
1-(2,3-Dibenzimidazol-2-ylpropyl)-2-methoxybenzene Is a Syk Inhibitor with Anti-Inflammatory Properties

 ,
, 
Abstract
:1. Introduction
2. Results
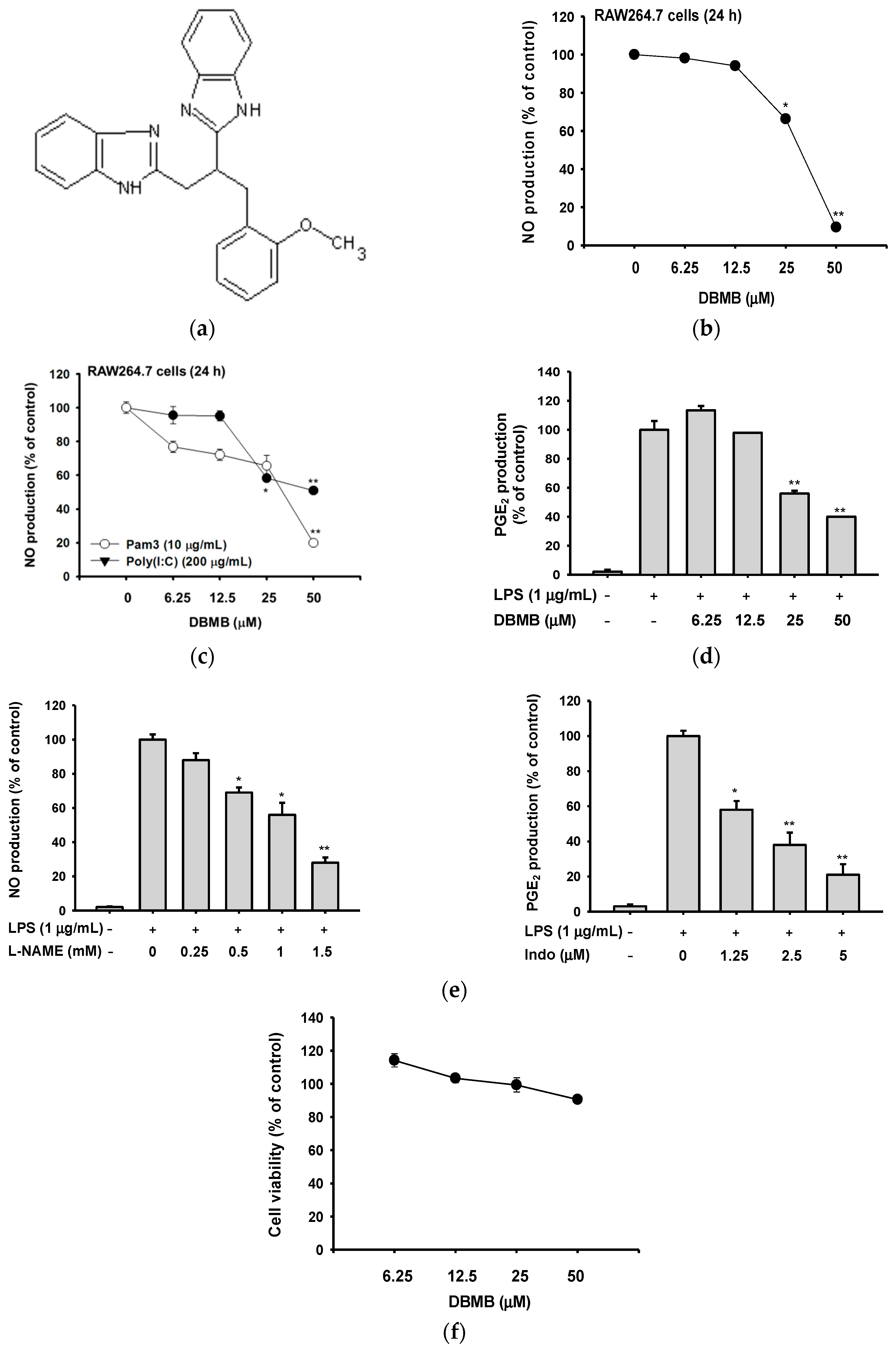
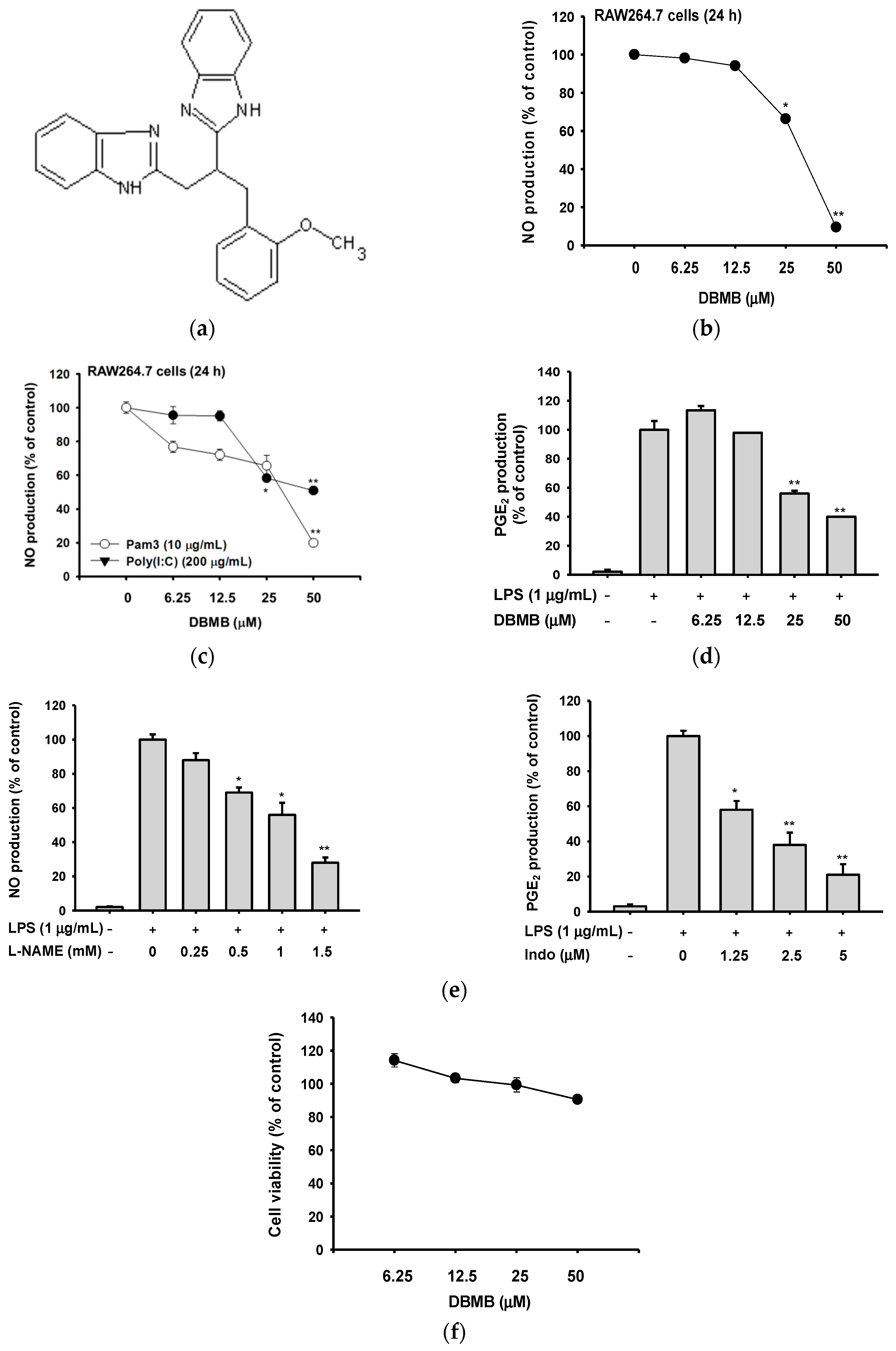
2.1. DBMB Suppressed the Production of Pro-Inflammatory Mediators
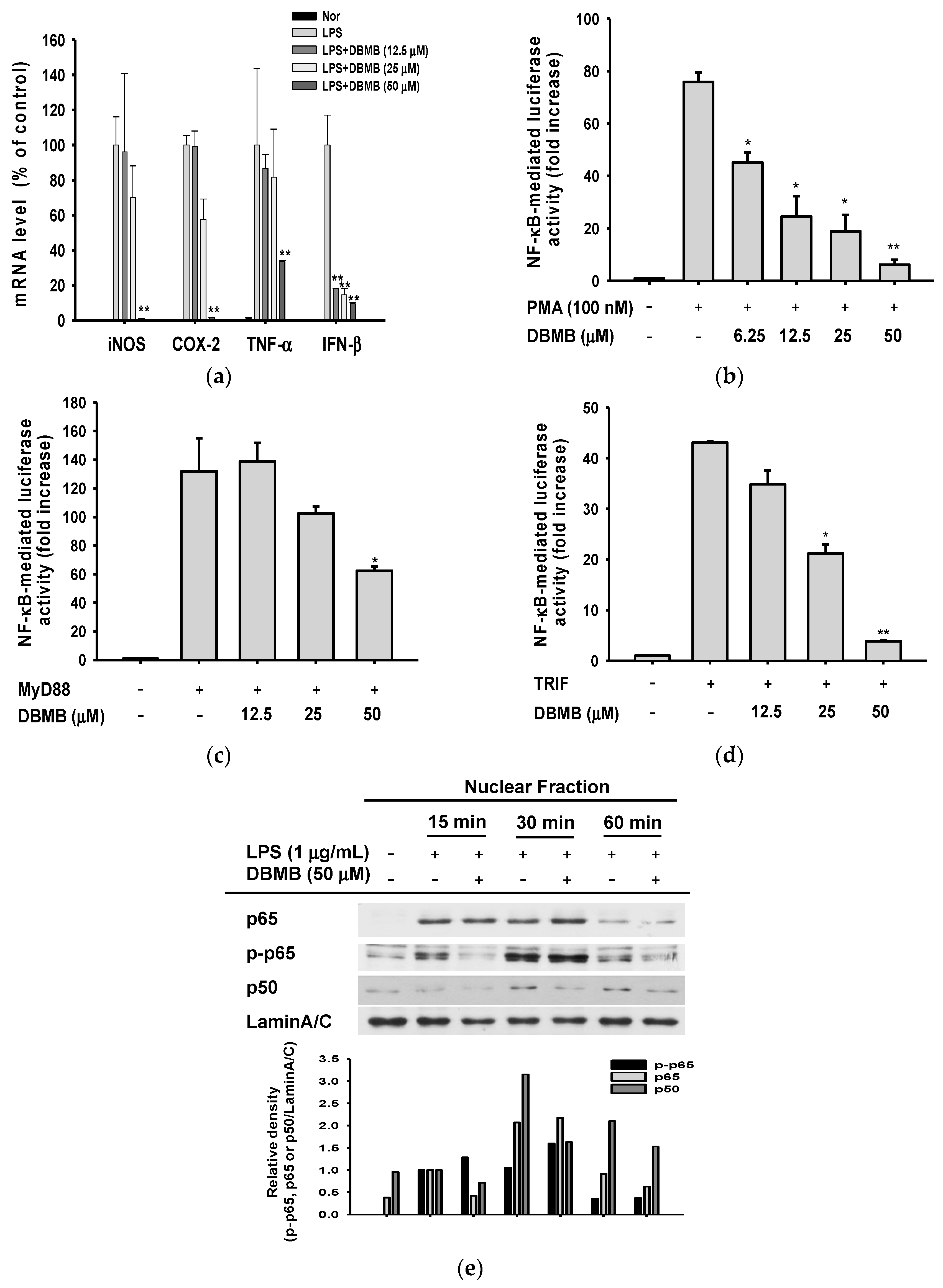
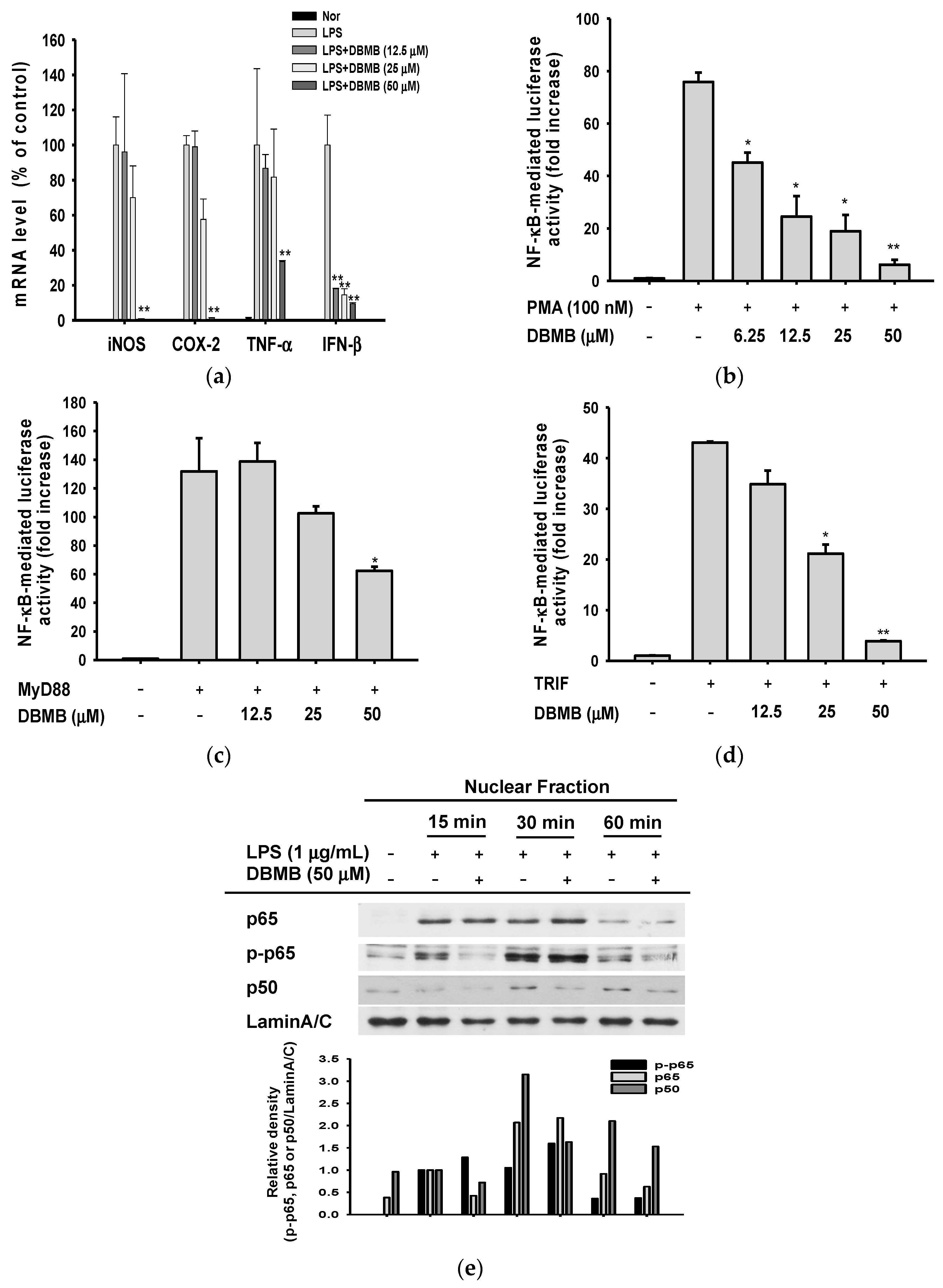
2.2. DBMB Inhibited Inflammatory Responses at the Transcriptional Level
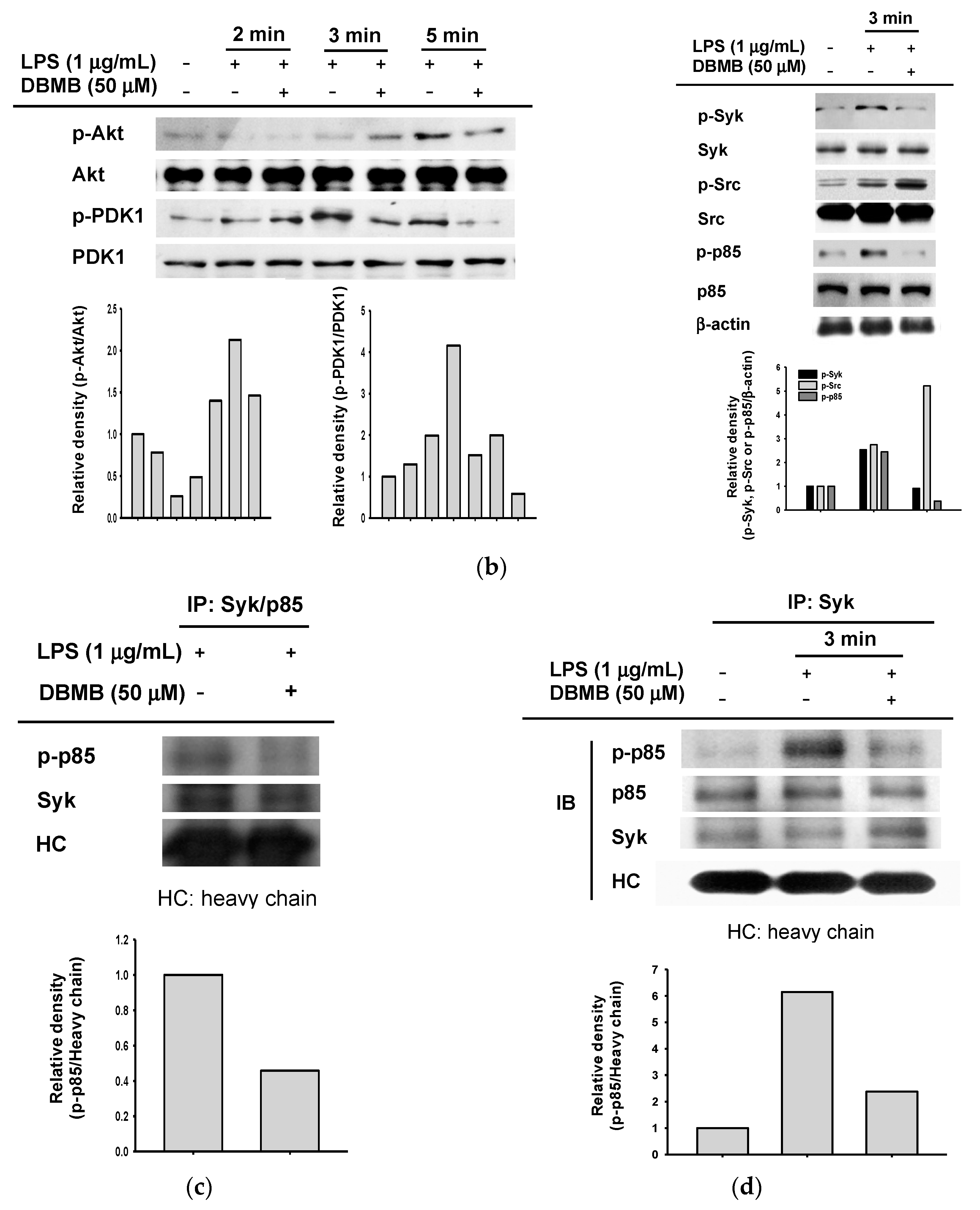
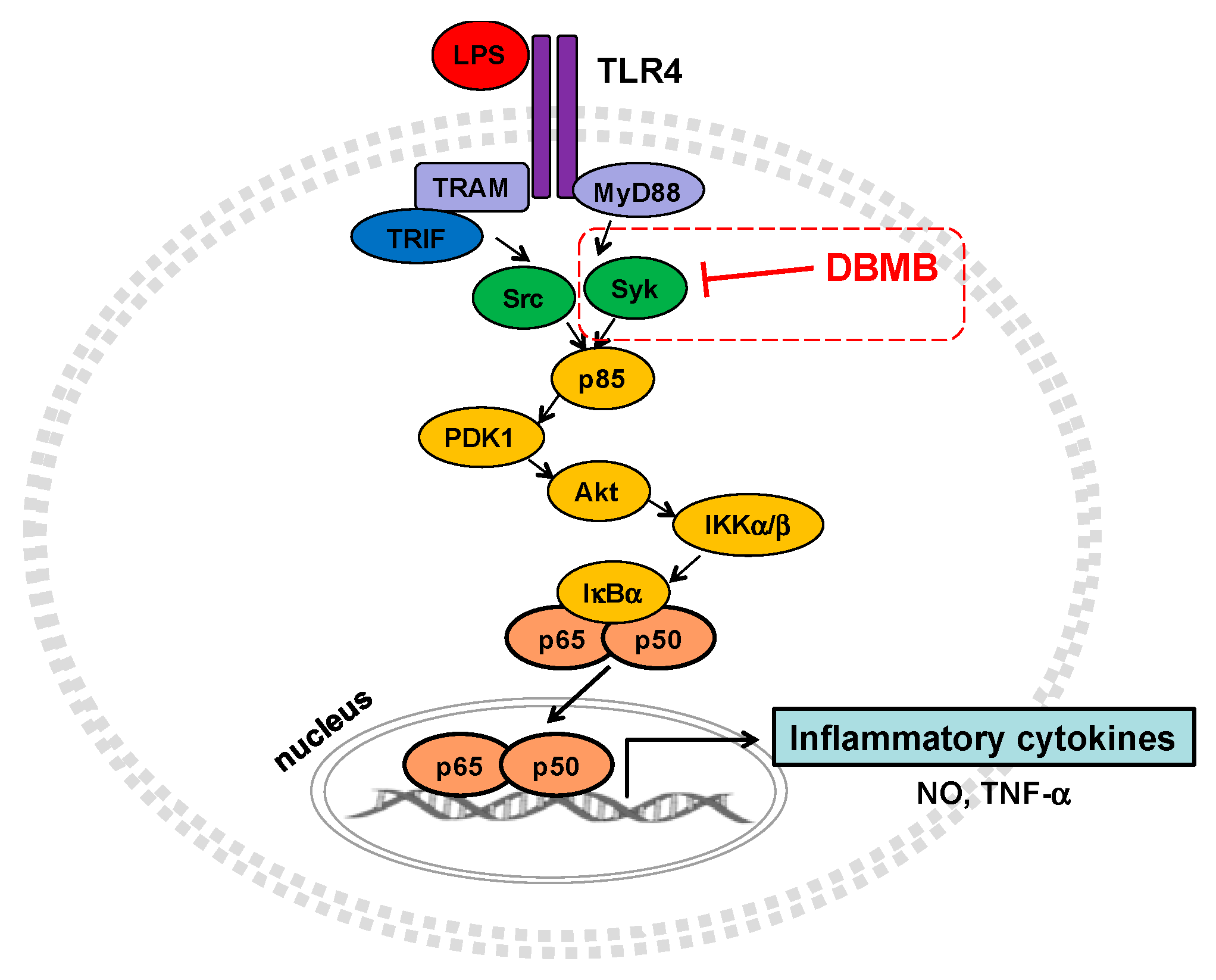
2.3. The NF-κB Signaling Cascade Was Modulated by DBMB
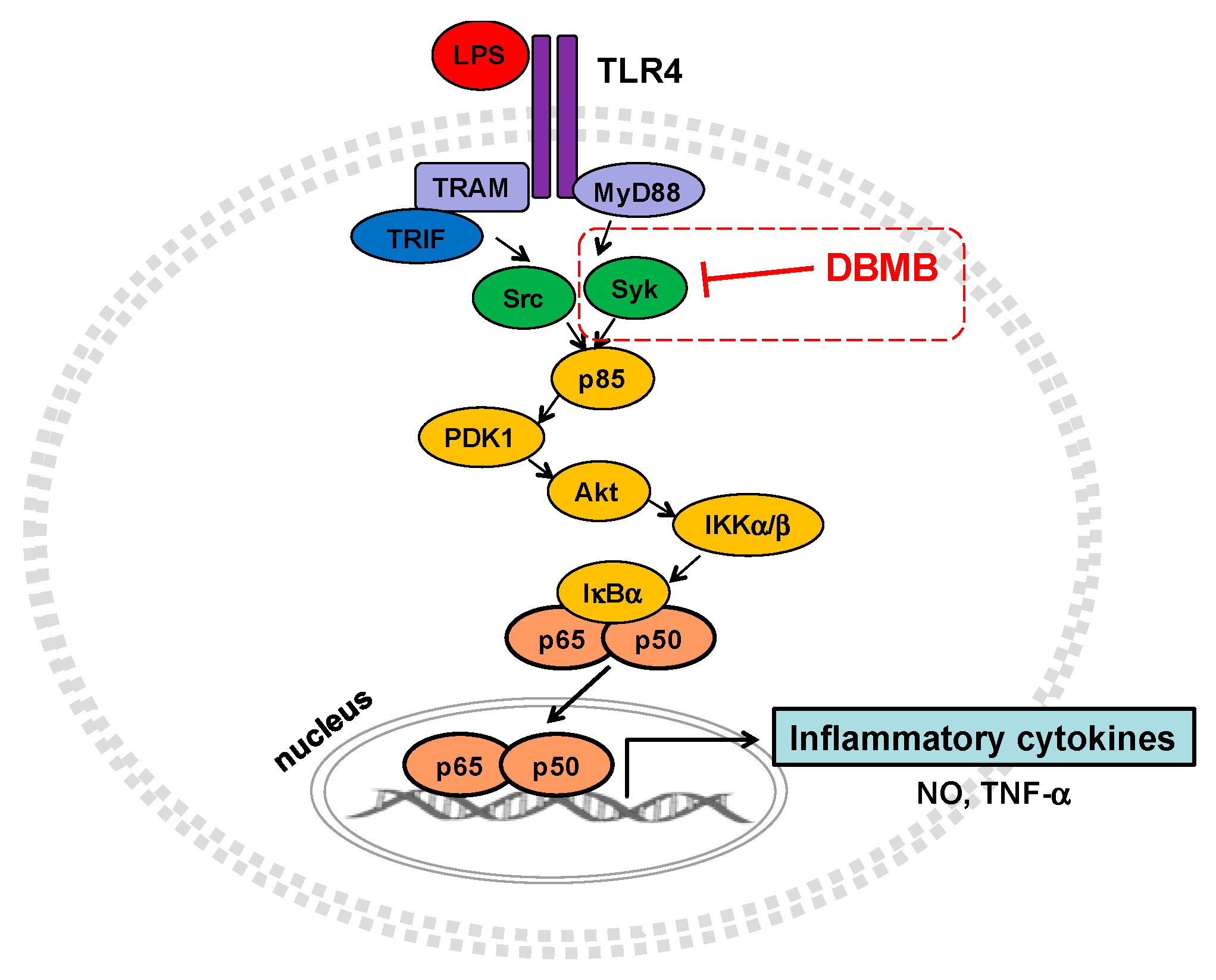
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture
4.3. NO and PGE2 Production
4.4. Cell Viability Test
4.5. mRNA Analysis by Quantitative Real-Time PCR
4.6. Preparation of Total Lysates, Nuclear Extracts, and Immunoblotting
4.7. Transfection of DNA and Luciferase Reporter Gene Activity Assay
4.8. In Vitro Syk Kinase Assay Using Immunoprecipitated Proteins
4.9. Immunoprecipitation
4.10. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| MTT | (3-4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| DBMB | 1-(2,3-dibenzimidazol-2-ylpropyl)-2-methoxybenzene |
| COX-2 | cyclooxygenase-2 |
| FBS | fetal bovine serum |
| Indo | indomethacin |
| iNOS | inducible nitric oxide synthase |
| IKK | IκB kinase |
| LPS | lipopolysaccharide |
| PAMPs | pathogen-associated molecular patterns |
| PBS | phosphate-buffered saline |
| PRRs | pattern recognition receptors |
| PI3K | phosphoinositide-3-kinase |
| PDK1 | phosphoinositide-dependent kinase-1 |
| Akt | protein kinase B |
| PKC | protein kinase C |
| TLRs | toll-like receptors |
| TNF | tumor necrosis factor |
References
- Kim, E.; Yang, W.S.; Kim, J.H.; Park, J.G.; Kim, H.G.; Ko, J.; Hong, Y.D.; Rho, H.S.; Shin, S.S.; Sung, G.-H. Lancemaside A from Codonopsis lanceolata modulates the inflammatory responses mediated by monocytes and macrophages. Mediat. Inflamm. 2014, 2014, 405158. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, O.; Akira, S. Pattern recognition receptors and inflammation. Cell 2010, 140, 805–820. [Google Scholar] [CrossRef] [PubMed]
