Action of Chitosan Against Xanthomonas Pathogenic Bacteria Isolated from Euphorbia pulcherrima

Abstract
:1. Introduction
2. Results and Discussion
2.1. Molecular Weight and Deacetylation Degree
2.2. Antibacterial Activity of Chitosan

{kind=link}
{kind=link}
| Strain number | Species identity | Strain source | OD600 reduction relative to the control (%) | |
|---|---|---|---|---|
| Chitosan A | Chitosan B | |||
| LMG849 | X. axonopodis pv. poinsettiicola | India | 24.20 ± 2.32 cd | 20.94 ± 2.29 c |
| LMG5401 | X. axonopodis pv. poinsettiicola | India | 49.95 ± 4.48 f | 52.52 ± 3.21 e |
| LMG5402 | X. arboricola pv. poinsettiicola | New Zealand | 61.88 ± 6.07 g | 14.35 ± 2.04 ab |
| LMG5403 | X. arboricola pv. poinsettiicola | New Zealand | 53.50 ± 4.74 fg | 15.40 ± 1.61 abc |
| LMG8675 | X. arboricola pv. poinsettiicola | New Zealand | 14.40 ± 1.25 ab | 16.35 ± 1.25 abc |
| LMG8676 | X. arboricola pv. poinsettiicola | New Zealand | 17.11 ± 1.77 abc | 16.07 ± 1.94 abc |
| R22578 | X. axonopodis pv. poinsettiicola | Hangzhou, China | 17.91 ± 1.61 abc | 19.75 ± 1.22 bc |
| R22579 | X. axonopodis pv. poinsettiicola | Hangzhou, China | 39.47 ± 4.16 e | 13.12 ± 1.28 a |
| R22580 | X. axonopodis pv. poinsettiicola | Hangzhou, China | 30.52 ± 3.27 de | 39.51 ± 3.16 d |
| HN-1 | X. axonopodis pv. poinsettiicola | Hainan, China | 11.17 ± 1.27 a | 34.53 ± 2.29 d |
