1. Introduction
Australia, like other banana-producing countries, is fighting to slow the spread of the destructive
Fusarium oxysporum f. sp.
cubense fungus (Fusarium wilt Tropical Race 4, TR4) through its AUD 583 million Cavendish-dominated banana industry [
1]. This soil-borne vascular-wilt disease has the capacity to devastate entire plantations with little prospect of recovery. At present, there is no treatment to eradicate the fungal chlamydospores which can remain dormant indefinitely in the soil or in the tissue of asymptomatic alternative host species [
2]. Currently, in Australia’s main production region of North Queensland, bananas are not permitted to be grown within 10 m of where an infected plant has been detected [
3]. No resistant Cavendish variety so far evaluated in the region has demonstrated itself to be agronomically competitive with the industry standard ‘Williams’ grown in the absence of the disease. TR4 remains a severe threat to the livelihood of the producers of Australia’s most widely consumed fruit [
4], and disease-resistant varieties are seen to be an important part of the solution in the medium to long term.
The Queensland Department of Agriculture and Fisheries (DAF) has had previous success using mutagenesis to develop the Cavendish selection ‘DPM25’, with resistance to Subtropical Race 4 (SR4) [
5], pioneering the way for future mutagenesis work within the organisation. A pilot study commenced in 2016 to better examine the potential for mutagenesis to help with developing a commercially viable TR4-resistant variety. This involved three Cavendish cultivars and Goldfinger, the latter being the sole focus of this research. Mutagenic treatments, such as gamma irradiation, have the potential to accelerate crop improvement through inducing novel allele combinations resulting in a genetically diverse population from which selections can be made [
6]. This is particularly useful for plants with a narrow genetic base and low fertility, like bananas. To date, there are four banana varieties registered with the International Atomic Energy Agency (IAEA) that have been bred through mutagenesis across various countries. These include Al-Beely (Sudan), Klue Hom Thong KU1 (Thailand), Novaria (Malaysia), and Pirama 1 (Indonesia) [
7].
Goldfinger (AAAB), also known as SH-3481 or FHIA-01, is a tetraploid hybrid which was originally developed through cross-pollinating Dwarf Prata (AAB, Pome, female parent) with the AA diploid SH-3142 (male parent). It was considered to have strong potential for commercialisation, as discussed by Rowe and Rosales [
8]. Goldfinger arrived in Australia in 1989 directly from Honduras, and subsequent field evaluations were encouraging [
9]. So, in 1995, it was commercially released as a Lady Finger (AAB, Pome) alternative for growers in the subtropics, due to its resistance to both Race 1 (R1) and SR4 of Fusarium wilt, along with its cold tolerance [
10]. Studies have also confirmed its resistance to TR4 [
11] and black Sigatoka (
Pseudocercospora fijiensis) leaf disease [
8], with less susceptibility to yellow Sigatoka (
Pseudocercospora musae) than Cavendish [
12]. Despite having sound agronomic qualities and disease resistance, Goldfinger struggled in the marketplace. Its inconsistent fruit quality, soft fruit texture, and lacklustre flavour failed to impress consumers and inhibited the variety’s commercial success [
13,
14,
15].
Goldfinger was included for mutagenesis because of its resistance to TR4. In the present study, improvement in its eating quality was sought whilst still retaining its disease resistance and other favourable features. This might then enhance its market prospects and, therefore, provide an alternative option to banana producers as the disease spreads. Goldfinger’s tolerance to yellow Sigatoka is also of potential significance to Australian banana growers, with informal observations in Phase II suggesting some plants appeared to have more leaf disease present than others. Therefore, assessing the selected variants’ reaction to this leaf disease was later included in the investigation to determine if mutagenesis had potentially impacted the plant’s level of resistance. Presented here is an overview of the project which was conducted over seven years and had multiple phases, from the initial gamma-irradiation treatment to arriving at the top four improved Goldfinger selections.
3. Results
3.1. Gamma Irradiation
The 50 and 75 Gy treatments were too high and resulted in a 100% plantlet mortality rate. There were high levels of survival in treatments of 10 and 20 Gy and little difference to the control. The 30 and 40 Gy treatments were more lethal doses, with survival at 57.5 and 55.0%, respectively, after two subculture cycles. Up to 50% of the surviving plants could subsequently be multiplied from the 30 Gy treatment, while few of the plants produced from the 40 Gy treatment were able to continue to multiply. It was therefore decided that the rate of 35 Gy would be the most appropriate for maximising mutations of the exposed Goldfinger plantlets while still inducing buds that survived and multiplied.
3.2. Morphological and Organoleptic Diversity Observed in the Goldfinger Variants
The field screening trial in Phase II revealed a substantial amount of morphological diversity throughout the irradiated plants (
Figure 1). Using the characterisations outlined by the International Plant Genetic Resources Institute [
36], Goldfinger can be described as having a slightly angled, compact bunch producing slightly curved, ridged fruit with a blunt-tipped fruit apex. The irradiation treatment impacted all aspects of the inflorescence, with a bunch angle ranging from vertical and perpendicular to the pseudostem, to horizontal. The curvature of the fruit and its size also varied and ranged from being short and wide with rounded fruit apices (gourd-shaped), to more slender with lengthily pointed ends. There was one bunch which only produced male flowers and had no fruit development altogether. The male bud of Goldfinger is described as a short heart shape with wide shoulders [
37]. Variants exhibited an array of differently shaped male buds which had the bracts attached with varying degrees of imbrication, angularity, and retention on the rachis, as well as changes to pigmentation. Distinct pigmentation differences were also observed on the pseudostem and leaf petioles of plants, variably flushed with shades of green, purple, and pink. Leaf habit was altered in some instances, and plants with more erect or drooping leaves contrasted with the more intermediate arrangement of the Goldfinger control.
