
Neurotrophins and Neuropathic Pain: Role in Pathobiology



Center for Integrated Preclinical Drug Development, The University of Queensland, St Lucia Campus, Brisbane, Queensland 4072, Australia
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Molecules 2015, 20(6), 10657-10688; https://doi.org/10.3390/molecules200610657
Submission received: 21 April 2015
/
Accepted: 3 June 2015
/
Published: 9 June 2015
Abstract
:Neurotrophins (NTs) belong to a family of trophic factors that regulate the survival, growth and programmed cell death of neurons. In mammals, there are four structurally and functionally related NT proteins, viz. nerve growth factor (NGF), brain-derived neurotrophic factor (BDNF), neurotrophin 3 and neurotrophin 4. Most research on NTs to date has focussed on the effects of NGF and BDNF signalling via their respective cognate high affinity neurotrophic tyrosine kinase viz TrkA and TrkB receptors. Apart from the key physiologic roles of NGF and BDNF in peripheral and central nervous system function, NGF and BDNF signalling via TrkA and TrkB receptors respectively have been implicated in mechanisms underpinning neuropathic pain. Additionally, NGF and BDNF signalling via the low-affinity pan neurotrophin receptor at 75 kDa (p75NTR) may also contribute to the pathobiology of neuropathic pain. In this review, we critically assess the role of neurotrophins signalling via their cognate high affinity receptors as well as the low affinity p75NTR in the pathophysiology of peripheral neuropathic and central neuropathic pain. We also identify knowledge gaps to guide future research aimed at generating novel insight on how to optimally modulate NT signalling for discovery of novel therapeutics to improve neuropathic pain relief.
1. Introduction
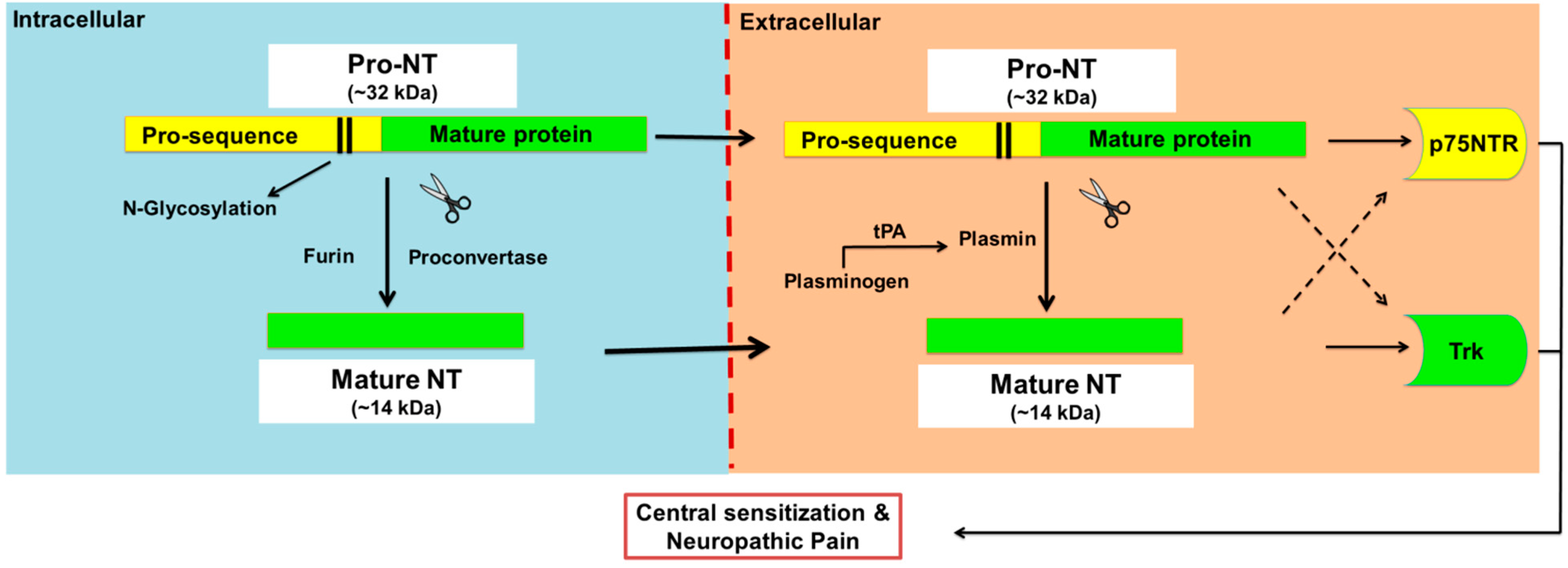
Neurotrophins (NTs) are a family of four structurally and functionally related proteins that regulate the growth, maintenance and apoptosis of neurons in the developing nervous system as well as injured neurons [1,2,3]. In mammals, there are four NTs, viz. nerve growth factor (NGF), brain-derived neurotrophic factor (BDNF), neurotrophin 3 (NT-3) and neurotrophin 4 with the latter also known as neurotrophin-5 (NT-4 or NT4/5) [1,2]. Initially, NTs are synthesised as precursor molecules called proneurotrophins (pro-NTs) at ~30–34 kDa, that undergo proteolytic cleavage in the endoplasmic reticulum and Golgi apparatus to produce C-terminally mature neurotrophins at ~13 kDa [4] (Figure 1).
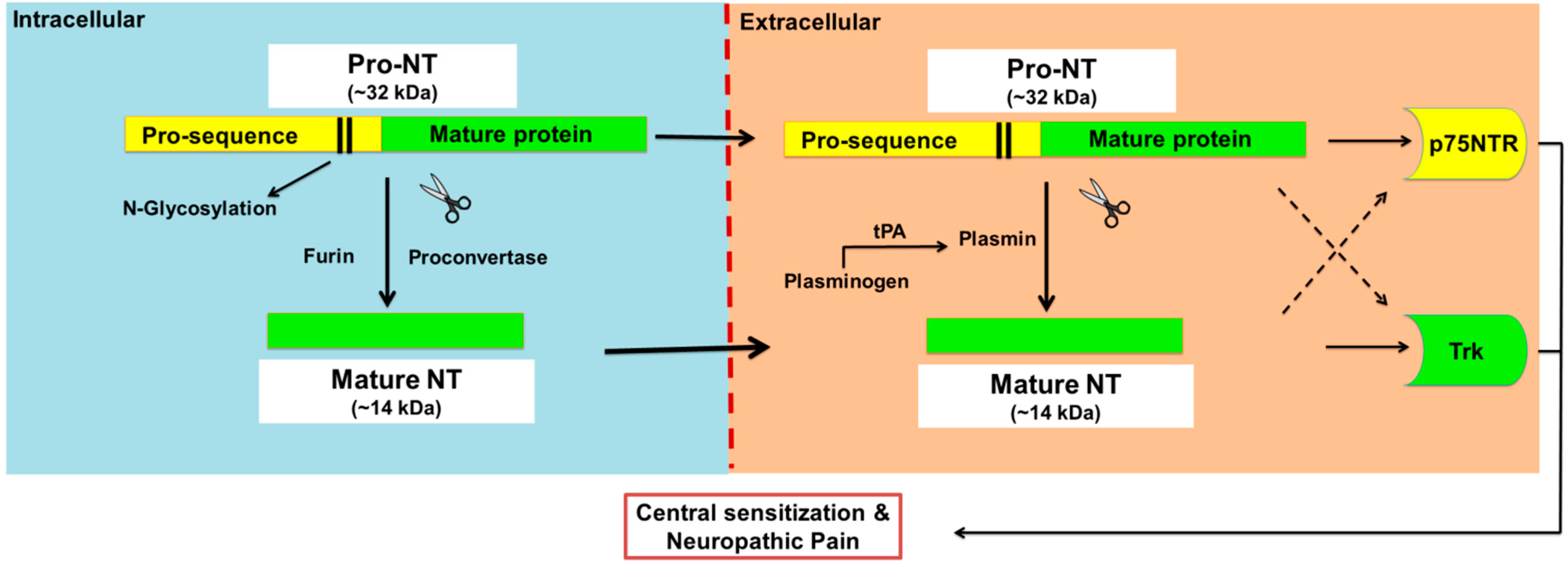
Figure 1.
Proteolytic cleavage of pro-neurotrophins to mature neurotrophins. Neurotrophins (NTs) are synthesised as glycosylated precursors of ~32 kDa, called pro-neurotrophins, pro-NTs. These precursor molecules are cleaved intracellularly by furin or proconvertase, and extracellularly by the action of plasmin to generate mature NTs. Plasmin is a serine protease enzyme that is formed from plasminogen by several enzymes including tissue plasminogen activator (tPA) and urokinase. Intact pro-NTs may also be released extracellularly and evoke direct biological activities that in general oppose that of the mature NT counterpart although there are exceptions. For example, pro-NTs may induce apoptosis whereas mature NTs are mainly associated with neuronal survival. However, both pro- and mature-NTs appear to mediate central sensitisation and neuropathic pain.
Figure 1.
Proteolytic cleavage of pro-neurotrophins to mature neurotrophins. Neurotrophins (NTs) are synthesised as glycosylated precursors of ~32 kDa, called pro-neurotrophins, pro-NTs. These precursor molecules are cleaved intracellularly by furin or proconvertase, and extracellularly by the action of plasmin to generate mature NTs. Plasmin is a serine protease enzyme that is formed from plasminogen by several enzymes including tissue plasminogen activator (tPA) and urokinase. Intact pro-NTs may also be released extracellularly and evoke direct biological activities that in general oppose that of the mature NT counterpart although there are exceptions. For example, pro-NTs may induce apoptosis whereas mature NTs are mainly associated with neuronal survival. However, both pro- and mature-NTs appear to mediate central sensitisation and neuropathic pain.
The biological effects of mature NTs are mediated via two major receptor types, viz neurotrophic tyrosine kinase (Trks) receptors and the pan neurotrophin receptor at 75 kDa (p75NTR) (Patapoutian and Reichardt, 2001). Each mature NT binds with high affinity to a specific Trk receptor. Specifically, NGF binds with high affinity to TrkA, BDNF and NT-4 bind with high affinity to TrkB, and NT-3 binds with high affinity to TrkC [5]. Apart from activation of TrkC, NT-3 also activates TrkA and TrkB, albeit with lower affinities [6] and all four mature NTs bind with similar affinity to the p75NTR [5] (Figure 2). By comparison, the biological actions of pro-NTs are relatively poorly understood [7]. Intact pro-NTs are high affinity ligands at both the p75NTR and the structurally distinct co-receptor, sortilin, in contrast to mature NTs [4,8]. Additionally, pro-NGF and pro-BDNF bind to TrkA [9] and TrkB [10] respectively, albeit with lower affinity than to the p75NTR [10,11]. This complexity of endogenous NT signalling has made investigation of potential pathobiological role(s) of pro-NTs particularly challenging.
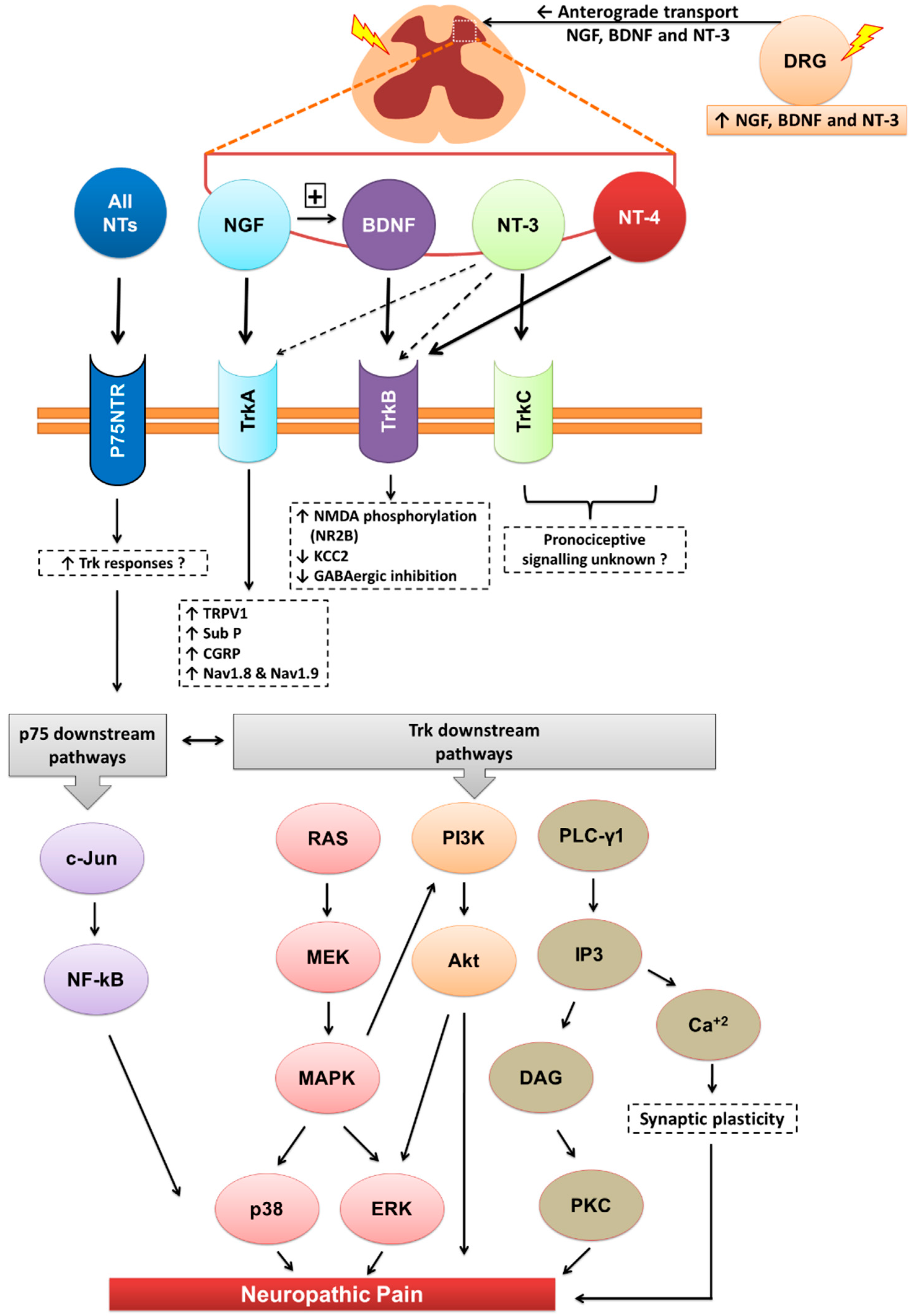
Figure 2.
Downstream signalling of neurotrophins and their receptors. Akt, Akt kinase; Ca+2, calcium ions; CGRP, calcitonin gene related peptide; DAG, diacylglycerol; DRG, dorsal root ganglia; ERK, extracellular-signal-regulated kinase; GABA, γ-Aminobutyric acid; IP3, inositol tris-phosphate; KCC2, potassium chloride cotransporter 2; MAPK, mitogen-activated protein kinases; MEK, mitogen-activated protein kinase/ERK kinase; Nav, sodium-ion voltage-gated channel; NMDA, N-methyl-d-aspartate; NT, neurotrophin; p75NTR, pan neurotrophin receptor at 75kDa; PI3K, phosphatidylinositol 3-kinase; PLC-γ1, phospholipase C-gamma-1; Ras, small GTP-binding protein; Sub-P, substance-P; Trk, tyrosine kinase receptor; TRPV1, transient receptor potential cation channel subfamily V member 1.
Figure 2.