| HN-18 | X. axonopodis pv. poinsettiicola | Hainan, China | 23.32 ± 2.43 bcd | 34.05 ± 2.21 d |
| HN-20 | X. axonopodis pv. poinsettiicola | Hainan, China | 9.26 ± 1.10 a | 16.45 ± 1.45 abc |
| LSD0.05 | 9.33 | 6.13 | ||
| ANOVA P-values | ||||
| Chitosan | <0.001 | |||
| Bacteria | <0.001 | |||
| Chitosan × Bacteria | <0.001 | |||
2.3. Antibacterial Mechanism of Chitosan
2.3.1. Integrity of Cell Membrane
| Chitosan (mg/mL) | The ratio release ( vs. control) at OD260 (%) | ||||
|---|---|---|---|---|---|
| 0.5 h | 1.0 h | 1.5 h | 2.0 h | ||
| A | |||||
| 0.1 | 104.05 ± 0.03 b | 103.74 ± 0.01 a | 103.07 ± 0.04 b | 103.23 ± 0.04 b | |
| 0.2 | 104.24 ± 0.04 c | 104.32 ± 0.04 c | 103.65 ± 0.02 c | 104.20 ± 0.04 d | |
| 0.3 | 103.08 ± 0.03 a | 104.09 ± 0.05 b | 102.66 ± 0.08 a | 103.65 ± 0.07 c | |
| 0.4 | 103.02 ± 0.04 a | 104.15 ± 0.08 b | 102.76 ± 0.02 a | 102.97 ± 0.04 a | |
| 0.5 | 104.92 ± 0.02 c | 105.82 ± 0.03 d | 105.56 ± 0.05 d | 106.33 ± 0.02 e | |
| B | |||||
| 0.1 | 103.77 ± 0.01 e | 104.44 ± 0.05 c | 103.06 ± 0.03 c | 103.67 ± 0.03 d | |
| 0.2 | 103.34 ± 0.03 d | 104.34 ± 0.06 c | 103.18 ± 0.07 c | 103.54 ± 0.07 d | |
| 0.3 | 101.99 ± 0.02 b | 103.53 ± 0.06 b | 102.64 ± 0.04 b | 103.01 ± 0.05 c | |
| 0.4 | 102.28 ± 0.01 c | 103.13 ± 0.07 a | 102.29 ± 0.10 a | 102.48 ± 0.06 a | |
| 0.5 | 101.62 ± 0.01 a | 103.21 ± 0.02 a | 102.45 ± 0.04 ab | 102.82 ± 0.06 b | |
