Role of Phosphatidyl-Serine in Bone Repair and Its Technological Exploitation

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Bone structure and function
2. Matrix Vesicles
3. Phospholipids and Calcification
4. A Role for Phosphatidyl-Serine

5. Technological Application of Artificial MVs
5.1. Calcium-binding phospholipids as coating molecules of medical implants: Early evidences of their osteointegrative potential
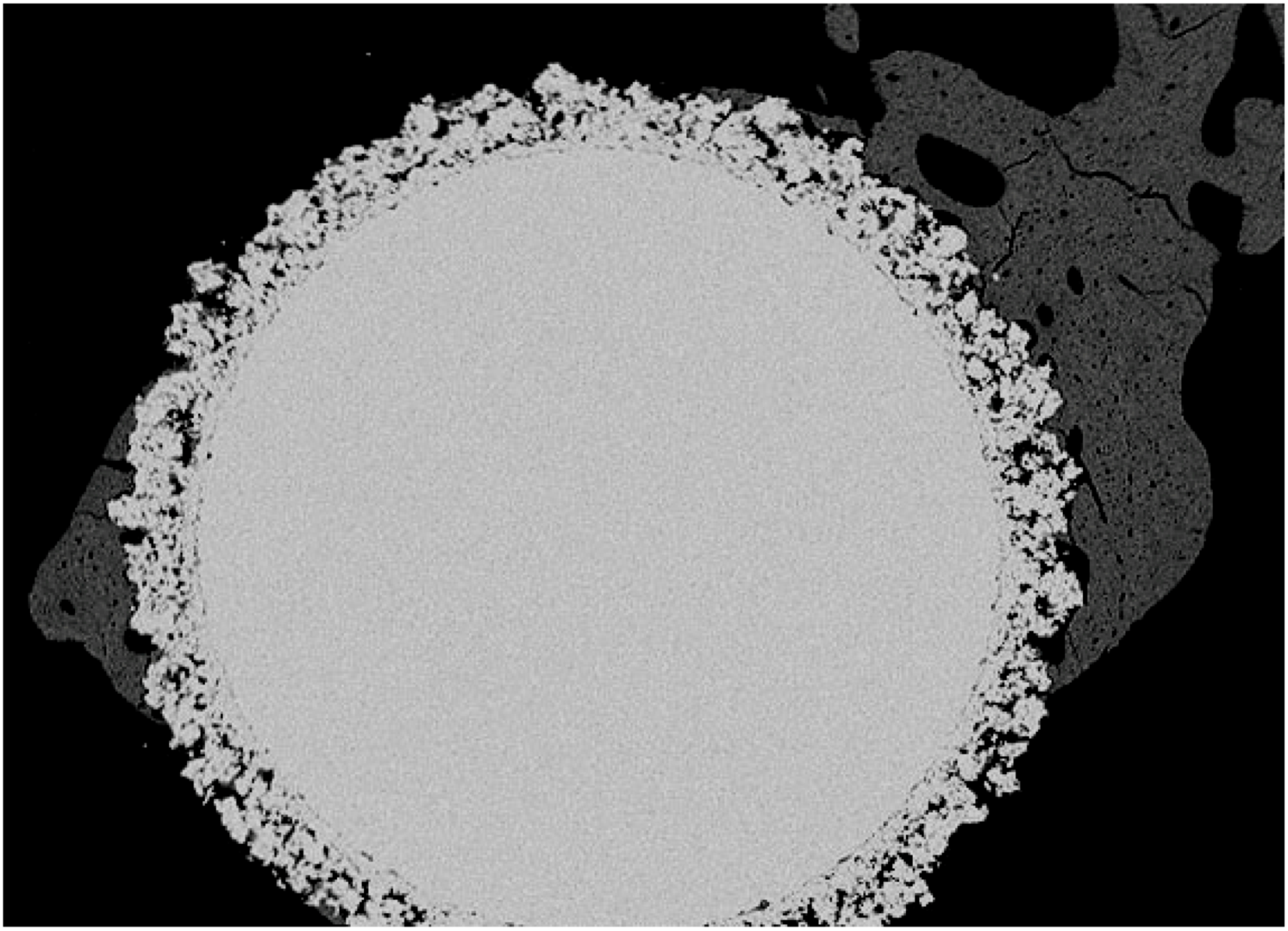
5.2. In vivo response to phosphatidyl-serine coating
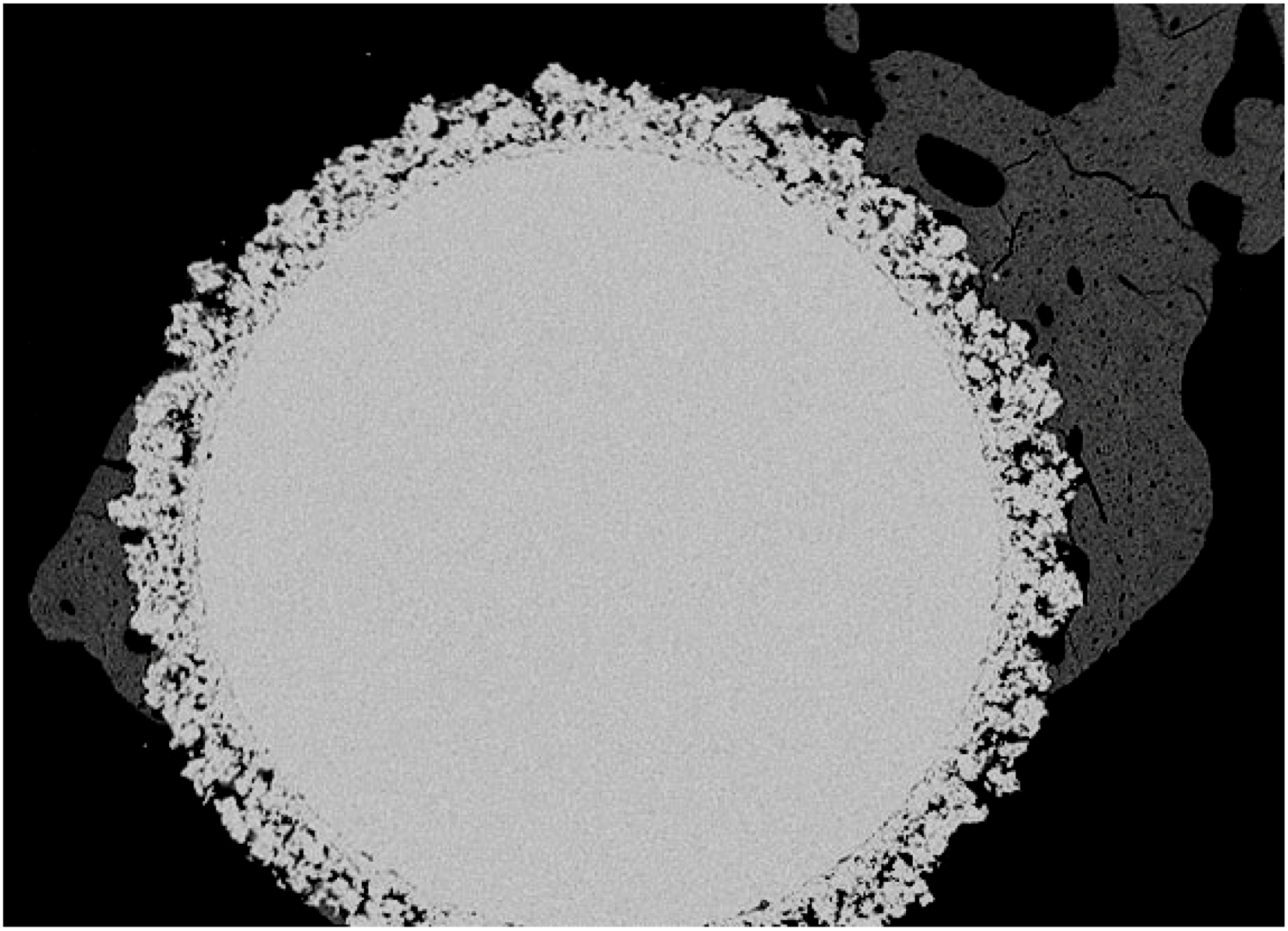

7. Conclusions
Acknowledgements
References
- Merolli, A.; Tranquilli, L.P. Hard tissues structures and functionality. In Biomimetic, Bioresponsive, and Bioactive Materials, 1st; Santin, M., Phillips, G.J., Eds.; Wiley: New York, NY, USA, 2010; in press. [Google Scholar]
- Gamble, J.G. The Musculoskeletal System. Physiological Basics; Raven Press: New York, NY, USA, 1988; p. 50. [Google Scholar]
- Arana-Chavez, V.E.; Massa, L.F. Odontoblasts: The cells forming and maintaining dentine. Int. J. Biochem. Cell Biol. 2004, 36, 1367–1373. [Google Scholar] [CrossRef]
- Wolff, J. The Law of Bone Remodelling; Springer-Verlag: Berlin, Heidelberg, Germany, 1986. [Google Scholar]
- Pauwels, F. Developmental effects of the functional adaptation of bone. Anat. Anz. 1976, 139, 213–220. [Google Scholar]
- Carter, D.R.; Orr, T.E. Skeletal development and bone functional adaptation. J. Bone Miner. Res. 1992, 7 (Suppl. 2), S389–S395. [Google Scholar]
- Ulstrup, A.K. Biomechanical concepts of fracture healing in weight-bearing long bones. Acta Orthop. Belg. 2008, 74, 291–302. [Google Scholar]
- Schindeler, A.; McDonald, M.M.; Bokko, P.; Little, D.G. Bone remodeling during fracture repair: The cellular picture. Semin. Cell Dev. Biol. 2008, 19, 459–466. [Google Scholar] [CrossRef]
- Isaksson, H.; Gröngröft, I.; Wilson, W.; van Donkelaar, C.C.; van Rietbergen, B.; Tami, A.; Huiskes, R.; Ito, K. Remodeling of fracture callus in mice is consistent with mechanical loading and bone remodeling theory. J. Orthop. Res. 2009, 27, 664–672. [Google Scholar] [CrossRef]
- Lauterburg, M.T.; Exner, G.U.; Jacob, H.A. Forces involved in lower limb lengthening: An in vivo biomechanical study. J. Orthop. Res. 2006, 24, 1815–22. [Google Scholar] [CrossRef]
