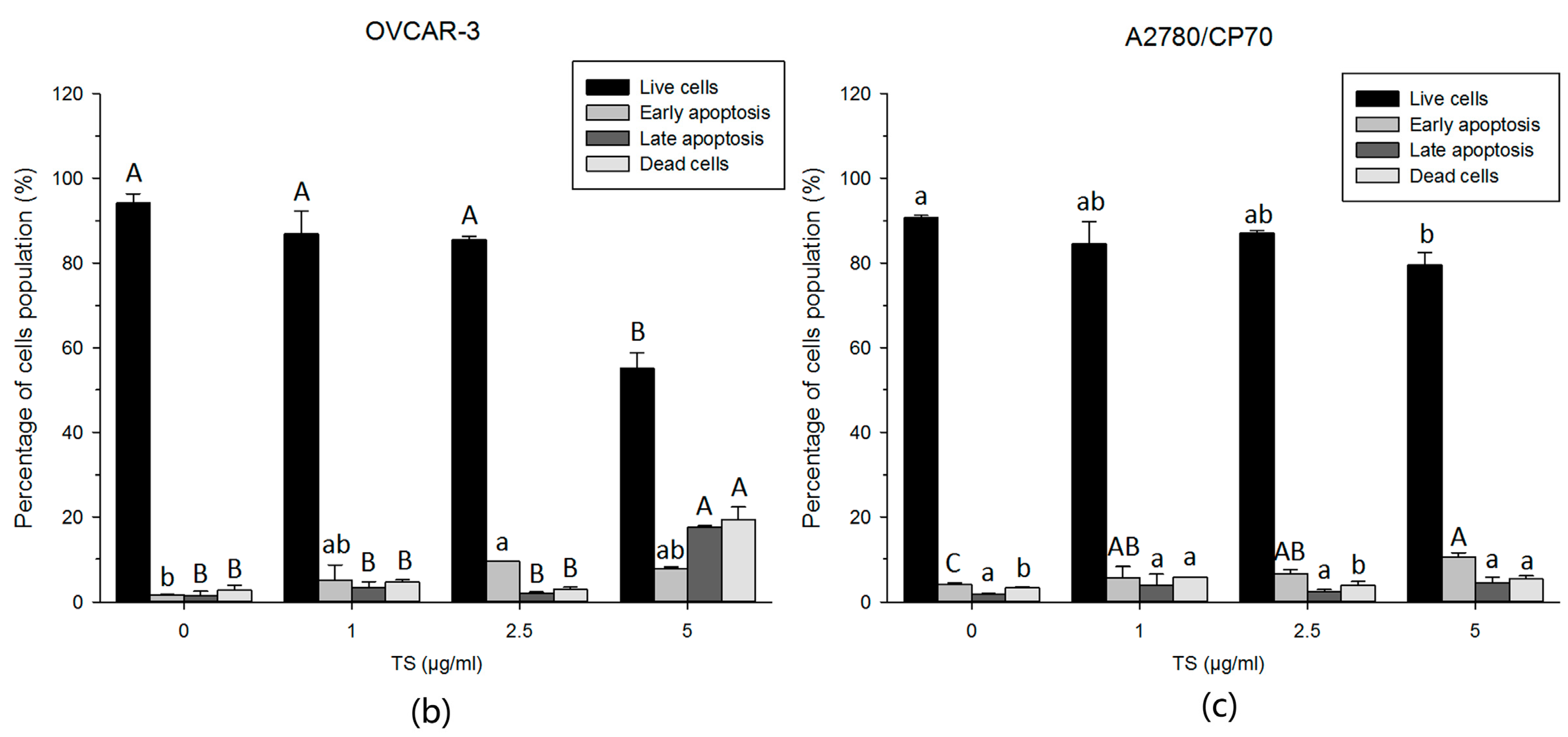
2.2. TS Induces Apoptosis in Both OVCAR-3 and A2780/CP70 Cells
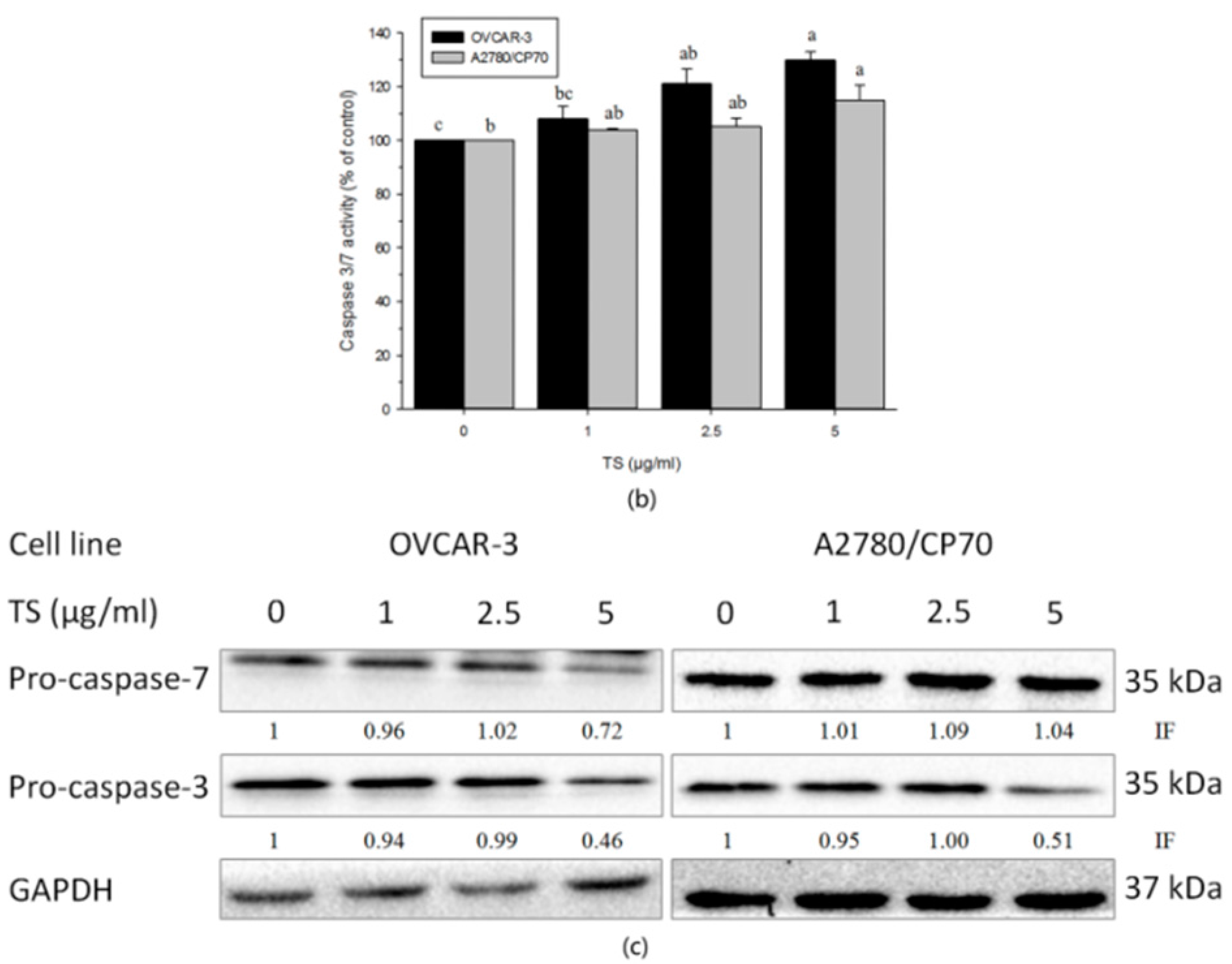
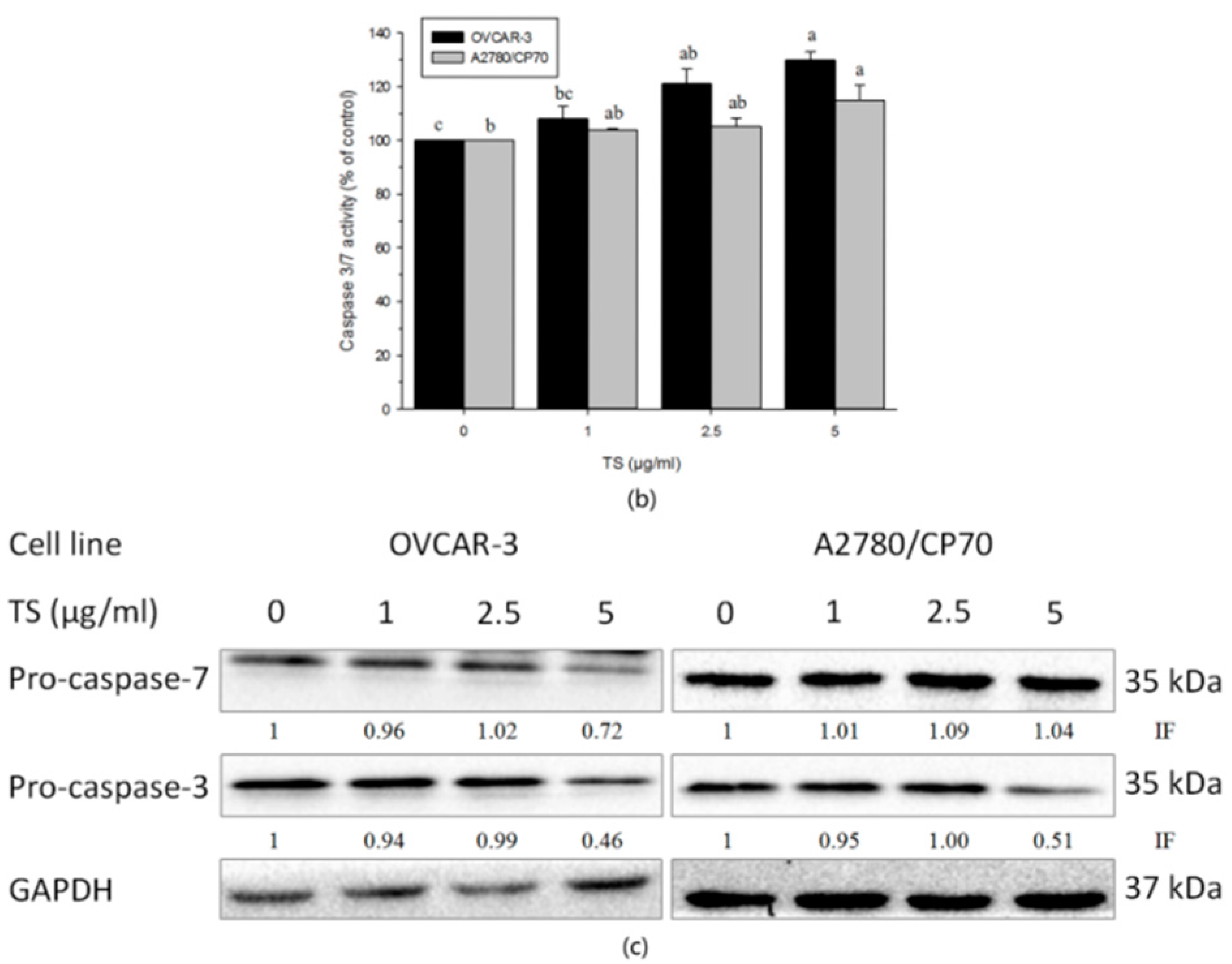
2.3. Effect of TS on the Intrinsic Apoptotic Pathway
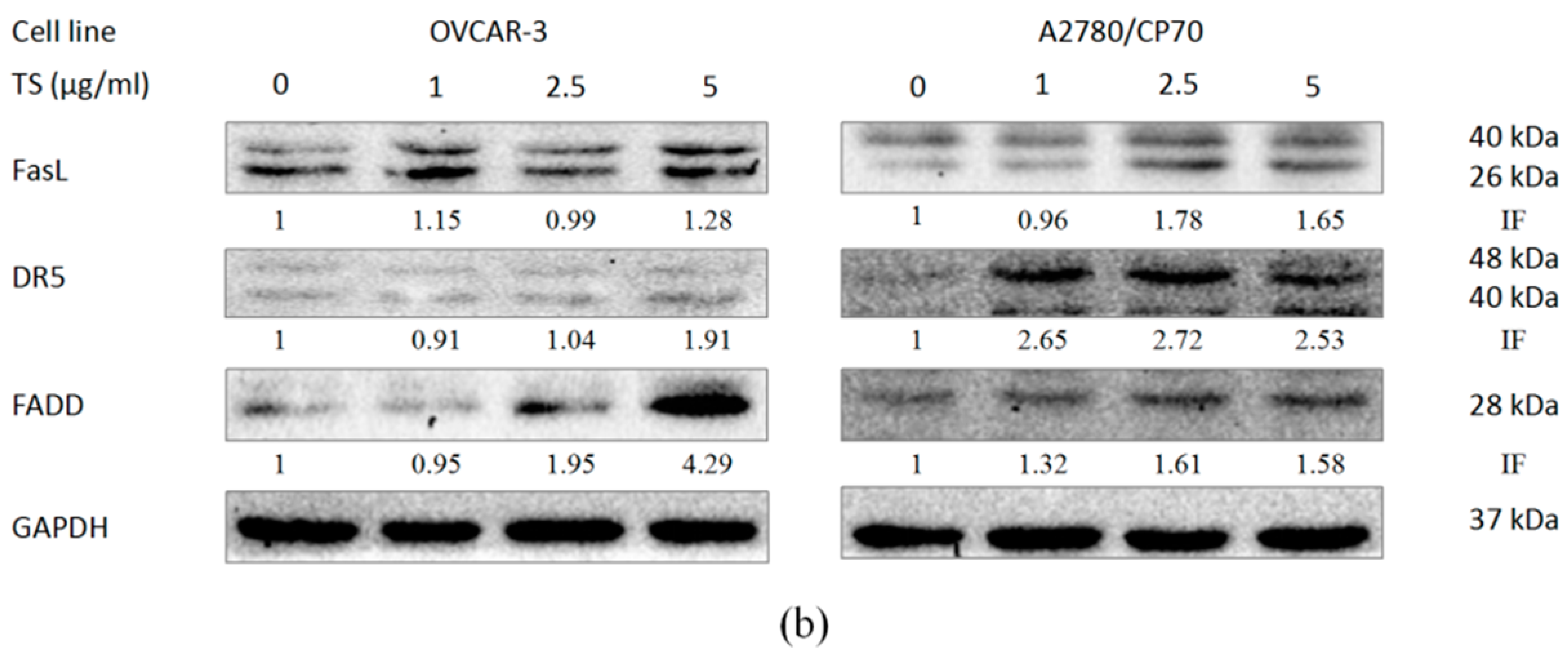
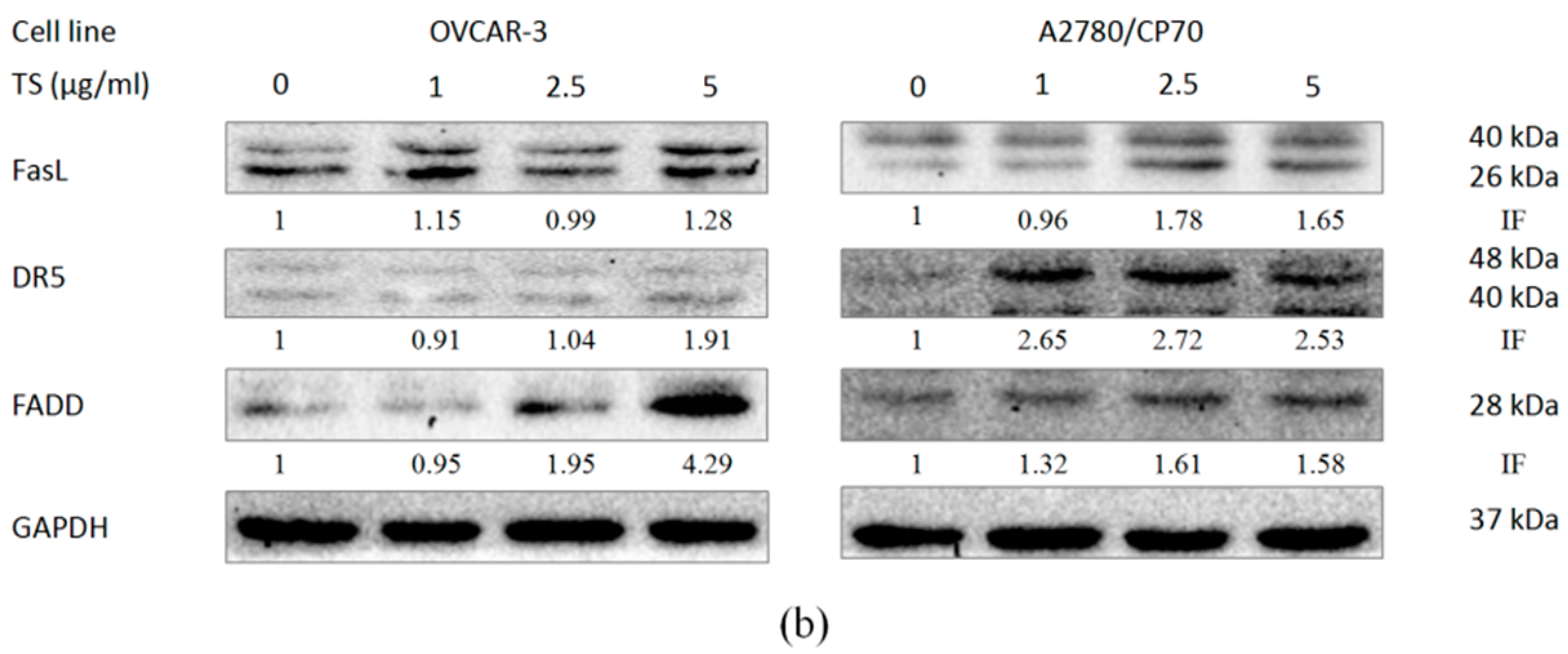
2.4. Effect of TS on the Extrinsic Apoptotic Pathway
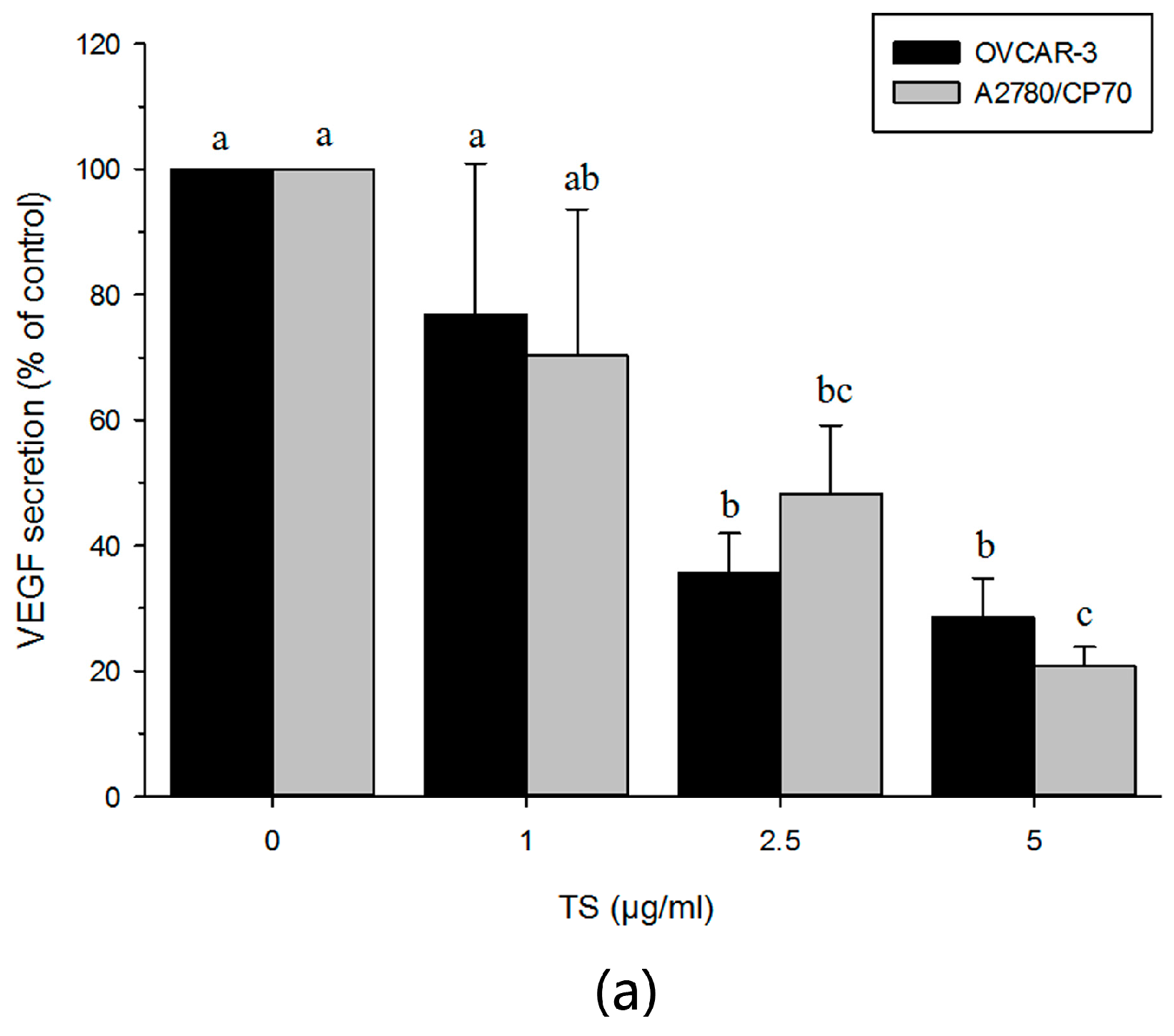
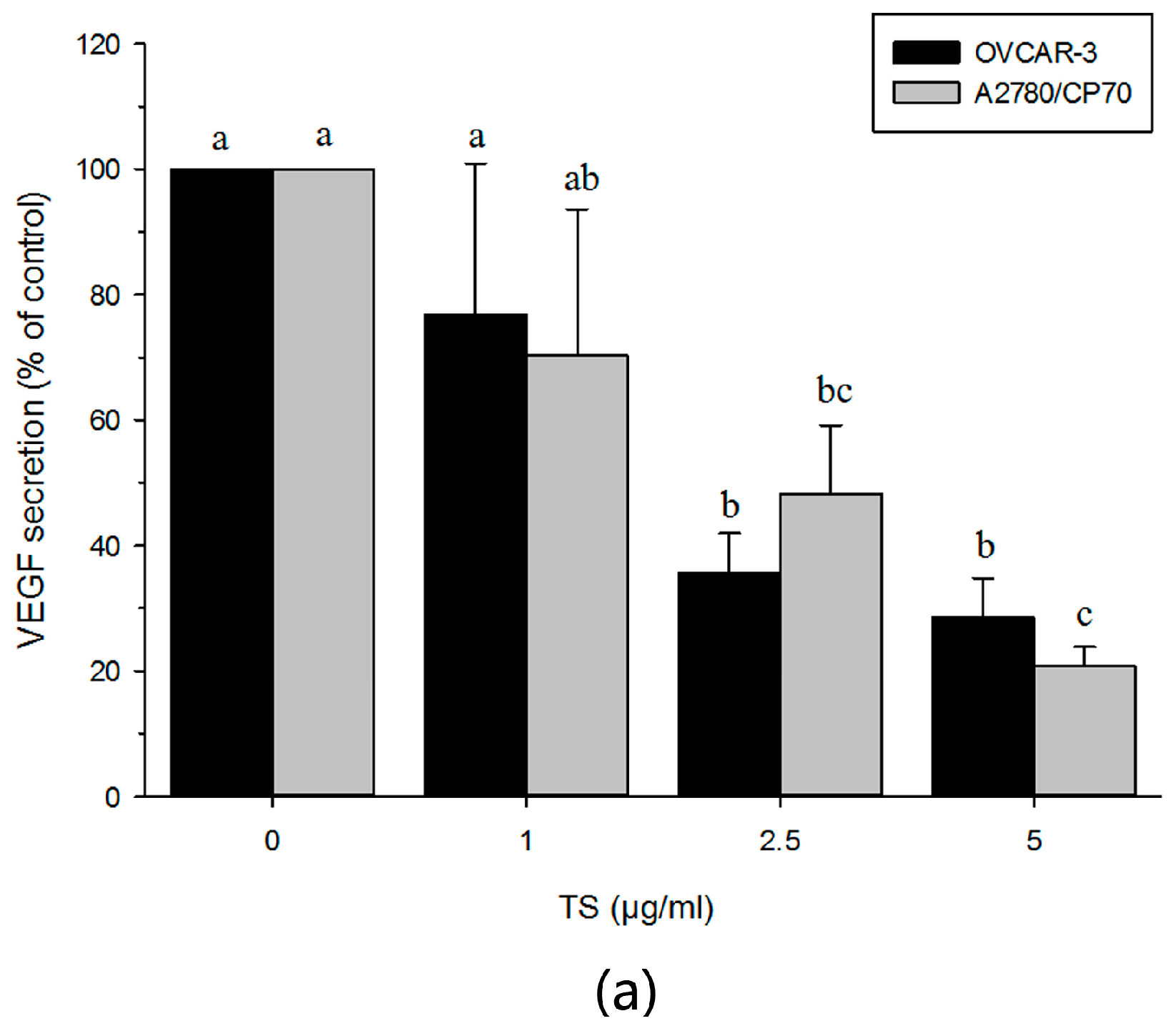
2.5. TS Reduces Tumor-Induced Angiogenesis In Vitro by Downregulating VEGF and HIF-1α
3. Discussion
4. Materials and Methods
4.1. General Experimental Procedures
4.2. Cell Culture
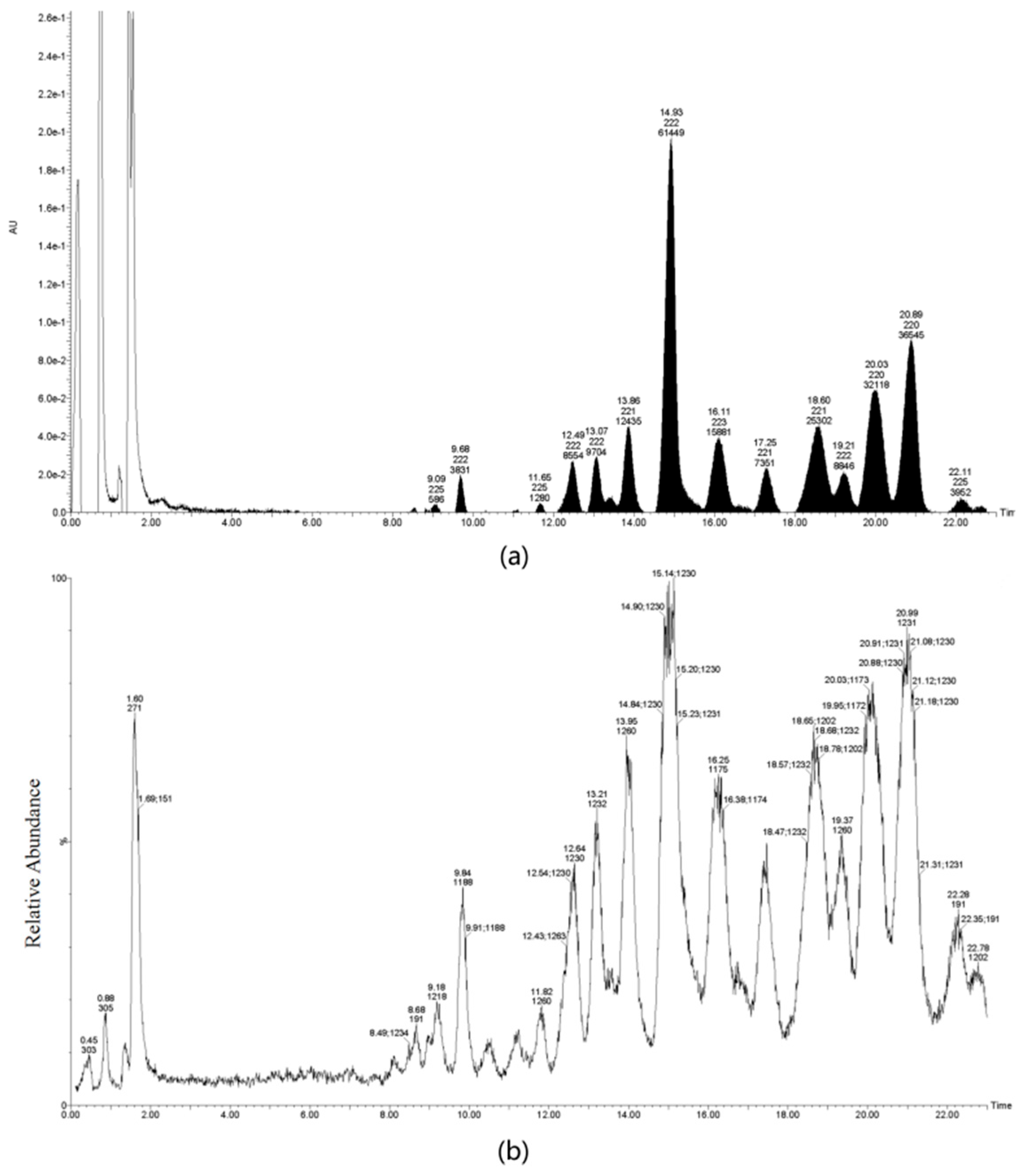
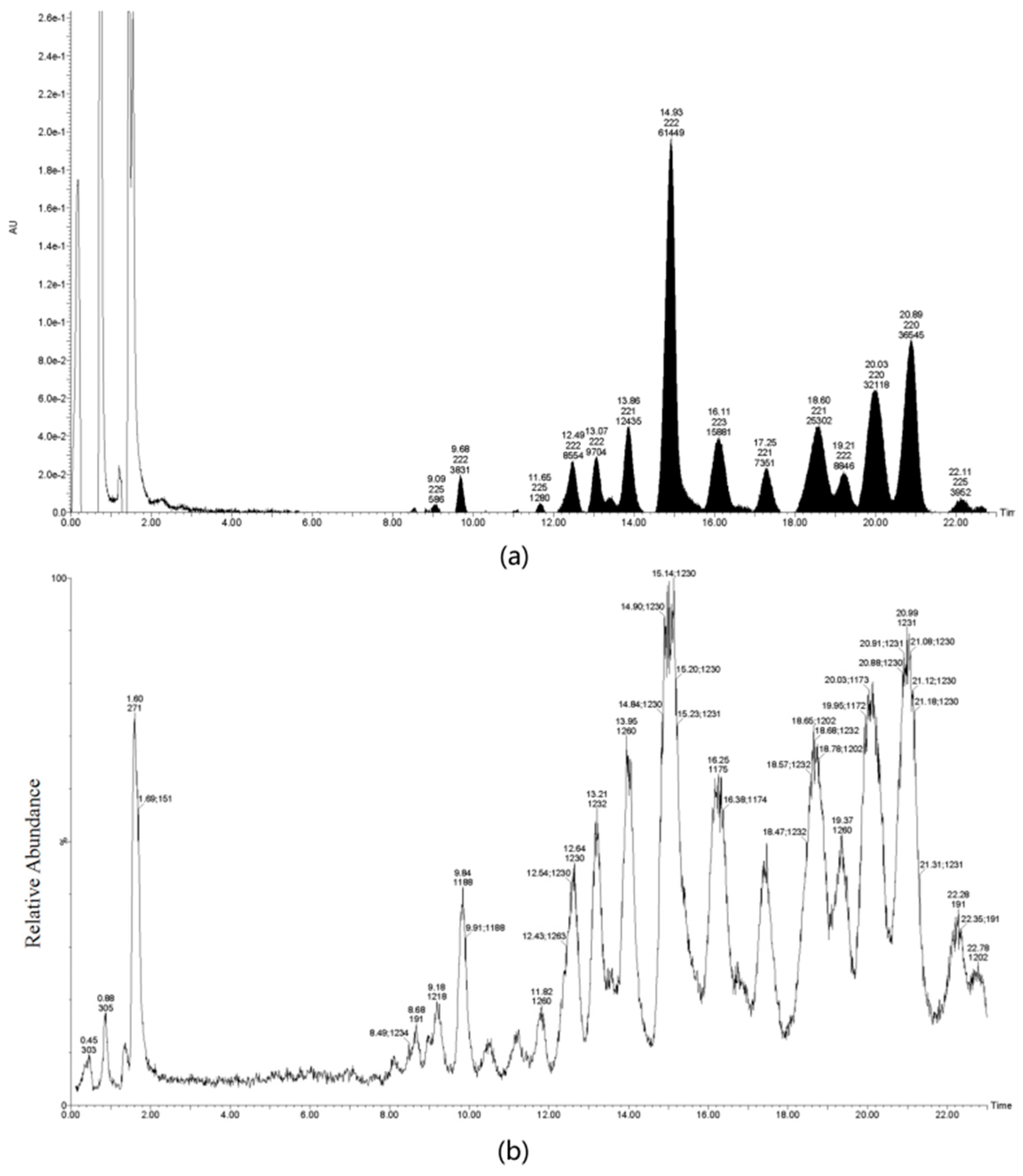
4.3. Extraction and Purification of TS
4.4. Cell Viability Assay
4.5. Colony Formation Assay
4.6. Apoptosis Assessment by Hoechst 33342 Staining
4.7. Caspase-3/7 Assay
4.8. Flow Cytometry Analysis of Cell Apoptosis
4.9. Enzyme-Linked Immunosorbent Assay
4.10. Western Blotting
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hanna, L.; Adams, M. Prevention of ovarian cancer. Best Pract. Res. Clin. Obstet. Gynaecol. 2006, 20, 339–362. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.R.; Lem, S. Ovarian cancer. Annu. Rev. Pathol. 2009, 4, 287–313. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2016. CA Cancer J. Clin. 2016, 66, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, L.A.; Huang, B.; Miller, R.W.; Tucker, T.; Goodrich, S.T.; Podzielinski, I.; DeSimone, C.P.; Ueland, F.R.; Nagell, J.R.; Seamon, L.G. Ten-year relative survival for epithelial ovarian cancer. Obstet. Gynecol. 2012, 120, 612–618. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.; Gore, M. The future of targeted therapies in ovarian cancer. Oncologist 2009, 14, 706–716. [Google Scholar] [CrossRef] [PubMed]