| LSD0.05 | 0.08 | 0.15 | 0.16 | 0.15 | |
| ANOVA P-values | |||||
| Chitosan | <0.001 | ||||
| Incubation time | <0.001 | ||||
| Chitosan × Incubation time | 0.56 | ||||
| Chitosan (mg/mL) | The ratio release at OD280 | |||||
|---|---|---|---|---|---|---|
| 0.5 h | 1.0 h | 1.5 h | 2.0 h | |||
| A | ||||||
| 0.1 | 99.64 ± 0.03 c | 100.49 ± 0.06 d | 100.25 ± 0.07 d | 100.30 ± 0.01 d | ||
| 0.2 | 100.58 ± 0.01 d | 102.01 ± 0.02 e | 100.88 ± 0.08 e | 99.83 ± 0.03 c | ||
| 0.3 | 98.87 ± 0.02 b | 99.63 ± 0.16 c | 99.11 ± 0.14 c | 98.77 ± 0.02 b | ||
| 0.4 | 98.32 ± 0.01 a | 98.14 ± 0.04 a | 97.97 ± 0.07 a | 97.37 ± 0.06 a | ||
| 0.5 | 98.32 ± 0.06 a | 98.48 ± 0.08 b | 98.72 ± 0.07 b | 98.84 ± 0.15 b | ||
| B | ||||||
| 0.1 | 92.24 ± 0.10 a | 92.59 ± 0.04 a | 93.33 ± 0.08 a | 94.23 ± 0.06 a | ||
| 0.2 | 97.61 ± 0.04 d | 98.18 ± 0.01 c | 98.61 ± 0.09 d | 98.11 ± 0.02 c | ||
| 0.3 | 97.38 ± 0.03 c | 97.26 ± 0.06 b | 98.27 ± 0.04 c | 98.79 ± 0.01 e | ||
| 0.4 | 98.15 ± 0.09 e | 98.35 ± 0.03 d | 98.25 ± 0.04 c | 98.30 ± 0.05 d | ||
| 0.5 | 96.70 ± 0.01 b | 97.26 ± 0.02 b | 97.27 ± 0.04 b | 96.76 ± 0.08 b | ||
| LSD0.05 | 0.15 | 0.13 | 0.22 | 0.14 | ||
| ANOVA P-values | ||||||
| Chitosan | <0.001 | |||||
| Incubation time | 0.65 | |||||
| Chitosan × Incubation time | 0.49 | |||||
2.3.2. Biofilm Formation