A two-sample
t-test comparing planting dates revealed that the Goldfinger controls in planting 1 (September) statistically differed from the control plants in planting 2 (November) when it came to measures such as cycle time and bunch weight. It was not unexpected for differences to be present between the plantings due to seasonal climatic effects. Given the differences between the two plantings (hereon in referred to as planting 1: P1 and planting 2: P2), each planting is analysed as a separate experiment. Across both planting dates, a number of variants either died before bunch emergence (9.7%), were alive but did not produce a bunch before the completion of the trial (3.6%), developed unfavourable bunch/plant characteristics and no harvest data was collected (10.3%), or developed necrotic blotches on green fruit and were positive for banana streak OL virus (BSOLV, K. Crew pers. comm) (1.6%) (
Figure 2). All 71 unirradiated Goldfinger control plants survived and were included in the postharvest assessments.
Among the variants which produced a bunch, emergence occurred between 24 and 46 weeks after field planting and 50% had bunched within 30 weeks of being planted (
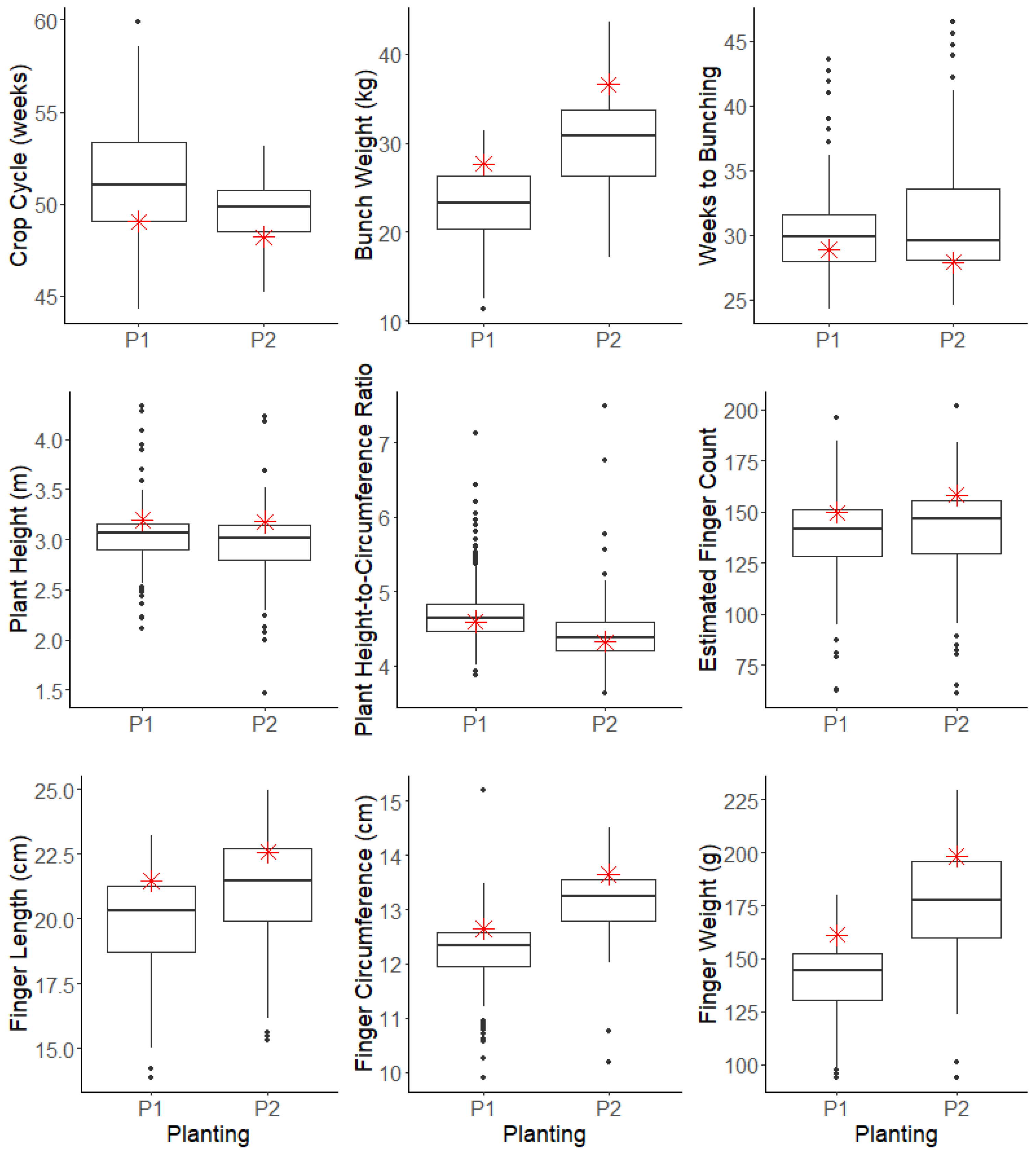
Figure 3). The mean time to the bunching of the unirradiated Goldfinger plants was one week longer in the first planting (P1 = 29 ± 0.2 weeks, P2 = 28 ± 0.3 weeks [mean ± SEM]). The spread of the variants’ crop cycle length was more condensed in P2, strongly influenced by removing any plants not ready for harvest by mid-December 2018, as their late bunch emergence was considered unfavourable for commercial production, as well as trial time constraints. Goldfinger controls in P1 had a planting-to-harvest length of 49 ± 0.4 weeks, slightly longer than those in P2 of 48 ± 0.2 weeks.
Pseudostem height and circumference measurements of the variants revealed considerable variation in relation to standard Goldfinger, which averaged a height of 3.2 m (SEMP1 = 0.05, SEMP2 = 0.04) in both planting dates. Overall, the tallest variant stood at 4.3 m, while the shortest was 1.5 m and the median height was similar in both plantings (P1 = 3.1 m, P2 = 3.0 m). The height-to-circumference ratio of Goldfinger was 4.6 ± 0.02 and 4.3 ± 0.05 in P1 and P2, respectively. In P1, 9% of variants fell within the ±1 SEM limits calculated from the 39 unirradiated Goldfinger control plants, while around half (53%) of the variants had greater pseudostem height-to-circumference ratios, and 38% had lower ratios. A similar spread was observed in the P2 variants: 53% higher, 32% lower, and the remaining 15% comparable to the unirradiated control.