- Leitao, M.M., Jr.; Chi, D.S. Surgical management of recurrent ovarian cancer. Int. J. Gynecol. Cancer 2009, 36, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Jamieson, E.R.; Lippard, S.J. Structure, recognition, and processing of Cisplatin-DNA adducts. Chem. Rev. 1999, 99, 2467–2498. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Ruiz, S.; Maksimovic-Ivanic, D.; Mijatovic, S.; Kaluderovic, G.N. On the discovery, biological effects, and use of Cisplatin and metallocenes in anticancer chemotherapy. Bioinorg. Chem. Appl. 2012, 5, 140284. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, R.; Kaye, S.B. Ovarian cancer: Strategies for overcoming resistance to chemotherapy. Nat. Rev. Cancer 2003, 3, 502–516. [Google Scholar] [CrossRef] [PubMed]
- Shin, K.; Kim, K.H.; Yoon, M.S.; Suh, D.S.; Lee, J.Y.; Kim, A.; Eo, W. Expression of interactive genes associated with apoptosis and their prognostic value for ovarian serous adenocarcinoma. Adv. Clin. Exp. Med. 2016, 25, 513–521. [Google Scholar] [PubMed]
- Wong, R.S. Apoptosis in cancer: From pathogenesis to treatment. J. Exp. Clin. Cancer Res. 2011, 30, 87. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.I.; Kim, J.H.; Lee, K.T.; Choi, J.H. Costunolide induces apoptosis in platinum-resistant human ovarian cancer cells by generating reactive oxygen species. Gynecol. Oncol. 2011, 123, 588–596. [Google Scholar] [CrossRef] [PubMed]
- Wahl, H.; Tan, L.; Griffith, K.; Choi, M.; Liu, J.R. Curcumin enhances Apo2L/TRAIL-induced apoptosis in chemoresistant ovarian cancer cells. Gynecol. Oncol. 2007, 105, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Giri, S.; Karakoti, A.; Graham, R.P.; Maguire, J.L.; Reilly, C.M.; Seal, S.; Rattan, R.; Shridhar, V. Nanoceria: A rare-earth nanoparticle as a novel anti-angiogenic therapeutic agent in ovarian cancer. PLoS ONE 2013, 8, e54578. [Google Scholar]
- Folkman, J.; Watson, K.; Ingber, D.; Hanahan, D. Induction of angiogenesis during the transition from hyperplasia to neoplasia. Nature 1989, 339, 58–61. [Google Scholar] [CrossRef] [PubMed]
