
Anti-HBV Activities of Three Compounds Extracted and Purified from Herpetospermum Seeds


Abstract
:1. Introduction
2. Results
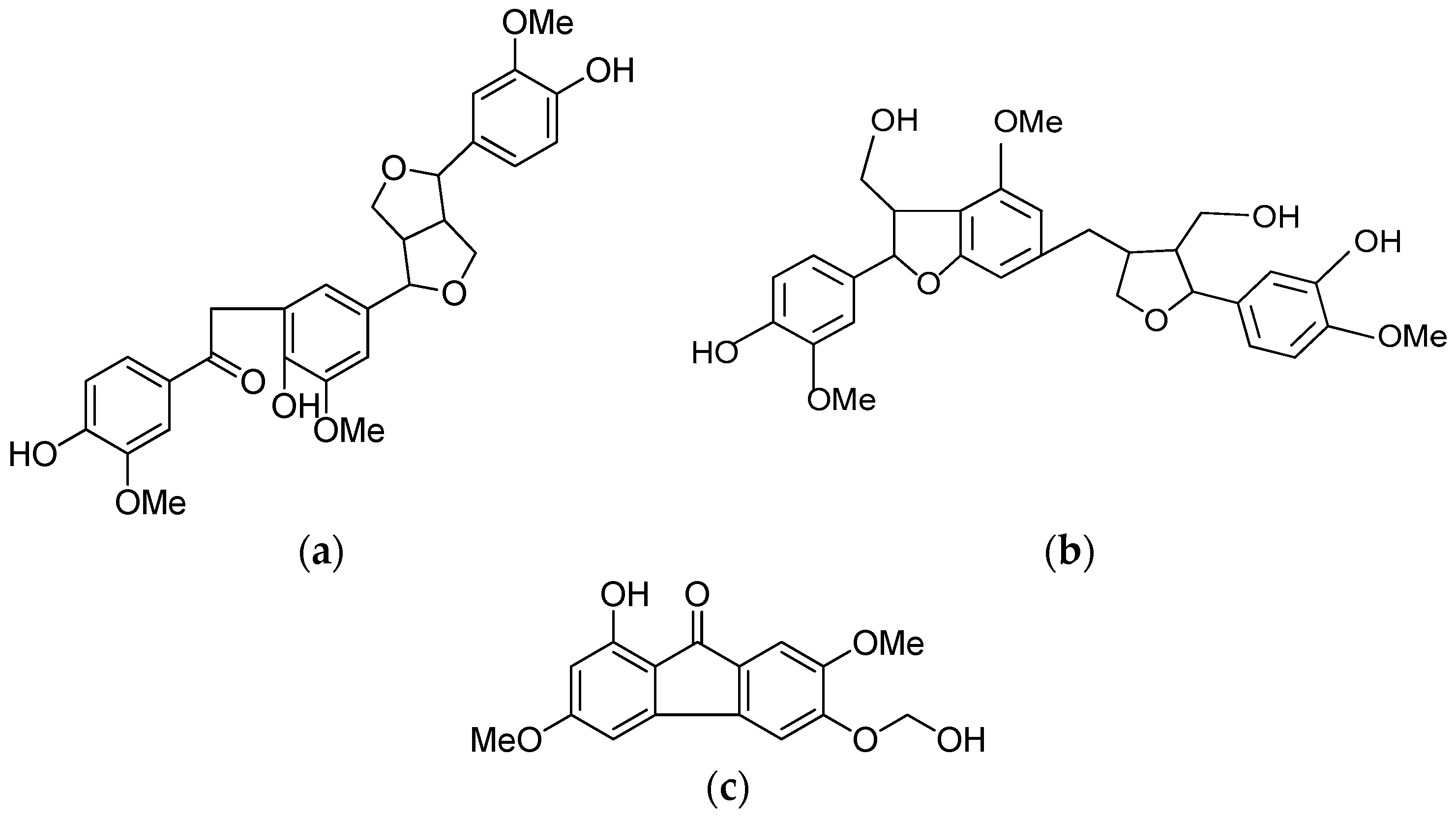
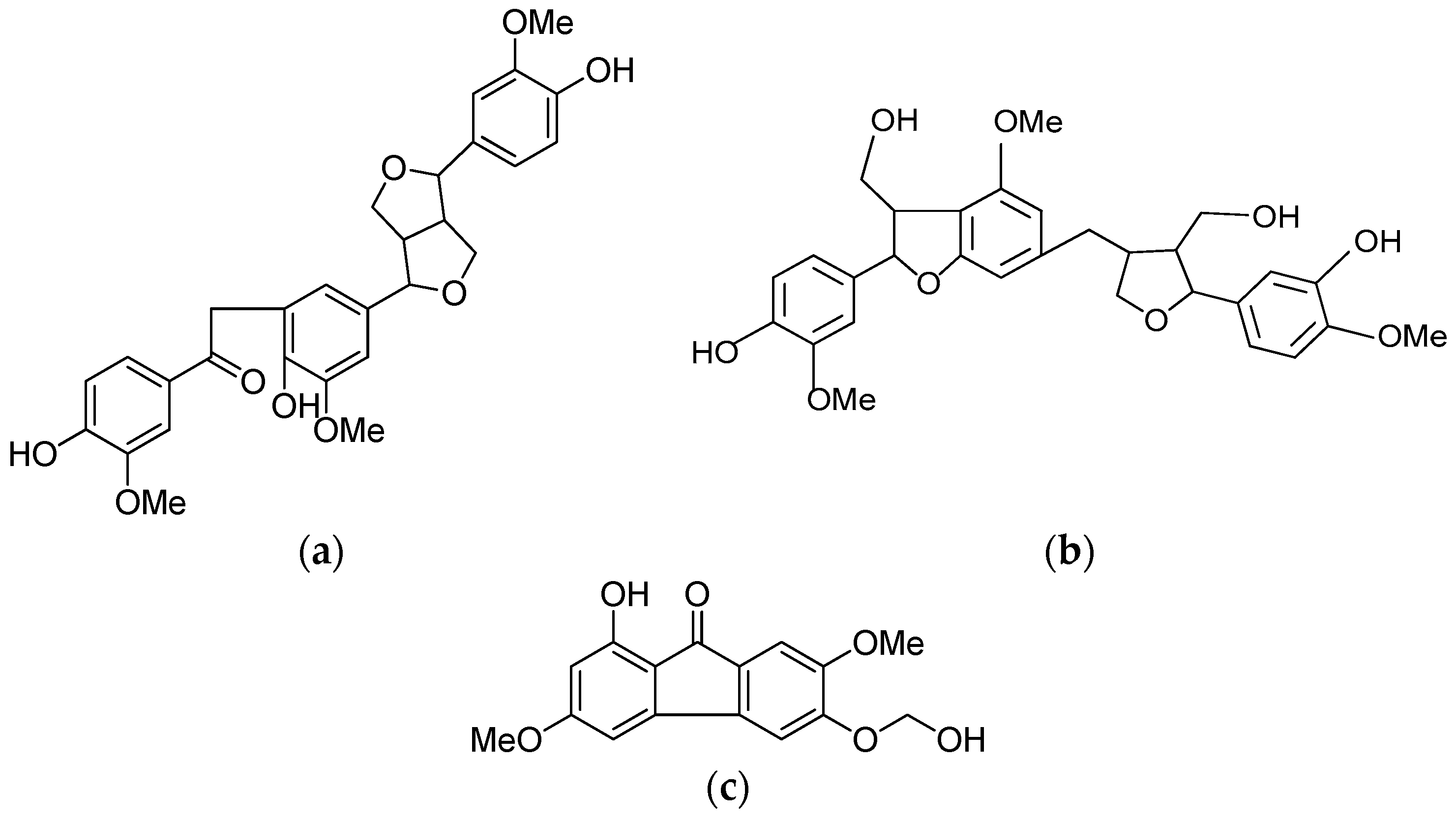
2.1. Isolation and Identification of HPO, HPT and HPF
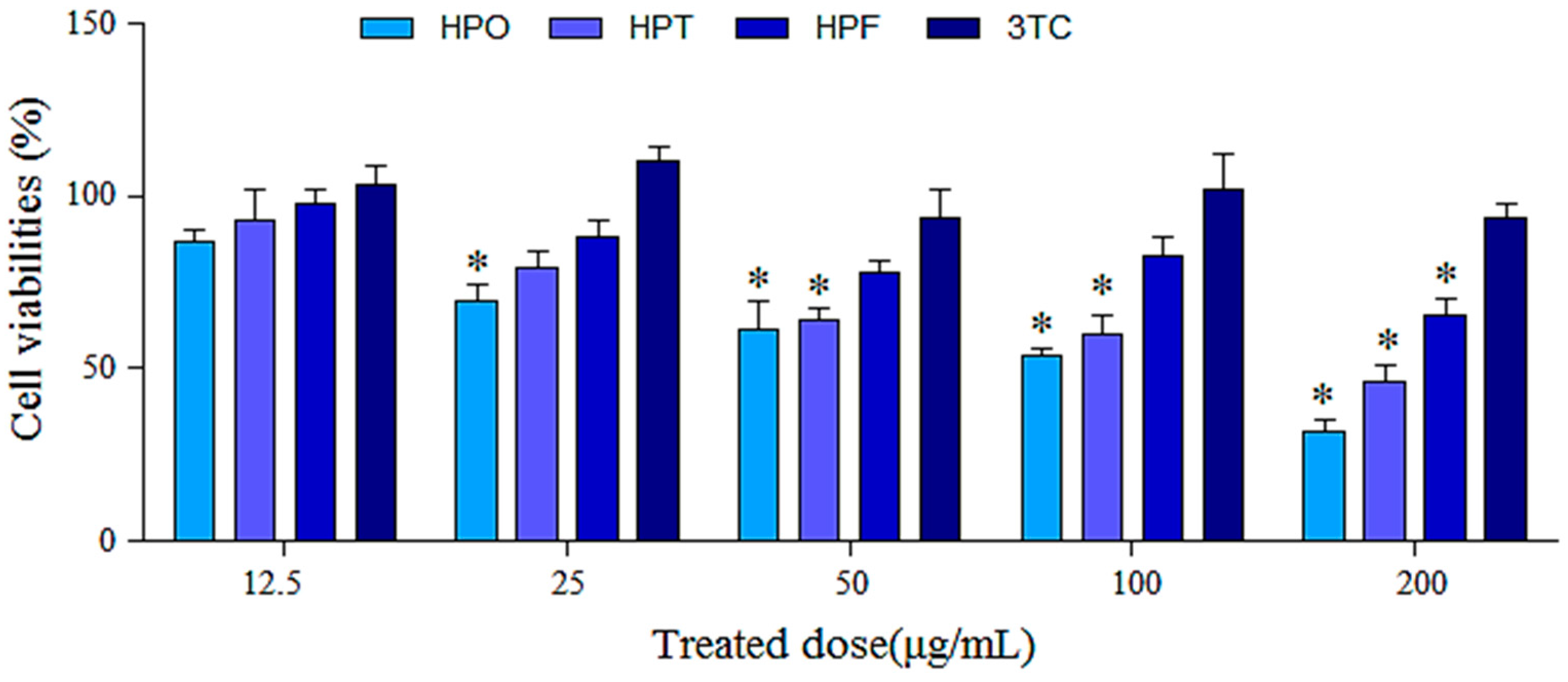
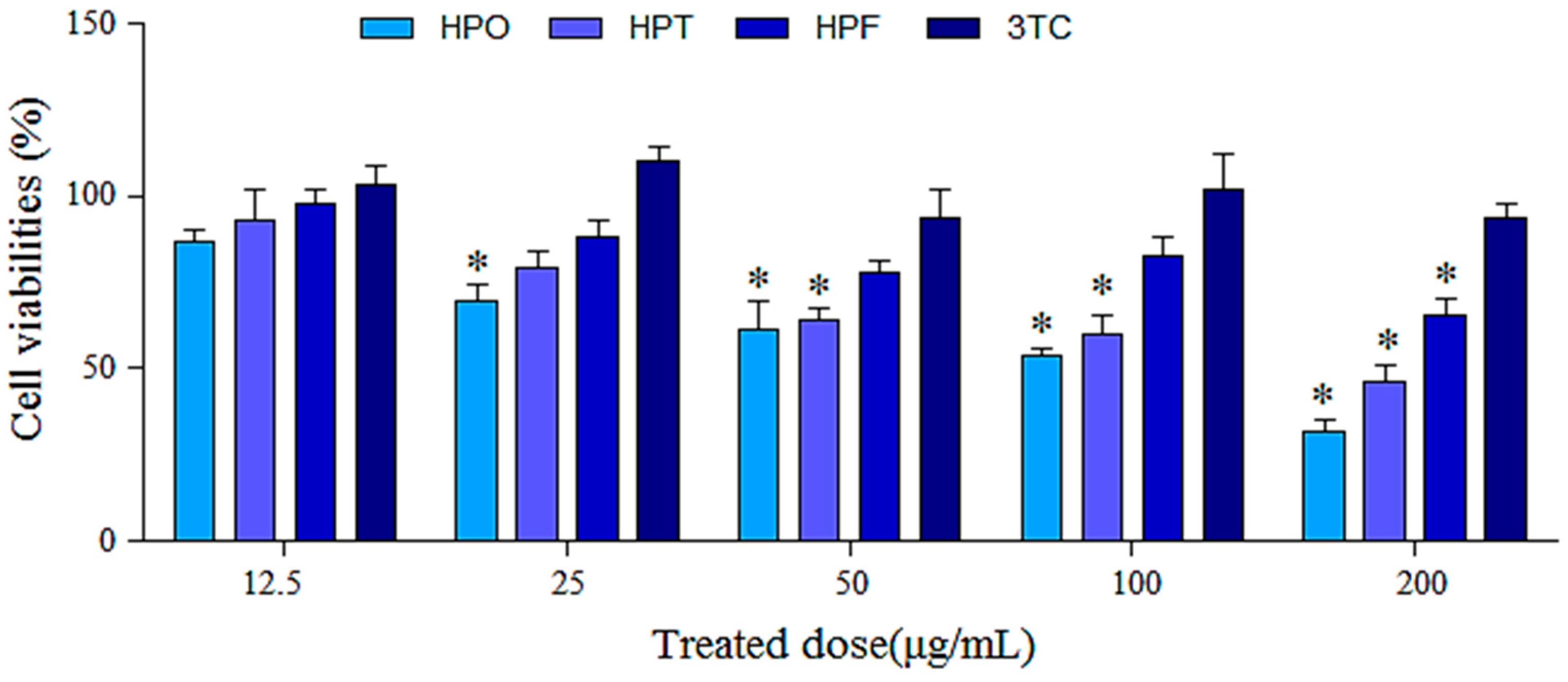
2.2. Cytotoxic Effects of Drugs on HepG2.2.15 Cells
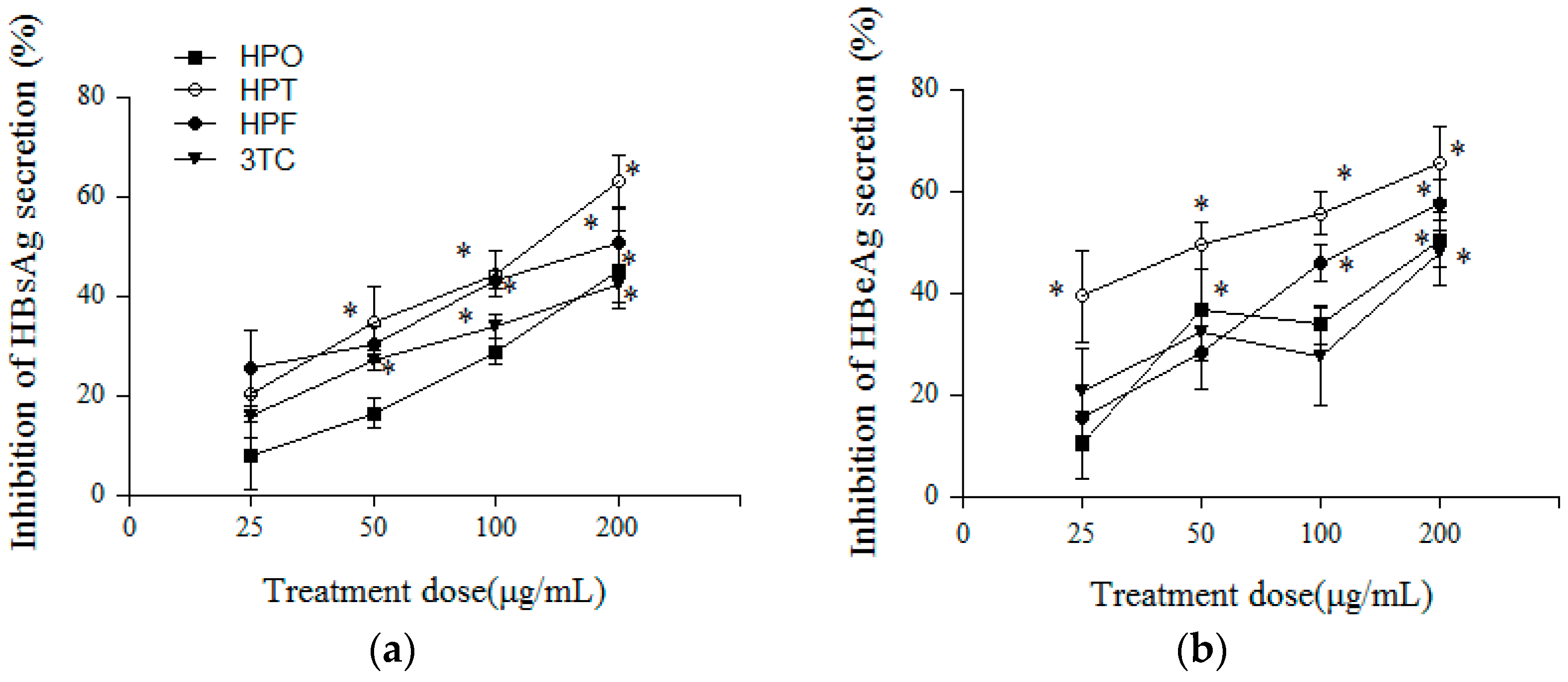
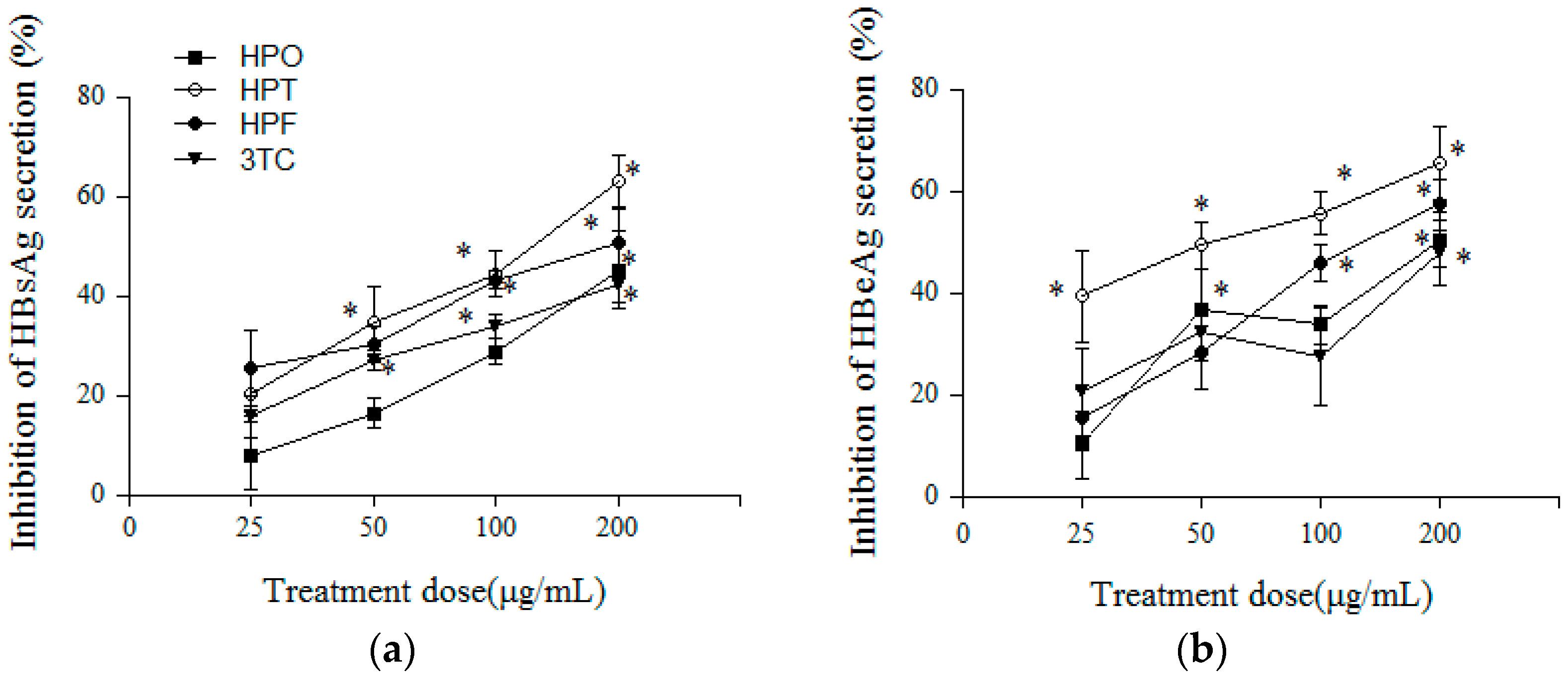
2.3. Inhibitory Effects of Three Compounds and 3TC on HBV Antigens
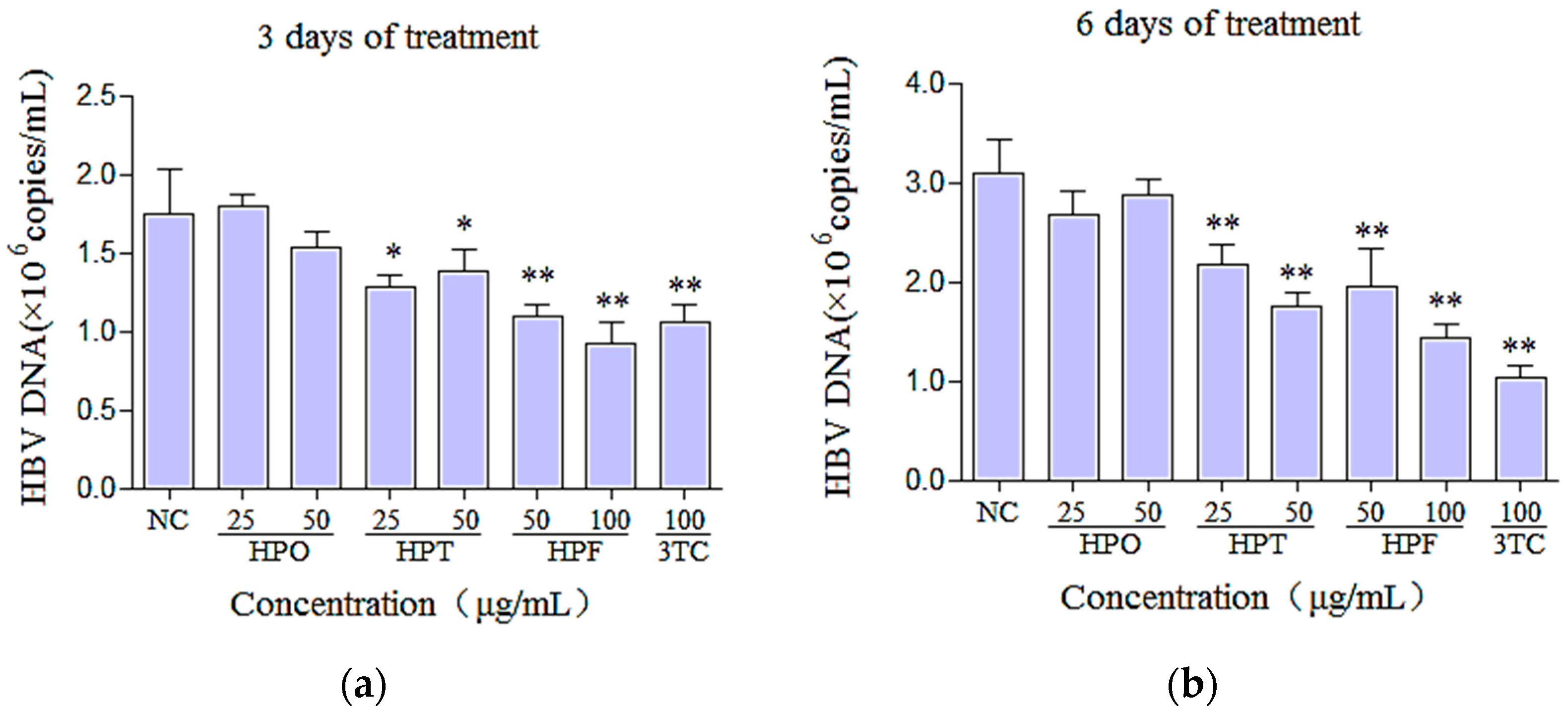
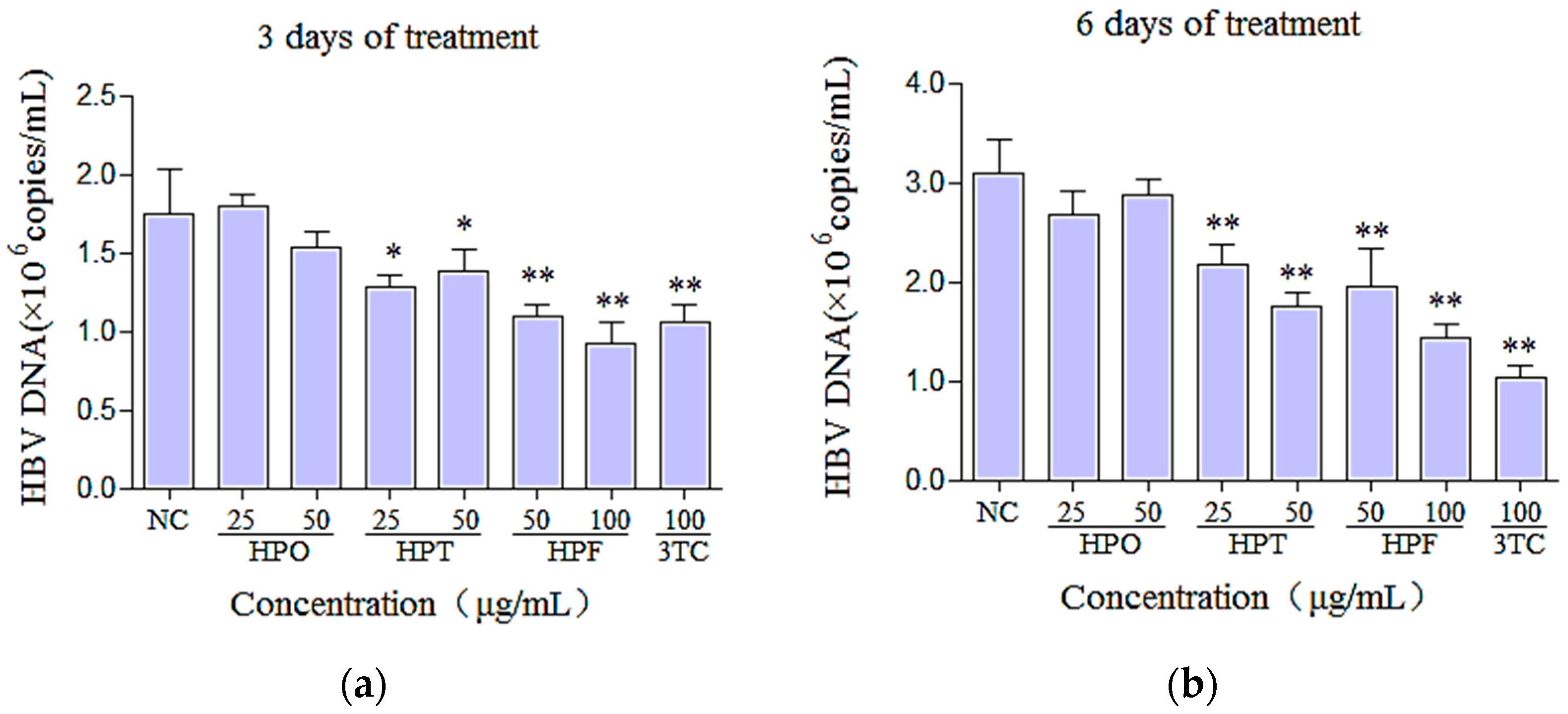
2.4. Effects of Three Compounds and 3TC on the Secretion of HBV DNA
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Plant Material
4.3. Isolation and Identification of Compounds
4.4. Cell Culture and Drug Treatment
4.5. MTT Assay
4.6. Determination of HBeAg and HBsAg
4.7. Determination of HBV DNA in the Supernatant
