Insecticidal Activities of Bark, Leaf and Seed Extracts of Zanthoxylum heitzii against the African Malaria Vector Anopheles gambiae


Abstract
:
1. Introduction
2. Results and Discussion
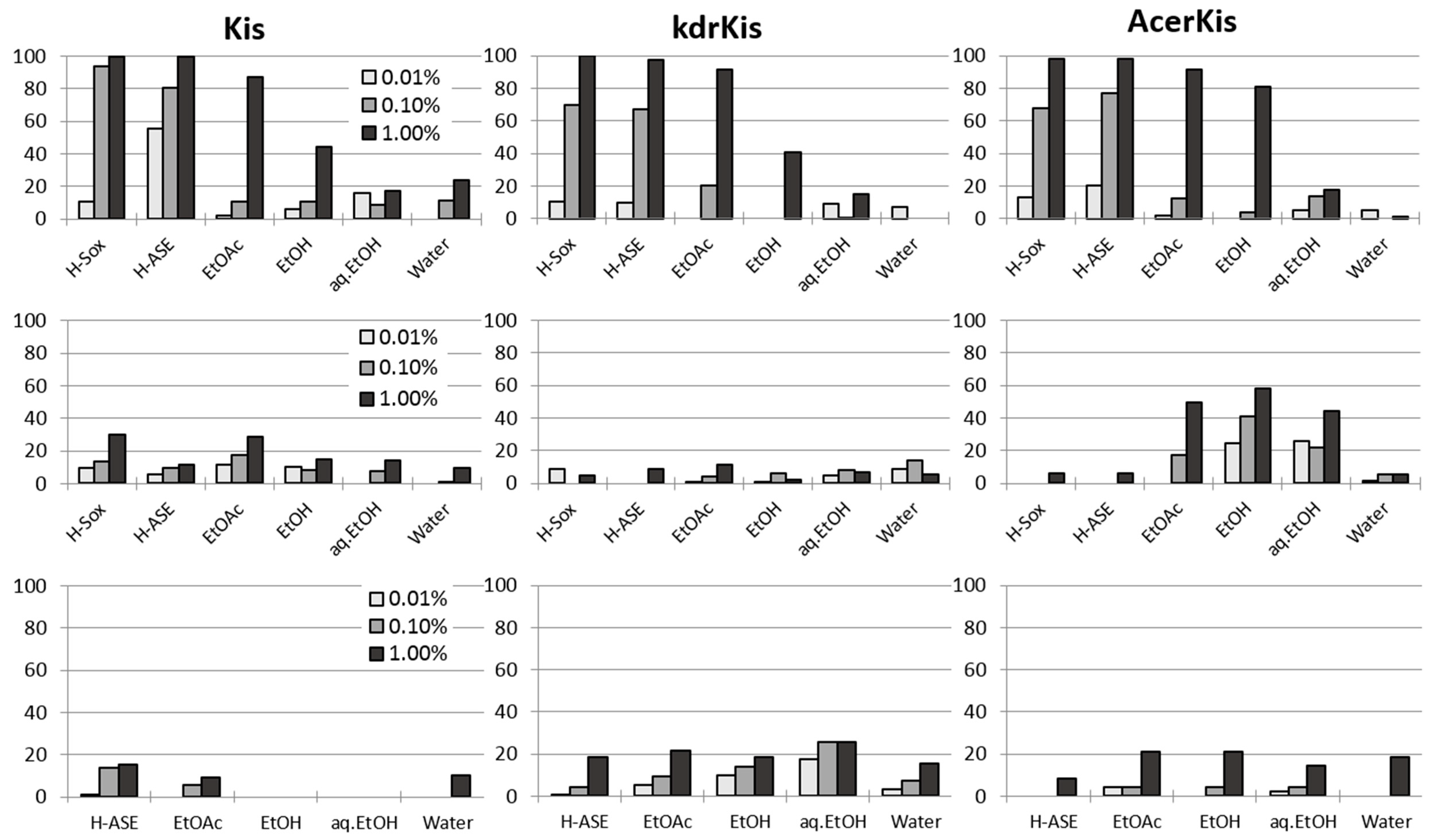
2.1. Extraction and Characterization of Active Fractions

{kind=link}
{kind=link}
{kind=link}
| # | Extraction Method | Solvent | Bark | Seed | Leaf |
|---|---|---|---|---|---|
| 1 | Soxhlet | Hexane | 2.0% | 29.0% | 2.7% |
| 2 | ASE | Hexane | 0.75% | 28.0% | 2.3% |
| 3 | ASE | Ethyl acetate | 0.4% | 1.9% | 1.5% |
| 4 | ASE | Ethanol (96%) | 2.4% | 7.9% | 13.5% |
| 5 | ASE | Ethanol/water (50%/50%) | 4.6% | 11.1% | 1.9% |
| 6 | ASE | Water (100%) | 1.6% | 2.3% | 5.8% |
2.2. Mosquito Bioassays
2.2.1. General Toxicity
2.2.2. Toxicity to Hexane Bark Extracts

| Group | Factors | n | Mortality a | df | F | p-Value | |
|---|---|---|---|---|---|---|---|
