Omega-3 Fatty Acids Protect Renal Functions by Increasing Docosahexaenoic Acid-Derived Metabolite Levels in SHR.Cg-Leprcp/NDmcr Rats, a Metabolic Syndrome Model

Abstract
:1. Introduction
2. Results and Discussion
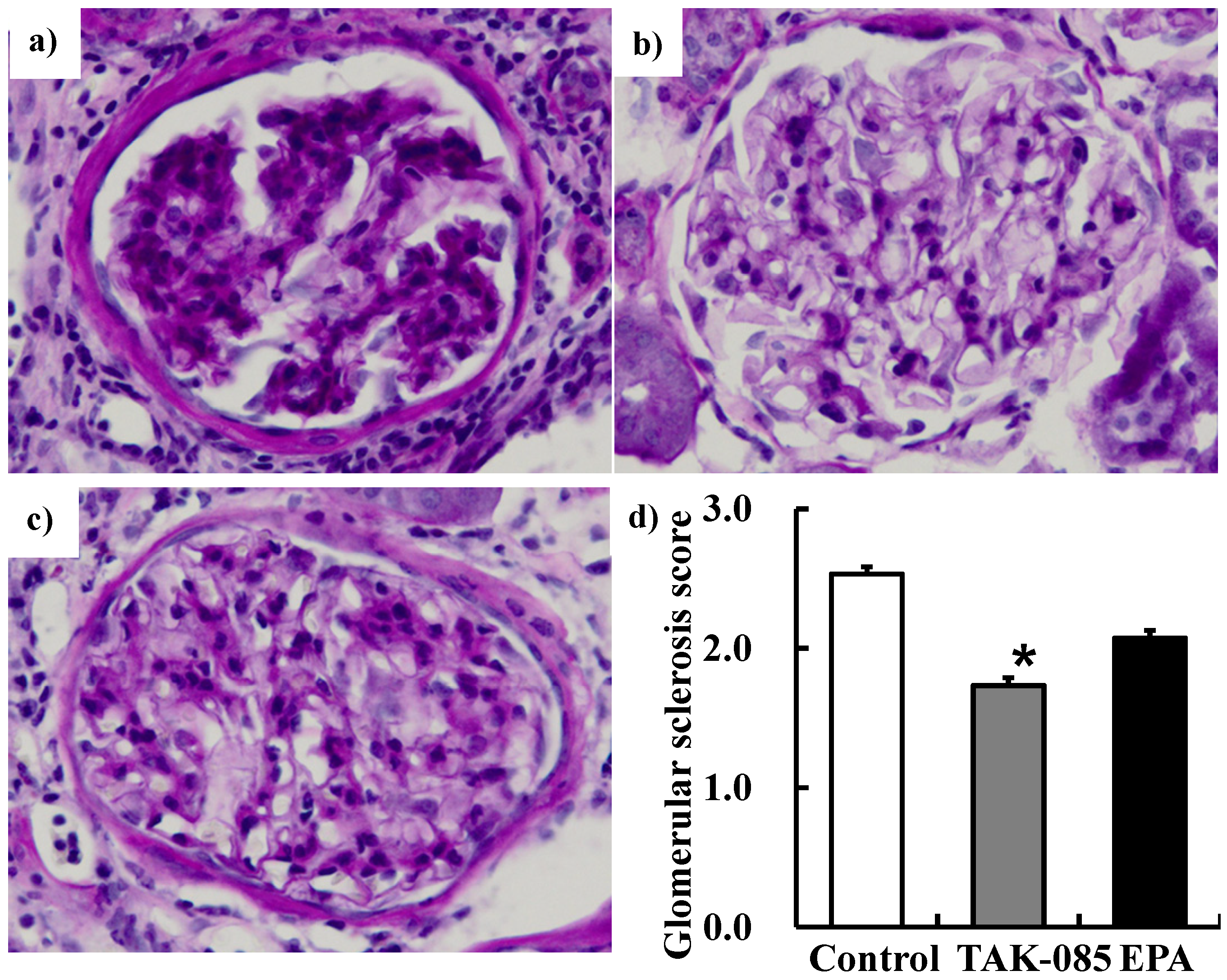
2.1. Effects of ω-3 PUFAs on Renal Function in the SHRcp Rats
2.1.1. Body Weight and Blood Pressure

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Control | TAK-085 | EPA |
|---|---|---|---|
| Body weight (g) | 591.5 ± 6.1 | 614.0 ± 6.9 | 595.6 ± 6.6 |
| SBP (mmHg) | 199.5 ± 4.5 | 192.9 ± 4.2 | 200.5 ± 3.6 |
| MBP (mmHg) | 165.5 ± 3.7 | 160.3 ± 3.1 | 163.3 ± 2.4 |
| DBP (mmHg) | 153.5 ± 3.8 | 145.7 ± 4.2 | 148.3 ± 3.4 |
| Creatinine (mg/dL) | 0.27 ± 0.01 | 0.30 ± 0.01 | 0.30 ± 0.01 |
| BUN (mg/dL) | 30.3 ± 0.9 | 31.5 ± 0.8 | 34.9 ± 1.3 |
| Triglyceride (mg/dL) | 266.9 ± 32.3 | 253.8 ± 13.1 | 341.3 ± 33.7 |
| Total cholesterol (mg/dL) | 368.3 ± 26.3 | 379.5 ± 10.4 | 422.6 ± 19.8 |
| HDL-cholesterol (mg/dL) | 51.3 ± 1.1 | 52.1 ± 1.2 | 55.5 ± 0.8 |
2.1.2. Renal Function Parameters
2.2. Effects of ω-3 PUFAs on Lipid Levels in SHRcp Rats
2.2.1. Plasma, Kidney, and Liver Fatty Acid Profiles
| Control | TAK-085 | EPA | |
|---|---|---|---|
| Water intake (mL/kg bw. day) | 64.9 ± 4.3 | 56.1 ± 3.3 | 61.2 ± 1.9 |
| Urine flow (mL/kg bw. day) | 36.8 ± 1.5 | 32.1 ± 2.4 | 37.5 ± 1.7 |
| Urinary albumin (mg/kg bw. day) | 163.6 ± 11.7 | 134.1 ± 12.1 | 172.5 ± 12.6 |
| Urinary creatinine (mg/kg bw. day) | 6.9 ± 0.7 | 8.9 ± 0.6 | 7.3 ± 0.6 |
| AC ratio | 25.7 ± 2.6 | 16.3 ± 2.4 * | 28.5 ± 6.0 |
| Creatinine clearance (mL/min) | 0.88 ± 0.08 | 1.27 ± 0.07 * | 0.96 ± 0.11 |

| (mol%) | Control | TAK-085 | EPA |
|---|---|---|---|
| PLA (16:0) | 26.03 ± 0.16 | 25.73 ± 0.21 | 25.43 ± 0.12 * |
| STA (18:0) | 6.63 ± 0.10 | 6.23 ± 0.06 | 6.32 ± 0.09 |
| OA (18:1n-9) | 25.20 ± 0.40 | 23.33 ± 0.17 * | 23.47 ± 0.27 * |
| LA (18:2n-6) | 18.45 ± 0.17 | 20.99 ± 0.24 * | 19.77 ± 0.13 * |
| ALA (18:3n-3) | 0.50 ± 0.02 | 0.58 ± 0.02 * | 0.59 ± 0.02 * |
| ARA (20:4n-6) | 19.48 ± 0.48 | 11.14 ± 0.30 * | 11.83 ± 0.31 * |
| EPA (20:5n-3) | 0.69 ± 0.04 | 4.11 ± 0.18 * | 6.89 ± 0.15 * |
| DPA (22:5n-3) | 1.00 ± 0.02 | 1.51 ± 0.02 * | 3.24 ± 0.05 * |