4.8. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kennedy, E.M.; Bassit, L.C.; Mueller, H.; Kornepati, A.V.; Bogerd, H.P.; Nie, T.; Chatterjee, P.; Javanbakht, H.; Schinazi, R.F.; Cullen, B.R. Suppression of hepatitis B virus DNA accumulation in chronically infected cells using a bacterial CRISPR/Cas RNA-guided DNA endonuclease. Virology 2015, 476, 196–205. [Google Scholar] [CrossRef] [PubMed]
- Park, N.H.; Song, I.H.; Chung, Y.H. Chronic hepatitis B in hepatocarcinogenesis. Postgrad. Med. J. 2006, 82, 507–515. [Google Scholar] [CrossRef] [PubMed]
- Elserag, H.B. Hepatocellular carcinoma: An epidemiologic view. J. Clin. Gastroenterol. 2002, 35, S72–S78. [Google Scholar] [CrossRef]
- Perrillo, R.P. Current treatment of chronic hepatitis B: Benefits and limitations. Semin. Liver Dis. 2005, 25 (Suppl. 1), 20–28. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Sun, L.M.; Liu, X.Q.; Li, B.; Wang, Q.; Dong, J.X. Anti-HBV active flavone glucosides from Euphorbia humifusa Willd. Fitoterapia 2010, 81, 799–802. [Google Scholar] [CrossRef] [PubMed]