- Van Roermund, P.M.; ter Haar Romeny, B.M.; Hoekstra, A.; Schoonderwoert, G.J.; Brandt, C.J.; van der Steen, S.P.; Roelofs, J.M.; Scholten, F.; Visser, W.J.; Renooij, W. Bone growth and remodeling after distraction epiphysiolysis of the proximal tibia of the rabbit. Effect of electromagnetic stimulation. Clin. Orthop. Relat. Res. 1991, 266, 304–312. [Google Scholar]
- Gamble, J.G. The Musculoskeletal System. Physiological Basics. Chapter 4: Structural Components; Raven Press: New York, NY, USA, 1988; pp. 57–80. [Google Scholar]
- Anderson, H.C. Vesicles associated with calcification in the matrix of epiphyseal cartilage. J. Cell Biol. 1969, 41, 59–72. [Google Scholar] [CrossRef]
- Anderson, H.C. Matrix vesicles and calcification. Curr. Rheumatol. Rep. 2003, 5, 222–226. [Google Scholar] [CrossRef]
- Anderson, H.C.; Garimella, R.; Tague, S.E. The role of matrix vesicles in growth plate development and biomineralization. Front. Biosci. 2005, 10, 822–837. [Google Scholar] [CrossRef]
- Anderson, H.C.; Clarke, H. The role of matrix vesicles in physiological and pathological calcification. Curr. Opin. Orthop. 2007, 18, 428–433. [Google Scholar] [CrossRef]
- Ali, S.Y.; Sajdera, S.W.; Anderson, H.C. Isolation and characterization of calcifying matrix vesicles from epiphyseal cartilage. Proc. Natl. Acad. Sci. USA 1970, 67, 1513–1520. [Google Scholar] [CrossRef]
- Thyberg, J.; Friberg, U. Ultrastructure and acid phosphatase activity of matrix vesicles and cytoplasmic dense bodies in the epiphyseal plate. J. Ultrastruct. Res. 1970, 33, 554–573. [Google Scholar] [CrossRef]
- Ali, S.Y.; Anderson, H.C.; Sajdera, S.W. Enzymic and electron-microscopic analysis of extracellular matrix vesicles associated with calcification in cartilage. Biochem. J. 1971, 122, 56P. [Google Scholar]
- Thyberg, J.; Friberg, U. Electron microscopic enzyme histochemical studies on the cellular genesis of matrix vesicles in the epiphyseal plate. J. Ultrastruct. Res. 1972, 41, 43–59. [Google Scholar] [CrossRef]
- Anderson, H.C.; Reynolds, J.J. Pyrophosphate stimulation of calcium uptake into cultured embryonic bones. Fine structure of matrix vesicles and their role in calcification. Dev. Biol. 1973, 34, 211–227. [Google Scholar]
- Anderson, H.C. Calcifying matrix vesicles of skeletal tissues. Annu. Meet. Am. Inst. Oral. Biol. 1974, 2–12. [Google Scholar]