The Goldfinger control bunches from the November planting (P2) were 25% heavier than those from September (P1). While procedures were in place to harvest fruit at a similar stage of maturity each week, it was, on average, harvested slightly fuller from the control plants in P2 (35 ± 0.3 mm vs. 38 ± 0.4 mm on the second proximal hand). There were also more fingers per bunch on Goldfinger control plants from P2 (159 ± 2.5 fingers) compared to P1 (150 ± 3.4 fingers). The smallest bunch recorded among all variants weighed 11.3 kg while the largest was 43.6 kg, with a median weight of 23.3 kg and 30.7 kg in P1 and P2, respectively.
Fruit size was evaluated by measuring the finger length, diameter, circumference, and weight on three fingers from the outer whorl of the second hand. For both planting dates, the majority of the fruit from the variants was smaller than that of the Goldfinger control, which was demonstrated through shorter finger lengths (77% in P1, 69% in P2) and reduced circumferences (74% in P1, 76% in P2) which subsequently weighed less (86% in P1, 73% in P2). The finger drop assessment revealed 74% of variants continued to demonstrate pedicel weakening when ripe, while 24% did not (the remaining 2% were not assessed).
In total, 72% of the 631 variants reached the postharvest assessment stage. DM and TSS were quantified as an indicator of the fruit’s sugar content. The sugar–acid ratio (TSS/TA) can give a clearer indication of the potentially perceived sweetness sensed on the tongue and consumer acceptability [
38]. A calibration error with the automatic titrator resulted in the loss of TA data for 27% of the variants involved in the postharvest assessments. Unaffected samples are incorporated into the TSS/TA ratio figure, along with the DM and TSS data (
Figure 4). Again, there was a statistical difference between the first and second planting of the Goldfinger controls when it came to fruit sensory characteristics. For example, the TSS score was 20.9 ± 0.15 °Brix in P1 and 22.5 ± 0.10 °Brix in P2. Not all the unirradiated Goldfinger bunches were included in the taste-testing sessions (P1,
n = 12; P2,
n = 4); however, the survey respondents gave the fruit in P1 a mean score of 1.51 ± 0.04 on the 3-point JAR scale for sweetness, with P2 scoring similarly at 1.67 ± 0.11.
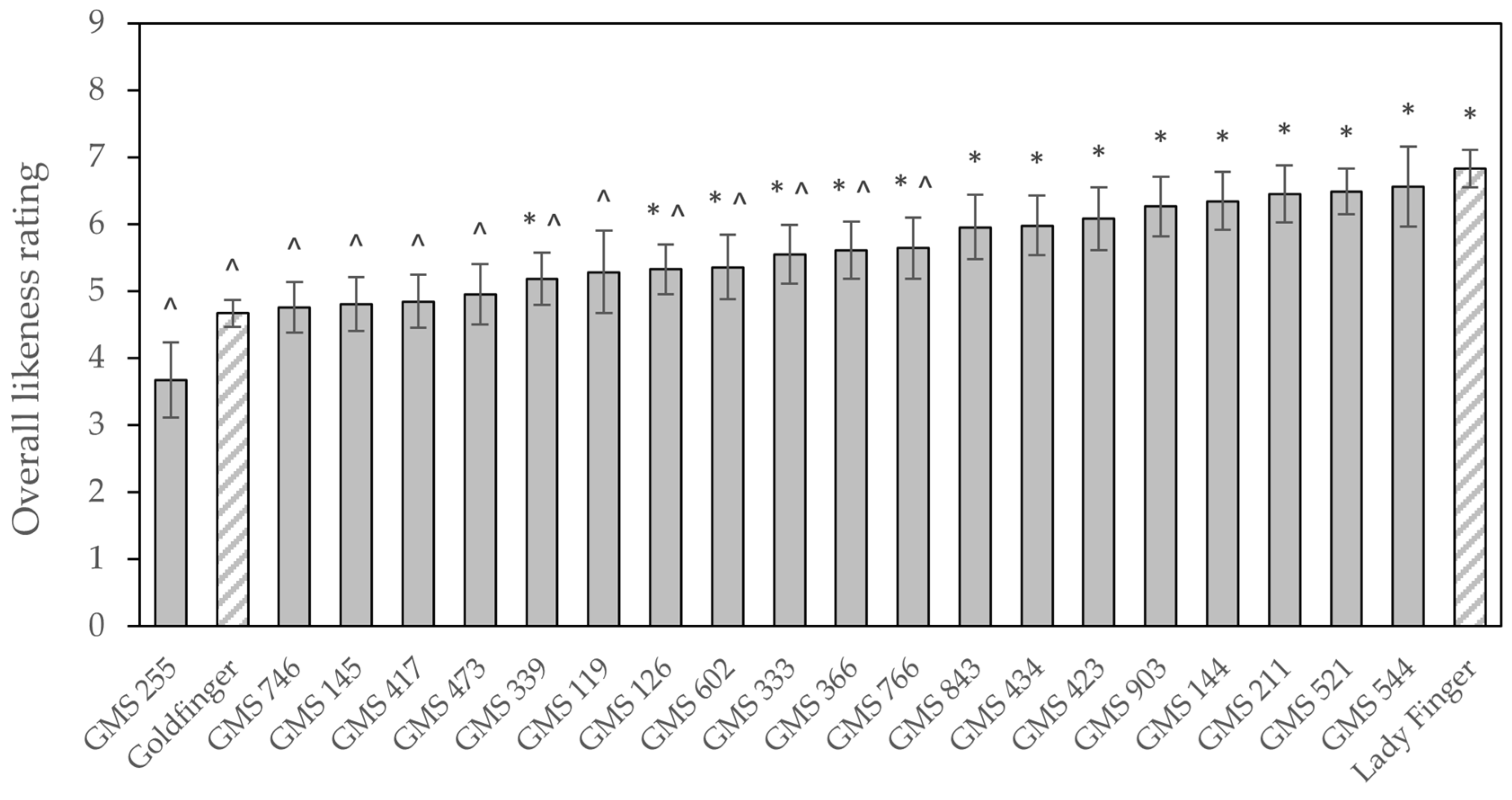
In total, 109 variants and 17 Goldfinger controls were included in 26 taste-testing sessions from July to December 2018, for which the median number of participants per session was eight. A further 291 variants were tasted in the vetting process, equating to 89% of the available variants being tasted by at least one person. The remaining fruit was excluded due to the plant displaying inferior agronomic characteristics and lacking strong commercial prospects. Like the diversity seen in the plants’ agronomic features, there was a considerable amount of variation in the fruits’ organoleptic characteristics. Goldfinger had a mean overall likeness rating of 5.5 ± 0.17; however, that varied from session to session, fluctuating between 4.1 and 6.6. For this reason, the variants were directly compared to the Goldfinger controls in their respective tasting sessions when it came to selecting the top 20 variants (data not presented).