- Parvez, M.K.; Arab, A.H.; Aldosari, M.S.; Alrehaily, A.J. Antiviral natural products against chronic hepatitis B: Recent developments. Curr. Pharm. Des. 2016, 22, 286–293. [Google Scholar] [CrossRef] [PubMed]
- Mcculloch, M.; Broffman, M.; Gao, J.; Colford, J.M., Jr. Chinese herbal medicine and interferon in the treatment of chronic hepatitis B: A meta-analysis of randomized, controlled trials. Am. J. Public Health 2002, 92, 1619–1628. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Li, J.; Huang, H.; Gao, W.; Zhuang, C.; Li, B.; Zhou, P.; Kong, D. Anti-hepatitis B virus activities of astragaloside IV isolated from Radix Astragali. Biol. Pharm. Bull. 2009, 32, 132–135. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Zhao, L.; You, Q.; Yang, Y.; Gu, H.; Song, G.; Lu, N.; Xin, J. Anti-hepatitis B virus activity of wogonin in vitro and in vivo. Antivir. Res. 2007, 74, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Wang, G. A review of non-nucleoside anti-hepatitis B virus agents. Eur. J. Med. Chem. 2014, 75, 267–281. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Zhang, Y.; Ding, X.R.; Chen, S.H.; Yang, J.; Wang, X.J.; Jia, G.L.; Chen, H.S.; Bo, X.C.; Wang, S.Q. Protocatechuic aldehyde inhibits hepatitis B virus replication both in vitro and in vivo. Antivir. Res. 2007, 74, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.; Yuan, Z.; Tan, R.; Zhang, X. Isolation of herpetin from Herpetospermum seed and hepatoprotective activity of liposomal herpetin against carbon tetrachloride-induced liver injury in mice. Pharmazie 2015, 70, 745–752. [Google Scholar] [PubMed]
- Zhang, M.; Dong, X.P.; Deng, Y.; Wang, H.; Li, X.N.; Song, Q. A new sesqui-norlignan from Herpetospermum pedunculosum. Acta Pharm. Sin. 2006, 41, 659–661. [Google Scholar]
- Jin, S.Y.; Lv, J.L.; Yuan, H.L.; Jin, S.X.; Fu, S.S.; Guo, J.J. Studies on anti-fatigue and anoxia-resistant effects of Herpetospermum caudigerum extracts in mice. Pharm. J. Chin. Peoples Lib. Army 2011, 27, 417–418. [Google Scholar]