- Sabroe, I.; Parker, L.; Dower, S.; Whyte, M. The role of TLR activation in inflammation. J. Pathol. 2008, 214, 126–135. [Google Scholar] [CrossRef] [PubMed]
- De Nardo, D. Toll-like receptors: Activation, signalling and transcriptional modulation. Cytokine 2015, 74, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Newton, K.; Dixit, V.M. Signaling in innate immunity and inflammation. Cold Spring Harbor Perspect. Biol. 2012, 4, a006049. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Xiao, Y.; Hu, H.; Zou, Q.; Li, Y.; Gao, Y.; Ge, W.; Cheng, X.; Sun, S.-C. Proinflammatory TLR signalling is regulated by a TRAF2-dependent proteolysis mechanism in macrophages. Nat. Commun. 2015, 6, 5930. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.H.; Park, J.; Kim, M. Gut microbiota-derived short-chain fatty acids, T cells, and inflammation. Immune Netw. 2014, 14, 277–288. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, F.; Mages, J.; Heit, A.; Lang, R.; Wagner, H. Transcriptional activation induced in macrophages by Toll-like receptor (TLR) ligands: From expression profiling to a model of TLR signaling. Eur. J. Immunol. 2004, 34, 2863–2873. [Google Scholar] [CrossRef] [PubMed]
- Yi, Y.S.; Son, Y.J.; Ryou, C.; Sung, G.H.; Kim, J.H.; Cho, J.Y. Functional roles of Syk in macrophage-mediated inflammatory responses. Mediat. Inflamm. 2014, 2014, 270302. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.Y.; Kim, J.H.; Cho, J.Y. Cytochalasin B modulates macrophage-mediated inflammatory responses. Biomol. Ther. 2014, 22, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.S.; Lee, J.; Kim, T.W.; Kim, J.H.; Lee, S.; Rhee, M.H.; Hong, S.; Cho, J.Y. Src/NF-κB-targeted inhibition of LPS-induced macrophage activation and dextran sodium sulphate-induced colitis by Archidendron clypearia methanol extract. J. Ethnopharmacol. 2012, 142, 287–293. [Google Scholar] [CrossRef] [PubMed]
- Takada, Y.; Mukhopadhyay, A.; Kundu, G.C.; Mahabeleshwar, G.H.; Singh, S.; Aggarwal, B.B. Hydrogen peroxide activates NF-κB through tyrosine phosphorylation of IκBα and serine phosphorylation of p65 evidence for the involvement of IκBα kinase and Syk protein-tyrosine kinase. J. Biol. Chem. 2003, 278, 24233–24241. [Google Scholar] [CrossRef] [PubMed]