Significant linear regressions were identified between the taste-testing feedback and the objective postharvest measurements of DM, TA, TSS, and the TSS/TA ratio. Although the relationships were statistically significant (p < 0.05), the percentage of variation explained did not exceed 12% and, therefore, is not considered to be of practical importance.
Results from the agronomic, postharvest, and taste-testing assessments identified 20 variants to progress to Phase II. Compromises were made as not all attributes outperformed Goldfinger in every category. In some instances, variants were selected because respondents found the taste to be notably ‘different’ to Goldfinger which warranted further exploration.
3.3. Evaluating the Characteristics of 20 Goldfinger Selections
The subsequent replicated trial of the top 20 selections enabled a more thorough examination of the plants’ agronomic and eating characteristics (
Supplementary Table S3 presents the agronomic data of the top 20 selections over three crop cycles). In the plant crop, bunches began emerging from the more established plants in March 2020 and the final harvest occurred in January 2021. Six of the selections (GMSs 144, 126, 417, 521, 544, and 903) had comparable bunch sizes to Goldfinger (27.3 ± 0.86 kg), while those of GMS 366 were significantly larger (33.4 ± 1.98 kg). The smallest bunch weight was from GMS 333 at 14.6 ± 1.27 kg, which was 30% smaller than the bunch observed in the original planting—an example of where the agronomic performance of the second-generation variants differed from the original parent material. When cycle time was accounted for, five of the variants (GMSs 126, 417, 521, 544, and 903) had a similar yield to Goldfinger per annum (27.0 ± 0.56 kg/12 months).
This next generation of material revealed a few undesirable characteristics which had gone undetected in the original selection of top performers. GMS 843 had several plants display severely fused fingers—to the point where whole hands in a bunch were unusable—while GMS 119 had fruit which retained a green tinge upon ripening and was unappealing to the consumer panel. The relatively tall (3.5 ± 0.07 m) and thin (54 ± 0.5 cm) pseudostem of GMS 255 contributed to 2 of the 10 plants snapping before bunch maturity; its brittle pseudostem also made harvesting difficult. Such observations undermine the commercial viability of these variants and make them undesirable candidates to pursue further.
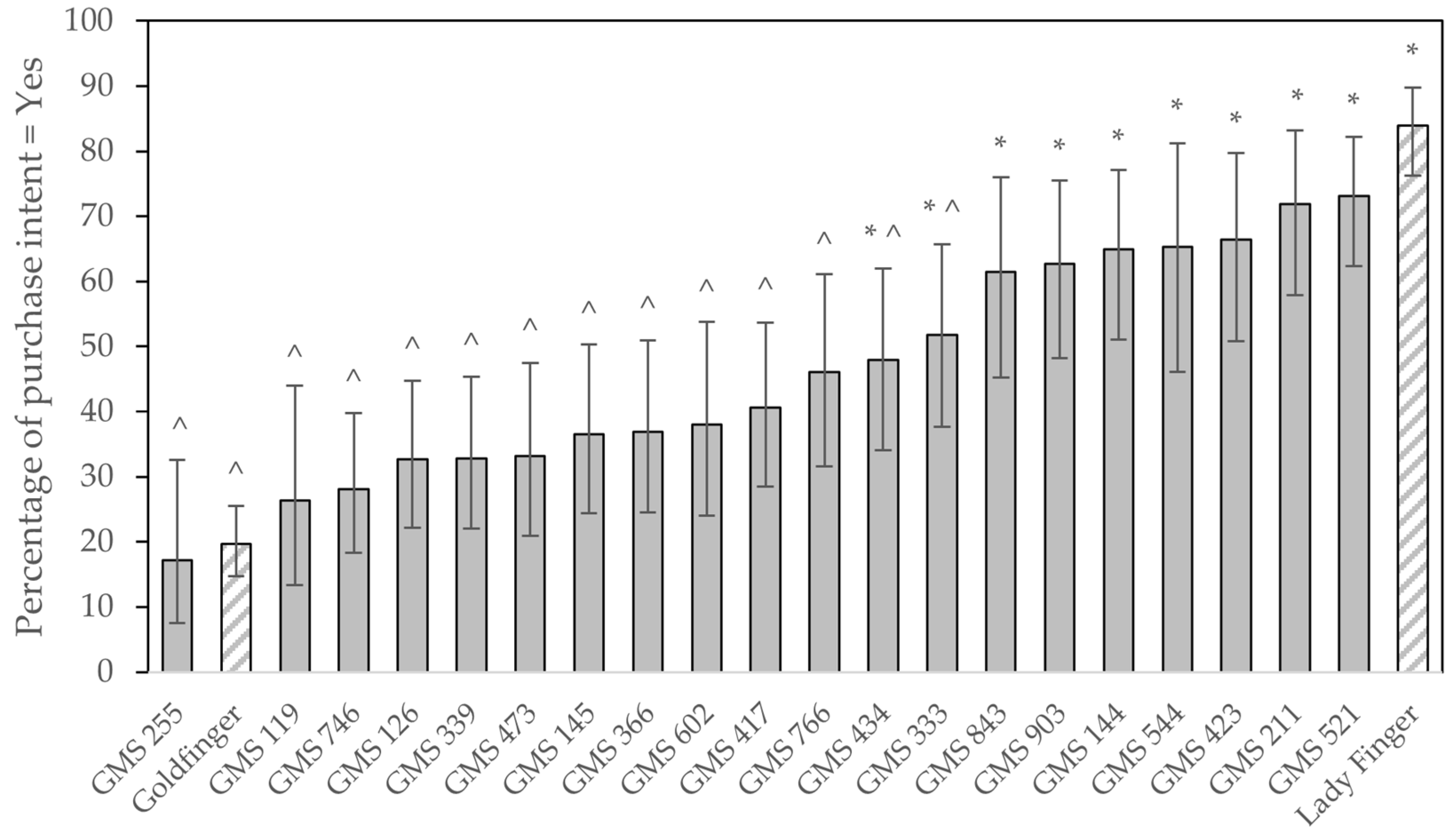
The Lady Finger comparison, ‘Dwarf Rossi’, achieved the highest mean overall likeness rating (6.8 ± 0.28) of all the varieties included in the taste panelling (
Figure 5), corresponding with 84% of respondents indicating they would purchase it if it were commercially available (
Figure 6). This was closely followed by GMS 544, with an average rating for overall likeness of 6.6 ± 0.60. Several comments were made by respondents that this variant had a nice flavour to it. The Goldfinger control rated poorly (4.7 ± 0.20), with only GMS 255 scoring below it at 3.7 ± 0.56. GMSs 211, 521, 144, and 903 joined GMS 544 in making up the five selections which respondents scored the highest for overall eating experience, and which also scored highly for the question regarding purchase intent. These variants were subsequently put forward for further investigation in the large-scale consumer and sensory evaluation. Only five selections were made due to needing to limit the number of samples sent for large-scale testing. GMS 423 was also rated highly in both survey questions; however, the plant’s cycle time was prolonged and its relatively small bunch weights meant it had a low yield per annum. This, coupled with fruit being unavailable at the correct time for the large-scale consumer and sensory panelling, meant it was not sent to Brisbane for further assessment.