| % | 95% C.I. | ||||||
| Mosquito strain | Kis | 51 | 18.4 § | 10.8–26.0 | df1 = 2 df2 = 150 | 0.24 | 0.784 |
| AcerKis | 51 | 19.9 § | 12.1–27.8 | ||||
| kdrKis | 51 | 16.2 § | 9.3–23.2 | ||||
| Plant part | Stem bark | 54 | 33.8 † | 23.6–44.0 | df1 = 2 df2 = 150 | 17.99 | <0.0001 |
| Seed | 54 | 11.1 ‡ | 7.6–14.7 | ||||
| Leaf | 45 | 7.9 ‡ | 5.4–10.3 | ||||
| Extract concentration | 0.01% | 51 | 6.4 α | 3.7–9.1 | df1 = 2 df2 = 150 | 14.49 | <0.0001 |
| 0.10% | 51 | 16.0 β | 9.3–22.7 | ||||
| 1.00% | 51 | 32.1 γ | 22.7–41.6 | ||||
| Extraction method/solvent | Soxhlet/hexane | 18 | 35.3 a | 15.5–55.2 | df1 = 5 df2 = 147 | 3.91 | 0.002 |
| ASE/hexane | 27 | 26.2 ac | 12.3–40.2 | ||||
| ASE/ethyl acetate | 27 | 19.9 bc | 9.0–30.8 | ||||
| ASE/96% ethanol | 27 | 15.5 bc | 7.4–23.6 | ||||
| ASE/50% ethanol-water | 27 | 12.1 bd | 7.9–16.3 | ||||
| ASE/100% water | 27 | 5.7 bd | 3.1–8.3 | ||||