| DHA (22:6n-3) | 1.36 ± 0.08 | 5.73 ± 0.11 * | 1.81 ± 0.03 * |
| n-6/n-3 | 10.80 ± 0.39 | 2.71 ± 0.07 * | 2.53 ± 0.05 * |
| DHA/ARA | 0.07 ± 0.01 | 0.52 ± 0.02 * | 0.15 ± 0.01 * |
| EPA/ARA | 0.04 ± 0.00 | 0.37 ± 0.02 * | 0.59 ± 0.03 * |
| SCD index | 3.82 ± 0.11 | 3.75 ± 0.05 | 3.72 ± 0.08 |
| (mol%) | Control | TAK-085 | EPA |
|---|---|---|---|
| PLA (16:0) | 28.48 ± 0.14 | 27.39 ± 0.16 * | 27.55 ± 0.17 * |
| STA (18:0) | 14.70 ± 0.31 | 14.48 ± 0.28 | 14.10 ± 0.37 |
| OA (18:1n-9) | 16.59 ± 0.93 | 14.38 ± 0.83 | 16.36 ± 1.05 |
| LA (18:2n-6) | 12.17 ± 0.18 | 15.26 ± 0.15 * | 14.38 ± 0.15 * |
| ALA (18:3n-3) | 0.26 ± 0.01 | 0.24 ± 0.01 | 0.26 ± 0.01 |
| ARA (20:4n-6) | 22.24 ± 0.61 | 17.37 ± 0.46 * | 16.87 ± 0.55 * |
| EPA (20:5n-3) | 0.24 ± 0.02 | 2.00 ± 0.05 * | 2.92 ± 0.11 * |
| DPA (22:5n-3) | 0.74 ± 0.02 | 1.34 ± 0.04 * | 2.70 ± 0.07 * |
| DHA (22:6n-3) | 1.45 ± 0.08 | 4.15 ± 0.10 * | 1.65 ± 0.06 |
| n-6/n-3 | 12.92 ± 0.36 | 4.25 ± 0.11 * | 4.16 ± 0.07 * |
| DHA/ARA | 0.07 ± 0.01 | 0.24 ± 0.01 * | 0.10 ± 0.01 * |
| EPA/ARA | 0.01 ± 0.01 | 0.12 ± 0.01 * | 0.17 ± 0.02 * |
| SCD index | 1.15 ± 0.09 | 1.01 ± 0.08 | 1.19 ± 0.11 |
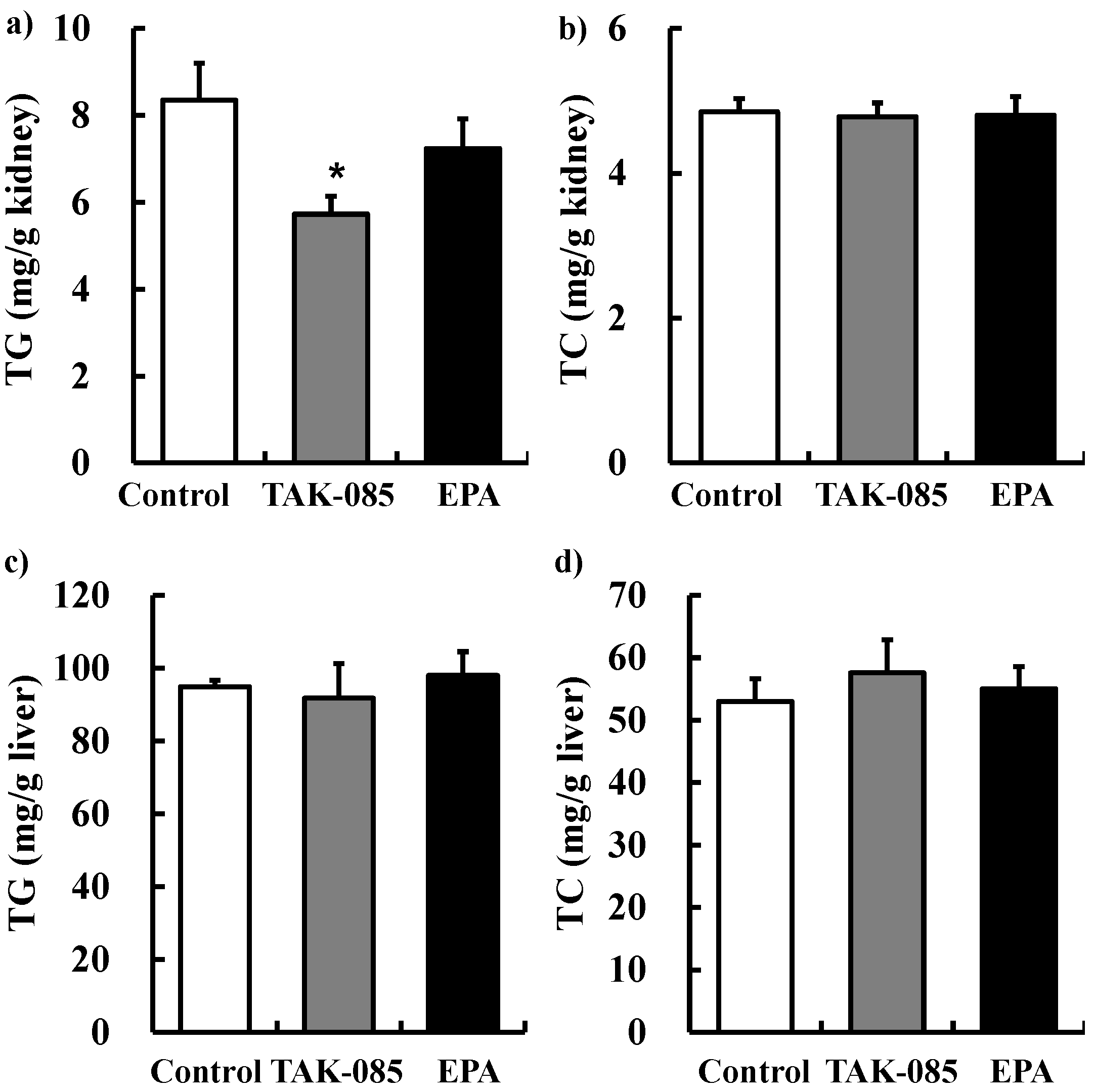
2.2.2. Kidney and Liver TG and TC Levels

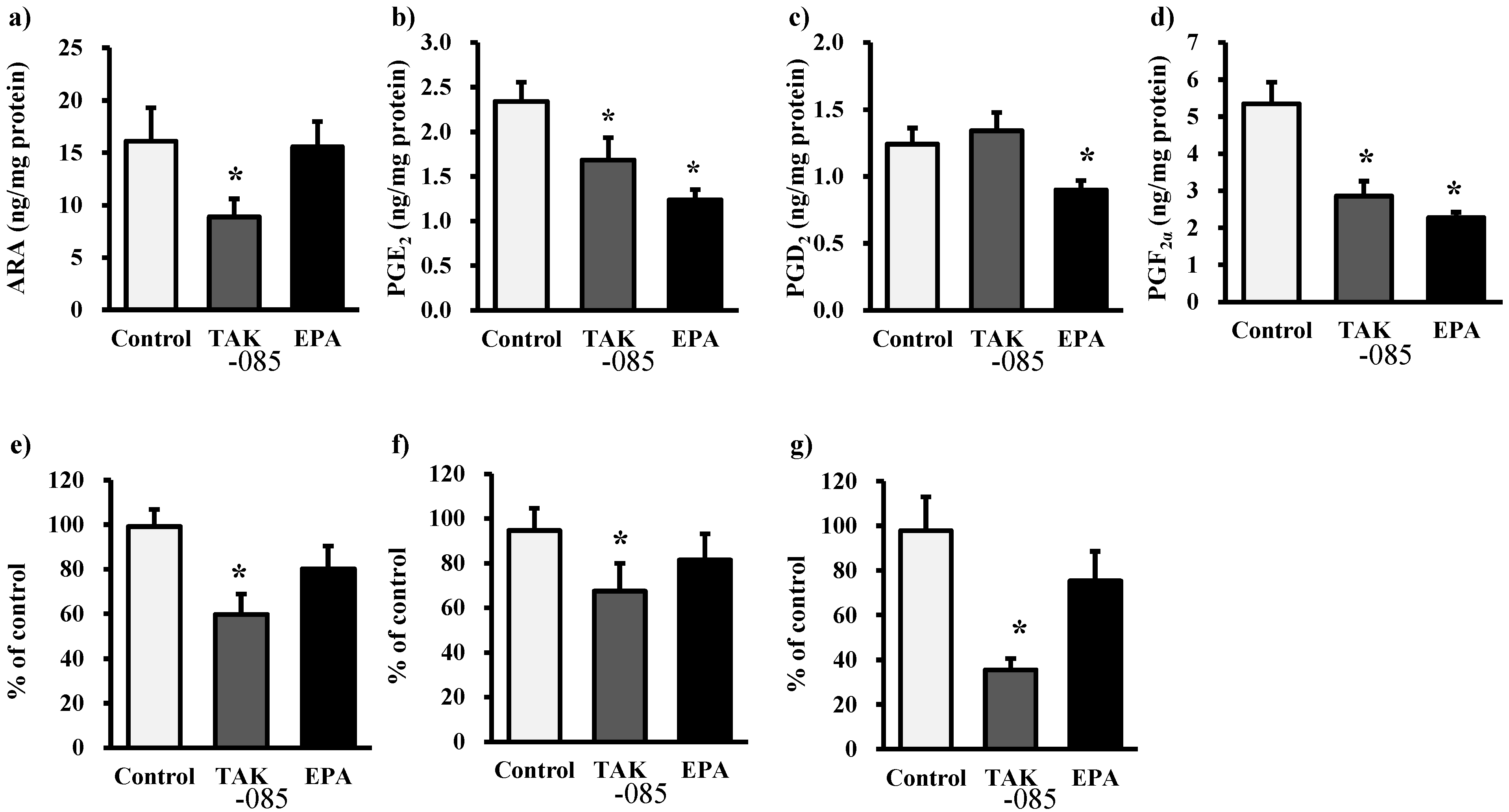
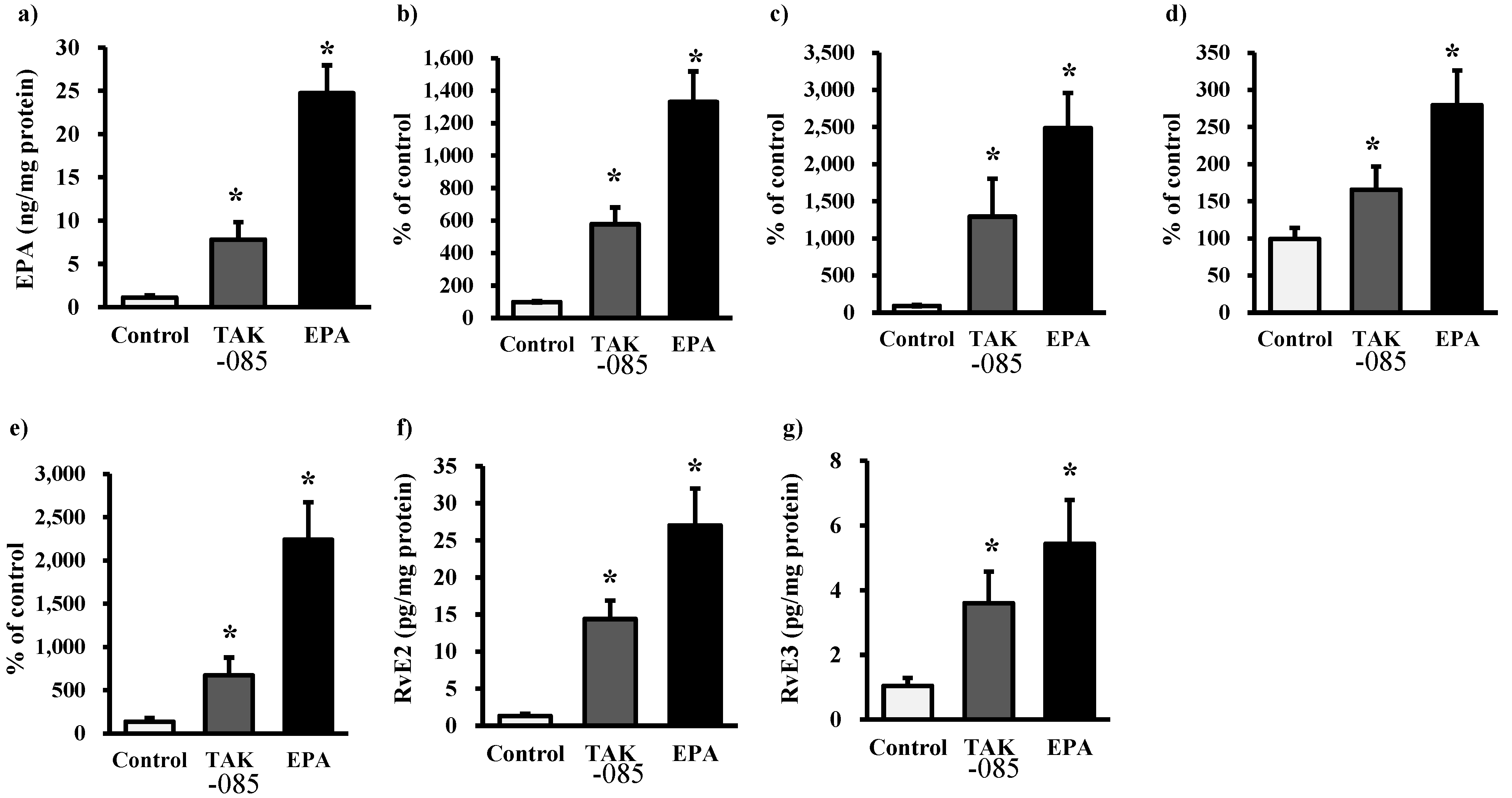
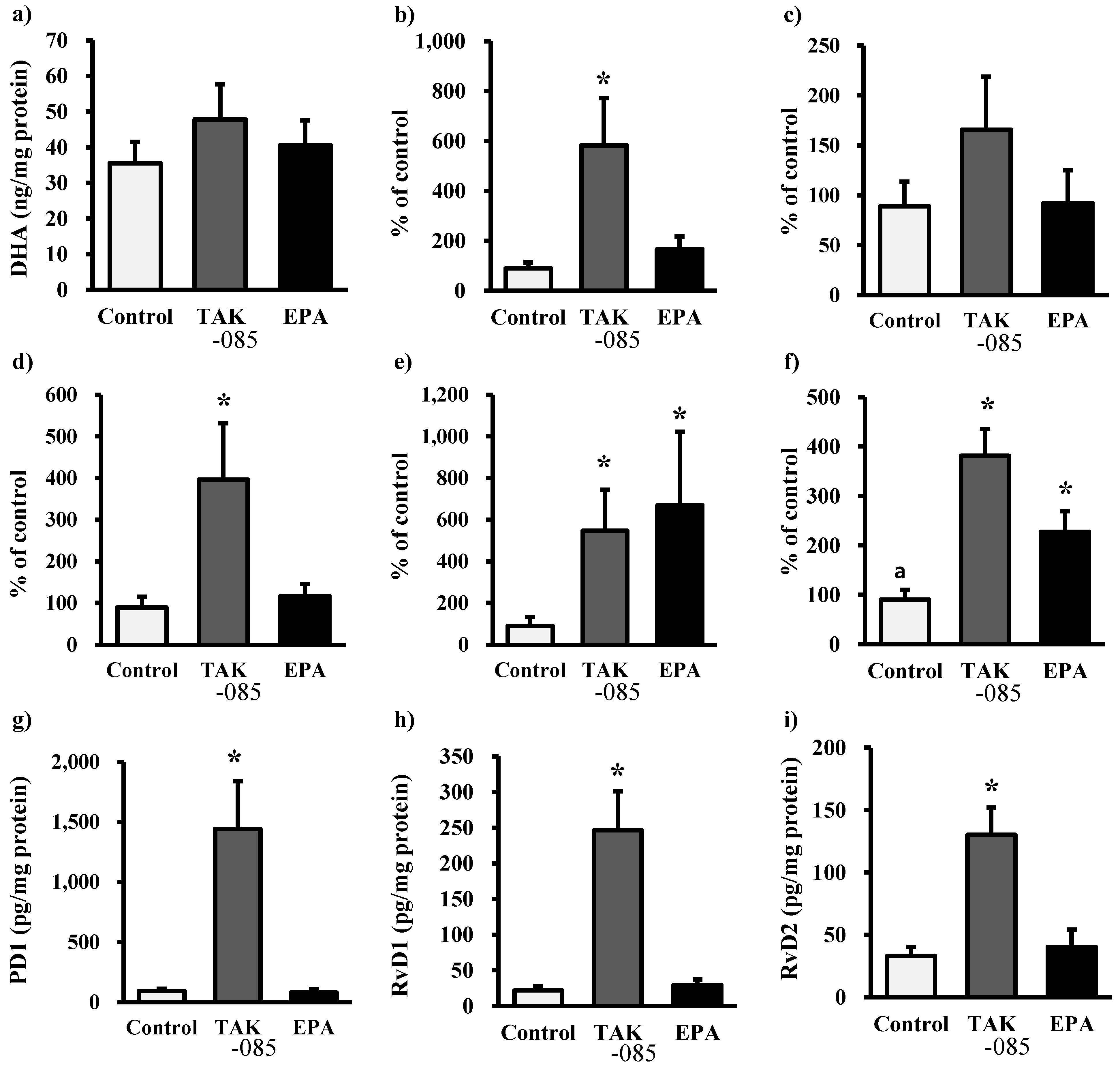
2.3. Levels of Eicosanoids and Docosanoids in the Kidney
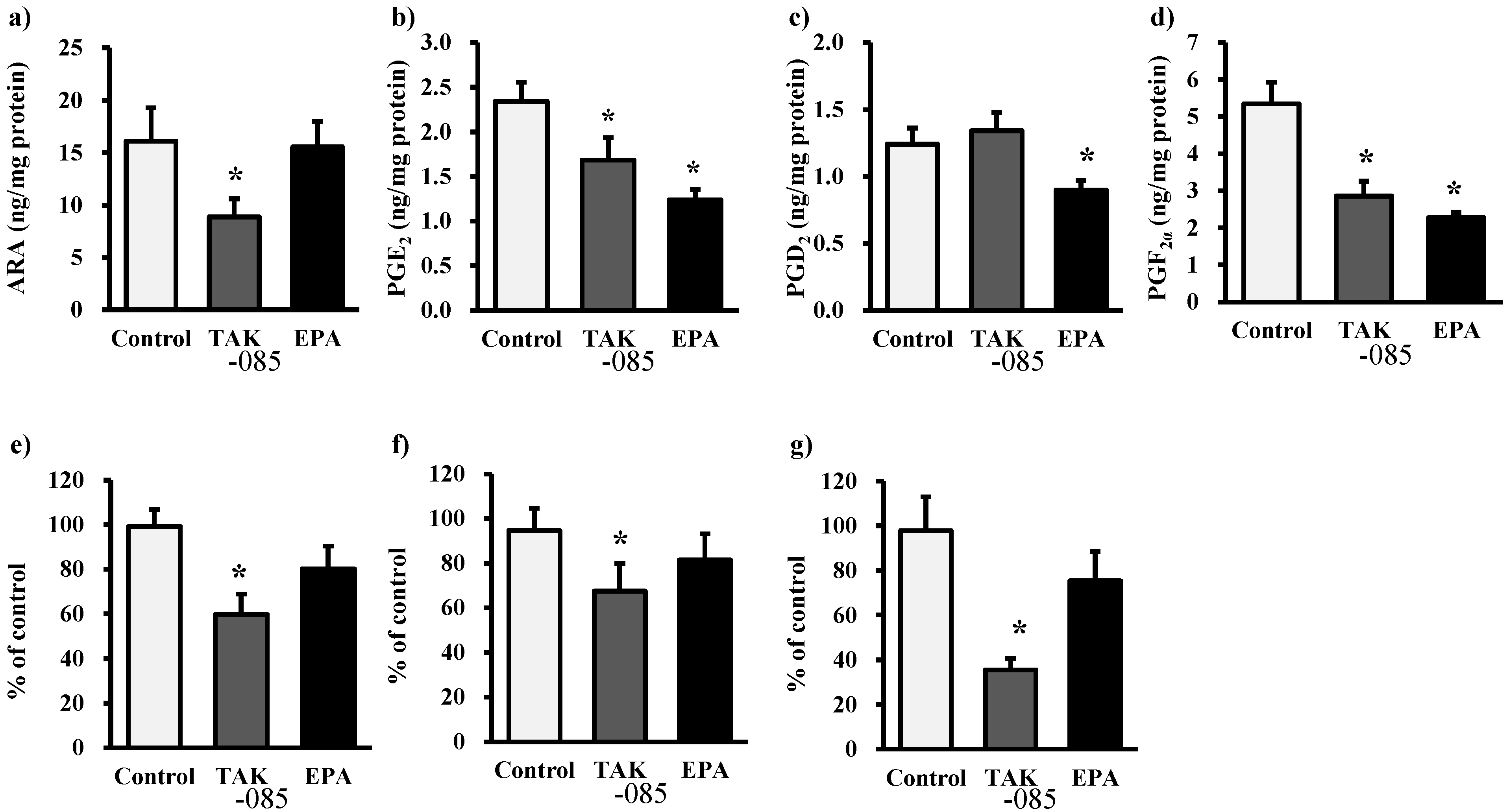
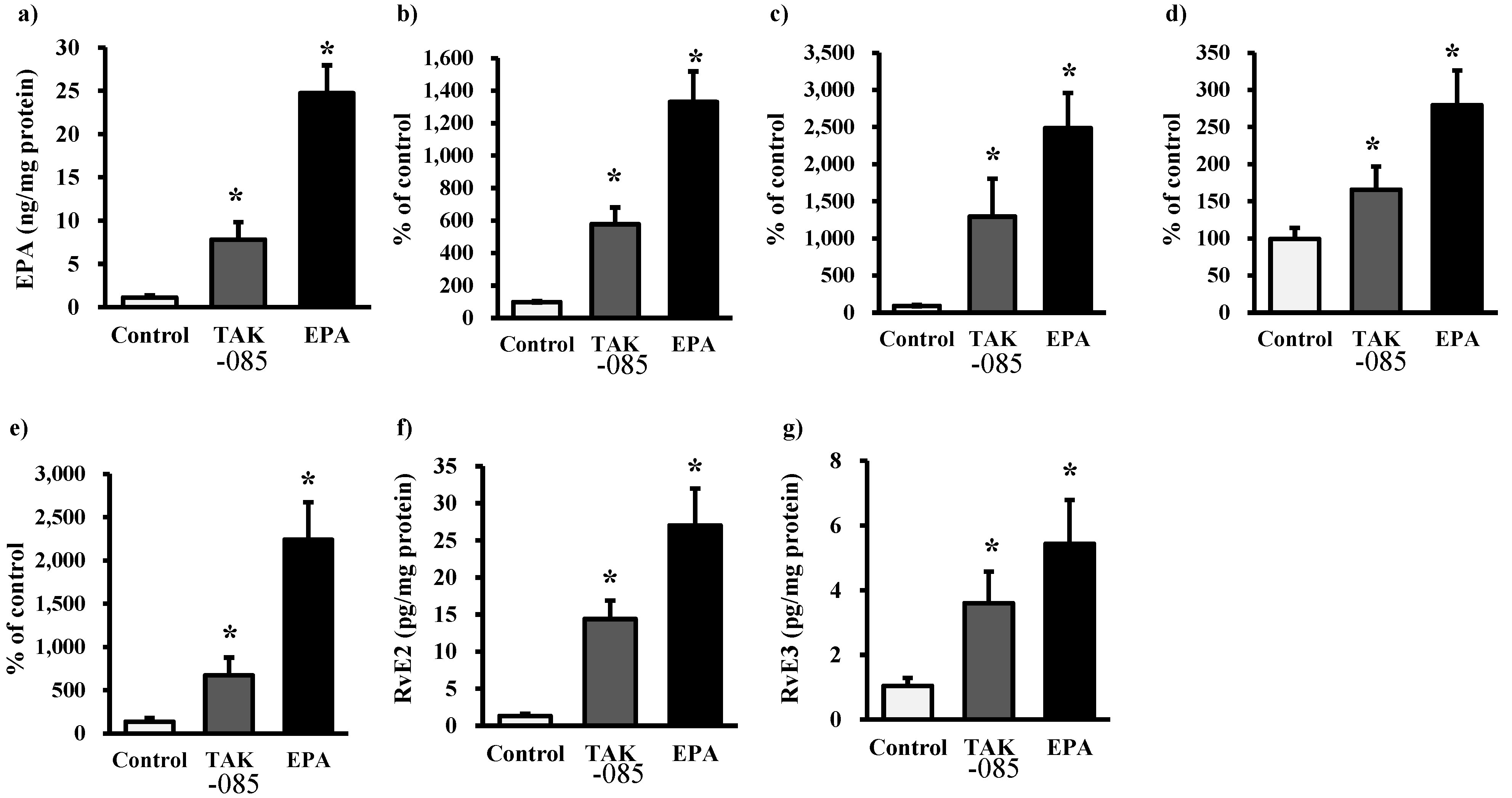
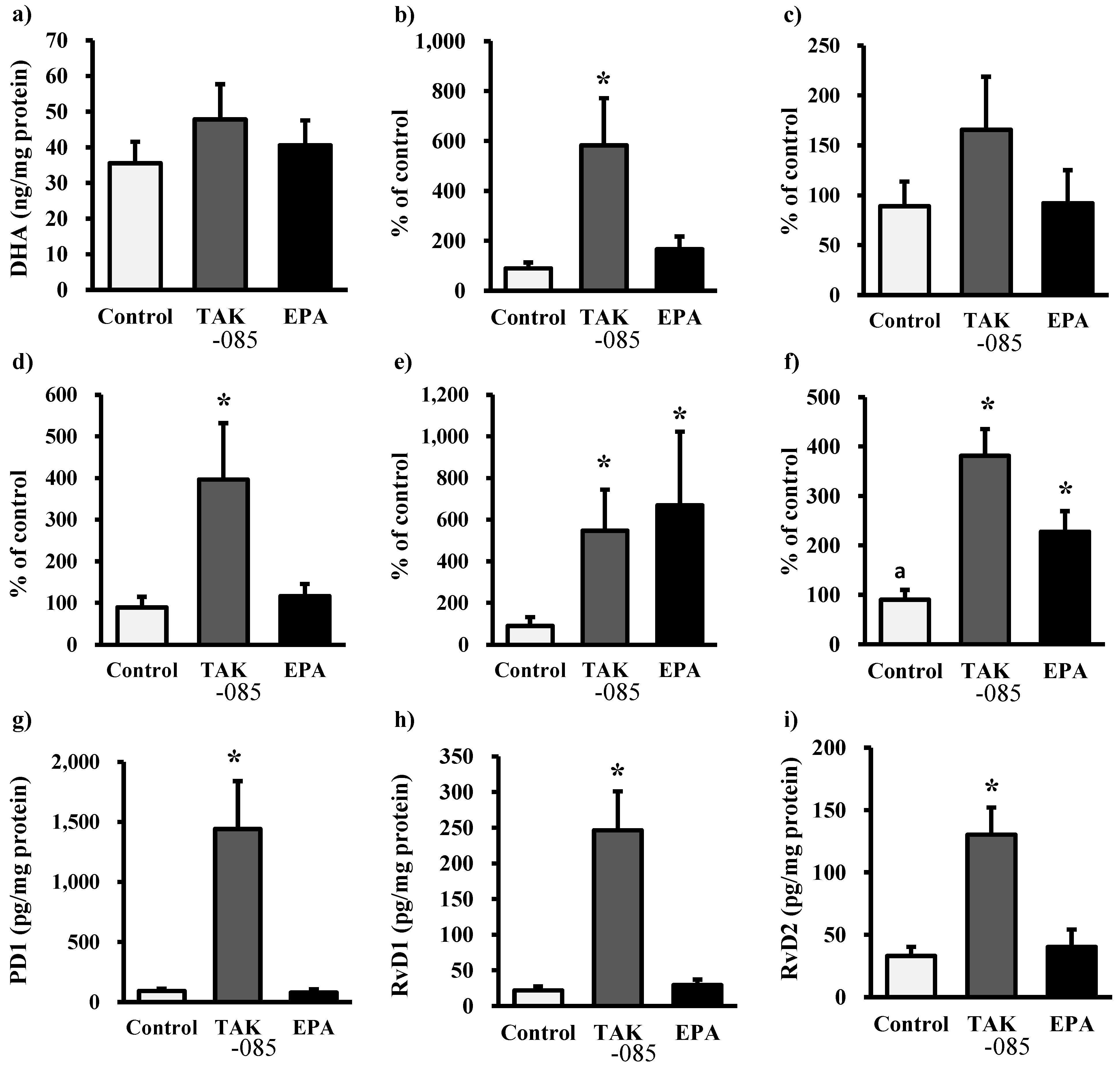
2.4. Discussion
3. Experimental
3.1. Animals
3.2. Measurement of Blood Pressure
3.3. Blood Sample Assays
3.4. Analysis of Fatty Acid profiles
3.5. TG and Total Cholesterol (TC) in the Kidney and Liver
3.6. ROS and Lipid Peroxidation (LPO) Measurement
3.7. Creatinine Clearance Measurement
3.8. Morphological Analysis
3.9. Analysis of Fatty Acid Metabolites
3.9.1. Sample Preparation
3.9.2. LC-ESI-MS–MS-Based Analysis
3.10. Statistical Analysis
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflictts of Interest
References