- Bonucci, E.; Gherardi, G. Histochemical and electron microscopy investigations on medullary bone. Cell Tissue Res. 1975, 163, 81–97. [Google Scholar]
- Rabinovitch, A.L.; Anderson, H.C. Biogenesis of matrix vesicles in cartilage growth plates. Fed. Proc. 1976, 35, 112–116. [Google Scholar]
- Bonucci, E.; Dearden, L.C. Matrix vesicles in aging cartilage. Fed. Proc. 1976, 35, 163–168. [Google Scholar]
- Koenig, H. The soluble acidic lipoproteins (SALPS) of storage granules. Matrix constituents which may bind stored molecules. Adv. Cytopharmacol. 1974, 2, 273–301. [Google Scholar]
- Peress, N.S.; Anderson, H.C.; Sajdera, S.W. The lipids of matrix vesicles from bovine fetal epiphyseal cartilage. Calcif. Tissue Res. 1974, 14, 275–81. [Google Scholar] [CrossRef]
- Wuthier, R.E. Lipid composition of isolated epiphyseal cartilage cells, membranes and matrix vesicles. Biochim. Biophys. Acta 1975, 409, 128–143. [Google Scholar] [CrossRef]
- Wuthier, R.E. Effect of phospholipids on the transformation of amorphous calcium phosphate to hydroxapatite in vitro. Calcif. Tissue Res. 1975, 19, 197–210. [Google Scholar] [CrossRef]
- Irving, J.T. Interrelations of matrix lipids, vesicles, and calcification. Fed. Proc. 1976, 35, 109–111. [Google Scholar]
- Wuthier, R.E. Lipids of matrix vesicles. Fed. Proc. 1976, 35, 117–121. [Google Scholar]
- Vogel, J.J.; Boyan-Salyers, B.D. Acidic lipids associated with the local mechanism of calcificaiton: A review. Clin. Orthop. Relat. Res. 1976, 118, 231–241. [Google Scholar]
- Boskey, A.L.; Posner, A.S. Effect of magnesium on lipid-induced calcification: An in vitro model for bone mineralization. Calcif. Tissue Int. 1980, 32, 139–143. [Google Scholar] [CrossRef]
- Wu, L.N.; Genge, B.R.; Kang, M.W.; Arsenault, A.L.; Wuthier, R.E. Changes in phospholipid extractability and composition accompany mineralization of chicken growth plate cartilage matrix vesicles. J. Biol. Chem. 2002, 277, 5126–5133. [Google Scholar]
- Bonucci, E. Electron microscope studies of the early stage of the calcification process: Role of matrix vesicles. Prog. Clin. Biol. Res. 1989, 295, 109–114. [Google Scholar]
- Bonucci, E.; Silvestrini, G.; Mocetti, P. MC22-33F monoclonal antibody shows unmasked polar head groups of choline-containing phospholipids in cartilage and bone. Eur. J. Histochem. 1997, 41, 177–190. [Google Scholar]