| Strain number | OD570 reduction relative to the control (%) | ||
|---|---|---|---|
| Chitosan A | Chitosan B | ||
| LMG849 | 18.05 ± 2.02 de | 28.56 ± 3.43 ef | |
| LMG5401 | 32.96 ± 3.52 g | 21.25 ± 2.37 cd | |
| LMG5402 | 27.53 ± 2.19 fg | 22.15 ± 3.46 de | |
| LMG5403 | 28.70 ± 2.18 g | 14.91 ± 1.77 abc | |
| LMG8675 | 7.31 ± 1.04 a | 16.79 ± 1.76 bcd | |
| LMG8676 | 9.36 ± 1.26 ab | 11.68 ± 1.13 ab | |
| R22578 | 14.13 ± 1.60 bcd | 10.37 ± 1.72 ab | |
| R22579 | 9.72 ± 0.64 ab | 11.73 ± 1.07 ab | |
| R22580 | 10.90 ± 1.04 abc | 9.47 ± 0.84 a | |
| HN-1 | 15.47 ± 1.75 cd | 12.82 ± 1.63 ab | |
| HN-18 | 28.09 ± 2.17 fg | 42.52 ± 3.57 g | |
| HN-20 | 23.07 ± 1.96 ef | 35.09 ± 2.96 f | |
| LSD0.05 | 5.51 | 6.84 | |
| ANOVA P-values | |||
| Chitosan | 0.25 | ||
| Bacteria | <0.001 | ||
| Chitosan × Bacteria | <0.001 | ||
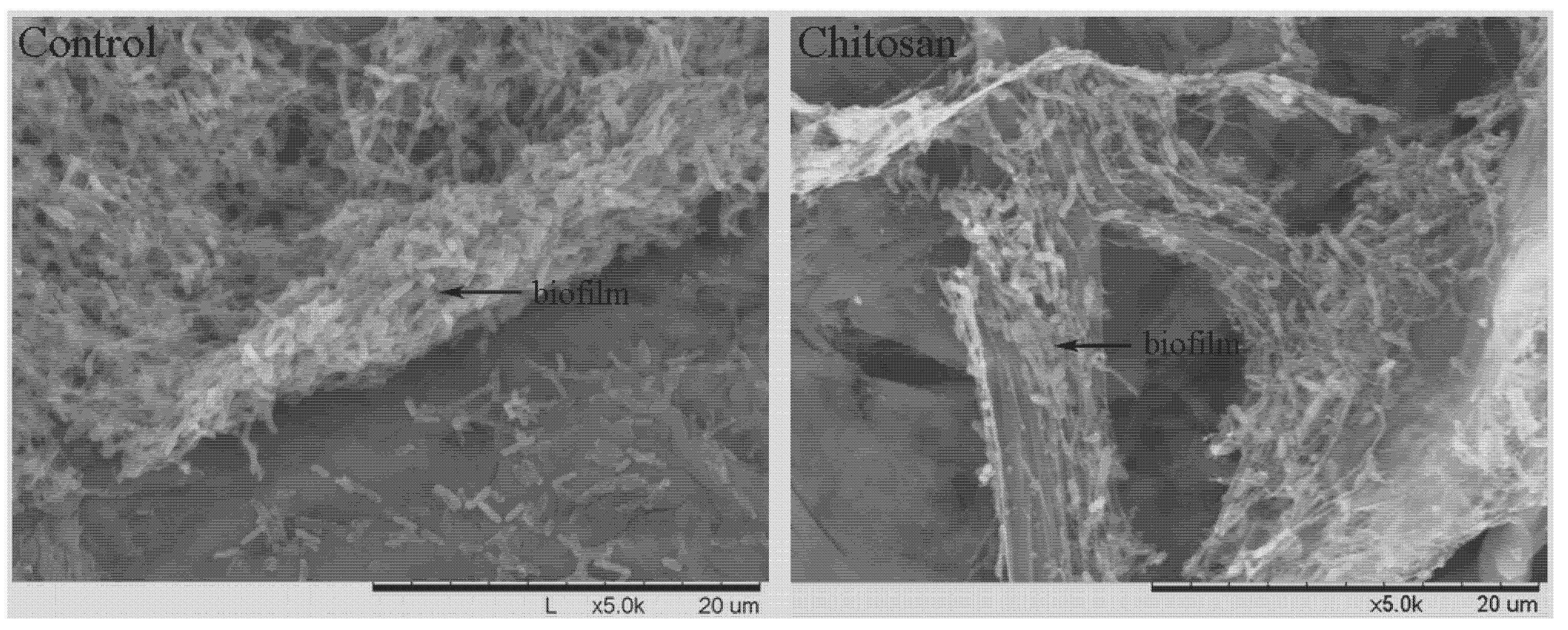
2.3.3. SEM

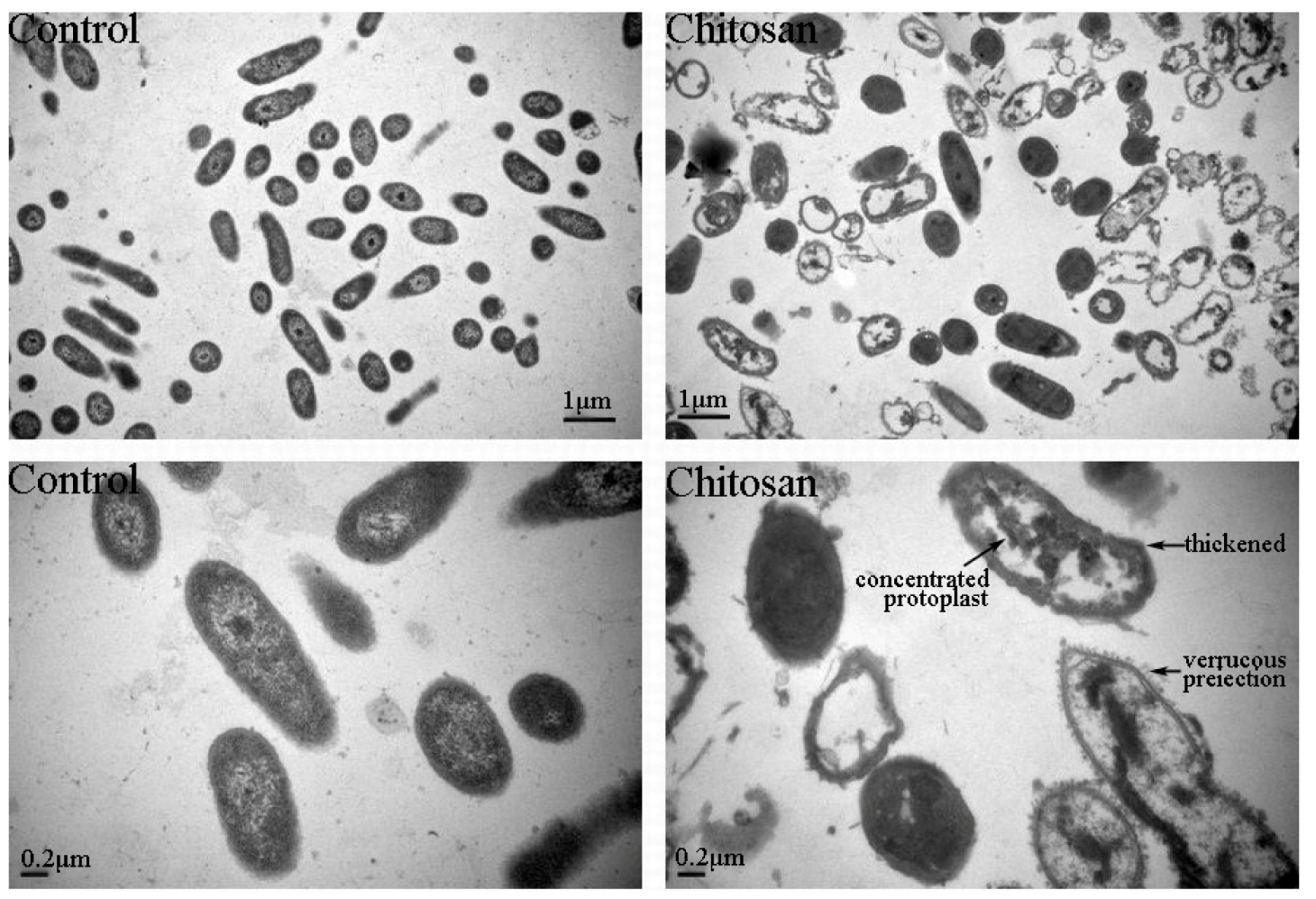
2.3.4. TEM

3. Experimental
3.1. Preparation of Chitosan Stock
3.2. Molecular Weight and Deacetylation Degree
3.3. Bacterial Strains
3.4. Inhibition Assessment
3.5. Integrity of Cell Membrane
3.6. Biofilm Formation
3.7. Scanning Electron Microscope (SEM)
3.8. Transmission Electron Microscope (TEM)
3.9. Statistical Analysis
4. Conclusions
Acknowledgements
Conflict of Interest
- Sample Availability: Samples of the compounds are available from the authors.