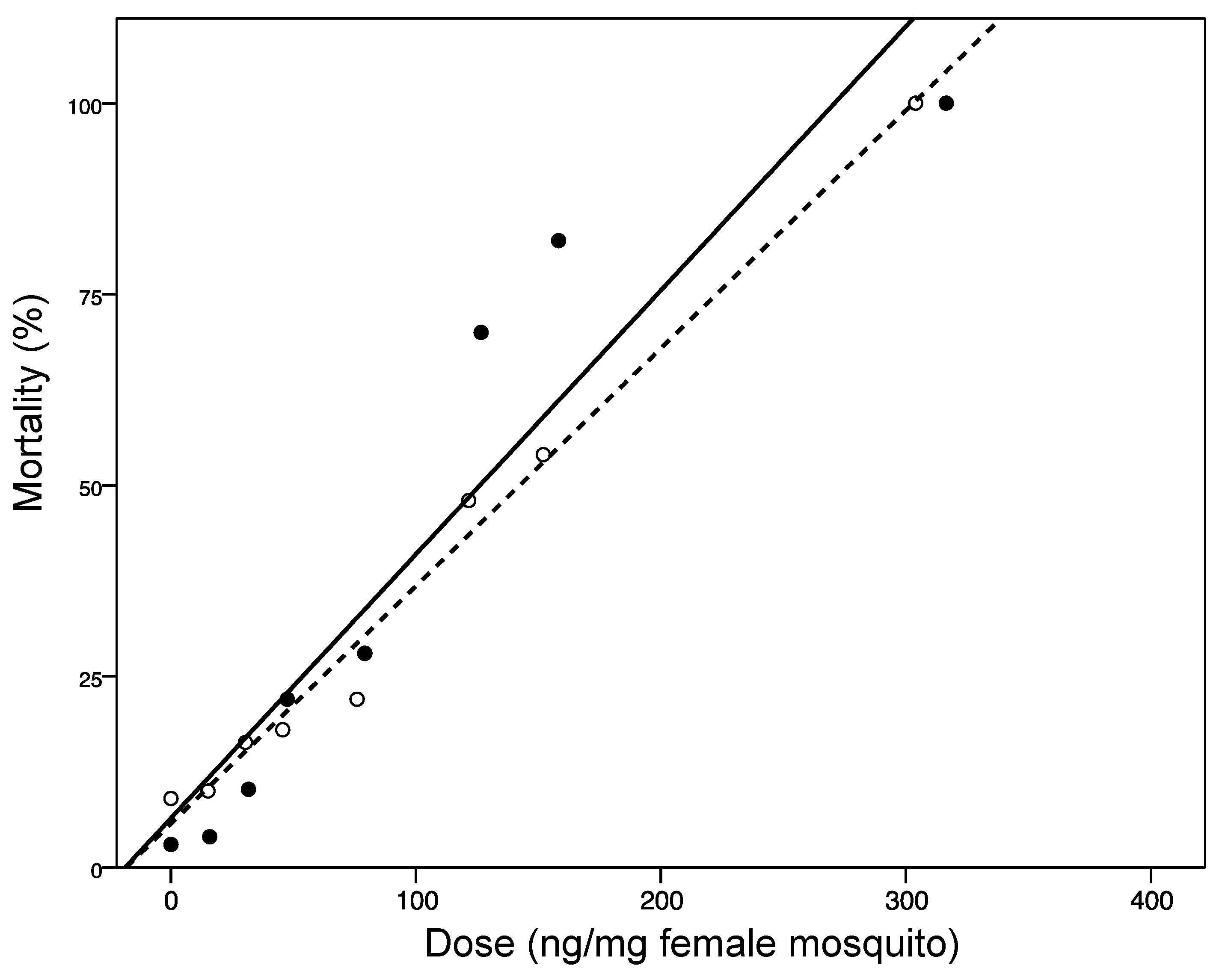
2.3. Discussion

| Extraction Method | Slope (±SD) | LD50 ng/mg Female (95% CI) | LD95 ng/mg Female (95% CI) |
|---|---|---|---|
| ASE | 4.55 (1.07) | 101.86 (74.75–117.93) | 234.05 (191.85–379.84) |
| Soxhlet | 6.50 (1.49) | 144.45 (124.99–161.59) | 258.71 (214.93–404.18) |
3. Experimental Section
3.1. Plant Material
3.2. Plant Preparation
3.3. Extraction
3.4. Mosquitoes
- Kis: The Kisumu strain, originating from Kenya, is free of any detectable insecticide resistance mechanisms.
- kdrKis: A pyrethroid and DDT resistant strain. This strain was obtained by introgression of L1014 F (kdr) into the genome of the susceptible Kisumu strain through successive backcrosses and selection with permethrin (1 mg/L). kdrKis has the same genetic background as the Kisumu strain but has the L1014F allele at the homozygous state.