- Thouverey, C.; Strzelecka-Kiliszek, A.; Balcerzak, M.; Buchet, R.; Pikula, S. Matrix vesicles originate from apical membrane microvilli of mineralizing osteoblast-like Saos-2 cells. J. Cell. Biochem. 2009, 106, 127–138. [Google Scholar] [CrossRef]
- Ennever, J.; Vogel, J.J.; Riggan, L.J. Phospholipids of a bone matrix calcification nucleator. J. Dent. Res. 1978, 57, 731–734. [Google Scholar] [CrossRef]
- Stewart, P.J.; Stern, P.H. Calcium/phosphatidylserine-stimulated protein phosphorylation in bone: Effect of parathyroid hormone. J. Bone Miner. Res. 1987, 2, 281–287. [Google Scholar] [CrossRef]
- Schwartz, Z.; Sela, J.; Ramirez, V.; Amir, D.; Boyan, B.D. Changes in extracellular matrix vesicles during healing of rat tibial bone: A morphometric and biochemical study. Bone 1989, 10, 53–60. [Google Scholar] [CrossRef]
- Genge, B.R.; Wu, L.N.; Wuthier, R.E. Identification of phospholipid-dependent calcium-binding proteins as constituents of matrix vesicles. J. Biol. Chem. 1989, 264, 10917–10921. [Google Scholar]
- Genge, B.R.; Wu, L.N.; Wuthier, R.E. Differential fractionation of matrix vesicle proteins. Further characterization of the acidic phospholipid-dependent Ca2(+)-binding proteins. J. Biol. Chem. 1990, 265, 4703–4710. [Google Scholar]
- Genge, B.R.; Wu, L.N.; Adkisson, H.D., 4th; Wuthier, R.E. Matrix vesicle annexins exhibit proteolipid-like properties. Selective partitioning into lipophilic solvents under acidic conditions. J. Biol. Chem. 1991, 266, 10678–10685. [Google Scholar]
- Genge, B.R.; Cao, X.; Wu, L.N.; Buzzi, W.R.; Showman, R.W.; Arsenault, A.L.; Ishikawa, Y.; Wuthier, R.E. Establishment of the primary structure of the major lipid-dependent Ca2+ binding proteins of chicken growth plate cartilage matrix vesicles: Identity with anchorin CII (annexin V) and annexin II. J. Bone Miner. Res. 1992, 7, 807–819. [Google Scholar]
- Wu, L.N.; Genge, B.R.; Dunkelberger, D.G.; LeGeros, R.Z.; Concannon, B.; Wuthier, R.E. Physicochemical characterization of the nucleational core of matrix vesicles. J. Biol. Chem. 1997, 272, 4404–4411. [Google Scholar]
- Von der Mark, K.; Mollenhauer, J. Annexin V interactions with collagen. Cell Mol. Life Sci. 1997, 53, 539–545. [Google Scholar] [CrossRef]
- Kirsch, T.; Nah, H.D.; Demuth, D.R.; Harrison, G.; Golub, E.E.; Adams, S.L.; Pacifici, M. Annexin V-mediated calcium flux across membranes is dependent on the lipid composition: Implications for cartilage mineralization. Biochemistry 1997, 36, 3359–3367. [Google Scholar]