Downstream signalling of neurotrophins and their receptors. Akt, Akt kinase; Ca+2, calcium ions; CGRP, calcitonin gene related peptide; DAG, diacylglycerol; DRG, dorsal root ganglia; ERK, extracellular-signal-regulated kinase; GABA, γ-Aminobutyric acid; IP3, inositol tris-phosphate; KCC2, potassium chloride cotransporter 2; MAPK, mitogen-activated protein kinases; MEK, mitogen-activated protein kinase/ERK kinase; Nav, sodium-ion voltage-gated channel; NMDA, N-methyl-d-aspartate; NT, neurotrophin; p75NTR, pan neurotrophin receptor at 75kDa; PI3K, phosphatidylinositol 3-kinase; PLC-γ1, phospholipase C-gamma-1; Ras, small GTP-binding protein; Sub-P, substance-P; Trk, tyrosine kinase receptor; TRPV1, transient receptor potential cation channel subfamily V member 1.
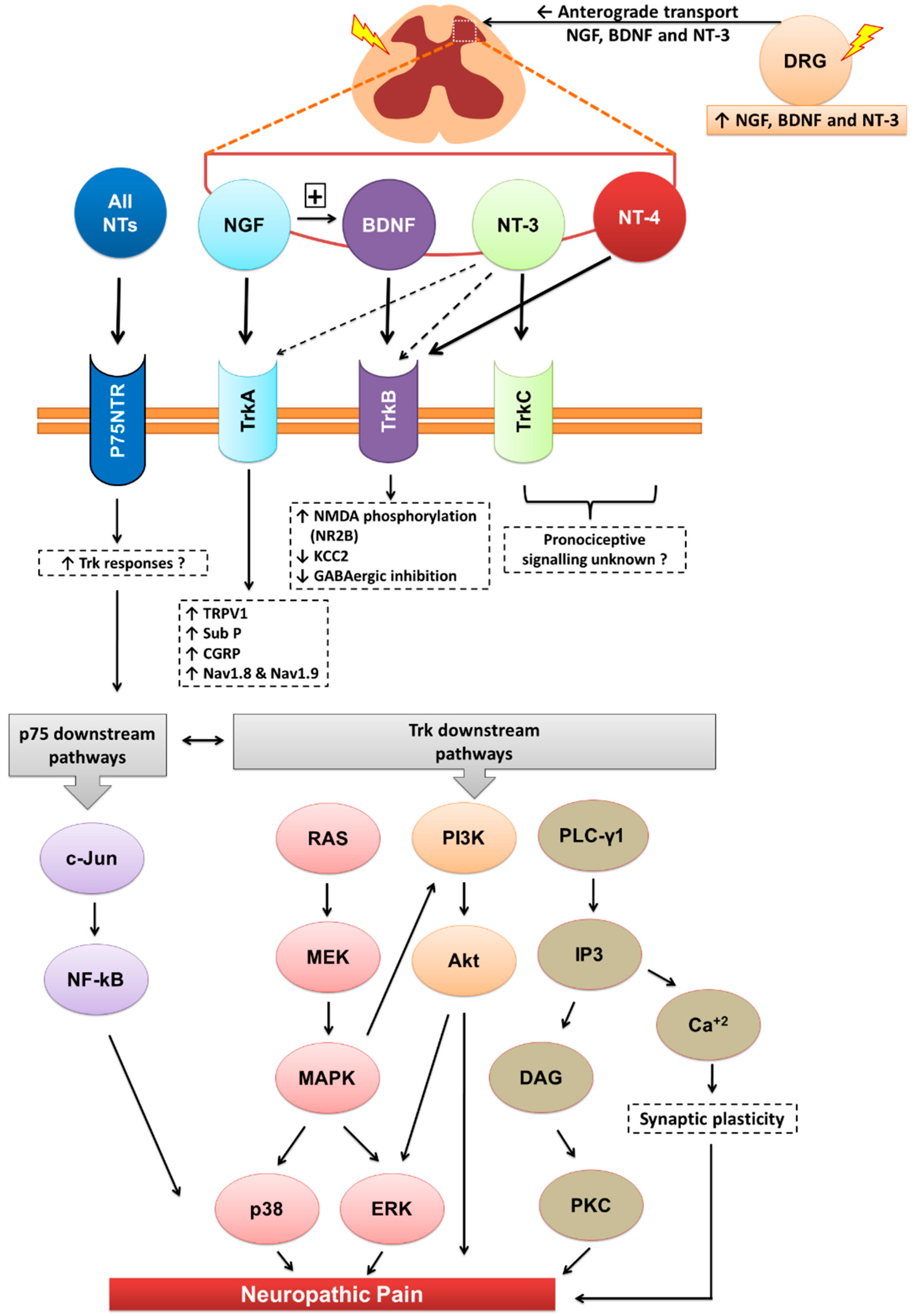
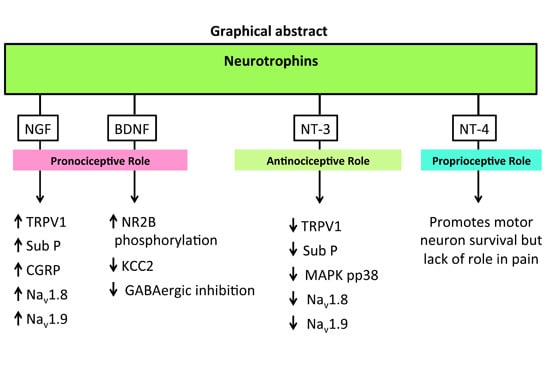
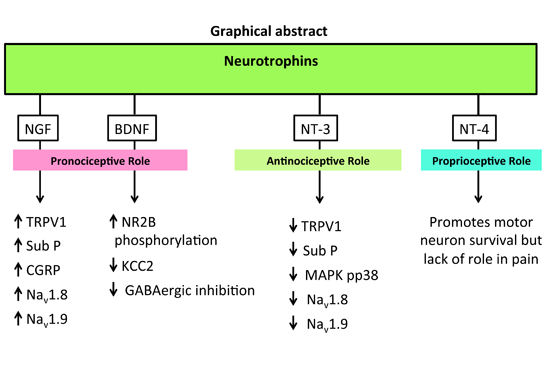
The neurotrophins (NTs) particularly NGF, BDNF and NT-3 are synthesised mainly in the dorsal root ganglia (DRG) with anterograde transported into the dorsal horn of the spinal cord. Apart from their roles in various physiological functions, they modulate central sensitisation in the spinal cord that underpins maintenance of neuropathic pain. NTs bind with high affinity to their respective cognate tryosine receptor kinase (Trk) receptors, namely, NGF with TrkA, BDNF and NT4 with TrkB, and NT3 with TrkC. NT3 may also activate TrkA and TrkB, but with lower affinity. All NTs including their precursors (pro-NTs) may bind to the p75NTR and this is the high affinity receptor for pro-NTs. NGF-TrkA signalling is mostly associated with upregulation of TRPV1 expression and insertion into the nociceptor membrane, substance-P, CGRP and the sodium channels, Nav1.8 and Nav1.9. BDNF-TrkB signalling results in NMDA receptor activation and downregulation of the potassium chloride cotransporter 2 (KCC2) and/or GABAergic inhibitory signalling mechanisms. NT-3 reportedly evokes antinociception whereas NT-4 appears to be predominantly involved in proprioception. Following Trk receptor activation, the downstream signalling mechanisms include the Ras-MAPK, PI3K and PLC-γ1 pathways whereas following activation of the p75NTR, downstream signalling associated with neuropathic pain appears to involve the c-Jun-NF-kB pathway. Additionally, activation of p75NTR may potentiate Trk-mediated signalling in neuropathic pain.
2. Neuropathic Pain
Neuropathic pain is defined by the International Association for the Study of Pain (IASP) as pain that arises as a direct consequence of a lesion or disease affecting the somatosensory system [12]. Neuropathic pain is often poorly alleviated by first- and second-line medications recommended by the Neuropathic Pain Special Interest Group of the IASP due to lack of efficacy and/or dose-limiting side-effects [13,14]. Hence, there is an urgent need to develop novel mechanism-based therapeutic agents that are highly efficacious and well tolerated to improve relief of neuropathic pain [13,14].
3. Neurotrophins and Neuropathic Pain
NTs contribute to the pathogenesis of neuropathic pain as they have key roles in the complex mechanisms that underpin peripheral and central sensitisation [15,16,17]. However, knowledge on the specific contribution of individual neurotrophins signalling via a particular Trk receptor and/or the p75NTR in the pathobiology of neuropathic pain is required to identify the optimal targets for use in discovery programs aimed at producing novel analgesics for improving neuropathic pain relief.
4. Nerve Growth Factor (NGF)
NGF was the first neurotrophin identified as having a key role in the survival and function of sensory and sympathetic neurons in the peripheral nervous system (PNS), as well as basal forebrain cholinergic neurons in the CNS [20,21].

{kind=link}
{kind=link}
{kind=link}
Table 1.
Expression of neurotrophins and their nociceptive role in rodent models of neuropathic pain.
| NTs | Rodent Models of Neuropathic Pain | |||||
|---|---|---|---|---|---|---|
| Tissue | Nerve Ligation/Axotomy(e.g., CCI, SNL) | CIPN | DPN | EAE (MS) | SCI | |
| NGF | DRG | ↑ [22,23] | ↑ [24] or ↓ [25] | ↑ [26] or ↓ [27] | NS | NS |
| SC | ↑ [23] | ↑ [24,28] | ↓ [27] | ↓ [29,30] (Correlation with pain not investigated) | Not changed [31] or ↑ [32] (Indirect evidence) | |
| Pronociception | ||||||
| ● Micro-injected NGF (50 µg) via a catheter into an L5 DRG of un-injured rodents, induced ipsilateral persistent mechanical allodynia [33] | ||||||
| ● Intrathecal (i.t.) infusion of NGF (12 µg/day) for 9 days induced thermal hyperalgesia in rats [34] | ||||||
| ● Intraplantar (i.pl.) injection of NGF (0.3–5 µg) into a hindpaw produced dose-dependent mechanical allodynia and thermal hyperalgesia that persisted for ≥1 week and 24–48 h respectively [35]. Both mechanical allodynia and thermal hyperalgesia were partially attenuated by a TRPV1 antagonist [35] suggesting that NGF-mediated hyperalgesia is driven at least in part by increased TRPV1 expression [35,36] | ||||||
| ● NGF (1.0 mg/kg s.c.) significantly increased Sub-P and CGRP expression levels in the dorsal horn of the spinal cord [37] thereby producing neuropathic pain [38] | ||||||
| ● Overexpression of NGF may induce dysregulation of DRG Na+ channel expression, particularly Nav 1.8 (SNS/PN3), Nav 1.9 (NaN), Nav 1.6, Nav 2.1 (NaG) as well as various Na+ channel subunits including α-I, α-II, β-I and β-II in sensory neurons leading to neuropathic pain symptoms [39,40,41,42 ] | ||||||
| Antinociception | ||||||
| ● I.t. infusion of NGF (125 ng/µL/h) for 7 days reversed mechanical allodynia and thermal hyperalgesia in the hindpaws of CCI-rats. The analgesic effects of NGF were correlated with neuroprotection and decreased astrocytosis [43] | ||||||
| BDNF | DRG | ↑ [44] | NS | ↑ [45] | ↑ [46] (Not investigated in relation with pain) | NS |
| SC | ↑ [44] | ↑ [28] | NS | ↑ [47] | ↑ [48] | |
| Pronociception | ||||||
| ● Microinjection of BDNF (27–270 pg) into the midbrain facilitated nociception dependent on phosphorylation of NMDA receptors [49] | ||||||
| ● Micro-injection of BDNF (50 μg) into an L5 DRG of control (non-injured) rodents induced persistent mechanical allodynia in the hindpaws [33] | ||||||
| ● I.pl. injection of BDNF (200 ng) into rodent hindpaws produced transient thermal hyperalgesia and was significantly less potent (p < 0.05) than a similar dose of NGF [36] | ||||||
| ● In rodent models of peripheral neuropathic pain, upregulated BDNF induced phosphorylation of the NR2B subunit of the NMDA receptor [50] that was accompanied by downregulation of Kv channels [45] as well as expression levels of KCC2 in lamina-I of the spinal cord, thereby disrupting GABAergic inhibition [51] | ||||||
| ● Migration of inflammatory cells into the spinal cord may contribute to upregulation of BDNF in rodent models of peripheral neuropathic pain (e.g., DPN) [52] or CNP (e.g., MS-neuropathic pain) [47] | ||||||
| Antinociception | ||||||
| ● BDNF-infusion (12 µg/day) into the midbrain for 1–11 days evoked antinociception in the tail flick test in rats [18]. The proposed mechanism was via activation of descending opiodergic and serotonergic inhibitory signalling [18] | ||||||
| NT-4 | DRG | NS | NS | |||
| SC | ↓ [53] (Short-term) or Unchanged [54] | |||||
| Lack of a role in neuropathic pain | ||||||
| ● NT-4 appears to have no effect on activity-dependent synaptic plasticity or neuropathic pain [55,56]. | ||||||
| ● In one study, transient thermal hyperalgesia was observed in rats followed by i.pl. injection of NT-4 (200 ng) however it was worn off by 24 h [36] | ||||||
| ● Although there was a significant decrease in NT-4 expression levels in rodent models of neuropathic pain, e.g., DPN (at sciatic nerve) [57] and EAE (at brain) [58], it’s possible role in the pathogenesis of neuropathic pain remains to be investigated. | ||||||
| NT-3 | DRG | ↑ [22] | ↓ [59] (Indirect evidence) | ↓ [60] | NS | NS |
| SC | ↑ [61] | NS | NS | ↑ [62] (Not investigated in relation with pain) | ↑ [63] or ↓ [31] (Not investigated in relation with pain) | |
| Pronociception | ||||||
| ● Micro-injection of NT-3 (50 µg) into the L5 DRGs of control non-injured rodents produced transient mechanical allodynia in the hindpaws [33] | ||||||
| ● I.t infusion of NT-3 at 200 ng/day for 20-days produced pronounced but delayed mechanical allodynia in the hindpaws of non-injured rats at days 10–20 after dose initiation [64] | ||||||
| Antinociception | ||||||
| ● NT-3 infusion (12 µg/day) into the midbrain for 1–11 days showed delayed but stable antinociception in the tail-flick test in rats [18] | ||||||
| ● I.t. administration of NT-3 (600 ng/μL/h) for 7 days suppressed the over-expression of TRPV1 channels, p38 MAPK and Na+ channels (Nav 1.8 and Nav 1.9) in the ipsilateral DRGs of CCI-rats [65,66] | ||||||
| ● Down-regulation of Kv channel gene expression in DRG neurons following sciatic nerve transection was reversed by ex vivo incubation of DRGs collected from nerve-injured rats, with NT-3 (100 ng/mL) [67] | ||||||
| ● Acute i.p. injection of NT-3 (10–20 mg/kg) evoked transient mechanical but not thermal hypoalgesia in the hindpaws of rats that appeared to be underpinned by inhibition of SP release in the spinal cord [68] | ||||||
| ● NT-3 (200 ng) injected locally into rodent hindpaws did not produce hyperalgesia in contrast to that evoked by either NGF or BDNF [36] | ||||||
BDNF, brain derived neurotrophic factor; CCI, chronic-constriction nerve injury; CGRP, calcitonin gene related peptide; CIPN, chemotherapy-induced peripheral neuropathy; DPN, diabetic peripheral neuropathy; DRG, dorsal root ganglia; EAE, experimental autoimmune encephalomyelitis; GABA, γ-Aminobutyric acid; h, h; I.pl, intraplantar; i.t, intrathecal; KCC2, potassium-chloride co-transporter; Kv, potassium voltage-gated channel; MAPK, mitogen-activated protein kinases; MS, multiple sclerosis; mg, milligram; mL, millilitre; ng, nanogram; Na+, sodium ion; Nav, sodium voltage-gated channel; NGF, nerve growth factor; NMDA, N-methyl-d-aspartate receptor; NS, not studied; NT, neurotrophins; s.c, subcutaneous; SC, spinal cord; SCI, spinal cord injury; SNL, spinal nerve ligation; Sub-P, substance-P; TRPV1, transient receptor potential cation channel subfamily V member 1; µg, microgram. µL, microliter; ↑, upregulation; ↓ downregulation.