References and Notes
- Tajik, H.; Moradi, M.; Rohani, S.M.R.; Erfani, A.M.; Jalali, F.S.S. Preparation of chitosan from brine shrimp (Artemia urmiana) cyst shells and effects of different chemical processing sequences on the physicochemical and functional properties of the product. Molecules 2008, 13, 1263–1274. [Google Scholar] [CrossRef]
- Li, B.; Wang, X.; Chen, R.; Huangfu, W.G.; Xie, G.L. Antibacterial activity of chitosan solution against Xanthomonas pathogenic bacteria isolated from Euphorbia pulcherrima. Carbohyd. Polym. 2008, 72, 287–292. [Google Scholar] [CrossRef]
- Chung, Y.C.; Yeh, J.Y.; Tsai, C.F. Antibacterial characteristics and activity of water-soluble chitosan derivatives prepared by the maillard reaction. Molecules 2011, 16, 8504–8514. [Google Scholar] [CrossRef]
- Orgaz, B.; Lobete, M.M.; Puga, C.H.; San Jose, C. Effectiveness of chitosan against mature biofilms formed by food related bacteria. Int. J. Mol. Sci. 2011, 12, 817–828. [Google Scholar] [CrossRef]
- Badawy, M.E.I. Effect of depolymerization degree of the natural biopolymer chitosan on some plant pathogenic bacteria and fungi. J. Pest Cont. Environ. Sci. 2007, 15, 69–85. [Google Scholar]
- Badawy, M.E.I. Structure and antimicrobial activity relationship of quaternary N-alkyl chitosan derivatives against some plant pathogens. J. Appl. Polym. Sci. 2010, 117, 960–969. [Google Scholar] [CrossRef]
- Fujimoto, T.; Tsuchiya, Y.; Terao, M.; Nakamura, K.; Yamamoto, M. Antibacterial effects of chitosan solution against Legionella pneumophila, Escherichia coli, and Staphylococcus aureus. Int. J. Food Microbiol. 2006, 112, 96–101. [Google Scholar] [CrossRef]
- Devlieghere, F.; Vermeulen, A.; Debevere, J. Chitosan: antimicrobial activity, interactions with food components and applicability as a coating on fruit and vegetables. Food Microbiol. 2004, 21, 703–714. [Google Scholar] [CrossRef]
- El Hadrami, A.; Adam, L.R.; El Hadrami, I.; Daayf, F. Chitosan in plant protection. Mar. Drugs 2010, 8, 968–987. [Google Scholar] [CrossRef]
- Helander, I.M.; Nurmiaho-Lassila, E.L.; Ahvenainen, R.; Rhoades, J.; Roller, S. Chitosan disrupts the barrier properties of the outer membrane of gram-negative bacteria. Int. J. Food Microbiol. 2001, 71, 235–244. [Google Scholar] [CrossRef]
- Li, B.; Liu, B.P.; Su, T.; Wang, F.; Tang, Q.M.; Fang, Y.; Xie, G.L.; Sun, G.C. Effect of chitosan solution on the inhibition of Pseudomonas fluorescens causing bacterial head rot of broccoli. Plant Pathol. J. 26, 189–193.