- Genge, B.R.; Wu, L.N.; Wuthier, R.E. Kinetic analysis of mineral formation during in vitro modeling of matrix vesicle mineralization: Effect of annexin A5, phosphatidylserine, and type II collagen. Anal. Biochem. 2007, 367, 159–66. [Google Scholar] [CrossRef]
- Genge, B.R.; Wu, L.N.; Wuthier, R.E. In vitro modeling of matrix vesicle nucleation: Synergistic stimulation of mineral formation by annexin A5 and phosphatidylserine. J. Biol. Chem. 2007, 282, 26035–26045. [Google Scholar] [CrossRef]
- Genge, B.R.; Wu, L.N.; Wuthier, R.E. Mineralization of annexin-5-containing lipid-calcium-phosphate complexes: Modulation by varying lipid composition and incubation with cartilage collagens. J. Biol. Chem. 2008, 283, 9737–9748. [Google Scholar]
- Wu, L.N.; Genge, B.R.; Wuthier, R.E. Differential effects of zinc and magnesium ions on mineralization activity of phosphatidylserine calcium phosphate complexes. J. Inorg. Biochem. 2009, 103, 948–62. [Google Scholar] [CrossRef]
- Borisenko, G.G.; Iverson, S.L.; Ahlberg, S.; Kagan, V.E.; Fadeel, B. Milk fat globule epidermal growth factor 8 (MFG-E8) binds to oxidized phosphatidylserine: Implications for macrophage clearance of apoptotic cells. Cell Death Differ. 2004, 11, 943–945. [Google Scholar] [CrossRef]
- Asano, K.; Miwa, M.; Miwa, K.; Hanayama, R.; Nagase, H.; Nagata, S.; Tanaka, M. Masking of phosphatidylserine inhibits apoptotic cell engulfment and induces autoantibody production in mice. J. Exp. Med. 2004, 200, 459–467. [Google Scholar] [CrossRef]
- Yoshida, H.; Kawane, K.; Koike, M.; Mori, Y.; Uchiyama, Y.; Nagata, S. Phosphatidylserine-dependent engulfment by macrophages of nuclei from erythroid precursor cells. Nature 2005, 437, 754–758. [Google Scholar]
- Wu, Y.; Tibrewal, N.; Birge, R.B. Phosphatidylserine recognition by phagocytes: A view to a kill. Trends Cell Biol. 2006, 16, 189–197. [Google Scholar] [CrossRef]
- Nakano, T.; Ishimoto, Y.; Kishino, J.; Umeda, M.; Inoue, K.; Nagata, K.; Ohashi, K.; Mizuno, K.; Arita, H. Cell adhesion to phosphatidylserine mediated by a product of growth arrest-specific gene 6. J. Biol. Chem. 1997, 272, 29411–29414. [Google Scholar]
- Ishimoto, Y.; Ohashi, K.; Mizuno, K.; Nakano, T. Promotion of the uptake of PS liposomes and apoptotic cells by a product of growth arrest-specific gene, gas6. J. Biochem. 2000, 127, 411–417. [Google Scholar] [CrossRef]