- Hoesel, B.; Schmid, J.A. The complexity of NF-κB signaling in inflammation and cancer. Mol. Cancer 2013, 12, 86. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.; Kim, B.M.; Chang, C.H. Co-stimulation of TLR4 and Dectin-1 induces the production of inflammatory cytokines but not TGF-β for Th17 cell differentiation. Immune Netw. 2014, 14, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-B.; Han, A.-R.; Park, E.-Y.; Kim, J.-Y.; Cho, W.; Lee, J.; Seo, E.-K.; Lee, K.-T. Inhibition of LPS-induced iNOS, COX-2 and cytokines expression by poncirin through the NF-κB inactivation in RAW 264.7 macrophage cells. Biol. Pharm. Bull. 2007, 30, 2345–2351. [Google Scholar] [CrossRef] [PubMed]
- Byeon, S.E.; Yi, Y.-S.; Oh, J.; Yoo, B.C.; Hong, S.; Cho, J.Y. The role of Src kinase in macrophage-mediated inflammatory responses. Mediat. Inflamm. 2012, 2012, 512926. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Yu, T.; Lian, Y.J.; Ma, R.; Yang, S.; Cho, J.Y. Nitric oxide synthase inhibitors: A review of patents from 2011 to the present. Expert Opin. Ther. Pat. 2015, 25, 49–68. [Google Scholar] [CrossRef] [PubMed]
- Zamora, R.; Vodovotz, Y.; Billiar, T.R. Inducible nitric oxide synthase and inflammatory diseases. Mol. Med. 2000, 6, 347–373. [Google Scholar] [PubMed]
- Ricciotti, E.; FitzGerald, G.A. Prostaglandins and inflammation. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 986–1000. [Google Scholar] [CrossRef] [PubMed]
- Siegfried, A.; Berchtold, S.; Manncke, B.; Deuschle, E.; Reber, J.; Ott, T.; Weber, M.; Kalinke, U.; Hofer, M.J.; Hatesuer, B. IFIT2 is an effector protein of type I IFN–mediated amplification of lipopolysaccharide (LPS)-induced TNF-α secretion and LPS-induced endotoxin shock. J. Immunol. 2013, 191, 3913–3921. [Google Scholar] [CrossRef] [PubMed]
- Kang, C.-D.; Han, C.-S.; Kim, K.-W.; Do, I.-R.; Kim, C.-M.; Kim, S.-H.; Lee, E.-Y.; Chung, B.-S. Activation of NF-κB mediates the PMA-induced differentiation of K562 cells. Cancer Lett. 1998, 132, 99–106. [Google Scholar] [CrossRef]