5. Pronociceptive Effects of NGF in Rodent Models
Following its local or systemic administration in rodents and humans, NGF produced non-inflammatory and long-lasting thermal and mechanical hyperalgesia, with these two pain behaviours mediated by distinct mechanisms [35,69,70,71]. Specifically, NGF-induced thermal hyperalgesia appeared to be underpinned by sensitisation of peripheral nociceptors whereas NGF-induced mechanical hyperalgesia was predominantly mediated by complex signalling in the spinal cord [69,72]. Cross-talk between sensory and adrenergic inputs from sympathetic neurons due to abnormal sprouting of axons from sensory and sympathetic DRG neurons to form baskets around large diameter neurons has also been implicated in the pathogenesis of peripheral neuropathic pain in rodents [22,73,74,75,76].
NGF has been shown to dynamically regulate the synthesis of multiple neurotransmitters and neuropeptides in sensory and sympathetic neurons [20]. These included norepinephrine (noradrenaline) in sympathetic neurons via selective induction of the key enzyme, tyrosine hydroxylase (TH) [77] and the pronociceptive neuropeptides, substance P (SP) and calcitonin gene-related peptide (CGRP) in sensory neurons in the dorsal root ganglia (DRGs) and spinal cord [78].
In rats, observations that localised hyperalgesia induced by intraplantar (i.pl.) NGF was attenuated partially by oral pre-treatment with a transient receptor potential vanilloid 1 (TRPV1) antagonist, implicated a role for TRPV1 at least in part, in mediating this pain behaviour [35]. These findings supported earlier work whereby upregulated NGF-TrkA signalling phosphorylated (activated) TRPV1 and promoted its insertion into the nociceptor cell membrane that in turn induced long-term hyperexcitability of primary afferent sensory nerve fibres [79]. In other work, a critical role for NGF-p75NTR signalling has been implicated in sensory nerve fibre hyperexitability and development of mechanical hyperalgesia induced by i.pl. NGF injection [80,81].
In primary sensory neurons, the cannabinoid CB1 receptor is co-localised with substance P, CGRP [82,83] and TRPV1 [83,84], expression levels of which are markedly increased in the cell membrane by retrograde transport of NGF to the cell bodies in the DRGs. In cultured neurons, CB1 receptor activation by cannabinoids attenuated NGF-induced TRPV1 sensitisation [85]. Hence, investigation of the extent to which cannabinoids attenuate NGF-induced TRPV1 upregulation and sensitisation of primary sensory neurons in rodent models of neuropathic pain is warranted. However, the widespread distribution of CB1 receptors in the CNS results in a plethora of cannabinoid-induced side effects including sedation, dependence, motor and cognitive impairments [86,87] that are impediments to the clinical exploitation of cannabinoid inhibition of the pronociceptive effects of NGF to alleviate chronic pain. Interestingly, endocannabinoid expression levels are significantly upregulated both spinally and supraspinally in rodent models of neuropathic pain [86,88,89]. Hence, a strategy for treatment of neuropathic pain worthy of future investigation is administration of a cannabinoid CB2 receptor agonist to down-regulate TRPV1 expression levels in primary sensory neurons and inhibit nociceptive input into the dorsal horn of the spinal cord [87]. This strategy has appeal as CB2 receptor expression is predominantly in peripheral components of the somatosensory system thereby avoiding CNS side effects [87].
6. Antinociceptive Effects of NGF in Rodent Models
Apart from variable reports on expression levels of NGF in the pathobiology of peripheral neuropathic pain conditions, the effect of intrathecal (i.t.) NGF on nociception in rodents is also controversial. For example, in one study thermal hyperalgesia was induced by i.t. NGF administration for 9-days at 12 µg/day [34] whereas in other work, chronic i.t. NGF infusion for 7-days at 125 ng/h induced pain relief in a rat model of mechanically-induced peripheral neuropathic pain [43]. The latter effected appeared to involve reduced reactive gliosis and restoration of homeostatic conditions in the spinal cord [43].
7. NGF: Role in Peripheral Neuropathic Pain
7.1. NGF in Rodent Models of Peripheral Nerve Ligation
In rodent models of peripheral neuropathic pain induced by peripheral nerve ligation, the pronociceptive action of NGF was reversed by an NGF antagonist [90,91,92]. In parallel with these reports, systemic administration of an antibody against the high affinity NGF receptor, TrkA, produced long lasting anti-allodynia (pain relief) in a mouse model of peripheral neuropathic pain induced by loose ligatures tied around a single sciatic nerve (CCI-mice) [93].
7.2. Rodent Models of PDN and CIPN: Controversial Reports of NGF Expression
Two to three decade ago, there were multiple reports of reduced levels of NGF and its regulated neuropeptides (CGRP and SP) in DRG sensory neurons and the spinal dorsal horn in rodent models of painful diabetic neuropathy (PDN) and chemotherapy induced peripheral neuropathy (CIPN), that are peripheral neuropathic pain models [25,28,94,95,96]. In PDN, peripheral neurodegeneration and impaired catecholaminergic neurotransmission were linked to decreased NGF expression [97]. Additionally, incubation of cultured adult rat DRG neurons with chemotherapy agents decreased neurite outgrowth and this was reversed by NGF treatment, suggesting that NGF treatment may be beneficial in CIPN [98]. In support of this notion, there was a correlation between CIPN severity in humans and the decrease in circulating NGF levels [99]. In rodent models of CIPN, NGF treatment reversed the behavioural and biochemical manifestations of pain [25,28,100]. In other work, NGF treatment reversed the downregulated expression of CGRP and SP in the lumbar DRGs of rodent models of PDN and CIPN and alleviated sensory deficits in these animals [25,95,96,100].
Conversely, more recent reports indicate that NGF expression is increased in rodent models of neuropathic pain or pain with a neuropathic component [24,101,102,103,104]. The upregulated levels of NGF appear to be located in Schwann cells and satellite glial cells in close proximity to injured primary sensory neurons [105,106]. NGF may also be released by invading mast cells, eosinophils, lymphocytes and macrophages at the site of peripheral nerve injury [22,107,108,109]. Following internalisation of the NGF-TrkA complex, its retrograde transport to the DRGs induced phenotypic changes in the peripheral and central terminals of sensory nerve fibres [38,110]. This in turn increased synthesis and release of pronociceptive mediators including SP, CGRP [111] and BDNF from TrkA-positive primary afferents [112]. The net effect was development of mechanical and/or thermal hyperalgesia in the hindpaws [112].
8. NGF: Role in Central Neuropathic Pain (CNP)
8.1. NGF in Spinal Cord Injury Induced Neuropathic Pain Rodent Model
Evidence supporting a role for NGF in the pathobiology of various CNP conditions is scarce. Following hemisection of the spinal cord in rats, mechanical and thermal allodynia were induced in the hindpaws [32,113]. These changes were accompanied by elevated levels of CGRP bilaterally in the spinal dorsal horn and by sprouting of CGRP-positive fine sensory afferents from laminae I-II into deeper laminae (III-IV) in the spinal cord [32,113]. A pathobiologic role for NGF in these neuroplastic changes in the spinal cord is supported by observations that i.t. treatment with an anti-NGF antibody for 2-weeks after spinal cord hemisection, prevented CGRP upregulation as well as the sprouting of CGRP-containing primary afferent C- and Aδ-fibres in the spinal cord [32,114].
8.2. NGF in Multiple Sclerosis-Associated Neuropathic Pain Mouse Model
Multiple sclerosis (MS) is an inflammatory demyelinating disease of the CNS where debilitating CNP may occur early in the disease course [115]. In patients with MS, upregulated synthesis of NGF by oligodendrocytes, astrocytes, microglia as well as by infiltrating T-lymphocytes and macrophages [116,117] has been proposed to have a role in the promotion of myelin repair as well as attenuation of neuroinflammation [117,118].
However, spinal dorsal horn expression levels of the pronociceptive peptides, CGRP and galanin, in an EAE mouse model of MS-associated neuropathic pain did not differ significantly from the corresponding levels in sham-mice administered Freund’s Complete Adjuvant (FCA) only [119]. Hence, these findings appeared to discount a role for upregulated NGF in the pathogenesis of MS-induced CNP [119]. However, the sham-mice had received FCA as adjuvant in the EAE-immunisation protocol, which itself has been shown to induce robust neuroinflammation [120,121] and increase CGRP expression [122]. Hence, an FCA-induced increase in CGRP expression levels in the spinal dorsal horn of sham-mice may have masked a similar effect by MOG35–55 in the EAE-mice. Hence, re-examination of this issue using an EAE immunisation protocol that uses an adjuvant (e.g., Quil A) that does not itself produce neuroinflammation [47,123] is warranted to assess the extent to which NGF has a role in the pathobiology of MS-associated neuropathic pain.
9. BDNF
BDNF is another neurotrophic factor implicated in the regulation of pronociceptive signalling in inflammatory and neuropathic pain conditions [124,125]. However, the pathobiologic role of BDNF-TrkB signalling in peripheral neuropathic pain [126], appears to be confined primarily to spinal and supraspinal sites [49,127,128]. This is in contrast to the predominant effects of NGF in the peripheral nervous system (PNS) [102]. Additionally, BDNF-p75NTR signalling, like NGF-p75NTR signalling, may also induce the downstream sphingomyelin-signalling cascade resulting in hyperexitability of small diameter sensory neurons [129].
10. Pronociceptive Effects of BDNF in Rodent Models
Increased BDNF expression in the spinal dorsal horn of rats with spinal nerve injury-induced neuropathic pain peaked at 24–48 h and was highly correlated with the onset of neuropathic pain behaviour [50,130]. Conversely, BDNF-mediated phosphorylation of the NR2B subunits of N-methyl-d-aspartate (NMDA) receptors was associated with the maintenance phase of neuropathic pain [50,130]. As the selective NR2B antagonist, Ro25-6981, markedly attenuated neuropathic pain in similarly nerve-injured rodents, a key role for NR2B-containing NMDA receptors in the maintenance of neuropathic pain, was affirmed [50].
The rostral ventromedial medulla (RVM) and the periaqueductal gray (PAG) matter are brain regions thought to have critical roles in descending facilitation of pronociceptive signalling in neuropathic pain [131]. In support of this notion, microinjection of BDNF (10–100 fmol) into the RVM facilitated nociception that was dependent on NMDA receptor activation [49]. Activity-dependent BDNF release from spinal neurons as well as supraspinal neurons in the hippocampus, PAG and RVM [49,132], was shown to underpin BDNF-induced descending facilitation [49,132]. Additionally, in a peripheral nerve ligation rat model of neuropathic pain, increased BDNF-TrkB signalling induced activation of phosphorylated-p44/p42 mitogen activated protein kinase (also called phospho-ERK, pERK) in the RVM that was correlated significantly with development of mechanical allodynia, a defining symptom of neuropathic pain [128].
11. Antinociceptive Effects of BDNF in Rodent Models
Contrary to the pronociceptive effects of BDNF in the RVM and PAG brain regions, antinociception was evoked by BDNF infusion (12 µg/day) into the midbrain of non-injured rats that appeared to be mediated by opioidergic signalling as well as increased serotoninergic activity both supraspinally and spinally [18]. Additionally, supraspinal release of BDNF particularly in the parabrachio-amygdala signalling pathway appears to have a key role in the mediation of morphine-induced analgesia [133]. In other work, neuropathic pain behaviour was abolished by recombinant adeno-associated viral (rAAV)-mediated BDNF over-expression in the spinal cord [134] or transplantation of genetically engineered BDNF-secreting neurons into the spinal cord of CCI-rats [135]. Hence, the net effects of sensory neuron derived BDNF on nociception in the dorsal horn are complex, in a manner similar to NGF [18,134,135].
Apart from pharmacological treatment of neuropathic pain, a considerable body of research in rodents shows that non-pharmacological interventions, such as electro-acupuncture (EA) at 2–10 Hz, alleviate neuropathic pain [136]. The underpinning analgesic mechanisms are complex and include attenuation of pronociceptive signalling and/or upregulation of antinociceptive signalling at multiple levels of the somatosensory system [136]. Of interest herein, EA-induced relief of neuropathic pain in rodents was accompanied by upregulated CNS expression of NTs particularly BDNF and NT-3 [137,138]. In neuropathic rats, EA-evoked pain relief was also accompanied by increased endogenous opioid peptide expression at peripheral nerve terminals, the spinal cord and brain, as well as enhanced descending noradrenergic and serotonergic neurotransmission [136,137,138,139,140]. This in turn inhibited NMDA receptor activation and glial cell activation in the spinal cord, with the net effect being decreased proinflammatory cytokine expression levels in the spinal dorsal horn [136,137,138,139,140].
12. BDNF: Role in Peripheral Neuropathic Pain
12.1. BDNF in Rodent Models of Peripheral Nerve Ligation
Peripheral nerve injury in rodents induced an upregulation in the synthesis and release of BDNF in small, medium and/or large diameter DRG neurons and in the superficial laminae of the spinal dorsal horn as well as in the gracile nuclei [44,124]. In more recent work, peripheral nerve injury on post-natal day 10 in infant rats did not induce neuropathic pain behaviour and this was correlated with a predominantly anti-inflammatory cytokine (IL-4 and IL-10) profile in the spinal dorsal horn of these animals [141]. However, once these same animals reached adolescence from post-natal day 21 onwards, the neuropathic pain behavioural phenotype became evident and this was accompanied by a switch from an antinociceptive to a pronociceptive cytokine/growth factor profile in the spinal dorsal horn [141]. Specifically, there was marked upregulation of spinal dorsal horn expression levels of BDNF and tumor necrosis factor alpha as well as a marked down-regulation of spinal dorsal horn expression levels of the anti-inflammatory cytokines, IL-4 and IL-10 [141]. These findings collectively suggest that although peripheral nerve injury in infancy may not appear to produce neuropathic pain due to powerful inhibition by anti-inflammatory cytokines in the spinal dorsal horn, there is a risk that it may emerge for no apparent reason in adolescence following switch of the neurochemical profile in the spinal dorsal horn from antinociceptive to pronociceptive [141].
Expression of TrkB, the high affinity BDNF receptor, was also upregulated in the lumbar DRGs and spinal dorsal horn [142,143]. In rodents with peripheral nerve injury, BDNF, like NGF, promoted sympathetic sprouting in the lumbar DRGs [74,144] and this was prevented by i.t. infusion of BDNF antiserum [145]. Affirming a role for augmented spinal BDNF-TrkB signalling in the pathobiology of neuropathic pain were observations that repeated i.t. administration of anti-BDNF or a BDNF-sequestering TrkB-Fc chimera protein, abolished neuropathic pain behaviours [55,146].
In rat neuropathic pain models, upregulated BDNF expression by an NGF-dependent mechanism in DRG neurons resulted in anterograde transport of BDNF by axonal mechanisms into the spinal dorsal horn [130,147,148], such that its release was restricted to TrkA-positive neurons in both lumbar DRGs and the spinal dorsal horn [112,147,149].