- Rajotte, I.; Hasanbasic, I.; Blostein, M. Gas6-mediated signaling is dependent on the engagement of its gamma-carboxyglutamic acid domain with phosphatidylserine. Biochem. Biophys. Res. Commun. 2008, 376, 70–73. [Google Scholar] [CrossRef]
- Hanayama, R.; Tanaka, M.; Miwa, K.; Nagata, S. Expression of developmental endothelial locus-1 in a subset of macrophages for engulfment of apoptotic cells. J. Immunol. 2004, 172, 3876–3882. [Google Scholar]
- Anderson, H.A.; Maylock, C.A.; Williams, J.A.; Paweletz, C.P.; Shu, H.; Shacter, E. Serum-derived protein S binds to phosphatidylserine and stimulates the phagocytosis of apoptotic cells. Nat. Immunol. 2003, 4, 87–91. [Google Scholar]
- Uehara, H.; Shacter, E. Auto-oxidation and oligomerization of protein S on the apoptotic cell surface is required for Mer tyrosine kinase-mediated phagocytosis of apoptotic cells. J. Immunol. 2008, 180, 2522–2530. [Google Scholar]
- Savill, J.; Gregory, C. Apoptotic PS to phagocyte TIM-4: Eat me. Immunity 2007, 27, 830–832. [Google Scholar] [CrossRef]
- Kobayashi, N.; Karisola, P.; Pena-Cruz, V.; Dorfman, D.M.; Jinushi, M.; Umetsu, S.E.; Butte, M.J.; Nagumo, H.; Chernova, I.; Zhu, B.; Sharpe, A.H.; Ito, S.; Dranoff, G.; Kaplan, G.G.; Casasnovas, J.M.; Umetsu, D.T.; Dekruyff, R.H.; Freeman, G.J. TIM-1 and TIM-4 glycoproteins bind phosphatidylserine and mediate uptake of apoptotic cells. Immunity 2007, 27, 927–940. [Google Scholar] [CrossRef]
- Santiago, C.; Ballesteros, A.; Martinez-Munoz, L.; Mellado, M.; Kaplan, G.G.; Freeman, G.J.; Casasnovas, J.M. Structures of T cell immunoglobulin mucin protein 4 show a metal-Ion-dependent ligand binding site where phosphatidylserine binds. Immunity 2007, 27, 941–951. [Google Scholar] [CrossRef]
- Miyanishi, M.; Tada, K.; Koike, M.; Uchiyama, Y.; Kitamura, T.; Nagata, S. Identification of Tim4 as a phosphatidylserine receptor. Nature 2007, 450, 435–439. [Google Scholar]
- Park, D.; Hochreiter-Hufford, A.; Ravichandran, KS. The phosphatidylserine receptor TIM-4 does not mediate direct signaling. Curr. Biol. 2009, 19, 346–351. [Google Scholar] [CrossRef]
- Braun, G.; Kohavi, D.; Amir, D.; Luna, M.; Caloss, R.; Sela, J.; Dean, D.D.; Boyan, B.D.; Schwartz, Z. Markers of primary mineralization are correlated with bone-bonding ability of titanium or stainless steel in vivo. Clin. Oral. Implants Res. 1995, 6, 1–13. [Google Scholar]