- Jones, E.; Adcock, I.M.; Ahmed, B.Y.; Punchard, N.A. Modulation of LPS stimulated NF-κB mediated nitric oxide production by PKCepsilon and JAK2 in RAW macrophages. J. Inflamm. 2007, 4, 23. [Google Scholar] [CrossRef] [PubMed]
- Viatour, P.; Merville, M.-P.; Bours, V.; Chariot, A. Phosphorylation of NF-κB and IκB proteins: Implications in cancer and inflammation. Trends Biochem. Sci. 2005, 30, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Tak, P.P.; Firestein, G.S. NF-κB: A key role in inflammatory diseases. J. Clin. Investig. 2001, 107, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.; Yoon, K.D.; Lee, W.-S.; Yang, W.S.; Kim, S.H.; Sung, N.Y.; Baek, K.-S.; Kim, Y.; Htwe, K.M.; Kim, Y.-D. Syk/Src-targeted anti-inflammatory activity of Codariocalyx motorius ethanolic extract. J. Ethnopharmacol. 2014, 155, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.Y.; Jeong, H.Y.; Kim, S.H.; Kim, H.G.; Nam, G.; Kim, J.P.; Yoon, D.H.; Hwang, H.; Kimc, T.W.; Hong, S. Methanol extract of Evodia lepta displays Syk/Src-targeted anti-inflammatory activity. J. Ethnopharmacol. 2013, 148, 999–1007. [Google Scholar] [CrossRef] [PubMed]
- Hatton, O.; Lambert, S.L.; Krams, S.M.; Martinez, O.M. Src kinase and Syk activation initiate PI3K signaling by a chimeric latent membrane protein 1 in Epstein-Barr virus (EBV) + B cell lymphomas. PLoS ONE 2012, 7, e42610. [Google Scholar] [CrossRef] [PubMed]
- Beitz, L.O.; Fruman, D.A.; Kurosaki, T.; Cantley, L.C.; Scharenberg, A.M. SYK is upstream of phosphoinositide 3-kinase in B cell receptor signaling. J. Biol. Chem. 1999, 274, 32662–32666. [Google Scholar] [CrossRef] [PubMed]
- Jiang, K.; Zhong, B.; Gilvary, D.L.; Corliss, B.C.; Vivier, E.; Hong-Geller, E.; Wei, S.; Djeu, J.Y. Syk regulation of phosphoinositide 3-kinase-dependent NK cell function. J. Immunol. 2002, 168, 3155–3164. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Park, J.G.; Lee, J.; Yang, W.S.; Park, G.W.; Kim, H.G.; Yi, Y.-S.; Baek, K.-S.; Sung, N.Y.; Hossen, M.J. The dietary flavonoid kaempferol mediates anti-inflammatory responses via the Src, Syk, IRAK1, and IRAK4 molecular targets. Mediat. Inflamm. 2015, 2015, 904142. [Google Scholar] [CrossRef] [PubMed]