- Burger, R.A. Overview of anti-angiogenic agents in development for ovarian cancer. Gynecol. Oncol. 2011, 121, 230–238. [Google Scholar] [CrossRef] [PubMed]
- Nieves, B.J.; D’Amore, P.A.; Bryan, B.A. The function of vascular endothelial growth factor. Biofactors 2009, 35, 332–337. [Google Scholar] [CrossRef] [PubMed]
- Galanis, A.; Pappa, A.; Giannakakis, A.; Lanitis, E.; Dangaj, D.; Sandaltzopoulos, R. Reactive oxygen species and HIF-1 signalling in cancer. Cancer Lett. 2008, 266, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Keith, B.; Simon, C. Hypoxia-inducible factors, stem cells, and cancer. Cell 2007, 129, 465–472. [Google Scholar] [CrossRef] [PubMed]
- De Cuba, E.M.V.; de Hingh, I.H.J.T.; Sluiter, N.R.; Kwakman, R.; Coupé, V.M.H.; Beliën, J.A.M. Angiogenesis-related markers and prognosis after cytoreductive surgery and hyperthermic intraperitoneal chemotherapy for metastatic colorectal cancer. Ann. Surg. Oncol. 2006, 23, 1601–1608. [Google Scholar] [CrossRef] [PubMed]
- Zhong, H.; De Marzo, A.M.; Laughner, E.; Lim, M.; Hilton, D.A.; Zagzag, D.; Buechler, P.; Isaacs, W.B.; Semenza, G.l.; Simons, J.W. Overexpression of hypoxia-inducible factor 1alpha in common human cancers and their metastases. Cancer Res. 1999, 59, 5830–5835. [Google Scholar] [PubMed]
- Joshi, R.; Sood, S.; Dogra, P.; Mahendru, M.; Kumar, D.; Bhangalia, S.; Pal, H.C.; Kumar, N.; Bhushan, S.; Gulati, A.; et al. In vitro cytotoxicity, antimicrobial and metal-chelating activity of triterpene saponins from tea seed grown in Kangra valley, India. Med. Chem. Res. 2013, 22, 4030–4038. [Google Scholar] [CrossRef]
- Zhao, W.; Li, N.; Zhang, X.; Wang, W.; Li, J.; Si, Y. Cancer chemopreventive theasaponin derivatives from the total tea seed saponin of Camellia sinensis. J. Funct. Foods 2015, 12, 192–198. [Google Scholar] [CrossRef]