- Taylor, M.G.; Simkiss, K.; Simmons, J.; Wu, L.N.; Wuthier, R.E. Structural studies of a phosphatidyl serine-amorphous calcium phosphate complex. Cell Mol. Life Sci. 1998, 54, 196–202. [Google Scholar] [CrossRef]
- Sela, J.; Gross, U.M.; Kohavi, D.; Shani, J.; Dean, D.D.; Boyan, B.D.; Schwartz, Z. Primary mineralization at the surfaces of implants. Crit. Rev. Oral Biol. Med. 2000, 11, 423–436. [Google Scholar] [CrossRef]
- Huang, J.S.; Wu, H.F.; Hong, J.M.; Chang, F.H.; Hong, K. Effect of liposomes on mineralization in rat osteoblast-enriched cultures. Kaohsiung J. Med. Sci. 1999, 15, 187–194. [Google Scholar]
- Camolezi, F.L.; Daghastanli, K.R.; Magalhaes, P.P.; Pizauro, J.M.; Ciancaglini, P. Construction of an alkaline phosphatase-liposome system: A tool for biomineralization study. Int. J. Biochem. Cell Biol. 2002, 34, 1091–1101. [Google Scholar] [CrossRef]
- Santin, M.; Rhys-Williams, W.; O'Reilly, J.; Davies, M.C.; Shakesheff, K.; Love, W.G.; Lloyd, A.W.; Denyer, S.P. Calcium-binding phospholipids as a coating material for implant osteointegration. J. R. Soc. Interface 2006, 3, 277–281. [Google Scholar] [CrossRef]
- Satsangi, A.; Satsangi, N.; Glover, R.; Satsangi, R.K.; Ong, J.L. Osteoblast response to phospholipid modified titanium surface. Biomaterials 2003, 24, 4585–4589. [Google Scholar] [CrossRef]
- Satsangi, N.; Satsangi, A.; Glover, R.; Ong, J.L.; Satsangi, R.K. Osteoblast response and calcium deposition on phospholipid modified surfaces. J. Mater. Sci. Mater. Med. 2004, 15, 693–697. [Google Scholar]
- Susin, C.; Qahash, M.; Hall, J.; Sennerby, L.; Wikesj, U.M. Histological and biomechanical evaluation of phosphorylcholine-coated titanium implants. J. Clin. Periodontol. 2008, 35, 270–275. [Google Scholar] [CrossRef]
- Bosetti, M.; Lloyd, A.W.; Santin, M.; Denyer, S.P.; Cannas, M. Effects of phosphatidylserine coatings on titanium on inflammatory cells and cell-induced mineralisation in vitro. Biomaterials 2005, 26, 7572–7578. [Google Scholar]
- Bosetti, M.; Santin, M.; Lloyd, A.W.; Denyer, S.P.; Sabbatini, M.; Cannas, M. Cell behaviour on phospholipids-coated surfaces. J. Mater. Sci. Mater. Med. 2007, 18, 611–617. [Google Scholar]