- Perkins, N.D. The diverse and complex roles of NF-κB subunits in cancer. Nat. Rev. Cancer 2012, 12, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Haseebuddin, M.; Young, M.; Colburn, N.H. Suppression of p65 phosphorylation coincides with inhibition of IκBα polyubiquitination and degradation. Mol. Carcinog. 2005, 44, 274–284. [Google Scholar] [CrossRef] [PubMed]
- Michael, K. How NF-kB is activated: The role of the IkB kinase (IKK) complex. Oncogene 1999, 18, 6867–6874. [Google Scholar]
- Lawrence, T. The nuclear factor NF-κB pathway in inflammation. Cold Spring Harbor Perspect. Biol. 2009, 1, a001651. [Google Scholar] [CrossRef] [PubMed]
- Kwon, D.J.; Bae, Y.S.; Ju, S.M.; Youn, G.S.; Choi, S.Y.; Park, J. Salicortin suppresses lipopolysaccharide-stimulated inflammatory responses via blockade of NF-κB and JNK activation in RAW 264.7 macrophages. BMB Rep. 2014, 47, 318–323. [Google Scholar] [CrossRef] [PubMed]
- Caamano, J.; Hunter, C.A. NF-κB family of transcription factors: Central regulators of innate and adaptive immune functions. Clin. Microbiol. Rev. 2002, 15, 414–429. [Google Scholar] [CrossRef] [PubMed]
- Hwang, Y.J.; Song, J.; Kim, H.R.; Hwang, K.A. Oleanolic acid regulates NF-κB signaling by suppressing MafK expression in RAW 264.7 cells. BMB Rep. 2014, 47, 524–529. [Google Scholar] [CrossRef] [PubMed]
- Crawford, J.M.; Watanabe, K. Cell adhesion molecules in inflammation and immunity: Relevance to periodontal diseases. Crit. Rev. Oral Biol. Med. 1994, 5, 91–123. [Google Scholar] [PubMed]
- Sung, N.Y.; Kim, M.-Y.; Cho, J.Y. Scutellarein reduces inflammatory responses by inhibiting Src kinase activity. Korean J. Physiol. Pharmacol. 2015, 19, 441–449. [Google Scholar] [CrossRef] [PubMed]
- Wan, D.; Jiao, L.; Yang, H.; Liu, S. Structural characterization and immunological activities of the water-soluble oligosaccharides isolated from the Panax ginseng roots. Planta 2012, 235, 1289–1297. [Google Scholar] [CrossRef] [PubMed]