- Yang, W.S.; Ko, J.; Kim, E.; Kim, J.H.; Park, J.G.; Sung, N.Y. 21-o-angeloyltheasapogenol E3, a novel triterpenoid saponin from the seeds of tea plants, inhibits macrophage-mediated inflammatory responses in a NF-κB-dependent manner. Mediat. Inflamm. 2014, 4, 658351. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, K.; Sagesaka, Y.M.; Suzuki, T.; Suzuki, Y. Inactivation of human type A and B influenza viruses by tea-seed saponins. Biosci. Biotechnol. Biochem. 2000, 64, 184–186. [Google Scholar] [CrossRef] [PubMed]
- Akagi, M.; Fukuishi, N.; Kan, T.; Sagesaka, Y.M.; Akagi, R. Anti-allergic effect of tea-leaf saponin (TLS) from tea leaves (Camellia sinensis var. sinensis). Biol. Pharm. Bull. 1997, 20, 565–567. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, H.; Hamao, M.; Nakamura, S.; Kon’i, H.; Murata, M.; Yoshikawa, M. Medicinal flowers. XXXIII. anti-hyperlipidemic and anti-hyperglycemic effects of chakasaponins I-III and structure of chakasaponin IV from flower buds of Chinese tea plant (Camellia sinensis). Chem. Phar. Bull. 2012, 60, 674–680. [Google Scholar] [CrossRef]
- Chattopadhyay, P.; Besra, S.E.; Gomes, A.; Das, M.; Sur, P.; Mitra, S.; Vedasiromoni, J.R. Anti-inflammatory activity of tea (Camellia sinensis) root extract. Life Sci. 2004, 74, 1839–1849. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, M.; Morikawa, T.; Nakamura, S.; Li, X.; Matsuda, H. Bioactive saponins and glycosides. XXV. acylated oleanane-type triterpene saponins from the seeds of tea plant (Camellia sinensis). Chem. Phar. Bull. 2007, 55, 57–63. [Google Scholar] [CrossRef]
- Li, N.; Ma, Z.J.; Chu, Y.; Wang, Y.; Li, X. Phytochemical analysis of the triterpenoids with cytotoxicity and QR inducing properties from the total tea seed saponin of Camellia sinensis. Fitoterapia 2013, 84, 321–325. [Google Scholar] [CrossRef] [PubMed]