12.2. BDNF in Rodent Models of PDN
Investigation of neurotrophins other than NGF in the pathogenesis of PDN is in its infancy [150]. In diabetic rats, increased BDNF expression in the lumbar DRGs [151] was linked with decreased expression of voltage-gated potassium (Kv) channels in the lumbar DRGs and primary sensory neuron hyperexcitability [45]. This decreased Kv channel expression in DRG neurons from diabetic rats was reversed by treatment with anti-BDNF antibody for 2–4 h [45]. Mimicking these observations in diabetic rats, mRNA levels for NGF, BDNF and NT-3 and their receptors were increased in peripheral nerves with axonal pathologies in patients with PDN [52]. The upregulated BDNF and NT-3 mRNA levels in the diseased segments of peripheral nerves was positively correlated with the extent of nerve invasion by T-cells and macrophages rather than axonal pathology or demyelination [52]. Thus neuroinflammation appears to be a key pathobiologic event in PDN.
12.3. BDNF in Rodent Models of CIPN
A role for BDNF in the pathogenesis of CIPN in rodent models or in patients is scant. In a single study using a rat model of CIPN, BDNF expression in the spinal dorsal horn was upregulated compared with the corresponding level in sham-animals [28]. Clearly, additional research is warranted.
13. BDNF: Role in Central Neuropathic Pain
13.1. BDNF in Spinal Cord Injury Induced Neuropathic Pain Rodent Model
In a rat model of spinal cord injury-induced CNP, a pathobiologic role for upregulated BDNF-TrkB signalling in the spinal cord is well established [48]. In particular, the truncated TrkB receptor isoform expressed by both neuronal and non-neuronal cells is implicated in the pathogenesis of SCI-induced CNP as expression levels were consistently increased in both white and grey matter in spinal cord tissue [48,152].
13.2. BDNF in Multiple Sclerosis-Associated Neuropathic Pain Mouse Model
In MS, upregulated BDNF expression in the CNS is due to infiltration of BDNF-containing T-cells and upregulated synthesis by microglia and reactive astrocytes rather than spinal cord neurons [153,154]. In recent work from our laboratory that used our novel optimised relapsing-remitting EAE (RR-EAE) mouse model of MS-neuropathic pain [123], mature BDNF was found to be down-regulated whereas its precursor, pro-BDNF, was upregulated in the lumbar spinal cord of RR-EAE mice exhibiting pain behaviour [47]. Thus the possibility that pro-BDNF has a pronociceptive role in the pathobiology of MS-associated neuropathic pain, requires future investigation.
14. NT-3
The biological effects of NT-3, the third member of the neurotrophin family, are produced via signalling by its high affinity receptor, TrkC, expressed predominantly on large diameter myelinated Aβ fibers in the DRGs and spinal cord, as well as on C-fibres [65]. NT-3 promotes neuronal growth and survival of sensory and sympathetic neurons in a manner similar to NGF [155,156]. NT-3 may also influence TrkA-positive neurons [157,158], but a functional role for NT-3-TrkA signalling remains to be elucidated.
15. Pronociceptive Effects of NT-3 in Rodent Models
Initially, NT-3-TrkC signalling was thought to be confined to the modulation of proprioception [159]. However, anterograde transport of NT-3 from the lumbar DRGs into the spinal cord suggested a role in the modulation of nociceptive signalling in the spinal dorsal horn [61,65], but this remains to be investigated.
Intrathecally administered NT-3 at low doses (200 ng/day) for 20-days produced pronounced mechanical allodynia in the hindpaws of non-injured rats at 10–20 days after dose initiation [64]. The long latency for development of mechanical allodynia suggested that it is underpinned by complex central mechanisms that are unclear [64]. In other work, delivery of NT-3 (50 µg over 7 days) by osmotic minipump into the L5 DRGs of uninjured rats evoked temporary mechanical hypersensitivity that was significant at 1 but not 3 or 7-days after dose initiation [33].
16. Antinociceptive Effects of NT-3 in Rodent Models
NT-3 has been shown to inhibit the pronociceptive effects of NGF [65]. However, exogenous NGF had no effect on NT-3 expression levels in rodent DRGs, in contrast to the regulatory effects of NGF on BDNF expression [160]. Acute i.p. injection of NT-3 (10 or 20 mg/kg) evoked mechanical but not thermal hypoalgesia in the rat hindpaws that was underpinned by inhibition of SP release in the spinal cord that persisted in the presence of pharmacological concentrations of NGF [68]. Observations that naloxone reversed NT-3 induced inhibition of SP release in isolated rat spinal cords, suggested that cross-talk between NT-3 and inhibitory opioidergic signaling, may underpin NT-3 evoked antinociception [68]. Interestingly, other work has suggested that NT-3 may non-competitively block the binding of NGF to its high affinity receptor, TrkA thereby inhibiting NGF-mediated pronociceptive activity [157]. Alternatively, NT-3 may downregulate TrkA but not TrkC [158]. Irrespective of the mechanism, the onset of NT-3 antinociception is slow suggesting complexity [68].
17. NT-3 in Peripheral Neuropathic Pain
17.1. NT-3 in Rodent Models of Peripheral Nerve Ligation
In CCI-rats, i.t. infusion of NT-3 for 7-days (600 ng/μL/h) both prevented and attenuated thermal, but not mechanical hyperalgesia in the ipsilateral (injured side) hindpaws [65]. This pain-relief was highly correlated with a marked decrease in the otherwise upregulated expression levels of TRPV1, phosphorylated (activated) p38 MAPK (pp38 MAPK) as well as voltage-gated sodium channels (Nav1.8 and Nav1.9) in the ipsilateral lumbar DRGs of these neuropathic rats [65,66]. Since these pronociceptive mediators were mainly driven by NGF, it is plausible to suggest that NT-3 is a powerful negative regulator of NGF and its pronociceptive activity [65].
17.2. NT-3 in Rodent Models of PDN
In the lumbar DRGs of STZ-induced diabetic rats, suboptimal NT-3 dependent neurotrophic support and diabetes-induced deficits in axonal transport of NT-3 contributed to large fibre neuropathy [60]. Reversal of NT-3 depletion by intramuscular injection of adenovirus-based NT-3 gene therapy in STZ-diabetic rats, prevented development of diabetic neuropathy for up to 5-weeks diabetes induction [161]. In rats with impaired peripheral nerve conduction velocity at 8-weeks post-STZ administration, treatment with NT-3 for one month progressively reversed the deficit and normalised peripheral nerve conduction velocities to match those of non-diabetic rats [162].
17.3. NT-3 in Rodent Model of CIPN
In a rat model of cisplatin-induced CIPN, s.c. treatment with NT-3 reversed impaired sensory nerve conduction velocity, corrected abnormal neurofilament protein (NF200) distribution in large DRG sensory neurons, and reversed the large reduction in myelinated nerve fibre density in the sural nerves [59]. Importantly, sensory nerve conduction velocity was not altered in control rats administered the same NT-3 dosing regimen [59].
18. NT-3: Role in Central Neuropathic Pain
18.1. NT-3 in Spinal Cord Injury Induced Neuropathic Pain Rodent Models
Treatment of rats with complete spinal cord transection at T10 with an AAV-BDNF construct at the lesion site induced thermal hyperalgesia in the hindpaws as well as spasticity [163]. By contrast, AAV-NT-3 delivery to the site of spinal cord transection did not alter nociceptive thresholds in response to applied noxious thermal (heat) stimuli to the hindpaws [163].
18.2. NT-3 in Multiple Sclerosis-Associated Neuropathic Pain Mouse Model
The pathobiological role of NT-3 in rodent models of MS-induced neuropathic pain has not been investigated. However, as clinical data showed that NT-3 concentrations in peripheral blood mononuclear cells (PBMCs) were correlated strongly with widely accepted measures of brain atrophy in patients with MS, the NT-3 concentration in PBMCs may potentially exert a neuroprotective role in MS, but this requires future investigation [164].
19. NT-4 (NT-4/5)
Neurotrophin-4 was first isolated from Xenopus ovary and from snake venom [165]. Subsequently, it was found in mammals and reported as NT-4 [166] or NT-5 [167] and so it is often referred to as NT-4/5. NT-4, like BDNF, is a ligand for the TrkB receptor that is also synthesised in the DRGs and expressed in the spinal cord [55]. However, the biological effects of NT-4 differ from those of BDNF [168]. As NT-4 is expressed predominantly by motor neurons in the ventral horn of the spinal cord [56,169], this highlights NT-4’s key role in the maintenance and survival of motor neurons [56,170]. This notion is further supported by a study showing that NT-4 does not have a role in nociceptive transmission in ex vivo spinal cord preparations from mice null for NT-4, such that there were no deficits in the ventral root potentials evoked by stimulating nociceptive primary afferents [56].
20. NT-4: Lack of a Role in Neuropathic Pain
Although there was a transient decrease in spinal cord levels of NT-4 in rats at 6 h post-sciatic nerve transection, NT-4 levels were normalised by 12 h [53]. Importantly, repeated i.t. injection of anti-NT-4 did not reverse thermal hyperalgesia in a mouse model of peripheral neuropathic pain, in contrast to the pain relief evoked by repeated i.t. administration of anti-BDNF [55]. These findings collectively affirm the notion that NT-4-TrkB signalling does not have a role in the modulation of nociception. Although i.pl. injection of NT-4 (200 ng) evoked transient thermal hyperalgesia in rats, it was reversed by 24 h [36]. Thermal nociceptive responses did not differ between NT-4 knockout and wild-type mice [171]. However, as morphine-induced analgesia was reduced in the knockout mice, NT4-TrkB signaling is implicated in opioid analgesia [171]. In other work, NT-4 expression levels were significantly reduced in the sciatic nerves of a rat model of PDN [57] and in the brain of EAE-mice [58]. Hence, the exact role of NT-4 in modulating nociceptive signaling remains to be defined.
21. Proneurotrophins, p75NTR and Sortilin: Potential Modulatory Role in Pain
21.1. Proneurotrophins
Pro-NTs, like mature NTs, may be released extracellularly to exert their distinct biological actions not only via activation of p75NTR [4], but also via direct activation of Trk receptors [10,172] and indirectly after endocytosis and enzymatic cleavage [173]. To date, there is no direct evidence of a pronociceptive role for pro-neurotrophins [7] and so this knowledge gap requires future investigation. Nevertheless, observations that pro-NGF is upregulated in structures in close proximity to sprouting sensory and sympathetic nerve fibres, suggest a possible role in the pathogenesis of peripheral neuropathic pain [174]. Likewise, in our recent work that used our optimised mouse model of MS-induced neuropathic pain, neuropathic pain behaviour was significantly correlated with upregulated pro-BDNF expression and decreased mature-BDNF expression in the lumbar spinal cord [47]. As pERK expression was also increased in the lumbar spinal dorsal horn of these mice, our findings together suggests a pronociceptive role for pro-BDNF-TrkB-ERK signalling in MS-associated CNP [47]. In agreement, others have shown that pro-BDNF may exert TrkB-mediated downstream signaling, particularly ERK activation [10] that is implicated in central sensitisation and neuropathic pain development [175]. Clearly, future investigations to affirm (or not) that pro-NTs have direct pronociceptive effects by using mice null for p75NTR and/or i.t. treatment with recombinant pro-NTs or anti-p75NTR, are warranted.
21.2. P75NTR
The p75NTR has diverse neuronal functions ranging from apoptosis to neuronal survival [176]. Apart from its traditionally known role in apoptosis, Schwann cell expression of the p75NTR suggested a possible role in remyelination after peripheral nerve injury [177]. In ~79% of all DRG neurons, co-expression of full length Trk receptor mRNA with p75 mRNA suggested involvement of the latter in transduction of the biological effects of NTs [178]. Pronociceptive signalling after L5 spinal nerve ligation in rats [179] induced neuropathic pain behaviour that was alleviated by pharmacological blockade of the p75NTR [179]. Upregulated expression of p75NTR, but not TrkA, in adjacent (L4) non-injured primary afferents was inhibited by anti-NGF [179]. Furthermore, mice null for p75NTR had a dramatic insensitivity to noxious thermal stimuli that was associated with decreased sensory nerve fibre innervations in the skin [180].
21.3. Sortilin
Sortilin, a type-1 membrane protein that is a member of the Vps10p-domain family of sorting receptors, is expressed widely in the PNS and the CNS [8]. A role for sortilin participation in NT signalling via complex formation with Trk receptors and/or p75NTR has been proposed [8,181]. It is well established that pro-NTs interact with the p75NTR-sortlin complex to mediate apoptosis [7]. Conversely, co-expression of sortilin with the p75NTR in sensory neurons unrelated to apoptosis, has been suggested to promote a range of biological activities including pronociceptive processing, but this remains largely unexplored [7].
22. Neurotrophin Induced Activation of Trk or p75 Receptors: Downstream Signalling
The NTs, particularly NGF and BDNF, signalling predominantly via their cognate high affinity receptors, TrkA and TrkB respectively, are implicated in the pathobiology of sensory neuron hyperexcitability and neuropathic pain [80]. Following NT-Trk receptor binding, dimerisation and autophosphorylation induced receptor activation and initiation of intracellular downstream signalling via multiple pathways [182]. These included Ras-mitogen-activated protein kinases (Ras-MAPK), phosphatidylinositol 3-kinase (PI3K) and phospholipase C-gamma-1 (PLC-γ1) [182]. Ras-MAPK activation promoted neuronal differentiation, neurite outgrowth and neuroplasticity via post-translational and transcriptional modifications [182,183]. Following NGF and BDNF induced activation of TrkA and TrkB respectively; augmented p38 MAPK and ERK signalling in the lumbar DRGs and spinal cord have been implicated in the pathobiology of chronic pain [127,184]. In other work, signalling via the Ras-MAPK pathway indirectly activated PI3K signalling [182], that in turn upregulated Akt signalling [185]. Activation of PLC-γ1, induced production of inositol tris-phosphate (IP3) and diacylglycerol (DAG) [186]. IP3 mobilised Ca2+ release from intracellular stores and activated Ca2+-regulated protein kinase C (PKC) as well as calmodulin-regulated protein kinase pathways [186]. By contrast, DAG production led to activation of DAG-regulated PKC isoforms [186]. Both IP3- and DAG-regulated signalling pathways were associated with synaptic and structural neuroplasticity [187] and PKC was implicated in the development of central sensitisation and persistent pain [143,188]. Despite the widespread expression of the p75NTR by sensory neurons in the DRGs and spinal cord, investigation of the contribution of NT-p75NTR to the pathobiology of neuropathic pain is in its relative infancy [80]. Generally, NT-p75NTR signalling is thought to primarily upregulate ceramide production and induce signalling via the Jun kinase pathway and induce NF-kappa B (NFκB) activation [189].
23. Clinical Studies Using rNTs or Anti-NTs for Pain Relief
Two decades ago, research in rodent neuropathic pain models suggested that exogenous NGF may alleviate PDN and CIPN [147]. However, in a randomised controlled clinical trial in patients with PDN, recombinant human NGF (rhNGF) produced pain/hyperalgesia at the injection site so severe that 22 patients out of 504 discontinued treatment [190]. The intolerable pronociceptive side-effects of rNGF hampered its testing in patients with CIPN [191].
Conversely, a randomised, double-blind, placebo-controlled clinical trial of tanezumab, a monoclonal antibody to NGF, showed that it produced significant analgesia in patients with osteoarthritis of the knee [11,192]. However, in a significant patient sub-group, osteoarthritis worsened such that some patients had to have total joint replacement [11,192]. Other frequent tanezumab-related adverse effects were arthralgia, pain in the extremities, paresthesia and peripheral edema [11,192]. Due to safety concerns, particularly the requirement for joint replacements in some patients, the United States Food and Drug Administration (FDA) placed all ongoing clinical trials of tanezumab for the relief of pain in patients with PDN [193], postherpetic neuralgia (PHN) [193] and low back pain [194] on clinical hold. However, very recently, the FDA has lifted this clinical hold [195,196,197]. An anti-TrkA monoclonal antibody, GBR900, is also in clinical trials as a potential novel analgesic that may have a superior safety profile compared with anti-NGF antibodies [198].