- Kielar, M.L.; Jeyarajah, D.R.; Zhou, X.J.; Lu, C.Y. Docosahexaenoic Acid Ameliorates Murine Ischemic Acute Renal Failure and Prevents Increases in mRNA Abundance for both TNF-Alpha and Inducible Nitric Oxide Synthase. J. Am. Soc. Nephrol. 2003, 14, 389–396. [Google Scholar] [CrossRef]
- Jia, Q.; Zhou, H.R.; Bennink, M.; Pestka, J.J. Docosahexaenoic Acid Attenuates Mycotoxin-Induced Immunoglobulin a Nephropathy, Interleukin-6 Transcription, and Mitogen-Activated Protein Kinase Phosphorylation in Mice. J. Nutr. 2004, 134, 3343–3349. [Google Scholar]
- Jia, Q.; Shi, Y.; Bennink, M.B.; Pestka, J.J. Docosahexaenoic Acid and Eicosapentaenoic Acid, but Not Alpha-Linolenic Acid, Suppress Deoxynivalenol-Induced Experimental IgA Nephropathy in Mice. J. Nutr. 2004, 134, 1353–1361. [Google Scholar]
- Parinyasiri, U.; Ong-Ajyooth, L.; Parichatikanond, P.; Ong-Ajyooth, S.; Liammongkolkul, S.; Kanyog, S. Effect of Fish Oil on Oxidative Stress, Lipid Profile and Renal Function in IgA Nephropathy. J. Med. Assoc. Thai. 2004, 87, 143–149. [Google Scholar]
- Mariee, A.D.; Abd-Ellah, M.F. Protective Effect of Docosahexaenoic Acid Against Cyclosporine A-Induced Nephrotoxicity in Rats: A Possible Mechanism of Action. Ren. Fail. 2011, 33, 66–71. [Google Scholar] [CrossRef]
- Thakkar, R.R.; Wang, O.L.; Zerouga, M.; Stillwell, W.; Haq, A.; Kissling, R.; Pierce, W.M.; Smith, N.B.; Miller, F.N.; Ehringer, W.D. Docosahexaenoic Acid Reverses Cyclosporin A-Induced Changes in Membrane Structure and Function. Biochim. Biophys. Acta 2000, 1474, 183–195. [Google Scholar]
- Taneda, S.; Honda, K.; Tomidokoro, K.; Uto, K.; Nitta, K.; Oda, H. Eicosapentaenoic Acid Restores Diabetic Tubular Injury through Regulating Oxidative Stress and Mitochondrial Apoptosis. Am. J. Physiol. Renal Physiol. 2010, 299, F1451–F1461. [Google Scholar] [CrossRef]
- Chin, H.J.; Fu, Y.Y.; Ahn, J.M.; Na, K.Y.; Kim, Y.S.; Kim, S.; Chae, D.W. Omacor, N-3 Polyunsaturated Fatty Acid, Attenuated Albuminuria and Renal Dysfunction with Decrease of SREBP-1 Expression and Triglyceride Amount in the Kidney of Type II Diabetic Animals. Nephrol. Dial. Transplant. 2010, 25, 1450–1457. [Google Scholar] [CrossRef]
- Bays, H. Clinical Overview of Omacor: A Concentrated Formulation of Omega-3 Polyunsaturated Fatty Acids. Am. J. Cardiol. 2006, 98, 71i–76i. [Google Scholar] [CrossRef]