- Kitagawa, I.; Hori, K.; Motozawa, T.; Murakami, Y. Structures of new acylated oleanene-type triterpene oligoglycosides, theasaponins E1 and E2, from the seeds of tea plant, Camellia sinensis (L.) O. KUNTZE. Chem. Pharm. Bull. 1998, 46, 1901–1906. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, M.; Morikawa, T.; Li, N.; Nagatomo, A.; Li, X.; Matsuda, H. Bioactive saponins and glycosides. XXIII. triterpene saponins with gastroprotective effect from the seeds of Camellia sinensis—Theasaponins E3, E4, E5, E6, and E7. Chem. Pharm. Bull. 2005, 53, 1559–1564. [Google Scholar] [CrossRef] [PubMed]
- Morikawa, T.; Li, N.; Nagatomo, A.; Matsuda, H.; Li, X. Triterpene saponins with gastroprotective effects from tea seed (the seeds of Camellia sinensis). J. Nat. Prod. 2006, 69, 185. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Feng, K.; Yang, X. In vitro antifungal activity and mechanism of action of tea polyphenols and tea saponin against Rhizopus stolonifer. J. Mol. Microbiol. Biotechnol. 2005, 25, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.D.; Chaudhary, N.; Seo, H.J.; Kim, M.Y.; Shin, T.S. Theasaponin E1 as an effective ingredient for anti-angiogenesis and anti-obesity effects. Biosci. Biotechnol. Biochem. 2014, 78, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Bhardwaj, J.; Chaudhary, N.; Seo, H.J.; Kim, M.Y.; Shin, T.S.; Kim, J.D. Immunomodulatory effect of tea saponin in immune T-cells and T-lymphoma cells via regulation of Th1, Th2 immune response and MAPK/ERK2 signaling pathway. Immunopharmacol. Immunotoxicol. 2014, 36, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Rankin, G.O.; Tu, Y.Y.; Chen, Y.C. Theaflavin-3,3′-digallate decreases human ovarian carcinoma OVCAR-3 cell-induced angiogenesis via Akt and Notch-1 pathways, not via MAPK pathways. Int. J. Oncol. 2016, 48, 281–292. [Google Scholar] [CrossRef] [PubMed]