Clinical studies of anti-BDNF antibodies for the relief of chronic inflammatory and neuropathic pain conditions are lacking. This may be due to the unfavourable side effects that emerged during the early phase clinical trials of anti-NGF. Although the safety of NT-3 was established in healthy subjects [199], a Phase-1 clinical trial of NT-3 for relief of DPN and CIPN was discontinued in 1997 but no reports were published [200]. Preclinical research to date indicates that NT-4 does not have a role in the modulation of nociception and this may explain the lack of clinical trials of NT-4 for relief of neuropathic pain.
24. Conclusions
Considerable research to date has implicated a pathobiologic role for NGF and BDNF in neuropathic pain, whereas NT-3 generally appeared to alleviate neuropathic pain in rodent models. The conventional view is that the effects of NGF, BDNF and NT-3 in neuropathic pain are evoked by signalling via their cognate high affinity receptors, viz TrkA, TrkB and TrkC respectively. However, NGF, BDNF and NT-3 signalling via the p75NTR may also modulate neuropathic pain by either direct or indirect mechanisms, but additional research is needed. A role for pro-NTs signalling via their high affinity receptor, the p75NTR, and/or via low affinity binding to TrkA, TrkB or TrkC to modulate pro-nociceptive signalling in neuropathic pain, has also been suggested. However, research on this topic is in its relative infancy.
Clinical research two decades ago showed that systemic administration of NGF evoked rather than relieved pain. This key observation re-directed efforts to the development of antibodies and small molecules that block rather than augment NGF-TrkA signalling, as a strategy to alleviate chronic inflammatory and neuropathic pain. Currently, multiple NGF antibody clinical trials in patients with a range of peripheral neuropathic pain conditions have recommenced following the decision of the US FDA in March 2015 to lift the clinical hold.
Apart from clinical trial investigation of NGF antibodies as novel analgesics for relief of neuropathic pain, small molecule TrkA inhibitors are also in development as they may have a reduced risk for serious adverse-effects due to selective blockade of NGF-TrkA signalling whilst not impeding NGF-p75NTR signalling. However this remains to be determined in future investigations.
Acknowledgments
Nematullah Khan (NK) was supported by an International PhD Scholarship funded by The University of Queensland (UQ), Australia.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ceni, C.; Unsain, N.; Zeinieh, M.P.; Barker, P.A. Neurotrophins in the regulation of cellular survival and death. Handb. Exp. Pharmacol. 2014, 220, 193–221. [Google Scholar] [PubMed]
- Skaper, S.D. The neurotrophin family of neurotrophic factors: An overview. Methods Mol. Biol. 2012, 846, 1–12. [Google Scholar] [PubMed]
- Davies, A.M. Neurotrophins giveth and they taketh away. Nat. Neurosci. 2008, 11, 627–628. [Google Scholar] [CrossRef] [PubMed]
- Teng, K.K.; Felice, S.; Kim, T.; Hempstead, B.L. Understanding proneurotrophin actions: Recent advances and challenges. Dev. Neurobiol. 2010, 70, 350–359. [Google Scholar] [CrossRef] [PubMed]
- Patapoutian, A.; Reichardt, L.F. Trk receptors: Mediators of neurotrophin action. Curr. Opin. Neurobiol. 2001, 11, 272–280. [Google Scholar] [CrossRef]
- Davies, A.M.; Minichiello, L.; Klein, R. Developmental changes in NT3 signalling via TrkA and TrkB in embryonic neurons. EMBO J. 1995, 14, 4482–4489. [Google Scholar] [PubMed]
- Lewin, G.R.; Nykjaer, A. Pro-neurotrophins, sortilin, and nociception. Eur. J. Neurosci. 2014, 39, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Nykjaer, A.; Willnow, T.E. Sortilin: A receptor to regulate neuronal viability and function. Trends Neurosci. 2012, 35, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Masoudi, R.; Ioannou, M.S.; Coughlin, M.D.; Pagadala, P.; Neet, K.E.; Clewes, O.; Allen, S.J.; Dawbarn, D.; Fahnestock, M. Biological activity of nerve growth factor precursor is dependent upon relative levels of its receptors. J. Biol. Chem. 2009, 284, 18424–18433. [Google Scholar] [CrossRef] [PubMed]
- Fayard, B.; Loeffler, S.; Weis, J.; Vögelin, E.; Krüttgen, A. The secreted brain-derived neurotrophic factor precursor pro-BDNF binds to TrkB and p75NTR but not to TrkA or TrkC. J. Neurosci. Res. 2005, 80, 18–28. [Google Scholar] [CrossRef] [PubMed]
- Lane, N.E.; Schnitzer, T.J.; Birbara, C.A.; Mokhtarani, M.; Shelton, D.L.; Smith, M.D.; Brown, M.T. Tanezumab for the treatment of pain from osteoarthritis of the knee. N. Engl. J. Med. 2010, 363, 1521–1531. [Google Scholar] [CrossRef] [PubMed]
- Treede, R.D.; Jensen, T.S.; Campbell, J.N.; Cruccu, G.; Dostrovsky, J.O.; Griffin, J.W.; Hansson, P.; Hughes, R.; Nurmikko, T.; Serra, J. Neuropathic pain: Redefinition and a grading system for clinical and research purposes. Neurology 2008, 70, 1630–1635. [Google Scholar] [CrossRef] [PubMed]
- Dworkin, R.H.; Levy, R.M.; Mackey, S.C.; Mayer, J.; Miaskowski, C.; Raja, S.N.; Rice, A.S.C.; Schmader, K.E.; Stacey, B.; Stanos, S.; et al. Recommendations for the pharmacological management of neuropathic pain: An overview and literature update. Mayo Clin. Proc. 2010, 85, S3–S14. [Google Scholar] [CrossRef] [PubMed]
- Dworkin, R.H.; O’Connor, A.B.; Kent, J.; Mackey, S.C.; Raja, S.N.; Stacey, B.R.; Levy, R.M.; Backonja, M.; Baron, R.; Harke, H.; et al. Interventional management of neuropathic pain: NeuPSIG recommendations. Pain 2013, 154, 2249–2261. [Google Scholar] [CrossRef] [PubMed]
- Nijs, J.; Meeus, M.; Versijpt, J.; Moens, M.; Bos, I.; Knaepen, K.; Meeusen, R. Brain-derived neurotrophic factor as a driving force behind neuroplasticity in neuropathic and central sensitization pain: A new therapeutic target? Expert Opin. Ther. Targets 2015, 19, 565–576. [Google Scholar] [CrossRef] [PubMed]
- McMahon, S.B.; Jones, N.G. Plasticity of pain signaling: Role of neurotrophic factors exemplified by acid-induced pain. J. Neurobiol. 2004, 61, 72–87. [Google Scholar] [CrossRef] [PubMed]
- Pezet, S.; McMahon, S.B. Neurotrophins: Mediators and modulators of pain. Annu. Rev. Neurosci. 2006, 29, 507–538. [Google Scholar] [CrossRef] [PubMed]
- Siuciak, J.A.; Altar, C.A.; Wiegand, S.J.; Lindsay, R.M. Antinociceptive effect of brain-derived neurotrophic factor and neurotrophin-3. Brain Res. 1994, 633, 326–330. [Google Scholar] [CrossRef]
- Siuciak, J.A.; Wong, V.; Pearsall, D.; Wiegand, S.J.; Lindsay, R.M. Bdnf produces analgesia in the formalin test and modifies neuropeptide levels in rat brain and spinal cord areas associated with nociception. Eur. J. Neurosci. 1995, 7, 663–670. [Google Scholar] [CrossRef] [PubMed]
- Aloe, L.; Rocco, M.; Bianchi, P.; Manni, L. Nerve growth factor: From the early discoveries to the potential clinical use. J. Transl. Med. 2012, 10, 239. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.S.; Nishimura, M.C.; Armanini, M.P.; Crowley, C.; Spencer, S.D.; Phillips, H.S. Disruption of a single allele of the nerve growth factor gene results in atrophy of basal forebrain cholinergic neurons and memory deficits. J. Neurosci. 1997, 17, 7288–7296. [Google Scholar] [PubMed]
- Zhou, X.F.; Deng, Y.S.; Chie, E.; Xue, Q.; Zhong, J.H.; McLachlan, E.M.; Rush, R.A.; Xian, C.J. Satellite-cell-derived nerve growth factor and neurotrophin-3 are involved in noradrenergic sprouting in the dorsal root ganglia following peripheral nerve injury in the rat. Eur. J. Neurosci. 1999, 11, 1711–1722. [Google Scholar] [CrossRef] [PubMed]
- Vivoli, E.; di Cesare Mannelli, L.; Salvicchi, A.; Bartolini, A.; Koverech, A.; Nicolai, R.; Benatti, P.; Ghelardini, C. Acetyl-l-carnitine increases artemin level and prevents neurotrophic factor alterations during neuropathy. Neuroscience 2010, 167, 1168–1174. [Google Scholar] [CrossRef] [PubMed]
- Nakahashi, Y.; Kamiya, Y.; Funakoshi, K.; Miyazaki, T.; Uchimoto, K.; Tojo, K.; Ogawa, K.; Fukuoka, T.; Goto, T. Role of nerve growth factor-tyrosine kinase receptor a signaling in paclitaxel-induced peripheral neuropathy in rats. Biochem. Biophys. Res. Commun. 2014, 444, 415–419. [Google Scholar] [CrossRef] [PubMed]
- Apfel, S.C.; Arezzo, J.C.; Lipson, L.; Kessler, J.A. Nerve growth factor prevents experimental cisplatin neuropathy. Ann. Neurol. 1992, 31, 76–80. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.T.; Dauch, J.R.; Hayes, J.M.; Hong, Y.; Feldman, E.L. Nerve growth factor mediates mechanical allodynia in a mouse model of type 2 diabetes. J. Neuropathol. Exp. Neurol. 2009, 68, 1229–1243. [Google Scholar] [CrossRef] [PubMed]
- Unger, J.W.; Klitzsch, T.; Pera, S.; Reiter, R. Nerve growth factor (NGF) and diabetic neuropathy in the rat: Morphological investigations of the sural nerve, dorsal root ganglion, and spinal cord. Exp. Neurol. 1998, 153, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Aloe, L.; Manni, L.; Properzi, F.; de Santis, S.; Fiore, M. Evidence that nerve growth factor promotes the recovery of peripheral neuropathy induced in mice by cisplatin: Behavioral, structural and biochemical analysis. Auton. Neurosci. 2000, 86, 84–93. [Google Scholar] [CrossRef]
- Calza, L.; Fernandez, M.; Giuliani, A.; Aloe, L.; Giardino, L. Thyroid hormone activates oligodendrocyte precursors and increases a myelin-forming protein and NGF content in the spinal cord during experimental allergic encephalomyelitis. Proc. Natl. Acad. Sci. USA 2002, 99, 3258–3263. [Google Scholar] [CrossRef] [PubMed]
- Calza, L.; Giardino, L.; Pozza, M.; Micera, A.; Aloe, L. Time-course changes of nerve growth factor, corticotropin-releasing hormone, and nitric oxide synthase isoforms and their possible role in the development of inflammatory response in experimental allergic encephalomyelitis. Proc. Natl. Acad. Sci. USA 1997, 94, 3368–3373. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.Y.; Kim, D.Y.; Yune, T.Y.; Shin, D.H.; Baek, S.B.; Kim, C.J. Treadmill exercise reduces spinal cord injury-induced apoptosis by activating the PI3K/Akt pathway in rats. Exp. Ther. Med. 2014, 7, 587–593. [Google Scholar] [PubMed]
- Christensen, M.D.; Hulsebosch, C.E. Spinal cord injury and anti-NGF treatment results in changes in cgrp density and distribution in the dorsal horn in the rat. Exp. Neurol. 1997, 147, 463–475. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.F.; Deng, Y.S.; Xian, C.J.; Zhong, J.H. Neurotrophins from dorsal root ganglia trigger allodynia after spinal nerve injury in rats. Eur J. Neurosci. 2000, 12, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Malcangio, M.; Ramer, M.S.; Boucher, T.J.; McMahon, S.B. Intrathecally injected neurotrophins and the release of substance P from the rat isolated spinal cord. Eur. J. Neurosci. 2000, 12, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Mills, C.D.; Nguyen, T.; Tanga, F.Y.; Zhong, C.; Gauvin, D.M.; Mikusa, J.; Gomez, E.J.; Salyers, A.K.; Bannon, A.W. Characterization of nerve growth factor-induced mechanical and thermal hypersensitivity in rats. Eur. J. Pain 2013, 17, 469–479. [Google Scholar] [CrossRef] [PubMed]
- Shu, X.; Mendell, L.M. Nerve growth factor acutely sensitizes the response of adult rat sensory neurons to capsaicin. Neurosci. Lett. 1999, 274, 159–162. [Google Scholar] [CrossRef]
- Malcangio, M.; Garrett, N.E.; Tomlinson, D.R. Nerve growth factor treatment increases stimulus-evoked release of sensory neuropeptides in the rat spinal cord. Eur. J. Neurosci. 1997, 9, 1101–1104. [Google Scholar] [CrossRef] [PubMed]
- Woolf, C.J. Phenotypic modification of primary sensory neurons: The role of nerve growth factor in the production of persistent pain. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1996, 351, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Gu, J.; Li, Y.Q.; Tao, Y.X. Are voltage-gated sodium channels on the dorsal root ganglion involved in the development of neuropathic pain? Mol. Pain 2011, 7, 16. [Google Scholar] [CrossRef] [PubMed]
- Waxman, S.G. The molecular pathophysiology of pain: Abnormal expression of sodium channel genes and its contributions to hyperexcitability of primary sensory neurons. Pain 1999, 81, S133–S140. [Google Scholar] [CrossRef]
- Fjell, J.; Cummins, T.R.; Fried, K.; Black, J.A.; Waxman, S.G. In vivo NGF deprivation reduces SNS expression and TTX-R sodium currents in IB4-negative DRG neurons. J. Neurophysiol. 1999, 81, 803–810. [Google Scholar] [PubMed]
- Fjell, J.; Cummins, T.R.; Dib-Hajj, S.D.; Fried, K.; Black, J.A.; Waxman, S.G. Differential role of GDNF and NGF in the maintenance of two TTX-resistant sodium channels in adult drg neurons. Mol. Brain Res. 1999, 67, 267–282. [Google Scholar] [CrossRef]
- Cirillo, G.; Cavaliere, C.; Bianco, M.R.; de Simone, A.; Colangelo, A.M.; Sellitti, S.; Alberghina, L.; Papa, M. Intrathecal NGF administration reduces reactive astrocytosis and changes neurotrophin receptors expression pattern in a rat model of neuropathic pain. Cell. Mol. Neurobiol. 2010, 30, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Ha, S.O.; Kim, J.K.; Hong, H.S.; Kim, D.S.; Cho, H.J. Expression of brain-derived neurotrophic factor in rat dorsal root ganglia, spinal cord and gracile nuclei in experimental models of neuropathic pain. Neuroscience 2001, 107, 301–309. [Google Scholar] [CrossRef]