- Li, B.; Su, T.; Chen, X.L.; Liu, B.P.; Zhu, B.; Fang, Y.; Qiu, W.; Xie, G.L. Effect of chitosan solution on the bacterial septicemia disease of Bombyx mori (Lepidoptera: Bombycidae) caused by Serratia marcescens. Appl. Ento. Zool. 45, 145–152.
- Li, B.; Yu, R.R.; Liu, B.P.; Tang, Q.M.; Zhang, G.Q.; Wang, Y.L.; Xie, G.L.; Sun, G.C. Characterization and comparison of Serratia marcescens isolated from edible cactus and from silkworm for virulence potential and chitosan susceptibility. Braz. J. Microbiol. 2011, 42, 96–104. [Google Scholar] [CrossRef]
- Chen, C.Z.S.; Cooper, S.L. Interactions between dendrimer biocides and bacterial membranes. Biomaterials 2002, 23, 3359–3368. [Google Scholar] [CrossRef]
- Didenko, L.V.; Gerasimenko, D.V.; Konstantinova, N.D.; Silkina, T.A.; Avdienko, I.D.; Bannikova, G.E.; Varlamov, V.P.B. Ultrastructural study of chitosan effects on Klebsiella and Staphylococci. Exp. Biol. Med. 2005, 140, 356–360. [Google Scholar] [CrossRef]
- Eaton, P.; Fernandes, J.C.; Pereira, E.; Pintado, M.E.; Malcata, F.X. Atomic force microscopy study of the antibacterial effects of chitosans on Escherichia coli and Staphylococcus aureus. Ultramicroscopy 2008, 108, 1128–1134. [Google Scholar] [CrossRef]
- Tang, H.; Zhang, P.; Kieft, T.L.; Ryan, S.J.; Baker, S.M.; Wiesmann, W.P.; Rogelj, S. Antibacterial action of a novel functionalized chitosan-arginine against Gram-negative bacteria. Acta Biomater. 2010, 6, 2562–2571. [Google Scholar] [CrossRef]
- Andres, Y.; Giraud, L.; Gerente, C.; Cloirec, P.L. Antibacterial effects of chitosan powder: mechanisms of action. Environ. Technol. 2007, 28, 1357–1363. [Google Scholar] [CrossRef]
- Goy, R.C.; de Britto, D.; Assis, O.B.G. A review of the antimicrobial activity of chitosan. Polim. Cienc. Tecnol. 2009, 19, 241–247. [Google Scholar]
- Je, J.Y.; Kim, S.K. Chitosan derivatives killed bacteria by disrupting the outer and inner membrane. J. Agric. Food Chem. 2006, 54, 6629–6633. [Google Scholar] [CrossRef]
- Liu, H.; Du, Y.M.; Wang, X.H.; Sun, L.P. Chitosan kills bacteria through cell membrane damage. Int. J. Food Microbiol. 2004, 95, 147–155. [Google Scholar] [CrossRef]
- Carlson, R.P.; Taffs, R.; Davison, W.M.; Stewart, P.S. Anti-biofilm properties of chitosan-coated surfaces. J. Biomater. Sci. Polym. Ed. 2008, 19, 1035–1046. [Google Scholar] [CrossRef]
- Martinez, L.R.; Mihu, M.R.; Han, G.; Frases, S.; Cordero, R.J.B.; Casadevall, A.; Friedman, A.J.; Friedman, J.M.; Nosanchuk, J.D. The use of chitosan to damage Cryptococcus neoformans biofilms. Biomaterials 2010, 31, 669–679. [Google Scholar] [CrossRef]
- Raafat, D.; von Bargen, K.; Haas, A.; Sahl, H.G. Insights into the mode of action of chitosan as an antibacterial compound. Appl. Environ. Microbiol. 2008, 74, 3764–3773. [Google Scholar] [CrossRef]