- Mócsai, A.; Ruland, J.; Tybulewicz, V.L. The SYK tyrosine kinase: A crucial player in diverse biological functions. Nat. Rev. Immunol. 2010, 10, 387–402. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.; Hsu, J.; Kim, Y.; Hu, D.-Q.; Xu, D.; Zhang, J.; Pashine, A.; Menke, J.; Whittard, T.; Romero, N. Selective inhibition of spleen tyrosine kinase (SYK) with a novel orally bioavailable small molecule inhibitor, RO9021, impinges on various innate and adaptive immune responses: Implications for SYK inhibitors in autoimmune disease therapy. Arthritis Res. Ther. 2013, 15, R146. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.; Ahn, H.M.; Shen, T.; Yoon, K.; Jang, H.-J.; Lee, Y.J.; Yang, H.M.; Kim, J.H.; Kim, C.; Han, M.H. Anti-inflammatory activity of ethanol extract derived from Phaseolus angularis beans. J. Ethnopharmacol. 2011, 137, 1197–1206. [Google Scholar] [CrossRef] [PubMed]
- Byeon, S.E.; Yu, T.; Yang, Y.; Lee, Y.G.; Kim, J.H.; Oh, J.; Jeong, H.Y.; Hong, S.; Yoo, B.C.; Cho, W.-J. Hydroquinone regulates hemeoxygenase-1 expression via modulation of Src kinase activity through thiolation of cysteine residues. Free Radic. Biol. Med. 2013, 57, 105–118. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Lee, Y.G.; Yoo, S.; Oh, J.; Jeong, D.; Song, W.K.; Yoo, B.C.; Rhee, M.H.; Park, J.; Cha, S.-H. Involvement of Src and the actin cytoskeleton in the antitumorigenic action of adenosine dialdehyde. Biochem. Pharmacol. 2013, 85, 1042–1056. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.G.; Chain, B.M.; Cho, J.Y. Distinct role of spleen tyrosine kinase in the early phosphorylation of inhibitor of κBα via activation of the phosphoinositide-3-kinase and Akt pathways. Int. J. Biochem. Cell Biol. 2009, 41, 811–821. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.Y.; Baik, K.U.; Jung, J.H.; Park, M.H. In vitro anti-inflammatory effects of cynaropicrin, a sesquiterpene lactone, from Saussurea lappa. Eur. J. Pharmacol. 2000, 398, 399–407. [Google Scholar] [CrossRef]
- Green, L.C.; Wagner, D.A.; Glogowski, J.; Skipper, P.L.; Wishnok, J.S.; Tannenbaum, S.R. Analysis of nitrate, nitrite, and [15N] nitrate in biological fluids. Anal. Biochem. 1982, 126, 131–138. [Google Scholar] [CrossRef]