- Ducheyne, P.; Healy, K.E. The effect of plasma-sprayed calcium phosphate ceramic coatings on the metal ion release from porous titanium and cobalt-chromium alloys. J. Biomed. Mater. Res. 1988, 22, 1137–1163. [Google Scholar] [CrossRef]
- Tranquilli Leali, P.; Merolli, A.; Palmacci, O.; Gabbi, C.; Cacchioni, A.; Gonizzi, G. Evaluation of different preparations of plasma-spray hydroxyapatite coating on titanium alloy and duplex stainless steel in the rabbit. J. Mater. Sci. Mater. Med. 1994, 5, 345–349. [Google Scholar] [CrossRef]
- Williams, D. Biomimetic surfaces: How man-made becomes man-like. Med. Device. Technol. 1995, 6, 6–8. [Google Scholar]
- Tudor, A.M.; Melia, C.D.; Davies, M.C.; Anderson, D.; Hastings, G.W.; Santos, J.D.; Monteiro, F. The analysis of biomedical hydroxyapatite powders and hydroxyapatite coatings on metallic implants by near-IR Fourier transform Raman Spectroscopy. Spectrochimica Acta 1993, 49A, 675–680. [Google Scholar]
- Merolli, A.; Moroni, A.; Faldini, C.; Tranquilli Leali, P.; Giannini, S. Histomorphological study of bone response to hydroxyapatite coating on stainless steel. J. Mater. Sci. Mater. Med. 2003, 14, 327–333. [Google Scholar] [CrossRef]
- Merolli, A.; Borsetti, M.; Giannotta, L.; Lloyd, A.W.; Denyer, S.P.; Rhys-Williams, W.; Love, W.G.; Gabbi, C.; Cacchioni, A.; Tranquilli Leali, P.; Cannas, M.; Santin, M. In vivo assessment of the osteointegrative potential of phosphatidylserine-based coatings. J. Mater. Sci. Mater. Med. 2006, 17, 789–794. [Google Scholar]
- Xu, C.; Wang, Y.; Yu, X.; Chen, X.; Li, X.; Yang, X.; Li, S.; Zhang, X.; Xiang, A.P. Evaluation of human mesenchymal stem cells response to biomimetic bioglass-collagen-hyaluronic acid-phosphatidylserine composite scaffolds for bone tissue engineering. J. Biomed. Mater. Res. A 2009, 88, 264–273. [Google Scholar]
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Merolli, A.; Santin, M. Role of Phosphatidyl-Serine in Bone Repair and Its Technological Exploitation. Molecules 2009, 14, 5367-5381. https://doi.org/10.3390/molecules14125367
Merolli A, Santin M. Role of Phosphatidyl-Serine in Bone Repair and Its Technological Exploitation. Molecules. 2009; 14(12):5367-5381. https://doi.org/10.3390/molecules14125367
Chicago/Turabian StyleMerolli, Antonio, and Matteo Santin. 2009. "Role of Phosphatidyl-Serine in Bone Repair and Its Technological Exploitation" Molecules 14, no. 12: 5367-5381. https://doi.org/10.3390/molecules14125367