- Cao, X.H.; Byun, H.S.; Chen, S.R.; Cai, Y.Q.; Pan, H.L. Reduction in voltage-gated K(+) channel activity in primary sensory neurons in painful diabetic neuropathy: Role of brain-derived neurotrophic factor. J. Neurochem. 2010, 114, 1460–1475. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Frost, E.E.; Begum, F.; Vora, P.; Au, K.; Gong, Y.; MacNeil, B.; Pillai, P.; Namaka, M. The role of dorsal root ganglia activation and brain-derived neurotrophic factor in multiple sclerosis. J. Cell Mol. Med. 2012, 16, 1856–1865. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Gordon, R.; Woodruff, T.M.; Smith, M.T. Antiallodynic effects of alpha lipoic acid in an optimized RR-EAE mouse model of MS-neuropathic pain are accompanied by attenuation of upregulated BDNF-TrkB-ERK signaling in the dorsal horn of the spinal cord. Pharm. Res. Perpect. 2015, 3, e00137. [Google Scholar]
- Wu, J.; Renn, C.L.; Faden, A.I.; Dorsey, S.G. TrkB.T1 contributes to neuropathic pain after spinal cord injury through regulation of cell cycle pathways. J. Neurosci. 2013, 33, 12447–12463. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Robbins, M.T.; Wei, F.; Zou, S.; Dubner, R.; Ren, K. Supraspinal brain-derived neurotrophic factor signaling: A novel mechanism for descending pain facilitation. J. Neurosci. 2006, 26, 126–137. [Google Scholar] [CrossRef] [PubMed]
- Geng, S.J.; Liao, F.F.; Dang, W.H.; Ding, X.; Liu, X.D.; Cai, J.; Han, J.S.; Wan, Y.; Xing, G.G. Contribution of the spinal cord BDNF to the development of neuropathic pain by activation of the NR2B-containing nmda receptors in rats with spinal nerve ligation. Exp. Neurol. 2010, 222, 256–266. [Google Scholar] [CrossRef] [PubMed]
- Coull, J.A.; Boudreau, D.; Bachand, K.; Prescott, S.A.; Nault, F.; Sik, A.; De Koninck, P.; De Koninck, Y. Trans-synaptic shift in anion gradient in spinal lamina-I neurons as a mechanism of neuropathic pain. Nature 2003, 424, 938–942. [Google Scholar] [CrossRef] [PubMed]
- Sobue, G.; Yamamoto, M.; Doyu, M.; Li, M.; Yasuda, T.; Mitsuma, T. Expression of mRNAs for neurotrophins (NGF, BDNF and NT-3) and their receptors (p75NGFR, Trk, TrkB, and TrkC) in human peripheral neuropathies. Neurochem. Res. 1998, 23, 821–829. [Google Scholar] [CrossRef] [PubMed]
- Funakoshi, H.; Frisen, J.; Barbany, G.; Timmusk, T.; Zachrisson, O.; Verge, V.M.; Persson, H. Differential expression of mrnas for neurotrophins and their receptors after axotomy of the sciatic nerve. J. Cell Biol. 1993, 123, 455–465. [Google Scholar] [CrossRef] [PubMed]
- Ramer, L.M.; McPhail, L.T.; Borisoff, J.F.; Soril, L.J.; Kaan, T.K.; Lee, J.H.; Saunders, J.W.; Hwi, L.P.; Ramer, M.S. Endogenous TrkB ligands suppress functional mechanosensory plasticity in the deafferented spinal cord. J. Neurosci. 2007, 27, 5812–5822. [Google Scholar] [CrossRef] [PubMed]
- Yajima, Y.; Narita, M.; Narita, M.; Matsumoto, N.; Suzuki, T. Involvement of a spinal brain-derived neurotrophic factor/full-length TrkB pathway in the development of nerve injury-induced thermal hyperalgesia in mice. Brain Res. 2002, 958, 338–346. [Google Scholar] [CrossRef]
- Heppenstall, P.A.; Lewin, G.R. BDNF but not NT-4 is required for normal flexion reflex plasticity and function. Proc. Natl. Acad. Sci. USA 2001, 98, 8107–8112. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Pena, A.; Botana, M.; Gonzalez, M.; Requejo, F. Expression of neurotrophins and their receptors in sciatic nerve of experimentally diabetic rats. Neurosci. Lett. 1995, 200, 37–40. [Google Scholar] [CrossRef]
- Aharoni, R.; Eilam, R.; Domev, H.; Labunskay, G.; Sela, M.; Arnon, R. The immunomodulator glatiramer acetate augments the expression of neurotrophic factors in brains of experimental autoimmune encephalomyelitis mice. Proc. Natl. Acad. Sci. USA 2005, 102, 19045–19050. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.Q.; Dybdal, N.; Shinsky, N.; Murnane, A.; Schmelzer, C.; Siegel, M.; Keller, G.; Hefti, F.; Phillips, H.S.; Winslow, J.W. Neurotrophin-3 reverses experimental cisplatin-induced peripheral sensory neuropathy. Ann. Neurol. 1995, 38, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Fernyhough, P.; Diemel, L.T.; Tomlinson, D.R. Target tissue production and axonal transport of neurotrophin-3 are reduced in streptozotocin-diabetic rats. Diabetologia 1998, 41, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.H.; Meng, Q.S.; Qi, J.G.; Zhang, W.M.; Chen, J.; Wu, L.F. NT-3 expression in spared DRG and the associated spinal laminae as well as its anterograde transport in sensory neurons following removal of adjacent DRG in cats. Neurochem. Res. 2008, 33, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Muhallab, S.; Lundberg, C.; Gielen, A.W.; Lidman, O.; Svenningsson, A.; Piehl, F.; Olsson, T. Differential expression of neurotrophic factors and inflammatory cytokines by myelin basic protein-specific and other recruited T cells infiltrating the central nervous system during experimental autoimmune encephalomyelitis. Scand. J. Immunol. 2002, 55, 264–273. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Shine, H.D. Neurotrophic factors expressed in both cortex and spinal cord induce axonal plasticity after spinal cord injury. J. Neurosci. Res. 2003, 74, 221–226. [Google Scholar] [CrossRef] [PubMed]
- White, D.M. Contribution of neurotrophin-3 to the neuropeptide Y-induced increase in neurite outgrowth of rat dorsal root ganglion cells. Neuroscience 1998, 86, 257–263. [Google Scholar] [CrossRef]
- Wilson-Gerwing, T.D.; Dmyterko, M.V.; Zochodne, D.W.; Johnston, J.M.; Verge, V.M. Neurotrophin-3 suppresses thermal hyperalgesia associated with neuropathic pain and attenuates transient receptor potential vanilloid receptor-1 expression in adult sensory neurons. J. Neurosci. 2005, 25, 758–767. [Google Scholar] [CrossRef] [PubMed]
- Wilson-Gerwing, T.D.; Stucky, C.L.; McComb, G.W.; Verge, V.M. Neurotrophin-3 significantly reduces sodium channel expression linked to neuropathic pain states. Exp. Neurol. 2008, 213, 303–314. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Choi, J.Y.; Kim, R.U.; Lee, Y.S.; Cho, H.J.; Kim, D.S. Downregulation of voltage-gated potassium channel alpha gene expression by axotomy and neurotrophins in rat dorsal root ganglia. Mol. Cells 2003, 16, 256–259. [Google Scholar] [PubMed]
- Malcangio, M.; Garrett, N.E.; Cruwys, S.; Tomlinson, D.R. Nerve growth factor- and neurotrophin-3-induced changes in nociceptive threshold and the release of substance P from the rat isolated spinal cord. J. Neurosci. 1997, 17, 8459–8467. [Google Scholar] [PubMed]
- Lewin, G.R.; Ritter, A.M.; Mendell, L.M. Nerve growth factor-induced hyperalgesia in the neonatal and adult rat. J. Neurosci. 1993, 13, 2136–2148. [Google Scholar] [PubMed]
- Rukwied, R.; Mayer, A.; Kluschina, O.; Obreja, O.; Schley, M.; Schmelz, M. NGF induces non-inflammatory localized and lasting mechanical and thermal hypersensitivity in human skin. Pain 2010, 148, 407–413. [Google Scholar] [CrossRef] [PubMed]
- Thompson, S.W.; Dray, A.; McCarson, K.E.; Krause, J.E.; Urban, L. Nerve growth factor induces mechanical allodynia associated with novel a fibre-evoked spinal reflex activity and enhanced neurokinin-1 receptor activation in the rat. Pain 1995, 62, 219–231. [Google Scholar] [CrossRef]
- Lewin, G.R.; Rueff, A.; Mendell, L.M. Peripheral and central mechanisms of NGF-induced hyperalgesia. Eur. J. Neurosci. 1994, 6, 1903–1912. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.G.; Munson, J.B.; Thompson, S.W. A role for nerve growth factor in sympathetic sprouting in rat dorsal root ganglia. Pain 1999, 79, 21–29. [Google Scholar] [CrossRef]
- Deng, Y.S.; Zhong, J.H.; Zhou, X.F. Effects of endogenous neurotrophins on sympathetic sprouting in the dorsal root ganglia and allodynia following spinal nerve injury. Exp. Neurol. 2000, 164, 344–350. [Google Scholar] [CrossRef] [PubMed]
- Mearow, K.M. The effects of NGF and sensory nerve stimulation on collateral sprouting and gene expression in adult sensory neurons. Exp. Neurol. 1998, 151, 14–25. [Google Scholar] [CrossRef] [PubMed]
- Yen, L.D.; Bennett, G.J.; Ribeiro-da-Silva, A. Sympathetic sprouting and changes in nociceptive sensory innervation in the glabrous skin of the rat hind paw following partial peripheral nerve injury. J. Comp. Neurol. 2006, 495, 679–690. [Google Scholar] [CrossRef] [PubMed]
- Otten, U.; Schwab, M.; Gagnon, C.; Thoenen, H. Selective induction of tyrosine hydroxylase and dopamine beta-hydroxylase by nerve growth factor: Comparison between adrenal medulla and sympathetic ganglia of adult and newborn rats. Brain Res. 1977, 133, 291–303. [Google Scholar] [CrossRef]
- Shadiack, A.M.; Sun, Y.; Zigmond, R.E. Nerve growth factor antiserum induces axotomy-like changes in neuropeptide expression in intact sympathetic and sensory neurons. J. Neurosci. 2001, 21, 363–371. [Google Scholar] [PubMed]
- Zhang, X.; Huang, J.; McNaughton, P.A. NGF rapidly increases membrane expression of TRPV1 heat-gated ion channels. EMBO J. 2005, 24, 4211–4223. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.H.; Nicol, G.D. NGF-mediated sensitization of the excitability of rat sensory neurons is prevented by a blocking antibody to the p75 neurotrophin receptor. Neurosci. Lett. 2004, 366, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Khodorova, A.; Nicol, G.D.; Strichartz, G. The p75NTR signaling cascade mediates mechanical hyperalgesia induced by nerve growth factor injected into the rat hind paw. Neuroscience 2013, 254, 312–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahluwalia, J.; Urban, L.; Bevan, S.; Capogna, M.; Nagy, I. Cannabinoid 1 receptors are expressed by nerve growth factor- and glial cell-derived neurotrophic factor-responsive primary sensory neurones. Neuroscience 2002, 110, 747–753. [Google Scholar] [CrossRef]
- Agarwal, N.; Pacher, P.; Tegeder, I.; Amaya, F.; Constantin, C.E.; Brenner, G.J.; Rubino, T.; Michalski, C.W.; Marsicano, G.; Monory, K.; et al. Cannabinoids mediate analgesia largely via peripheral type 1 cannabinoid receptors in nociceptors. Nat. Neurosci. 2007, 10, 870–879. [Google Scholar] [CrossRef] [PubMed]
- Anand, U.; Otto, W.R.; Sanchez-Herrera, D.; Facer, P.; Yiangou, Y.; Korchev, Y.; Birch, R.; Benham, C.; Bountra, C.; Chessell, I.P.; et al. Cannabinoid receptor CB2 localisation and agonist-mediated inhibition of capsaicin responses in human sensory neurons. Pain 2008, 138, 667–680. [Google Scholar] [CrossRef] [PubMed]
- McDowell, T.S.; Wang, Z.Y.; Singh, R.; Bjorling, D. CB1 cannabinoid receptor agonist prevents NGF-induced sensitization of TRPV1 in sensory neurons. Neurosci. Lett. 2013, 551, 34–38. [Google Scholar] [CrossRef] [PubMed]
- Jhaveri, M.D.; Richardson, D.; Chapman, V. Endocannabinoid metabolism and uptake: Novel targets for neuropathic and inflammatory pain. Br. J. Pharmacol. 2007, 152, 624–632. [Google Scholar] [CrossRef] [PubMed]
- Luongo, L.; Maione, S.; di Marzo, V. Endocannabinoids and neuropathic pain: Focus on neuron-glia and endocannabinoid-neurotrophin interactions. Eur. J. Neurosci. 2014, 39, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Petrosino, S.; Palazzo, E.; de Novellis, V.; Bisogno, T.; Rossi, F.; Maione, S.; di Marzo, V. Changes in spinal and supraspinal endocannabinoid levels in neuropathic rats. Neuropharmacology 2007, 52, 415–422. [Google Scholar] [CrossRef] [PubMed]
- Guindon, J.; Lai, Y.; Takacs, S.M.; Bradshaw, H.B.; Hohmann, A.G. Alterations in endocannabinoid tone following chemotherapy-induced peripheral neuropathy: Effects of endocannabinoid deactivation inhibitors targeting fatty-acid amide hydrolase and monoacylglycerol lipase in comparison to reference analgesics following cisplatin treatment. Pharmacol. Res. 2013, 67, 94–109. [Google Scholar] [PubMed]
- Owolabi, J.B.; Rizkalla, G.; Tehim, A.; Ross, G.M.; Riopelle, R.J.; Kamboj, R.; Ossipov, M.; Bian, D.; Wegert, S.; Porreca, F.; et al. Characterization of antiallodynic actions of ALE-0540, a novel nerve growth factor receptor antagonist, in the rat. J. Pharmacol. Exp. Ther. 1999, 289, 1271–1276. [Google Scholar] [PubMed]
- Ro, L.S.; Chen, S.T.; Tang, L.M.; Jacobs, J.M. Effect of NGF and anti-NGF on neuropathic pain in rats following chronic constriction injury of the sciatic nerve. Pain 1999, 79, 265–274. [Google Scholar] [CrossRef]
- Wild, K.D.; Bian, D.; Zhu, D.; Davis, J.; Bannon, A.W.; Zhang, T.J.; Louis, J.C. Antibodies to nerve growth factor reverse established tactile allodynia in rodent models of neuropathic pain without tolerance. J. Pharmacol. Exp. Ther. 2007, 322, 282–287. [Google Scholar] [CrossRef] [PubMed]
- Ugolini, G.; Marinelli, S.; Covaceuszach, S.; Cattaneo, A.; Pavone, F. The function neutralizing anti-TrkA antibody MNAC13 reduces inflammatory and neuropathic pain. Proc. Natl. Acad. Sci. USA 2007, 104, 2985–2990. [Google Scholar] [CrossRef] [PubMed]
- Verge, V.M.; Riopelle, R.J.; Richardson, P.M. Nerve growth factor receptors on normal and injured sensory neurons. J. Neurosci. 1989, 9, 914–922. [Google Scholar] [PubMed]
- Verge, V.M.; Richardson, P.M.; Wiesenfeld-Hallin, Z.; Hokfelt, T. Differential influence of nerve growth factor on neuropeptide expression in vivo: A novel role in peptide suppression in adult sensory neurons. J. Neurosci. 1995, 15, 2081–2096. [Google Scholar] [PubMed]
- Schmidt, Y.; Unger, J.W.; Bartke, I.; Reiter, R. Effect of nerve growth factor on peptide neurons in dorsal root ganglia after taxol or cisplatin treatment and in diabetic (db/db) mice. Exp. Neurol. 1995, 132, 16–23. [Google Scholar] [CrossRef]