- Kitagawa, N.; Morikawa, T.; Motai, C.; Ninomiya, K.; Okugawa, S.; Nishida, A.; Yoshikawa, M.; Muraoka, O. The antiproliferative effect of chakasaponins I and II, floratheasaponin A, and epigallocatechin3-o-gallate isolated from Camellia sinensis on human digestive tract carcinoma cell lines. Int. J. Mol. Sci. 2016, 17, 1979. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Jiang, P.; Wang, P.; Yang, C.S.; Wang, X.; Feng, Q. EGCG enhances cisplatin sensitivity by regulating expression of the copper and cisplatin influx transporter CTR1 in ovary cancer. PLoS ONE 2015, 10, e0125402. [Google Scholar]
- Harpole, J.L.; Tucci, M.; Benghuzzi, H. Pathophysiological effects of thymoquinone and epigallocatechin-3-gallate on SKOV-3 ovarian cancer like cell line. Biomed. Sci. Instrum. 2015, 51, 31–39. [Google Scholar] [PubMed]
- Wang, F.; Chang, Z.; Fan, Q.; Wang, L. Epigallocatechin-3-gallate inhibits the proliferation and migration of human ovarian carcinoma cells by modulating p38 kinase and matrix metalloproteinase-2. Mol. Med. Rep. 2014, 9, 1085–1089. [Google Scholar] [CrossRef] [PubMed]
- Tu, Y.Y.; Kim, E.; Gao, Y.; Rankin, G.O.; Li, B.; Chen, Y.C. Theaflavin-3,3′-digallate induces apoptosis and G2 cell cycle arrest through the Akt/MDM2/p53 pathway in cisplatin-resistant ovarian cancer A2780/CP70 cells. Int. J. Oncol. 2016, 48, 2657–2665. [Google Scholar] [PubMed]
- Brunelle, J.K.; Letai, A. Control of mitochondrial apoptosis by the Bcl-2 family. J. Cell Sci. 2009, 122, 437–441. [Google Scholar] [CrossRef] [PubMed]
- Winterhoff, B.; Freyer, L.; Hammond, E.; Giri, S.; Mondal, S.; Roy, D.; Teoman, A.; Mullany, S.A.; Hoffmann, R.; von Bismarck, A.; et al. PG545 enhances anti-cancer activity of chemotherapy in ovarian models and increases surrogate biomarkers such as VEGF in preclinical and clinical plasma samples. Eur. J. Cancer 2015, 51, 879–892. [Google Scholar] [CrossRef] [PubMed]
- Kinghorn, A.D.; Chin, Y.W.; Swanson, S.M. Discovery of natural product anticancer agents from biodiverse organisms. Curr. Opin. Drug Discov. Dev. 2009, 12, 189–196. [Google Scholar]