- Gerlier, D.; Thomasset, N. Use of MTT colorimetric assay to measure cell activation. J. Immunol. Methods 1986, 94, 57–63. [Google Scholar] [CrossRef]
- Baek, K.S.; Hong, Y.D.; Kim, Y.; Sung, N.Y.; Yang, S.; Lee, K.M.; Park, J.Y.; Park, J.S.; Rho, H.S.; Shin, S.S.; et al. Anti-inflammatory activity of AP-SF, a ginsenoside-enriched fraction, from Korean ginseng. J. Ginseng Res. 2015, 39, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Lee, J.; Rhee, M.H.; Yu, T.; Baek, K.S.; Sung, N.Y.; Kim, Y.; Yoon, K.; Kim, J.H.; Kwak, Y.S.; et al. Molecular mechanism of protopanaxadiol saponin fraction-mediated anti-inflammatory actions. J. Ginseng Res. 2015, 39, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.Y.; Yoo, B.C.; Cho, J.Y. Ginsenoside-Rp1-induced apolipoprotein A-1 expression in the LoVo human colon cancer cell line. J. Ginseng Res. 2014, 38, 251–255. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.; Lee, J.; Lee, Y.G.; Byeon, S.E.; Kim, M.H.; Sohn, E.-H.; Lee, Y.J.; Lee, S.G.; Cho, J.Y. In vitro and in vivo anti-inflammatory effects of ethanol extract from Acer tegmentosum. J. Ethnopharmacol. 2010, 128, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Seo, H.S.; Sikder, M.A.; Lee, H.J.; Ryu, J.; Lee, C.J. Apigenin inhibits tumor necrosis factor-α-induced production and gene expression of mucin through regulating nuclear factor-κB signaling pathway in airway epithelial cells. Biomol. Ther. 2014, 22, 525–531. [Google Scholar] [CrossRef] [PubMed]
- Shen, T.; Lee, J.; Park, M.H.; Lee, Y.G.; Rho, H.S.; Kwak, Y.-S.; Rhee, M.H.; Park, Y.C.; Cho, J.Y. Ginsenoside Rp1, a ginsenoside derivative, blocks promoter activation of iNOS and COX-2 genes by suppression of an IKKβ-mediated NF-κB pathway in HEK293 cells. J. Ginseng Res. 2011, 35, 200. [Google Scholar] [CrossRef] [PubMed]
- Song, S.B.; Tung, N.H.; Quang, T.H.; Ngan, N.T.T.; Kim, K.E.; Kim, Y.H. Inhibition of TNF-α-mediated NF-κB transcriptional activity in HepG2 cells by dammarane-type saponins from Panax ginseng leaves. J. Ginseng Res. 2012, 36, 146. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-O.; Jeong, D.; Kim, M.-Y.; Cho, J.Y. ATP-binding pocket-targeted suppression of Src and Syk by luteolin contributes to its anti-inflammatory action. Mediat. Inflamm. 2015, 501, 967053. [Google Scholar] [CrossRef] [PubMed]