- Hellweg, R.; Hartung, H.D. Endogenous levels of nerve growth factor (NGF) are altered in experimental diabetes mellitus: A possible role for NGF in the pathogenesis of diabetic neuropathy. J. Neurosci. Res. 1990, 26, 258–267. [Google Scholar] [CrossRef] [PubMed]
- Konings, P.N.; Makkink, W.K.; van Delft, A.M.; Ruigt, G.S. Reversal by NGF of cytostatic drug-induced reduction of neurite outgrowth in rat dorsal root ganglia in vitro. Brain Res. 1994, 640, 195–204. [Google Scholar] [CrossRef]
- Cavaletti, G.; Bogliun, G.; Marzorati, L.; Zincone, A.; Piatti, M.; Colombo, N.; Franchi, D.; la Presa, M.T.; Lissoni, A.; Buda, A.; et al. Early predictors of peripheral neurotoxicity in cisplatin and paclitaxel combination chemotherapy. Ann. Oncol. 2004, 15, 1439–1442. [Google Scholar] [CrossRef] [PubMed]
- Apfel, S.C.; Arezzo, J.C.; Brownlee, M.; Federoff, H.; Kessler, J.A. Nerve growth factor administration protects against experimental diabetic sensory neuropathy. Brain Res. 1994, 634, 7–12. [Google Scholar] [CrossRef]
- Watson, J.J.; Allen, S.J.; Dawbarn, D. Targeting nerve growth factor in pain: What is the therapeutic potential? BioDrugs 2008, 22, 349–359. [Google Scholar] [CrossRef] [PubMed]
- Mantyh, P.W.; Koltzenburg, M.; Mendell, L.M.; Tive, L.; Shelton, D.L. Antagonism of nerve growth factor-TrkA signaling and the relief of pain. Anesthesiology 2011, 115, 189–204. [Google Scholar] [CrossRef] [PubMed]
- McKelvey, L.; Shorten, G.D.; O’Keeffe, G.W. Nerve growth factor-mediated regulation of pain signalling and proposed new intervention strategies in clinical pain management. J. Neurochem. 2013, 124, 276–289. [Google Scholar] [CrossRef] [PubMed]
- Romero, M.I.; Rangappa, N.; Li, L.; Lightfoot, E.; Garry, M.G.; Smith, G.M. Extensive sprouting of sensory afferents and hyperalgesia induced by conditional expression of nerve growth factor in the adult spinal cord. J. Neurosci. 2000, 20, 4435–4445. [Google Scholar] [PubMed]
- Taniuchi, M.; Clark, H.B.; Johnson, E.M., Jr. Induction of nerve growth factor receptor in schwann cells after axotomy. Proc. Natl. Acad. Sci. USA 1986, 83, 4094–4098. [Google Scholar] [CrossRef] [PubMed]
- Taniuchi, M.; Clark, H.B.; Schweitzer, J.B.; Johnson, E.M., Jr. Expression of nerve growth factor receptors by schwann cells of axotomized peripheral nerves: Ultrastructural location, suppression by axonal contact, and binding properties. J. Neurosci. 1988, 8, 664–681. [Google Scholar] [PubMed]
- Lindholm, D.; Heumann, R.; Meyer, M.; Thoenen, H. Interleukin-1 regulates synthesis of nerve growth factor in non-neuronal cells of rat sciatic nerve. Nature 1987, 330, 658–659. [Google Scholar] [CrossRef] [PubMed]
- Heumann, R.; Lindholm, D.; Bandtlow, C.; Meyer, M.; Radeke, M.J.; Misko, T.P.; Shooter, E.; Thoenen, H. Differential regulation of mrna encoding nerve growth factor and its receptor in rat sciatic nerve during development, degeneration, and regeneration: Role of macrophages. Proc. Natl. Acad. Sci. USA 1987, 84, 8735–8739. [Google Scholar] [CrossRef] [PubMed]
- Ramer, M.S.; Kawaja, M.D.; Henderson, J.T.; Roder, J.C.; Bisby, M.A. Glial overexpression of NGF enhances neuropathic pain and adrenergic sprouting into drg following chronic sciatic constriction in mice. Neurosci. Lett. 1998, 251, 53–56. [Google Scholar] [CrossRef]
- Woolf, C.J.; Safieh-Garabedian, B.; Ma, Q.P.; Crilly, P.; Winter, J. Nerve growth factor contributes to the generation of inflammatory sensory hypersensitivity. Neuroscience 1994, 62, 327–331. [Google Scholar] [CrossRef]
- Donnerer, J.; Schuligoi, R.; Stein, C. Increased content and transport of substance P and calcitonin gene-related peptide in sensory nerves innervating inflamed tissue: Evidence for a regulatory function of nerve growth factor in vivo. Neuroscience 1992, 49, 693–698. [Google Scholar] [CrossRef]
- Michael, G.J.; Averill, S.; Nitkunan, A.; Rattray, M.; Bennett, D.L.; Yan, Q.; Priestley, J.V. Nerve growth factor treatment increases brain-derived neurotrophic factor selectively in TrkA-expressing dorsal root ganglion cells and in their central terminations within the spinal cord. J. Neurosci. 1997, 17, 8476–8490. [Google Scholar] [PubMed]
- Ondarza, A.B.; Ye, Z.; Hulsebosch, C.E. Direct evidence of primary afferent sprouting in distant segments following spinal cord injury in the rat: Colocalization of GAP-43 and CGRP. Exp. Neurol. 2003, 184, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Gwak, Y.S.; Nam, T.S.; Paik, K.S.; Hulsebosch, C.E.; Leem, J.W. Attenuation of mechanical hyperalgesia following spinal cord injury by administration of antibodies to nerve growth factor in the rat. Neurosci. Lett. 2003, 336, 117–120. [Google Scholar] [CrossRef]
- Khan, N.; Smith, M.T. Multiple sclerosis-induced neuropathic pain: Pharmacological management and pathophysiological insights from rodent EAE models. Inflammopharmacology 2014, 22, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Althaus, H.H. Remyelination in multiple sclerosis: A new role for neurotrophins? Prog. Brain Res. 2004, 146, 415–432. [Google Scholar] [PubMed]
- Colafrancesco, V.; Villoslada, P. Targeting NGF pathway for developing neuroprotective therapies for multiple sclerosis and other neurological diseases. Arch. Ital. Biol. 2011, 149, 183–192. [Google Scholar] [PubMed]
- Laudiero, L.B.; Aloe, L.; Levi-Montalcini, R.; Buttinelli, C.; Schilter, D.; Gillessen, S.; Otten, U. Multiple sclerosis patients express increased levels of beta-nerve growth factor in cerebrospinal fluid. Neurosci. Lett. 1992, 147, 9–12. [Google Scholar] [CrossRef]
- Olechowski, C.J.; Truong, J.J.; Kerr, B.J. Neuropathic pain behaviours in a chronic-relapsing model of experimental autoimmune encephalomyelitis (EAE). Pain 2009, 141, 156–164. [Google Scholar] [CrossRef] [PubMed]
- Hui, J.; Zhang, Z.J.; Zhang, X.; Shen, Y.; Gao, Y.J. Repetitive hyperbaric oxygen treatment attenuates complete Freund’s adjuvant-induced pain and reduces glia-mediated neuroinflammation in the spinal cord. J. Pain 2013, 14, 747–758. [Google Scholar] [CrossRef] [PubMed]
- Raghavendra, V.; Tanga, F.Y.; DeLeo, J.A. Complete Freunds adjuvant-induced peripheral inflammation evokes glial activation and proinflammatory cytokine expression in the cns. Eur. J. Neurosci. 2004, 20, 467–473. [Google Scholar] [CrossRef] [PubMed]
- Seybold, V.S.; Galeazza, M.T.; Garry, M.G.; Hargreaves, K.M. Plasticity of calcitonin gene related peptide neurotransmission in the spinal cord during peripheral inflammation. Can. J. Physiol. Pharmacol. 1995, 73, 1007–1014. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Woodruff, T.M.; Smith, M.T. Establishment and characterization of an optimized mouse model of multiple sclerosis-induced neuropathic pain using behavioral, pharmacologic, histologic and immunohistochemical methods. Pharmacol. Biochem. Behav. 2014, 126, 13–27. [Google Scholar] [CrossRef] [PubMed]
- Obata, K.; Noguchi, K. Bdnf in sensory neurons and chronic pain. Neurosci. Res. 2006, 55, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Groth, R.; Aanonsen, L. Spinal brain-derived neurotrophic factor (BDNF) produces hyperalgesia in normal mice while antisense directed against either BDNF or TrkB, prevent inflammation-induced hyperalgesia. Pain 2002, 100, 171–181. [Google Scholar] [CrossRef]
- Wang, X.; Ratnam, J.; Zou, B.; England, P.M.; Basbaum, A.I. TrkB signaling is required for both the induction and maintenance of tissue and nerve injury-induced persistent pain. J. Neurosci. 2009, 29, 5508–5515. [Google Scholar] [CrossRef] [PubMed]
- Pezet, S.; Malcangio, M.; McMahon, S.B. Bdnf: A neuromodulator in nociceptive pathways? Brain Res. Rev. 2002, 40, 240–249. [Google Scholar] [CrossRef]
- Geranton, S.M.; Tochiki, K.K.; Chiu, W.W.; Stuart, S.A.; Hunt, S.P. Injury induced activation of extracellular signal-regulated kinase (ERK) in the rat rostral ventromedial medulla (RVM) is age dependant and requires the lamina-I projection pathway. Mol. Pain 2010, 6, 54. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.H.; Chi, X.X.; Nicol, G.D. Brain-derived neurotrophic factor enhances the excitability of rat sensory neurons through activation of the p75 neurotrophin receptor and the sphingomyelin pathway. J. Physiol. 2008, 586, 3113–3127. [Google Scholar] [CrossRef] [PubMed]
- Kerr, B.J.; Bradbury, E.J.; Bennett, D.L.; Trivedi, P.M.; Dassan, P.; French, J.; Shelton, D.B.; McMahon, S.B.; Thompson, S.W. Brain-derived neurotrophic factor modulates nociceptive sensory inputs and nmda-evoked responses in the rat spinal cord. J. Neurosci. 1999, 19, 5138–5148. [Google Scholar] [PubMed]
- Ossipov, M.H.; Dussor, G.O.; Porreca, F. Central modulation of pain. J. Clin. Investig. 2010, 120, 3779–3787. [Google Scholar] [CrossRef] [PubMed]
- Balkowiec, A.; Katz, D.M. Cellular mechanisms regulating activity-dependent release of native brain-derived neurotrophic factor from hippocampal neurons. J. Neurosci. 2002, 22, 10399–10407. [Google Scholar] [PubMed]
- Sarhan, M.; Pawlowski, S.A.; Barthas, F.; Yalcin, I.; Kaufling, J.; Dardente, H.; Zachariou, V.; DiLeone, R.J.; Barrot, M.; Veinante, P. BDNF parabrachio-amygdaloid pathway in morphine-induced analgesia. Int. J. Neuropsychopharmacol. 2013, 16, 1649–1660. [Google Scholar] [CrossRef] [PubMed]
- Eaton, M.J.; Blits, B.; Ruitenberg, M.J.; Verhaagen, J.; Oudega, M. Amelioration of chronic neuropathic pain after partial nerve injury by adeno-associated viral (AAV) vector-mediated over-expression of BDNF in the rat spinal cord. Gene Ther. 2002, 9, 1387–1395. [Google Scholar] [CrossRef] [PubMed]
- Cejas, P.J.; Martinez, M.; Karmally, S.; McKillop, M.; McKillop, J.; Plunkett, J.A.; Oudega, M.; Eaton, M.J. Lumbar transplant of neurons genetically modified to secrete brain-derived neurotrophic factor attenuates allodynia and hyperalgesia after sciatic nerve constriction. Pain 2000, 86, 195–210. [Google Scholar] [CrossRef]
- Zhang, R.; Lao, L.; Ren, K.; Berman, B.M. Mechanisms of acupuncture-electroacupuncture on persistent pain. Anesthesiology 2014, 120, 482–503. [Google Scholar] [CrossRef] [PubMed]
- Xue, C.; Xie, L.; Li, X.; Cai, J.; Gu, Z.; Wang, K. Analgesic mechanism of electroacupuncture in a rat L5 spinal nerve ligation model. Exp. Ther. Med. 2015, 9, 987–991. [Google Scholar] [CrossRef] [PubMed]
- Tu, W.; Wang, W.; Xi, H.; He, R.; Gao, L.; Jiang, S. Regulation of neurotrophin-3 and interleukin-1 and inhibition of spinal glial activation contribute to the analgesic effect of electroacupuncture in chronic neuropathic pain states of rats. Evid. Based Complement. Alternat. Med. 2014. [Google Scholar] [CrossRef]
- Kim, S.K.; Park, J.H.; Bae, S.J.; Kim, J.H.; Hwang, B.G.; Min, B.I.; Park, D.S.; Na, H.S. Effects of electroacupuncture on cold allodynia in a rat model of neuropathic pain: Mediation by spinal adrenergic and serotonergic receptors. Exp. Neurol. 2005, 195, 430–436. [Google Scholar] [CrossRef] [PubMed]
- Gim, G.T.; Lee, J.H.; Park, E.; Sung, Y.H.; Kim, C.J.; Hwang, W.W.; Chu, J.P.; Min, B.I. Electroacupuncture attenuates mechanical and warm allodynia through suppression of spinal glial activation in a rat model of neuropathic pain. Brain Res. Bull. 2011, 86, 403–411. [Google Scholar] [CrossRef] [PubMed]
- McKelvey, R.; Berta, T.; Old, E.; Ji, R.R.; Fitzgerald, M. Neuropathic pain is constitutively suppressed in early life by anti-inflammatory neuroimmune regulation. J. Neurosci. 2015, 35, 457–466. [Google Scholar] [CrossRef] [PubMed]
- Foster, E.; Robertson, B.; Fried, K. TrkB-like immunoreactivity in rat dorsal root ganglia following sciatic nerve injury. Brain Res. 1994, 659, 267–271. [Google Scholar] [CrossRef]
- Narita, M.; Yajima, Y.; Aoki, T.; Ozaki, S.; Mizoguchi, H.; Tseng, L.F.; Suzuki, T. Up-regulation of the TrkB receptor in mice injured by the partial ligation of the sciatic nerve. Eur. J. Pharmacol. 2000, 401, 187–190. [Google Scholar] [CrossRef]
- Deng, Y.S.; Zhong, J.H.; Zhou, X.F. BDNF is involved in sympathetic sprouting in the dorsal root ganglia following peripheral nerve injury in rats. Neurotox Res. 1999, 1, 311–322. [Google Scholar] [CrossRef]
- Hayashida, K.; Clayton, B.A.; Johnson, J.E.; Eisenach, J.C. Brain derived nerve growth factor induces spinal noradrenergic fiber sprouting and enhances clonidine analgesia following nerve injury in rats. Pain 2008, 136, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Yajima, Y.; Narita, M.; Usui, A.; Kaneko, C.; Miyatake, M.; Narita, M.; Yamaguchi, T.; Tamaki, H.; Wachi, H.; Seyama, Y.; et al. Direct evidence for the involvement of brain-derived neurotrophic factor in the development of a neuropathic pain-like state in mice. J. Neurochem. 2005, 93, 584–594. [Google Scholar] [CrossRef] [PubMed]
- Apfel, S.C.; Kessler, J.A. Neurotrophic factors in the treatment of peripheral neuropathy. Ciba Found. Symp. 1996, 196, 98–108, discussion 108–112. [Google Scholar] [PubMed]
- Zhou, X.F.; Rush, R.A. Endogenous brain-derived neurotrophic factor is anterogradely transported in primary sensory neurons. Neuroscience 1996, 74, 945–953. [Google Scholar] [CrossRef]
- Matsuoka, Y.; Yang, J. Selective inhibition of ERK1/2 blocks NGF to BDNF signaling and suppresses the development of and reverses already established pain behavior in rats. Neuroscience 2012, 206, 224–236. [Google Scholar] [CrossRef] [PubMed]