- Zhang, M.; Hagiwara, S.; Matsumoto, M.; Gu, L.; Tanimoto, M.; Nakamura, S.; Kaneko, S.; Gohda, T.; Qian, J.; Horikoshi, S.; et al. Effects of Eicosapentaenoic Acid on the Early Stage of Type 2 Diabetic Nephropathy in KKA(Y)/Ta Mice: Involvement of Anti-Inflammation and Antioxidative Stress. Metabolism 2006, 55, 1590–1598. [Google Scholar] [CrossRef]
- Garman, J.H.; Mulroney, S.; Manigrasso, M.; Flynn, E.; Maric, C. Omega-3 Fatty Acid Rich Diet Prevents Diabetic Renal Disease. Am. J. Physiol. Renal Physiol. 2009, 296, F306–F316. [Google Scholar]
- Huang, S.; Konieczkowski, M.; Schelling, J.R.; Sedor, J.R. Interleukin-1 Stimulates Jun N-Terminal/Stress-Activated Protein Kinase by an Arachidonate-Dependent Mechanism in Mesangial Cells. Kidney Int. 1999, 55, 1740–1749. [Google Scholar] [CrossRef]
- Kiritoshi, S.; Nishikawa, T.; Sonoda, K.; Kukidome, D.; Senokuchi, T.; Matsuo, T.; Matsumura, T.; Tokunaga, H.; Brownlee, M.; Araki, E. Reactive Oxygen Species from Mitochondria Induce Cyclooxygenase-2 Gene Expression in Human Mesangial Cells: Potential Role in Diabetic Nephropathy. Diabetes 2003, 52, 2570–2577. [Google Scholar] [CrossRef]
- Goto, S.; Nakamura, H.; Morooka, H.; Terao, Y.; Shibata, O.; Sumikawa, K. Role of Reactive Oxygen in Phospholipase A2 Activation by Ischemia/Reperfusion of the Rat Kidney. J. Anesth. 1999, 13, 90–93. [Google Scholar] [CrossRef]
- Ferreri, N.R.; An, S.J.; McGiff, J.C. Cyclooxygenase-2 Expression and Function in the Medullary Thick Ascending Limb. Am. J. Physiol. 1999, 277, F360–F368. [Google Scholar]
- Fischer, D.B.; Christman, J.W.; Badr, K.F. Fifteen-S-Hydroxyeicosatetraenoic Acid (15-S-HETE) Specifically Antagonizes the Chemotactic Action and Glomerular Synthesis of Leukotriene B4 in the Rat. Kidney Int. 1992, 41, 1155–1160. [Google Scholar] [CrossRef]
- Hong, S.; Lu, Y. Omega-3 Fatty Acid-Derived Resolvins and Protectins in Inflammation Resolution and Leukocyte Functions: Targeting Novel Lipid Mediator Pathways in Mitigation of Acute Kidney Injury. Front. Immunol. 2013, 4, 13. [Google Scholar]