- Gang, S.; Shen, C.Y.; Ling, C.; Zhang, L.H.; Ling, Q.; Juan-Juan, L.I.; Yuan, H.L.; Jin, H. Hepatoprotective effects of total lignans of Herpetospermum caudigerum nanosuspension capsules. J. Chin. Pharm. Sci. 2015, 50, 1038–1042. [Google Scholar]
- Li, L.; Yuan, H.L.; Wu, Q. Clinical study on effect of ganneng dripping pill in treating chronic hepatitis B. Chin. J. Integr. Tradit. West. Med. 2006, 26, 788–790. [Google Scholar]
- Yuan, H.L.; Yang, M.; Li, X.Y.; You, R.H.; Liu, Y.; Zhu, J.; Xie, H.; Xiao, X.H. Hepatitis B virus inhibiting constituents from Herpetospermum caudigerum. ChemInform 2007, 54, 1592–1594. [Google Scholar] [CrossRef]
- Wen, G.J.; Fan, X.J.; Zhang, C.T.; Feng, Q.S.; Jiang, W.U.; Zhou, X.H. Discussion about traditional Chinese medicine anti-hepatitis B virus and its research ideas. Lishizhen Med. Mater. Med. Res. 2012, 1, 096. [Google Scholar]
- Marcellin, P.; Asselah, T. Viral hepatitis: Impressive advances but still a long way to eradication of the disease. Liver Int. 2014, 34 (Suppl. 1), 1–3. [Google Scholar] [CrossRef] [PubMed]