- AcerKis: An organophosphate and carbamate resistant strain. This strain was obtained by introgression of insensitive acetylcholinesterase (Ace1R) into the genome of the Kisumu strain through successive backcrosses and selection with propoxur (10 mg/L). AcerKis has the same genetic background as the Kisumu strain but differs by the presence of Ace1R allele (G119S) at homozygous state [45].
3.5. Mosquito Bioassays
3.5.1. Step 1—General Toxicity
3.5.2. Step 2—Toxicity to Hexane Bark Extracts
3.6. Data Analysis
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kilama, W.L. Ethical perspective on malaria research for Africa. Acta Trop. 2005, 95, 276–284. [Google Scholar] [CrossRef] [PubMed]
- Sachs, J.; Malaney, P. The economic and social burden of malaria. Nature 2002, 415, 680–685. [Google Scholar] [CrossRef] [PubMed]
- WHO. Guidelines for Testing Mosquito Adulticides for Indoor Residual Spraying and Treatment of Mosquito Nets; World Health Organization: Geneva, Switzerland, 2006. [Google Scholar]
- WHO. Who Recommended Insecticide Products Treatment of Mosquito Nets for Malaria Vector Control (Updated December 2007). Available online: http://www.who.int/entity/whopes/Insecticides_ITN_Malaria_ok3.pdf?ua=1 (accessed on 1 June 2014).
- WHO. WHO Recommended Insecticides for Indoor Residual Spraying against Malaria Vectors. (Updated: 25 October 2013). Available online: http://www.who.int/entity/whopes/Insecticides_IRS_Malaria_25_Oct_2013.pdf?ua=1 (accessed on 1 June 2014).
- WHO. Who Recommended Long-Lasting Insecticidal Nets (Updated 6 February 2014). Available online: http://www.who.int/entity/whopes/Long_lasting_insecticidal_nets_06_Feb_2014.pdf?ua=1 (accessed on 1 June 2014).
- Corbel, V.; N’Guessan, R. Distribution, mechanisms, impact and management of insecticide resistance in malaria vectors: A pragmatic review. In Anopheles Mosquitoes—New Insights into Malaria Vectors; Manguin, P.S., Ed.; InTech: Rijeka, Croatia, 2013; Available online: http://www.intechopen.com/books/anopheles-mosquitoes-new-insights-into-malaria-vectors/distribution-mechanisms-impact-and-management-of-insecticide-resistance-in-malaria-vectors-a-pragmat (accessed on 17 September 2014).
- Ranson, H.; N’Guessan, R.; Lines, J.; Moiroux, N.; Nkuni, Z.; Corbel, V. Pyrethroid resistance in African anopheline mosquitoes: What are the implications for malaria control? Trends Parasitol. 2011, 27, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Hemingway, J.; Beaty, B.J.; Rowland, M.; Scott, T.W.; Sharp, B.L. The innovative vector control consortium: Improved control of mosquito-borne diseases. Trends Parasitol. 2006, 22, 308–312. [Google Scholar] [CrossRef] [PubMed]
- Trape, J.F.; Tall, A.; Diagne, N.; Ndiath, O.; Ly, A.B.; Faye, J.; Dieye-Ba, F.; Roucher, C.; Bouganali, C.; Badiane, A.; et al. Malaria morbidity and pyrethroid resistance after the introduction of insecticide-treated bednets and artemisinin-based combination therapies: A longitudinal study. Lancet Infect. Dis. 2011, 11, 925–932. [Google Scholar] [CrossRef] [PubMed]
- Corbel, V.; Duchon, S.; Zaim, M.; Hougard, J.M. Dinotefuran: A potential neonicotinoid insecticide against resistant mosquitoes. J. Med. Entomol. 2004, 41, 712–717. [Google Scholar] [CrossRef] [PubMed]
- N’Guessan, R.; Corbel, V.; Bonnet, J.; Yates, A.; Asidi, A.; Boko, P.; Odjo, A.; Akogbeto, M.; Rowland, M. Evaluation of indoxacarb, an oxadiazine insecticide for the control of pyrethroid-resistant Anopheles gambiae (Diptera: Culicidae). J. Med. Entomol. 2007, 44, 270–276. [Google Scholar] [CrossRef] [PubMed]