Many of the initial observations in the replicated plant crop continued to present throughout the following nurse-suckered ratoon cycles. After the poor performance of GMS 255 in the plant crop and first ratoon, it was excluded from the second ratoon analysis. Of the remaining 19 selections, 6 of the variants which had produced bunches as large or heavier than Goldfinger in the plant crop had remained comparable in size by the second ratoon (but none yielded higher). Calculated from the date of nurse-suckering which includes the first and second crop cycles, seven of the variants achieved a similar yield per annum to Goldfinger. Four of these variants (GMSs 126, 366, 417, and 903) overlapped with the six that had similar bunch weights, while the other three (GMSs 119, 339 and 521) were assisted by having larger bunches in the first ratoon and slightly faster (but not significantly different) cycle times when compared to Goldfinger.
In the second ratoon, none of the variants were significantly taller than Goldfinger, but seven were shorter by between 8 and 28%. Two variants, GMSs 473 and 211, had height-to-pseudostem circumference ratios larger than Goldfinger, while the ratio for GMS 119 was smaller (meaning the plant had a stouter stature). The average finger length of fruit from the third hand was significantly shorter than Goldfinger for most variants, except for GMSs 126, 366, and 417.
The mature bunches and agronomic characteristics of the five variants selected from the top 20 are presented in
Figure 7 and
Table 2, respectively. Three of the five selections (GMSs 144, 544, and 903) had comparable bunch weights to Goldfinger in the second ratoon, averaging 34.9 ± 0.94 kg, 34.4 ± 2.47 kg, and 38.6 ± 1.46 kg, respectively. The slightly faster cycle time of GMS 521 (19.9 ± 0.003 months from nurse-suckering to second ratoon harvest), coupled with a heavier average bunch weight in the first ratoon, meant that over the two crop cycles, its average yield of 40.9 ± 0.98 kg/12 months was not significantly different to Goldfinger (46.6 ± 0.89 kg/12 months). Variant 903 was also similar, with yields of 48.5 ± 1.95 kg/12 months. With a 10% reduction in height, GMS 903 was the only one out of the five significantly shorter than Goldfinger (3.8 ± 0.10 m vs. 4.2 ± 0.02 m); however, its height-to-circumference ratio remained proportional to that of Goldfinger. Finger length was measured on the top, third, middle, and bottom hands. Across all categories, the five selections had significantly shorter finger lengths by between 8 and 18%, apart from the top hands of GMSs 544 and 903 which were not significantly different to Goldfinger (data from the third hand are presented in
Table 2).
3.4. Sensory and Consumer Analysis of Five Goldfinger Selections
To identify the key differences in fruit appearance, aroma, flavour, and texture, a sensory descriptive analysis created a lexicon of 26 attributes (three appearance, eight aroma, four texture/mouthfeel, nine flavour, and two aftertaste/afterfeel attributes) (
Supplementary Table S2). The analysis of appearance highlighted the round shape of the Cavendish and Lady Finger varieties, contrasting with the more oval shape of the remaining samples (
Figure 8—Appearance). Goldfinger and GMS 521 displayed uniformly coloured flesh, while GMSs 544, 211, 144, and 903 stood out for their glossy or moist appearance. Distinct aromatic profiles were noted (
Figure 8—Aroma). Goldfinger, Lady Finger, and GMS 211 exhibited robust sappy/resinous and sour/citrus aromas, with GMS 211 also carrying a perfumed aroma. Cavendish had an intense aroma overall, dominated by confectionary and musty notes. GMSs 144, 521, 903, and 544 presented milder aromas with hints of confectionary and tropical fruits. Regarding flavour (
Figure 8—Flavour), Cavendish displayed sweet, confectionary, and starchy notes, with subtle sour and fruity undertones. Goldfinger and Lady Finger boasted fruity, sour, and sappy/resinous flavours, with Lady Finger notably sweeter than Goldfinger. GMSs 544, 521, 144, and 903 exhibited weaker sappy/resinous and starchy flavours, with overall low flavour persistence. GMS 211 showcased strong sour and fruity flavours, possessing the most intense overall flavour profile with a sappy/resinous note. Distinct texture characteristics were also observed (
Figure 8—Texture). Cavendish featured a chalky texture, while Goldfinger and GMS 144 were characterised by soft textures. GMSs 521, 544, and 903 provided a mouth-coating sensation with medium firmness. Lady Finger and GMS 211 were lumpy and astringent, with GMS 211 also displaying a gritty texture.