- Verge, V.M.K.; Andreassen, C.S.; Arnason, T.G.; Andersen, H. Mechanisms of Disease: Role of Neurotrophins in Diabetes and Diabetic Neuropathy. In Handbook of Clinical Neurology; Douglas, W.Z., Rayaz, A.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2014; Volume 126, pp. 443–460. [Google Scholar]
- Fernyhough, P.; Diemel, L.T.; Brewster, W.J.; Tomlinson, D.R. Altered neurotrophin mrna levels in peripheral nerve and skeletal muscle of experimentally diabetic rats. J. Neurochem. 1995, 64, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- Liebl, D.J.; Huang, W.; Young, W.; Parada, L.F. Regulation of Trk receptors following contusion of the rat spinal cord. Exp. Neurol. 2001, 167, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Stadelmann, C.; Kerschensteiner, M.; Misgeld, T.; Bruck, W.; Hohlfeld, R.; Lassmann, H. BDNF and gp145TrkB in multiple sclerosis brain lesions: Neuroprotective interactions between immune and neuronal cells? Brain 2002, 125, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Gielen, A.; Khademi, M.; Muhallab, S.; Olsson, T.; Piehl, F. Increased brain-derived neurotrophic factor expression in white blood cells of relapsing-remitting multiple sclerosis patients. Scand. J. Immunol. 2003, 57, 493–497. [Google Scholar] [CrossRef] [PubMed]
- Bradbury, E.J.; Khemani, S.; Von, R.; King; Priestley, J.V.; McMahon, S.B. NT-3 promotes growth of lesioned adult rat sensory axons ascending in the dorsal columns of the spinal cord. Eur. J. Neurosci. 1999, 11, 3873–3883. [Google Scholar] [CrossRef] [PubMed]
- Francis, N.; Farinas, I.; Brennan, C.; Rivas-Plata, K.; Backus, C.; Reichardt, L.; Landis, S. NT-3, like NGF, is required for survival of sympathetic neurons, but not their precursors. Dev. Biol. 1999, 210, 411–427. [Google Scholar] [CrossRef] [PubMed]
- Ivanisevic, L.; Zheng, W.; Woo, S.B.; Neet, K.E.; Saragovi, H.U. TrkA receptor “hot spots” for binding of NT-3 as a heterologous ligand. J. Biol. Chem. 2007, 282, 16754–16763. [Google Scholar] [CrossRef] [PubMed]
- Gratto, K.A.; Verge, V.M.K. Neurotrophin-3 down-regulates TrkA mRNA, NGF high-affinity binding sites, and associated phenotype in adult DRG neurons. Eur J. Neurosci. 2003, 18, 1535–1548. [Google Scholar] [CrossRef] [PubMed]
- Wright, D.E.; Zhou, L.; Kucera, J.; Snider, W.D. Introduction of a neurotrophin-3 transgene into muscle selectively rescues proprioceptive neurons in mice lacking endogenous neurotrophin-3. Neuron 1997, 19, 503–517. [Google Scholar] [CrossRef]
- Apfel, S.C.; Wright, D.E.; Wiideman, A.M.; Dormia, C.; Snider, W.D.; Kessler, J.A. Nerve growth factor regulates the expression of brain-derived neurotrophic factor mRNA in the peripheral nervous system. Mol. Cell. Neurosci. 1996, 7, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Pradat, P.F.; Kennel, P.; Naimi-Sadaoui, S.; Finiels, F.; Orsini, C.; Revah, F.; Delaere, P.; Mallet, J. Continuous delivery of neurotrophin 3 by gene therapy has a neuroprotective effect in experimental models of diabetic and acrylamide neuropathies. Hum. Gene Ther. 2001, 12, 2237–2249. [Google Scholar] [CrossRef] [PubMed]
- Mizisin, A.P.; Calcutt, N.A.; Tomlinson, D.R.; Gallagher, A.; Fernyhough, P. Neurotrophin-3 reverses nerve conduction velocity deficits in streptozotocin-diabetic rats. J. Peripher. Nerv. Syst. 1999, 4, 211–221. [Google Scholar] [PubMed]
- Boyce, V.S.; Park, J.; Gage, F.H.; Mendell, L.M. Differential effects of brain-derived neurotrophic factor and neurotrophin-3 on hindlimb function in paraplegic rats. Eur. J. Neurosci. 2012, 35, 221–232. [Google Scholar] [CrossRef] [PubMed]
- Kalinowska-Lyszczarz, A.; Pawlak, M.A.; Michalak, S.; Paprzycki, W.; Losy, J. Immune cell NT-3 expression is associated with brain atrophy in multiple sclerosis patients. J. NeuroImmunol. 2011, 240–241, 109–113. [Google Scholar] [CrossRef] [PubMed]
- Hallbook, F.; Ibanez, C.F.; Persson, H. Evolutionary studies of the nerve growth factor family reveal a novel member abundantly expressed in xenopus ovary. Neuron 1991, 6, 845–858. [Google Scholar] [CrossRef]
- Ip, N.Y.; Ibáñez, C.F.; Nye, S.H.; McClain, J.; Jones, P.F.; Gies, D.R.; Belluscio, L.; le Beau, M.M.; Espinosa, R.; Squinto, S.P. Mammalian neurotrophin-4: Structure, chromosomal localization, tissue distribution, and receptor specificity. Proc. Natl. Acad. Sci. USA 1992, 89, 3060–3064. [Google Scholar] [CrossRef] [PubMed]
- Berkemeier, L.R.; Winslow, J.W.; Kaplan, D.R.; Nikolics, K.; Goeddel, D.V.; Rosenthal, A. Neurotrophin-5: A novel neurotrophic factor that activates Trk and TrkB. Neuron 1991, 7, 857–866. [Google Scholar] [CrossRef]
- Hibbert, A.P.; Morris, S.J.; Seidah, N.G.; Murphy, R.A. Neurotrophin-4, alone or heterodimerized with brain-derived neurotrophic factor, is sorted to the constitutive secretory pathway. J. Biol. Chem. 2003, 278, 48129–48136. [Google Scholar] [CrossRef] [PubMed]
- Buck, C.R.; Seburn, K.L.; Cope, T.C. Neurotrophin expression by spinal motoneurons in adult and developing rats. J. Comp. Neurol. 2000, 416, 309–318. [Google Scholar] [CrossRef]
- Stephens, H.E.; Belliveau, A.C.; Gupta, J.S.; Mirkovic, S.; Kablar, B. The role of neurotrophins in the maintenance of the spinal cord motor neurons and the dorsal root ganglia proprioceptive sensory neurons. Int. J. Dev. Neurosci. 2005, 23, 613–620. [Google Scholar] [CrossRef] [PubMed]
- Lucas, G.; Hendolin, P.; Harkany, T.; Agerman, K.; Paratcha, G.; Holmgren, C.; Zilberter, Y.; Sairanen, M.; Minichiello, L.; Castren, E.; et al. Neurotrophin-4 mediated TRKB activation reinforces morphine-induced analgesia. Nat. Neurosci. 2003, 6, 221–222. [Google Scholar] [CrossRef] [PubMed]
- Sakuragi, S.; Tominaga-Yoshino, K.; Ogura, A. Involvement of TrkB- and p75NTR-signaling pathways in two contrasting forms of long-lasting synaptic plasticity. Sci. Rep. 2013, 3. [Google Scholar] [CrossRef] [PubMed]
- Boutilier, J.; Ceni, C.; Pagdala, P.C.; Forgie, A.; Neet, K.E.; Barker, P.A. Proneurotrophins require endocytosis and intracellular proteolysis to induce TrkA activation. J. Biol. Chem. 2008, 283, 12709–12716. [Google Scholar] [CrossRef] [PubMed]
- Peleshok, J.C.; Ribeiro-da-Silva, A. Neurotrophic factor changes in the rat thick skin following chronic constriction injury of the sciatic nerve. Mol. Pain 2012, 8, 1. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.J.; Ji, R.R. c-FOS and pERK, which is a better marker for neuronal activation and central sensitization after noxious stimulation and tissue injury? Open Pain J. 2009, 2, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Ibanez, C.F.; Simi, A. P75 neurotrophin receptor signaling in nervous system injury and degeneration: Paradox and opportunity. Trends Neurosci. 2012, 35, 431–440. [Google Scholar] [CrossRef] [PubMed]
- Tomita, K.; Kubo, T.; Matsuda, K.; Fujiwara, T.; Yano, K.; Winograd, J.M.; Tohyama, M.; Hosokawa, K. The neurotrophin receptor p75NTR in schwann cells is implicated in remyelination and motor recovery after peripheral nerve injury. Glia 2007, 55, 1199–1208. [Google Scholar] [CrossRef] [PubMed]
- Karchewski, L.A.; Kim, F.A.; Johnston, J.; McKnight, R.M.; Verge, V.M. Anatomical evidence supporting the potential for modulation by multiple neurotrophins in the majority of adult lumbar sensory neurons. J. Comp. Neurol. 1999, 413, 327–341. [Google Scholar] [CrossRef]
- Obata, K.; Katsura, H.; Sakurai, J.; Kobayashi, K.; Yamanaka, H.; Dai, Y.; Fukuoka, T.; Noguchi, K. Suppression of the p75 neurotrophin receptor in uninjured sensory neurons reduces neuropathic pain after nerve injury. J. Neurosci. 2006, 26, 11974–11986. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.F.; Li, E.; Huber, L.J.; Landis, S.C.; Sharpe, A.H.; Chao, M.V.; Jaenisch, R. Targeted mutation of the gene encoding the low affinity NGF receptor p75 leads to deficits in the peripheral sensory nervous system. Cell 1992, 69, 737–749. [Google Scholar] [CrossRef]
- Vaegter, C.B.; Jansen, P.; Fjorback, A.W.; Glerup, S.; Skeldal, S.; Richner, M.; Erdmann, B.; Nyengaard, J.R.; Tessarollo, L.; Lewin, G.R.; et al. Sortilin associates with trk receptors to enhance anterograde transport and signaling by neurotrophins. Nat. Neurosci. 2011, 14, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, D.R.; Miller, F.D. Neurotrophin signal transduction in the nervous system. Curr Opin Neurobiol. 2000, 10, 381–391. [Google Scholar] [CrossRef]
- Ji, R.R.; Kawasaki, Y.; Zhuang, Z.Y.; Wen, Y.R.; Zhang, Y.Q. Protein kinases as potential targets for the treatment of pathological pain. Handb. Exp. Pharmacol. 2007, 359–389. [Google Scholar]
- Zhuang, Z.Y.; Gerner, P.; Woolf, C.J.; Ji, R.R. Erk is sequentially activated in neurons, microglia, and astrocytes by spinal nerve ligation and contributes to mechanical allodynia in this neuropathic pain model. Pain 2005, 114, 149–159. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.T.; Tu, H.Y.; Xin, W.J.; Liu, X.G.; Zhang, G.H.; Zhai, C.H. Activation of phosphatidylinositol 3-kinase and protein kinase B/Akt in dorsal root ganglia and spinal cord contributes to the neuropathic pain induced by spinal nerve ligation in rats. Exp. Neurol. 2007, 206, 269–279. [Google Scholar] [CrossRef] [PubMed]
- Huang, E.J.; Reichardt, L.F. Trk receptors: Roles in neuronal signal transduction. Annu. Rev. Biochem. 2003, 72, 609–642. [Google Scholar] [CrossRef] [PubMed]
- Canossa, M.; Griesbeck, O.; Berninger, B.; Campana, G.; Kolbeck, R.; Thoenen, H. Neurotrophin release by neurotrophins: Implications for activity-dependent neuronal plasticity. Proc. Natl. Acad. Sci. USA 1997, 94, 13279–13286. [Google Scholar] [CrossRef] [PubMed]
- Coderre, T.J. Contribution of protein kinase c to central sensitization and persistent pain following tissue injury. Neurosci. Lett. 1992, 140, 181–184. [Google Scholar] [CrossRef]
- Reichardt, L.F. Neurotrophin-regulated signalling pathways. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2006, 361, 1545–1564. [Google Scholar] [CrossRef] [PubMed]
- Apfel, S.C.; Schwartz, S.; Adornato, B.T.; Freeman, R.; Biton, V.; Rendell, M.; Vinik, A.; Giuliani, M.; Stevens, J.C.; Barbano, R.; et al. Efficacy and safety of recombinant human nerve growth factor in patients with diabetic polyneuropathy: A randomized controlled trial. rhNGF clinical investigator group. JAMA 2000, 284, 2215–2221. [Google Scholar] [CrossRef] [PubMed]
- Cavaletti, G.; Marmiroli, P. The role of growth factors in the prevention and treatment of chemotherapy-induced peripheral neurotoxicity. Curr. Drug Saf. 2006, 1, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.T.; Murphy, F.T.; Radin, D.M.; Davignon, I.; Smith, M.D.; West, C.R. Tanezumab reduces osteoarthritic knee pain: Results of a randomized, double-blind, placebo-controlled phase-III trial. J. Pain 2012, 13, 790–798. [Google Scholar] [CrossRef] [PubMed]
- Bramson, C.; Herrmann, D.N.; Carey, W.; Keller, D.; Brown, M.T.; West, C.R.; Verburg, K.M.; Dyck, P.J. Exploring the role of tanezumab as a novel treatment for the relief of neuropathic pain. Pain Med. 2015. [Google Scholar] [CrossRef] [PubMed]
- Katz, N.; Borenstein, D.G.; Birbara, C.; Bramson, C.; Nemeth, M.A.; Smith, M.D.; Brown, M.T. Efficacy and safety of tanezumab in the treatment of chronic low back pain. Pain 2011, 152, 2248–2258. [Google Scholar] [CrossRef] [PubMed]
- Schnitzer, T.J.; Marks, J.A. A systematic review of the efficacy and general safety of antibodies to ngf in the treatment of oa of the hip or knee. Osteoarthr. Cartil. 2015, 23, S8–S17. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.T.; Herrmann, D.N.; Goldstein, M.; Burr, A.M.; Smith, M.D.; West, C.R.; Verburg, K.M.; Dyck, P.J. Nerve safety of tanezumab, a nerve growth factor inhibitor for pain treatment. J. Neurol. Sci. 2014, 345, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Danehy, S. Pfizer and Lilly Preparing to Resume Phase-3 Chronic Pain Program for Tanezumab. Available online: http://www.pfizer.com/news/press-release/press-release-detail/pfizer_and_lilly_preparing_to_resume_phase_3_chronic_pain_program_for_tanezumab (accessed on 20 April 2015).
- Lange, J. Glenmark’s Novel Monoclonal Antibody gbr 900 for Treatment of Chronic Pain Entering Human Trials. Available online: http://www.prnewswire.com/news-releases/glenmarks-novel-monoclonal-antibody-gbr-900-for-treatment-of-chronic-pain-entering-human-trials-257130311.html (accessed on 20 April 2015).
- Chaudhry, V.; Giuliani, M.; Petty, B.G.; Lee, D.; Seyedsadr, M.; Hilt, D.; Cornblath, D.R. Tolerability of recombinant-methionyl human neurotrophin-3 (r-metHuNT3) in healthy subjects. Muscle Nerve 2000, 23, 189–192. [Google Scholar] [CrossRef]
- Leinninger, G.M.; Vincent, A.M.; Feldman, E.L. The role of growth factors in diabetic peripheral neuropathy. J. Peripher. Nerv. Syst. 2004, 9, 26–53. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Khan, N.; Smith, M.T. Neurotrophins and Neuropathic Pain: Role in Pathobiology. Molecules 2015, 20, 10657-10688. https://doi.org/10.3390/molecules200610657
AMA Style
Khan N, Smith MT. Neurotrophins and Neuropathic Pain: Role in Pathobiology. Molecules. 2015; 20(6):10657-10688. https://doi.org/10.3390/molecules200610657
Chicago/Turabian StyleKhan, Nemat, and Maree T. Smith. 2015. "Neurotrophins and Neuropathic Pain: Role in Pathobiology" Molecules 20, no. 6: 10657-10688. https://doi.org/10.3390/molecules200610657