- Yamaguchi, Y.; Yoshikawa, N.; Kagota, S.; Nakamura, K.; Haginaka, J.; Kunitomo, M. Elevated Circulating Levels of Markers of Oxidative-Nitrative Stress and Inflammation in a Genetic Rat Model of Metabolic Syndrome. Nitric Oxide 2006, 15, 380–386. [Google Scholar] [CrossRef]
- Andrade, L.; Campos, S.B.; Seguro, A.C. Hypercholesterolemia Aggravates Radiocontrast Nephrotoxicity: Protective Role of L-Arginine. Kidney Int. 1998, 53, 1736–1742. [Google Scholar] [CrossRef]
- Van Buren, P.N.; Toto, R.D. The Pathogenesis and Management of Hypertension in Diabetic Kidney Disease. Med. Clin. North Am. 2013, 97, 31–51. [Google Scholar] [CrossRef]
- Kroger, J.; Schulze, M.B. Recent Insights into the Relation of Delta5 Desaturase and Delta6 Desaturase Activity to the Development of Type 2 Diabetes. Curr. Opin. Lipidol. 2012, 23, 4–10. [Google Scholar] [CrossRef]
- Merino, D.M.; Ma, D.W.; Mutch, D.M. Genetic Variation in Lipid Desaturases and its Impact on the Development of Human Disease. Lipids Health. Dis. 2010, 9. [Google Scholar] [CrossRef]
- Gronn, M.; Christensen, E.; Hagve, T.A.; Christophersen, B.O. Effects of Dietary Purified Eicosapentaenoic Acid (20:5 (N-3)) and Docosahexaenoic Acid (22:6 (N-3)) on Fatty Acid Desaturation and Oxidation in Isolated Rat Liver Cells. Biochim. Biophys. Acta 1992, 1125, 35–43. [Google Scholar]
- Li, S.; Nagothu, K.; Ranganathan, G.; Ali, S.M.; Shank, B.; Gokden, N.; Ayyadevara, S.; Megyesi, J.; Olivecrona, G.; Chugh, S.S.; et al. Reduced Kidney Lipoprotein Lipase and Renal Tubule Triglyceride Accumulation in Cisplatin-Mediated Acute Kidney Injury. Am. J. Physiol. Renal Physiol. 2012, 303, F437–F448. [Google Scholar] [CrossRef]
- Lee, H.S. Mechanisms and Consequences of Hypertriglyceridemia and Cellular Lipid Accumulation in Chronic Kidney Disease and Metabolic Syndrome. Histol. Histopathol. 2011, 26, 1599–1610. [Google Scholar]
- Bobulescu, I.A. Renal Lipid Metabolism and Lipotoxicity. Curr. Opin. Nephrol. Hypertens. 2010, 19, 393–402. [Google Scholar] [CrossRef]
- Ntambi, J.M. Regulation of Stearoyl-CoA Desaturase by Polyunsaturated Fatty Acids and Cholesterol. J. Lipid Res. 1999, 40, 1549–1558. [Google Scholar]
- Kim, H.J.; Vaziri, N.D. Sterol Regulatory Element-Binding Proteins, Liver X Receptor, ABCA1 Transporter, CD36, Scavenger Receptors A1 and B1 in Nephrotic Kidney. Am. J. Nephrol. 2009, 29, 607–614. [Google Scholar] [CrossRef]