- Muzzarelli, R.A.A.; Rocchetti, R. Determination of the degree of acetylation of chitosans by first derivative ultraviolet spectrophotometry. Carbohyd. Polym. 1985, 5, 461–472. [Google Scholar] [CrossRef]
- Li, B.; Yu, R.R.; Tang, Q.M.; Su, T.; Chen, X.L.; Zhu, B.; Wang, Y.L.; Xie, G.L.; Sun, G.C. Biofilm formation ability of Paenibacillus polymyxa and Paenibacillus macerans and their inhibitory effect against tomato bacterial wilt. Afr. J. Microbiol. Res. 2011, 5, 4260–4266. [Google Scholar]
- Li, X.F.; Feng, X.Q.; Yang, S.; Fu, G.Q.; Wang, T.P.; Su, Z.X. Chitosan kills Escherichia coli through damage to be of cell membrane mechanism. Carbohyd. Polym. 2010, 79, 493–499. [Google Scholar] [CrossRef]
- Li, Y.H.; Tan, N.; Aspiras, M.B.; Lau, P.C.Y.; Lee, J.H.; Ellen, R.P.; Cvitkovitch, D.G. A quorum-sensing signaling system essential for genetic competence in Streptococcus mutans is involved in biofilm formation. J. Bacteriol. 2002, 184, 2699–2708. [Google Scholar] [CrossRef]
- Loo, C.Y.; Corliss, D.A.; Ganeshkumar, N. Streptococcus gordonii biofilm formation: identification of genes that code for biofilm phenotypes. J. Bacteriol. 2000, 182, 1374–1382. [Google Scholar] [CrossRef]
- Rabea, E.I.; Badawy, M.E.T.; Stevens, C.V.; Smagghe, G.; Steurbaut, W. Chitosan as antimicrobial agent: Applications and mode of action. Biomacromolecules 2003, 4, 1457–1465. [Google Scholar] [CrossRef]
- Ren, D.; Sims, J.J.; Wood, T.K. Inhibition of biofilm formation and swarming of Bacillus subtilis by (5Z)-4-bromo-5-(bromomethylene)-3-butyl-2(5H)-furanone. Lett. Appl. Microbiol. 2002, 34, 293–299. [Google Scholar] [CrossRef]
- Pasquantonio, G.; Greco, C.; Prenna, M.; Ripa, C.; Vitali, L.A.; Petrelli, D.; Di Luca, M.C.; Ripa, S. Antibacterial activity and anti-biofilm effect of chitosan against strains of Streptococcus mutans isolated in dental plaque. Int. J. Immunopathol. Pharmacol. 2008, 21, 993–997. [Google Scholar]
- Shrestha, A.; Shi, Z.; Neoh, K.G.; Kishen, A. Nanoparticulates for antibiofilm treatment and effect of aging on its antibacterial activity. J. Endod. 2010, 36, 1030–1035. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wang, Y.; Li, L.; Li, B.; Wu, G.; Tang, Q.; Ibrahim, M.; Li, H.; Xie, G.; Sun, G. Action of Chitosan Against Xanthomonas Pathogenic Bacteria Isolated from Euphorbia pulcherrima. Molecules 2012, 17, 7028-7041. https://doi.org/10.3390/molecules17067028
Wang Y, Li L, Li B, Wu G, Tang Q, Ibrahim M, Li H, Xie G, Sun G. Action of Chitosan Against Xanthomonas Pathogenic Bacteria Isolated from Euphorbia pulcherrima. Molecules. 2012; 17(6):7028-7041. https://doi.org/10.3390/molecules17067028
Chicago/Turabian StyleWang, Yanli, Liping Li, Bin Li, Guoxing Wu, Qiaomei Tang, Muhammad Ibrahim, Hongye Li, Guanlin Xie, and Guochang Sun. 2012. "Action of Chitosan Against Xanthomonas Pathogenic Bacteria Isolated from Euphorbia pulcherrima" Molecules 17, no. 6: 7028-7041. https://doi.org/10.3390/molecules17067028