The consumer analysis results indicate that GMSs 144, 521, 544, and 903 scored at parity with industry-standard bananas, Cavendish and Lady Finger, in terms of flavour, texture, and overall liking (
Table 3). Consumers particularly appreciated the sweet and sour balance in these variants. These four selections were preferred over Goldfinger (5.8 ± 0.12), which was not favoured due to its less sweet taste, stronger sappy/resin note, and chalkier texture. GMS 211 scored lower (5.4 ± 0.20) than other samples due to its intense sour and sappy/resin flavour, astringent mouthfeel, and unpleasant lumpy and gritty texture, with a small number of respondents reporting to have found one or two under-developed seeds in the fruit. For appearance, consumers rated Cavendish highest (7.1 ± 0.14), describing it as a “perfect” banana, while GMS 211 scored lowest for external appearance (5.1 ± 0.17), criticised for being too short, wide, and misshapen. Regarding aroma, all samples were liked, with Cavendish and GMSs 521 and 544 scoring the highest (6.7 ± 0.17, 6.5 ± 0.13, and 6.2 ± 0.14, respectively), and GMS 211 scoring the lowest (5.6 ± 0.16). Cavendish had the most familiar banana aroma. In texture, GMS 211 scored significantly lower than others (5.7 ± 0.20), while GMSs 521, 544, 903, and 144 scored at parity with Lady Finger and Cavendish. For flavour, GMSs 521, 544, 144, and 903 scored at parity with Lady Finger and Cavendish, while Goldfinger and GMS 211 scored significantly lower.
The analysis of JAR data, ease-of-peeling, and aroma ripeness (
Supplementary Table S4) showed preferences for various aspects including colour, shape, peel, aroma ripeness, firmness, moistness, and sweetness. External colour played a significant role in consumer preference, with Cavendish, Goldfinger, and GMSs 544 and 521 being preferred, with the peel colour reflecting ideal ripeness. On the other hand, GMSs 903, 211, 144, and Lady Finger were described as looking over-ripe by one-third of consumers. Most consumers (>70%) found the internal colour to be just right across all samples. Regarding shape and size, consumers prefer bananas that are curved and of medium size, with Cavendish exhibiting the preferred shape and size. GMS 211 and Lady Finger were considered too small, while Goldfinger was deemed too big. Additionally, one-third of consumers found Goldfinger, Lady Finger, and GMSs 144, 903, 544, 521, and 211 to be too straight. In terms of peel characteristics Cavendish and GMSs 544, 521, and 903 were easy to peel, whereas Goldfinger, Lady Finger, and GMSs 144 and 211 were more difficult. Ideal peel thickness was found in Cavendish, Lady Finger, and GMSs 144, 903, and 211. Whereas Goldfinger and GMSs 521 and 544 had peel that was considered too thick.
Of this consumer cohort, Cavendish is purchased by 96% of participants, and Lady Finger by 50% of participants The findings suggest that individuals consuming both varieties are accustomed to more diverse banana flavour attributes and are, therefore, more accustomed to the slight sourness found in some varieties. Conversely, those exclusively consuming Cavendish tend to be less receptive to such unique flavour profiles. Regular Cavendish consumers favoured GMSs 544, 144, 903, and 521 over Lady Finger, Goldfinger, and GMS 211. Regular Lady Finger consumers favoured Cavendish and GMSs 521, 544, and 903 over GMSs 144 and 211 and Goldfinger.
3.5. Screening for Yellow Sigatoka Resistance
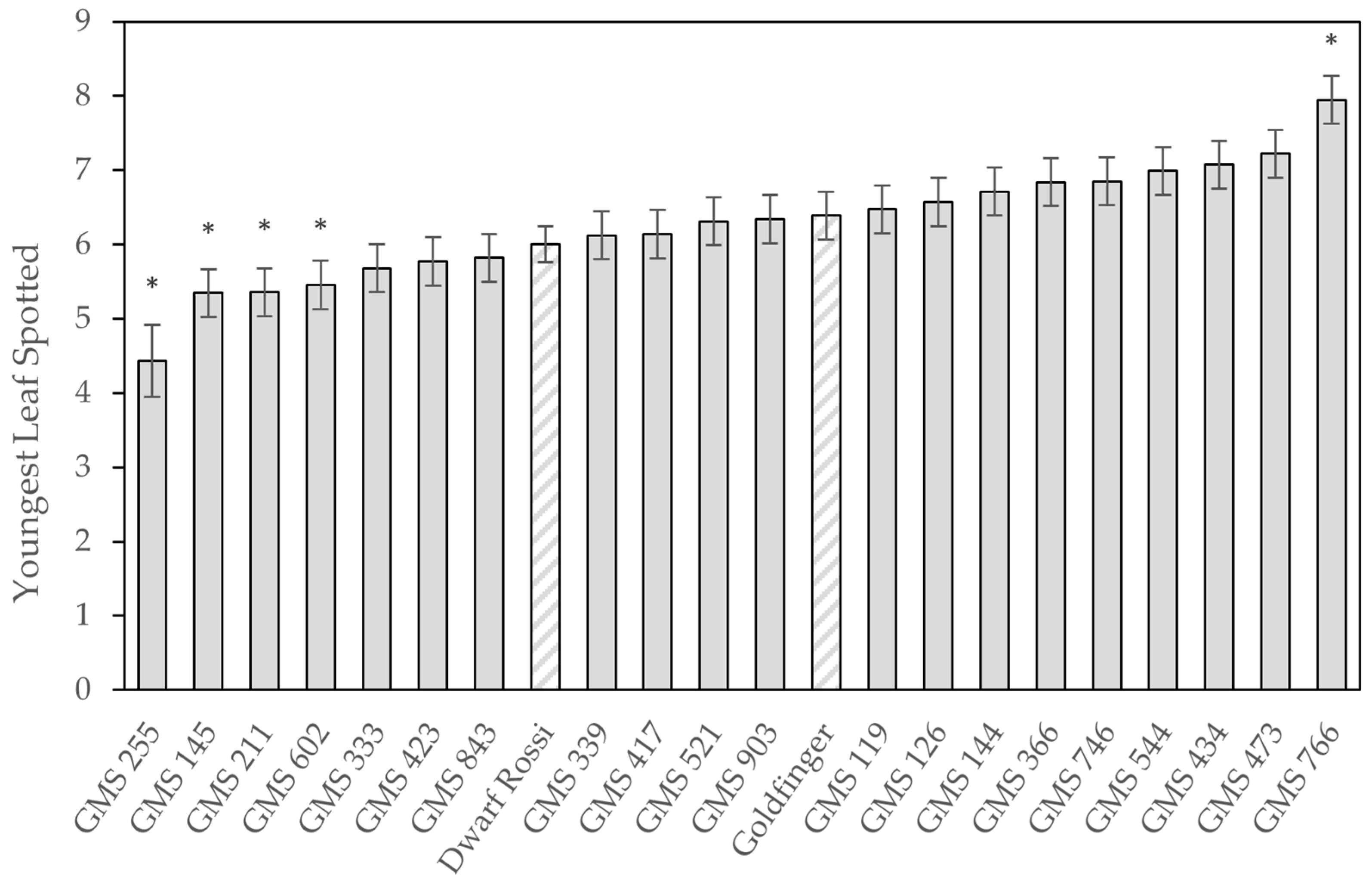
The average YLS of Goldfinger was 6.4 ± 0.32, which was similar to 15 of the 20 mutant selections (
Figure 9). GMS 766 was the only selection that demonstrated significantly better resistance to leaf spot, having a YLS of 8.0 ± 0.32, while its YL33 was also almost two leaves more than Goldfinger (11.3 ± 0.59 vs. 9.83 ± 0.59 leaves); however, the latter was not deemed significantly different. On the other end of the spectrum, GMSs 602, 211, 145, and 255 had YLS values of between 5.4 and 4.4, demonstrating they were significantly more susceptible to the disease. GMS 333 also performed poorly in the YL33 ratings, with necrotic lesions covering one-third of the leaf area from leaf 8.0 ± 0.59.
There was no significant difference between the LER for the variants, nor was there any evidence of a significant relationship between LER and the YLS. This suggests disease pressure was not compounded by slow leaf emergence. The dwarf Lady Finger selection ‘Dwarf Rossi’ was not significantly different to Goldfinger in its reaction to yellow Sigatoka.
4. Discussion
The amount of phenotypic and organoleptic variability observed in this study has demonstrated that mutagenesis can further enhance diversity and offer scope for continued improvement for banana varieties with limited commercial potential. This rapid breeding approach assists with overcoming some of the constraints faced by conventional banana breeding programs with regard to low reproductive fertility and the amount of time required from seed to seed in triploid lines [