- N’Guessan, R.; Corbel, V.; Bonnet, J.; Akogbeto, M.; Yates, A.; Rowland, M. Indoxacarb and chlorfenapyr: Alternative insecticides with potential for use on nets against pyrethroid resistant Anopheles gambiae and Culex quinquefasciatus mosquitoes MIM-RN-268086. Acta Trop. 2005, 95, S133. [Google Scholar]
- Shaalan, E.A.; Canyon, D.; Younes, M.W.; Abdel-Wahab, H.; Mansour, A.H. A review of botanical phytochemicals with mosquitocidal potential. Environ. Int. 2005, 31, 1149–1166. [Google Scholar] [CrossRef] [PubMed]
- Adesina, S.K. The Nigerian Zanthoxylum; chemical and biological values. Afr. J. Tradit. Complement. Altern. Med. 2005, 2, 282–301. [Google Scholar]
- Lye, K.A.; Bukenya-Ziraba, R.; Tabuti, J.R.S.; Waako, P.J. Plant-Medicinal Dictionary for East Africa; Department of Botany; Makerere University: Kampala, Uganda, 2008; pp. 418–420. [Google Scholar]
- Garcez, W.S.; Garcez, F.R.; da Silva, L.; Hamerski, L. Larvicidal activity against Aedes aegypti of some plants native to the West-Central region of Brazil. Bioresour. Technol. 2009, 100, 6647–6650. [Google Scholar] [CrossRef] [PubMed]
- Ginesta, E.; Cunat, P.; Primo, J.; Primoyufera, E. Compounds with ovicidal effect isolated from Fagara zanthoxyloides Lam. Biosci. Biotechnol. Biochem. 1994, 58, 936–937. [Google Scholar] [CrossRef]
- He, W.D.; van Puyvelde, L.; de Kimpe, N.; Verbruggen, L.; Anthonissen, K.; van der Flaas, M.; Bosselaers, J.; Mathenge, S.G.; Mudida, F.P. Chemical constituents and biological activities of Zanthoxylum usambarense. Phytother. Res. 2002, 16, 66–70. [Google Scholar] [CrossRef] [PubMed]
- Kubo, I.; Matsumoto, T.; Klocke, J.A.; Kamikawa, T. Molluscicidal and insecticidal activities of isobutylamides isolated from Fagara macrophylla. Experientia 1984, 40, 340–341. [Google Scholar] [CrossRef]
- Navarrete, A.; Flores, A.; Sixtos, C.; Reyes, B. Analisis isobolografico de la interaccion entre α-sanshool, sesamina, asarinina, fagaramida y piperina sobre la actividad larvicida en Culex quinquefasciatus Say. Rev. Soc. Quim. México 2003, 47, 178–185. [Google Scholar]
- Nissanka, A.P.K.; Karunaratne, V.; Bandara, B.M.R.; Kumar, V.; Nakanishi, T.; Nishi, M.; Inada, A.; Tillekeratne, L.M.V.; Wijesundara, D.S.A.; Gunatilaka, A.A.L. Antimicrobial alkaloids from Zanthoxylum tetraspermum and caudatum. Phytochemistry 2001, 56, 857–861. [Google Scholar] [CrossRef] [PubMed]
- Pitasawat, B.; Champakaew, D.; Choochote, W.; Jitpakdi, A.; Chaithong, U.; Kanjanapothi, D.; Rattanachanpichai, E.; Tippawangkosol, P.; Riyong, D.; Tuetun, B.; et al. Aromatic plant-derived essential oil: An alternative larvicide for mosquito control. Fitoterapia 2007, 78, 205–210. [Google Scholar] [CrossRef] [PubMed]
- Talontsi, F.M.; Matasyoh, J.C.; Ngoumfo, R.M.; Chepkorir, R. Mosquito larvicidal activity of alkaloids from Zanthoxylum lemairei against the malaria vector Anopheles gambiae. Pestic. Biochem. Physiol. 2011, 99, 82–85. [Google Scholar] [CrossRef]
- Tiwary, M.; Naik, S.N.; Tewary, D.K.; Mittal, P.K.; Yadav, S. Chemical composition and larvicidal activities of the essential oil of Zanthoxylum armatum DC (Rutaceae) against three mosquito vectors. J. Vector Borne Dis. 2007, 44, 198–204. [Google Scholar] [PubMed]
- Tafokou, J.R.B. Zanthoxylum heitzii (Aubrév. & Pellegr.) P.G. Waterman. Available online: http://database.prota.org/PROTAhtml/Zanthoxylum%20heitzii_En.htm (accessed on 1 June 2014).