- Li, B.; Chen, D.; Li, W.; Xiao, D. 20(S)-Protopanaxadiol saponins inhibit SKOV3 cell migration. Oncol. Lett. 2016, 11, 1693–1698. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Qi, J.; Zhang, J.; Wang, F.; Fan, L. Effects of paris polyphylla saponin VII plus silica nano composite on ovarian cancer drug resistance in vitro. Zhonghua Yi Xue Za Zhi 2015, 95, 1859–1861. [Google Scholar] [PubMed]
- Yang, M.; Zou, J.; Zhu, H.; Liu, S.; Wang, H.; Bai, P.; Xiao, X. Paris saponin II inhibits human ovarian cancer cell-induced angiogenesis by modulating NF-κB signaling. Oncol. Rep. 2015, 33, 2190–2198. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Yang, M.; Xiao, J.; Zou, J.; Huang, Q.; Yang, K.; Zhang, B.; Yang, F.; Liu, S.; Wang, H.; et al. Paris Saponin II suppresses the growth of human ovarian cancer xenografts via modulating VEGF-mediated angiogenesis and tumor cell migration. Cancer Chemother. Pharmacol. 2014, 73, 807–818. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Zou, J.; Bui-Nguyen, T.M.; Bai, P.; Gao, L.; Liu, J.; Liu, S.; Xiao, J.; Chen, X.; Zhang, X.; et al. Paris saponin II of Rhizoma Paridis--a novel inducer of apoptosis in human ovarian cancer cells. Biosci. Trends 2012, 6, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.H.; Nao, J.F.; Zhang, M.; He, P. 20(s)-ginsenoside Rg3 promotes apoptosis in human ovarian cancer HO-8910 cells through PI3K/Akt and XIAP pathways. Tumour Biol. 2014, 35, 11985–11994. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liu, T.; Zhao, L.; Chen, W.; Hou, H.; Ye, Z.; Li, X. Ginsenoside 20 (S)-Rg3 inhibits the warburg effect through STAT3 pathways in ovarian cancer cells. Int. J. Oncol. 2015, 46, 775–781. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhao, L.; Zhang, Y.; Chen, W.; Liu, D.; Hou, H.; Ding, L.; Li, X. Ginsenoside 20(S)-Rg3 targets HIF-1α to block hypoxia-induced epithelial-mesenchymal transition in ovarian cancer cells. PLoS ONE 2014, 9, e103887. [Google Scholar] [CrossRef] [PubMed]