39,
40]. It is also difficult to progress further with a tetraploid like Goldfinger using traditional breeding methods, as additional crosses risk diluting the desirable traits inherited from the triploid female parent. Such tetraploids have been described as “dead end” targets of a primary cross by Stover and Buddenhagen [
41]. Previous mutation work in
Musa spp. has focused on improving agronomic characteristics, like plant height [
42,
43] and yield [
5,
44,
45], and the disease resistance [
46,
47] of commercially relevant varieties. While these outcomes were not the primary focus of the present study, the phenotypic variation observed in the initial field planting of 631 variants (Phase II) demonstrates the 35 Gy irradiation treatment was effective in generating a population of Goldfinger mutant lines from which such improved agronomic selections could be made. For example, the plant height-to-circumference ratio can be an indicator of sturdiness. Although Goldfinger is generally considered to be a robust plant with no notable issues with pseudostem snapping or leaning [
48], just over one-third of irradiated plants had a lower height-to-circumference ratio, which, in the case of identifying variants with better wind resilience, would be an initial starting pool for further selections.
The early culling performed in vitro was an important step in reducing the amount of plant mortality seen in-field; however, it takes multiple cycles after exposure to gamma irradiation for the full effects to be seen. In tissue culture, plants may initially appear to survive but subsequently do not produce buds or eventually die. Alternatively, plants may start to produce buds which later grow abnormally or the plants which arose from the buds do not continue to multiply. The level of irradiation exposure is a balance of mutation effect, survival, and ability to produce stable buds that develop into plants that multiply. In total, 25.2% of field-planted material did not make it to harvest maturity by the completion of the trial. Along with premature plant death, late bunch emergence, and unmarketable agronomic characteristics, contributing to this was the small percentage of plants which presented with BSOLV symptoms. The genome of some BSV species is integrated into the banana B genome as activatable elements (known as endogenous viral DNA); when activated, the episomal infection can be transmitted through mealybug vectors and vegetative propagation [
49,
50]. The activation of endogenous sequences is thought to be triggered by stress events, including micropropagation via tissue culture [
51]. It has been reported in other FHIA hybrid evaluations over the years [
52] and has been found to have a transmission rate of 100% into sucker material shooting from the infected parent [
53]; therefore, the plants displaying any BSV symptoms were unsuitable for further progression in the selection process.
It is well established that planting dates can influence the subsequent phenological development of banana plants due to seasonal variations in climatic conditions [
54,
55]. The two field planting dates in Phase II were two months apart, which was found to have significantly influenced the cycle time and yield of the unirradiated Goldfinger control plants. The slightly fuller fingers on the second proximal hand of the control plants in P2 (38 ± 0.4 mm) compared to P1 (35 ± 0.3 mm) were likely influenced by more rapid fruit filling rates during periods of warmer weather. This brings to light the issue of comparing the productivity of cultivars which have not been grown under the same conditions, as well as the importance of carrying out studies over several ratoon cycles to better gauge plant performance. By making initial comparisons of the variants with the controls from their respective planting dates, and then carrying out a replicated trial originating from sucker material over three more crop cycles, there is greater confidence in the accuracy of the data presented on the mutant selections’ agronomic performance. In addition, ensuring the stability and heritability of genetic changes is an important part of crop breeding programs. In vitro mutagenesis can lead to the formation of chimeras, with plants requiring three to four generations of multiplication to allow for disassociation [
56] and the subsequent removal of any abnormal or outlying plants from analysis and further propagation. Tissue culture propagation can now more confidently be used to increase the availability of these selections for further trial work.