- Jian, G.U.; Jia-Chuan, L.I.; Fan, L.N. The protective effect and its mechanism of total Lignans from Tibetan medicinal Herpetospermum seeds on Concanavalin A-induced liver injury in mice. J. Southwest Univ. Natl. 2014, 3, 010. [Google Scholar]
- Shen, B.; Chen, H.; Shen, C.; Xu, P.; Li, J.; Shen, G.; Yuan, H.; Han, J. Hepatoprotective effects of lignans extract from Herpetospermum caudigerum against CCl4-induced acute liver injury in mice. J. Ethnopharmacol. 2015, 164, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Ringelhan, M.; Heikenwalder, M.; Protzer, U. Direct effects of hepatitis B virus-encoded proteins and chronic infection in liver cancer development. Dig. Dis. 2013, 31, 138–151. [Google Scholar] [CrossRef] [PubMed]
- Mirko Tarocchi, S.P.; Marroncini, G.; Galli, A. Molecular mechanism of hepatitis B virus-induced hepatocarcinogenesis. World J. Gastroenterol. 2014, 20, 11630–11640. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Abdel-Hafiz, H.; Suhail, M.; Al-Mars, A.; Zakaria, M.K.; Fatima, K.; Ahmad, S.; Azhar, E.; Chaudhary, A.; Qadri, I. Hepatitis B virus, HBx mutants and their role in hepatocellular carcinoma. World J. Gastroenterol. 2014, 7, 10238–10248. [Google Scholar] [CrossRef] [PubMed]