- Betti, J.L.; Lejoly, J. Contribution to the knowledge of medicinal plants of the Dja biosphere reserve, Cameroon: Plants used for treating jaundice. J. Med. Plant Res. 2009, 3, 1056–1065. [Google Scholar]
- Betti, J.L. An ethnobotanical study of medicinal plants among the baka pygmies in the Dja biosphere reserve, Cameroon. Afr. Study Monogr. 2004, 25, 1–27. [Google Scholar]
- Betti, J.L. Medicinal plants sold in Yaoundé markets, Cameroon. Afr. Study Monogr. 2002, 23, 47–64. [Google Scholar]
- Ahmad, S. Flindersine from Fagara heitzii. J. Nat. Prod. 1984, 47, 391–392. [Google Scholar] [CrossRef]
- Bongui, J.B.; Blanckaert, A.; Elorari, A.; Seguin, E. Constituents of Zanthoxylum heitzii (Rutaceae). Biochem. Syst. Ecol. 2005, 33, 845–847. [Google Scholar] [CrossRef]
- Mbaze, L.M.; Lado, J.A.; Wansi, J.D.; Shiao, T.C.; Chiozem, D.D.; Mesaik, M.A.; Choudhary, M.I.; Lacaille-Dubois, M.A.; Wandji, J.; Roy, R.; et al. Oxidative burst inhibitory and cytotoxic amides and lignans from the stem bark of Fagara heitzii (Rutaceae). Phytochemistry 2009, 70, 1442–1447. [Google Scholar] [CrossRef] [PubMed]
- Ngouela, S.; Tsamo, E.; Connolly, J.D. Lignans and other constituents of Zanthoxylum heitzii. Phytochemistry 1994, 37, 867–869. [Google Scholar] [CrossRef]
- Mikolo, B.; Matos, L.; Massamba, D.; Mamonekene, V.; Miller, T. Extracts from the bark of Fagara heitzii (Aubr. et Pel.) (Rutaceae) tree are toxic to two weevils and the American cockroach. Entomol. Res. 2009, 39, 401–405. [Google Scholar] [CrossRef]
- Mengome, L.E.; Akue, J.P.; Souza, A.; Tchoua, G.R.F.; Emvo, E.N. In vitro activities of plant extracts on human loa loa isolates and cytotoxicity for eukaryotic cells. Parasitol. Res. 2010, 107, 643–650. [Google Scholar] [CrossRef] [PubMed]
- Eto, B.; Pyebi-Oyoubi, P.; Louma-Eyougha, A. Composition Pharmaceutique Utilisable Notamment en Tant qu´Antiviral, Antibacterien et pour Stimuler les Defenses Immunitaires. Eur. Patent EP 1,675,605 B1, 9 April 2008. [Google Scholar]
- Dzoyem, J.P.; Guru, S.K.; Pieme, C.A.; Kuete, V.; Sharma, A.; Khan, I.A.; Saxena, A.K.; Vishwakarma, R.A. Cytotoxic and antimicrobial activity of selected Cameroonian edible plants. BMC Complement. Altern. Med. 2013, 13, 78. [Google Scholar] [CrossRef] [PubMed]
- Bonnet, J.; Corbel, V.; Darriet, F.; Chandre, F.; Hougard, J.M. Topical applications of pyrethroid and organophosphate mixtures revealed positive interactions against pyrethroid-resistant Anopheles gambiae. J. Am. Mosq. Control Assoc. 2004, 20, 438–443. [Google Scholar] [PubMed]