- Day, T.W.; Huang, S.; Safa, A.R. c-FLIP knockdown induces ligand-independent DR5-, FADD-,caspase-8-, and caspase-9-dependent apoptosis in breast cancer cells. Biochem. Pharmacol. 2008, 76, 1694–1704. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Gao, Y.; Rankin, G.O.; Rojanasakul, Y.; Cutler, S.J.; Tu, Y.Y.; Chen, Y.C. Chaetoglobosin K induces apoptosis and G2 cell cycle arrest through p53-dependent pathway in cisplatin-resistant ovarian cancer cells. Cancer Lett. 2015, 356, 418–433. [Google Scholar] [CrossRef] [PubMed]
- Ren, W.; Mi, D.; Yang, K.; Cao, N.; Tian, J.; Li, Z.; Bin, M. The expression of hypoxia-inducible factor-1α and its clinical significance in lung cancer: A systematic review and meta-analysis. Swiss Med. Wkly. 2013, 143, w13855. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Not available. |


© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jia, L.-Y.; Wu, X.-J.; Gao, Y.; Rankin, G.O.; Pigliacampi, A.; Bucur, H.; Li, B.; Tu, Y.-Y.; Chen, Y.C. Inhibitory Effects of Total Triterpenoid Saponins Isolated from the Seeds of the Tea Plant (Camellia sinensis) on Human Ovarian Cancer Cells. Molecules 2017, 22, 1649. https://doi.org/10.3390/molecules22101649
Jia L-Y, Wu X-J, Gao Y, Rankin GO, Pigliacampi A, Bucur H, Li B, Tu Y-Y, Chen YC. Inhibitory Effects of Total Triterpenoid Saponins Isolated from the Seeds of the Tea Plant (Camellia sinensis) on Human Ovarian Cancer Cells. Molecules. 2017; 22(10):1649. https://doi.org/10.3390/molecules22101649
Chicago/Turabian StyleJia, Ling-Yan, Xue-Jin Wu, Ying Gao, Gary O. Rankin, Alexa Pigliacampi, Heather Bucur, Bo Li, You-Ying Tu, and Yi Charlie Chen. 2017. "Inhibitory Effects of Total Triterpenoid Saponins Isolated from the Seeds of the Tea Plant (Camellia sinensis) on Human Ovarian Cancer Cells" Molecules 22, no. 10: 1649. https://doi.org/10.3390/molecules22101649