- Levrero, M.; Zucman-Rossi, J. Mechanisms of HBV-induced hepatocellular carcinoma. J. Hepatol. 2016, 64, S84–S101. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Cui, L.; Wang, Y.; Yang, G.; He, J.; Hao, R.; Fan, C.; Qu, M.; Liu, Z.; Wang, M. Hepatitis B e antigen and its precursors promote the progress of hepatocellular carcinoma by interacting with NUMB and decreasing p53 activity. Hepatology 2016, 64, S386–S387. [Google Scholar] [CrossRef]
- Geng, C.-A.; Wang, L.-J.; Zhang, X.-M.; Ma, Y.-B.; Huang, X.-Y.; Luo, J.; Guo, R.-H.; Zhou, J.; Shen, Y.; Zuo, A.-X.; et al. Anti-hepatitis B virus active lactones from the traditional Chinese herb: Swertia mileensis. Chemistry 2011, 17, 3893–3903. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the compounds (HPO, HPT, and HPF) are available from the authors.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | TC50 (μg/mL) | HBsAg | HBeAg | ||
|---|---|---|---|---|---|
| IC50 (μg/mL) | TI | IC50 (μg/mL) | TI | ||
| HPT | 146.74 | 114.48 | 1.28 | 57.75 | 2.54 |
| HPO | 91.15 | 240.31 | 0.38 | 185.64 | 0.49 |
| HPF | 469.92 | 176.99 | 2.66 | 134.53 | 3.49 |
| 3TC | - | 305.38 | - | 312.00 | - |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gong, P.-Y.; Yuan, Z.-X.; Gu, J.; Tan, R.; Li, J.-C.; Ren, Y.; Hu, S. Anti-HBV Activities of Three Compounds Extracted and Purified from Herpetospermum Seeds. Molecules 2017, 22, 14. https://doi.org/10.3390/molecules22010014
Gong P-Y, Yuan Z-X, Gu J, Tan R, Li J-C, Ren Y, Hu S. Anti-HBV Activities of Three Compounds Extracted and Purified from Herpetospermum Seeds. Molecules. 2017; 22(1):14. https://doi.org/10.3390/molecules22010014
Chicago/Turabian StyleGong, Pu-Yang, Zhi-Xiang Yuan, Jian Gu, Rui Tan, Jia-Chuan Li, Yan Ren, and Sha Hu. 2017. "Anti-HBV Activities of Three Compounds Extracted and Purified from Herpetospermum Seeds" Molecules 22, no. 1: 14. https://doi.org/10.3390/molecules22010014