- Tanaka, N.; Zhang, X.; Sugiyama, E.; Kono, H.; Horiuchi, A.; Nakajima, T.; Kanbe, H.; Tanaka, E.; Gonzalez, F.J.; Aoyama, T. Eicosapentaenoic Acid Improves Hepatic Steatosis Independent of PPARalpha Activation through Inhibition of SREBP-1 Maturation in Mice. Biochem. Pharmacol. 2010, 80, 1601–1612. [Google Scholar] [CrossRef]
- Zaima, N.; Sugawara, T.; Goto, D.; Hirata, T. Trans Geometric Isomers of EPA Decrease LXRalpha-Induced Cellular Triacylglycerol Via Suppression of SREBP-1c and PGC-1beta. J. Lipid Res. 2006, 47, 2712–2717. [Google Scholar] [CrossRef]
- Caputo, M.; Zirpoli, H.; Torino, G.; Tecce, M.F. Selective Regulation of UGT1A1 and SREBP-1c mRNA Expression by Docosahexaenoic, Eicosapentaenoic, and Arachidonic Acids. J. Cell. Physiol. 2011, 226, 187–193. [Google Scholar] [CrossRef]
- Fujimoto, Y.; Shimada, S.; Fujikawa, T.; Sakuma, S.; Fujita, T. Triacylglycerol Lipase Mediated Release of Arachidonic Acid for Prostaglandin Synthesis in Rabbit Kidney Medulla Microsomes. Prostaglandins Leukot. Essent. Fatty Acids 1991, 42, 251–256. [Google Scholar] [CrossRef]
- Qu, X.; Zhang, X.; Yao, J.; Song, J.; Nikolic-Paterson, D.J.; Li, J. Resolvins E1 and D1 Inhibit Interstitial Fibrosis in the Obstructed Kidney Via Inhibition of Local Fibroblast Proliferation. J. Pathol. 2012, 228, 506–519. [Google Scholar]
- Oh, S.F.; Dona, M.; Fredman, G.; Krishnamoorthy, S.; Irimia, D.; Serhan, C.N. Resolvin E2 Formation and Impact in Inflammation Resolution. J. Immunol. 2012, 188, 4527–4534. [Google Scholar] [CrossRef]
- Hassan, I.R.; Gronert, K. Acute Changes in Dietary Omega-3 and Omega-6 Polyunsaturated Fatty Acids have a Pronounced Impact on Survival Following Ischemic Renal Injury and Formation of Renoprotective Docosahexaenoic Acid-Derived Protectin D1. J. Immunol. 2009, 182, 3223–3232. [Google Scholar] [CrossRef]
- Duffield, J.S.; Hong, S.; Vaidya, V.S.; Lu, Y.; Fredman, G.; Serhan, C.N.; Bonventre, J.V. Resolvin D Series and Protectin D1 Mitigate Acute Kidney Injury. J. Immunol. 2006, 177, 5902–5911. [Google Scholar]