- Jeong, D.; Yi, Y.-S.; Sung, G.-H.; Yang, W.S.; Park, J.G.; Yoon, K.; Yoon, D.H.; Song, C.; Lee, Y.; Rhee, M.H. Anti-inflammatory activities and mechanisms of Artemisia asiatica ethanol extract. J. Ethnopharmacol. 2014, 152, 487–496. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the compound, 1-(2,3-dibenzimidazol-2-ylpropyl)-2-methoxybenzene, are available from the authors.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name (Real-time PCR) | Sequence (5′ to 3′) | |
|---|---|---|
| iNOS | F | GGAGCCTTTAGACCTCAACAGA |
| R | TGAACGAGGAGGGTGGTG | |
| TNF-α | F | TGCCTATGTCTCAGCCTCTTC |
| R | GAGGCCATTTGGGAACTTCT | |
| COX-2 | F | GGGAGTCTGGAACATTGTGAA |
| R | GCACATTGTAAGTAGGTGGACTGT | |
| IFN-β | F | TCCAAGAAAGGACGAACATTCG |
| R | GAGGCCATTTGGGAACTTCT | |
| GAPDH | F | CAATGAATACGGCTACAGCAAC |
| R | AGGGAGATGCTCAGTGTTGG |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, E.; Son, Y.-J.; Yang, Y.; Shen, T.; Kim, I.; Aravinthan, A.; Kim, J.-H.; Cho, J.Y. 1-(2,3-Dibenzimidazol-2-ylpropyl)-2-methoxybenzene Is a Syk Inhibitor with Anti-Inflammatory Properties. Molecules 2016, 21, 508. https://doi.org/10.3390/molecules21040508
Kim E, Son Y-J, Yang Y, Shen T, Kim I, Aravinthan A, Kim J-H, Cho JY. 1-(2,3-Dibenzimidazol-2-ylpropyl)-2-methoxybenzene Is a Syk Inhibitor with Anti-Inflammatory Properties. Molecules. 2016; 21(4):508. https://doi.org/10.3390/molecules21040508
Chicago/Turabian StyleKim, Eunji, Young-Jin Son, Yanyan Yang, Ting Shen, Ikyon Kim, Adithan Aravinthan, Jong-Hoon Kim, and Jae Youl Cho. 2016. "1-(2,3-Dibenzimidazol-2-ylpropyl)-2-methoxybenzene Is a Syk Inhibitor with Anti-Inflammatory Properties" Molecules 21, no. 4: 508. https://doi.org/10.3390/molecules21040508
APA StyleKim, E., Son, Y.-J., Yang, Y., Shen, T., Kim, I., Aravinthan, A., Kim, J.-H., & Cho, J. Y. (2016). 1-(2,3-Dibenzimidazol-2-ylpropyl)-2-methoxybenzene Is a Syk Inhibitor with Anti-Inflammatory Properties. Molecules, 21(4), 508. https://doi.org/10.3390/molecules21040508