- Duchon, S.; Bonnet, J.; Marcombe, S.; Zaim, M.; Corbel, V. Pyrethrum: A mixture of natural pyrethrins has potential for malaria vector control. J. Med. Entomol. 2009, 46, 516–522. [Google Scholar] [CrossRef] [PubMed]
- WHO. Interim Position Statement: The Role of Larviciding for Malaria Control in Sub-Saharan Africa; WHO: Geneva, Switzerland, 2012. [Google Scholar]
- Andemo, A.; Yewhalaw, D.; Alemayehu, B.; Ambelu, A. Evaluation of the mosquitocidal effect of Birbira (Mellitia ferruginea) seed extract against Anopheles arabiensis (Diptera: Culicidae) from Ethiopia. Acta Trop. 2014, 136, 68–73. [Google Scholar] [CrossRef] [PubMed]
- Isman, M.B.; Grieneisen, M.L. Botanical insecticide research: Many publications, limited useful data. Trends Plant Sci. 2014, 19, 140–145. [Google Scholar] [CrossRef] [PubMed]
- The Plant List. Available online: http://www.theplantlist.org (accessed on 18 September 2014).
- Tailfer, Y. la Forêt Dense d'Afrique Centrale, Identification Pratique des Principaux Arbres; Centre Technique de Coopération Agricole et Rurale (CTA): Wageningen, The Netherlands, 1989; Volume 2, pp. 465–1271. [Google Scholar]
- Djogbenou, L.; Weill, M.; Hougard, J.M.; Raymond, M.; Akogbeto, M.; Chandre, F. Characterization of insensitive acetylcholinesterase (ace-1R) in Anopheles gambiae (Diptera: Culicidae): Resistance levels and dominance. J. Med. Entomol. 2007, 44, 805–810. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the Zanthoxylum heitzii bark used are available from the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Overgaard, H.J.; Sirisopa, P.; Mikolo, B.; Malterud, K.E.; Wangensteen, H.; Zou, Y.-F.; Paulsen, B.S.; Massamba, D.; Duchon, S.; Corbel, V.; et al. Insecticidal Activities of Bark, Leaf and Seed Extracts of Zanthoxylum heitzii against the African Malaria Vector Anopheles gambiae. Molecules 2014, 19, 21276-21290. https://doi.org/10.3390/molecules191221276
Overgaard HJ, Sirisopa P, Mikolo B, Malterud KE, Wangensteen H, Zou Y-F, Paulsen BS, Massamba D, Duchon S, Corbel V, et al. Insecticidal Activities of Bark, Leaf and Seed Extracts of Zanthoxylum heitzii against the African Malaria Vector Anopheles gambiae. Molecules. 2014; 19(12):21276-21290. https://doi.org/10.3390/molecules191221276
Chicago/Turabian StyleOvergaard, Hans J., Patcharawan Sirisopa, Bertin Mikolo, Karl E. Malterud, Helle Wangensteen, Yuan-Feng Zou, Berit S. Paulsen, Daniel Massamba, Stephane Duchon, Vincent Corbel, and et al. 2014. "Insecticidal Activities of Bark, Leaf and Seed Extracts of Zanthoxylum heitzii against the African Malaria Vector Anopheles gambiae" Molecules 19, no. 12: 21276-21290. https://doi.org/10.3390/molecules191221276