- Tian, H.; Lu, Y.; Shah, S.P.; Wang, Q.; Hong, S. 14S,21R-Dihydroxy-Docosahexaenoic Acid Treatment Enhances Mesenchymal Stem Cell Amelioration of Renal Ischemia/Reperfusion Injury. Stem Cells Dev. 2012, 21, 1187–1199. [Google Scholar] [CrossRef]
- Serhan, C.N.; Gotlinger, K.; Hong, S.; Arita, M. Resolvins, Docosatrienes, and Neuroprotectins, Novel Omega-3-Derived Mediators, and their Aspirin-Triggered Endogenous Epimers: An Overview of their Protective Roles in Catabasis. Prostaglandins Other Lipid Mediat. 2004, 73, 155–172. [Google Scholar] [CrossRef]
- Wanninger, J.; Bauer, S.; Eisinger, K.; Weiss, T.S.; Walter, R.; Hellerbrand, C.; Schaffler, A.; Higuchi, A.; Walsh, K.; Buechler, C. Adiponectin Upregulates Hepatocyte CMKLR1 which is Reduced in Human Fatty Liver. Mol. Cell. Endocrinol. 2012, 349, 248–254. [Google Scholar] [CrossRef]
- Hao, C.M.; Breyer, M.D. Physiologic and Pathophysiologic Roles of Lipid Mediators in the Kidney. Kidney Int. 2007, 71, 1105–1115. [Google Scholar] [CrossRef]
- Muirhead, E.E. Renal Vasodepressor Mechanisms: The Medullipin System. J. Hypertens. 1993, 11 (Suppl. 5), S53–S58. [Google Scholar]
- Badr, K.F. Lipoxygenases as Therapeutic Targets in the Acute and Subacute Phases of Glomerular Immune Injury. Contrib. Nephrol. 1996, 118, 113–125. [Google Scholar]
- Hashimoto, M.; Shinozuka, K.; Gamoh, S.; Tanabe, Y.; Hossain, M.S.; Kwon, Y.M.; Hata, N.; Misawa, Y.; Kunitomo, M.; Masumura, S. The Hypotensive Effect of Docosahexaenoic Acid is Associated with the Enhanced Release of ATP from the Caudal Artery of Aged Rats. J. Nutr. 1999, 129, 70–76. [Google Scholar]
- Bligh, E.G.; Dyer, W.J. A Rapid Method of Total Lipid Extraction and Purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Katakura, M.; Hashimoto, M.; Tanabe, Y.; Shido, O. Hydrogen-Rich Water Inhibits Glucose and Alpha,Beta -Dicarbonyl Compound-Induced Reactive Oxygen Species Production in the SHR.Cg-Leprcp/NDmcr Rat Kidney. Med. Gas. Res. 2012, 2. [Google Scholar] [CrossRef]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for Lipid Peroxides in Animal Tissues by Thiobarbituric Acid Reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein Measurement with the Folin Phenol Reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar]
- Uehara, Y.; Hirawa, N.; Kawabata, Y.; Suzuki, T.; Ohshima, N.; Oka, K.; Ikeda, T.; Goto, A.; Toyo-oka, T.; Kizuki, K. Long-Term Infusion of Kallikrein Attenuates Renal Injury in Dahl Salt-Sensitive Rats. Hypertension 1994, 24, 770–778. [Google Scholar] [CrossRef]
- Arita, M. Mediator Lipidomics in Acute Inflammation and Resolution. J. Biochem. 2012, 152, 313–319. [Google Scholar] [CrossRef]
- Isobe, Y.; Arita, M.; Matsueda, S.; Iwamoto, R.; Fujihara, T.; Nakanishi, H.; Taguchi, R.; Masuda, K.; Sasaki, K.; Urabe, D.; et al. Identification and Structure Determination of Novel Anti-Inflammatory Mediator Resolvin E3, 17,18-Dihydroxyeicosapentaenoic Acid. J. Biol. Chem. 2012, 287, 10525–10534. [Google Scholar] [CrossRef]
- Juman, S.; Hashimoto, M.; Katakura, M.; Inoue, T.; Tanabe, Y.; Arita, M.; Miki, T.; Shido, O. Effects of Long-Term Oral Administration of Arachidonic Acid and Docosahexaenoic Acid on the Immune Functions of Young Rats. Nutrients 2013, 5, 1949–1961. [Google Scholar] [CrossRef]
- Sample Availability: Contact the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Katakura, M.; Hashimoto, M.; Inoue, T.; Mamun, A.A.; Tanabe, Y.; Iwamoto, R.; Arita, M.; Tsuchikura, S.; Shido, O. Omega-3 Fatty Acids Protect Renal Functions by Increasing Docosahexaenoic Acid-Derived Metabolite Levels in SHR.Cg-Leprcp/NDmcr Rats, a Metabolic Syndrome Model. Molecules 2014, 19, 3247-3263. https://doi.org/10.3390/molecules19033247
Katakura M, Hashimoto M, Inoue T, Mamun AA, Tanabe Y, Iwamoto R, Arita M, Tsuchikura S, Shido O. Omega-3 Fatty Acids Protect Renal Functions by Increasing Docosahexaenoic Acid-Derived Metabolite Levels in SHR.Cg-Leprcp/NDmcr Rats, a Metabolic Syndrome Model. Molecules. 2014; 19(3):3247-3263. https://doi.org/10.3390/molecules19033247
Chicago/Turabian StyleKatakura, Masanori, Michio Hashimoto, Takayuki Inoue, Abdullah Al Mamun, Yoko Tanabe, Ryo Iwamoto, Makoto Arita, Satoru Tsuchikura, and Osamu Shido. 2014. "Omega-3 Fatty Acids Protect Renal Functions by Increasing Docosahexaenoic Acid-Derived Metabolite Levels in SHR.Cg-Leprcp/NDmcr Rats, a Metabolic Syndrome Model" Molecules 19, no. 3: 3247-3263. https://doi.org/10.3390/molecules19033247