It is important to scrutinise commercially important traits and ensure the selected variants are acceptably productive while also appealing to consumer preferences. Early flowering and bunch yield are cited as two important production parameters selected for in banana breeding programs [
44], and it is useful to examine both in concert for crops, like bananas, which are ratooned. The average yield per annum over several cycles revealed 12 of the 19 selections (excluding GMS 255) were less productive than Goldfinger by the second ratoon through a combination of smaller bunch weights and/or longer cycle times. This included two of the top four mutant selections. However, upon the commercialisation of any new cultivar, ways to adjust production and management practices to best suit the plant’s behaviour, such as the optimum planting density, harvest criteria, and ripening protocols, need to be explored. Investigative trials on growers’ properties are currently underway to examine the commercial performance of the four Goldfinger selections, which will provide answers to some of these questions. Furthermore, the likely disease resistance offered by an improved Goldfinger selection could help reduce the costs of production while also providing a product that fulfils a market niche sought by more environmentally conscious consumers, as discussed by Roberston and Daniells [
15].
As cited earlier, mutation breeding in
Musa spp. has largely focused on manipulating agronomic features and disease resistance. No detailed postharvest assessments have been reported in the literature as it was not the targeted change sought and it was likely assumed that fruit quality had remained relatively unaffected. Arranging such a large number of unique variants to be taste-tested was arguably the biggest challenge in this project. Taste panelling is often performed on much smaller sample sizes (e.g.,
n = 8 [
57];
n = 3 [
58]); it would be unfeasible and impractical to hire a sensory panel or consumer audience for evaluating each of the 454 variants which made it to the postharvest assessment stage. Although protocols were in place to standardise factors which can influence fruit quality and eating experience—such as fruit maturity at harvest [
22], ripening and storage conditions [
59], and the stage of ripeness at assessment [
23]—the very nature of this work (to increase variation) means that each variant could potentially have a unique set of parameters suited to optimise its fruit quality. As there was only one opportunity to examine each variant, the possibility of overlooking promising candidates was an inherent risk. Despite these limitations, by prioritising fruit from plants with satisfactory agronomic characteristics over poorer performers, or those which had notable phenotypic differences in the bunch or fruit compared to Goldfinger, and by holding screening sessions to remove unpalatable fruit from further group taste-testing, there was success in identifying a selection of variants which satisfy consumer taste preferences. In 2006, a spontaneous mutation was identified in a commercial planting of Goldfinger in New South Wales, Australia. The plant’s relatively small fruit and large bunch size were noteworthy, and the ripened fruit was perceived to have a desirable flavour and texture [
60]. This variety is currently marketed as ‘Little Gem
®’ through the fresh produce supplier, Perfection Fresh Australia. This discovery emphasises the importance of ensuring a wide diversity of bunch and fruit phenotypes in the taste panelling to capture those which may have linked changes in the fruit’s eating characteristics. A link between altered plant phenotype and eating quality was also investigated by Msogoya and Grout [
58]. An off-type selection of a local cooking banana cv. ‘Uganda’ was evaluated for its performance both cooked and ripe; however, in contrast to the case of ‘Litte Gem
®’, this new selection was less favoured compared to its popular true-to-type parent.
It was originally postulated that correlations between TSS, TA, or DM and the taste-testing feedback could be used to assist with the retrospective selection of promising variants that were unable to be taste-tested. For example, Bugaud et al. [
61] identified some measurable chemical predictors of taste and texture in dessert bananas, while Akankwasa et al. [
62] identified some associations between consumer assessments (namely colour and texture) and quantitative instrumental measurements for steamed East African highland bananas. Although statistically significant relationships were identified in the present study, these relationships were weaker than those identified by Bugaud et al. [
61] and may not be of practical importance. Therefore, if possible, including a larger number of variants in the taste panelling would be recommended for future work as it remains a critical step in selecting variants which align with consumer preferences.
The findings of the sensory and consumer research highlight the importance of considering sensory attributes and consumer preferences in banana varietal development. In understanding consumer preferences for flavour, texture, appearance, and overall liking, banana producers and marketers can optimise product offerings to meet consumer demand effectively and enhance satisfaction. The four variants (GMSs 144, 521, 544, and 903) that performed well across these categories present opportunities for an improved Goldfinger selection to make its way into commercial production. However, attention should be paid to attributes such as appearance, aroma, and peelability, which also influence consumer perceptions and preferences. As apparent in the data, no sample appeared at the extreme liking end of the 9-point hedonic scale. It is important to note that when assessing food products on any structured line scale, consumers may exhibit a tendency to avoid utilising the extreme ends of the scale due to several underlying factors. Firstly, cognitive biases such as anchoring and central tendency might influence consumers’ perceptions, leading them to lean towards the middle range of the scale as a default response [
63]. Additionally, individuals may possess varying degrees of familiarity and experience with the product, in this case, different banana varieties, which could impact their ability to discern differences and as a result limit their willingness to assign extreme ratings [
64]. Furthermore, sociocultural influences and personal preferences may shape consumers’ expectations and perceptions, potentially leading them to avoid extreme ratings to conform to perceived norms or avoid perceived social risks associated with unconventional preferences [
65]. These psychological and sociocultural dynamics underscore the complex nature present in consumer hedonic evaluations and may warrant further investigation to better understand consumer behaviour in fruit preference assessment.
While Goldfinger has resistance to black Sigatoka, this is not the case for yellow Sigatoka, to which it is better described as having an intermediate reaction [
66]. Of particular interest was how the new selections compared to Goldfinger and the possible effect mutagenesis may have had on the plants’ reaction to this disease. This screening work was initially not included in the project; however, observations were made during Phase II (after all bunches had been harvested and leaf spot control had ceased) that some plants appeared more symptomatic than others. The variation among the top 20 selections demonstrates that mutagenesis can significantly change a plant’s level of yellow Sigatoka resistance. This result supports the need to now screen the top four selections to ensure their TR4, SR4, and R1 resistance have not been compromised. Mutation breeding has purportedly been used to introduce tolerance to Fusarium wilt into susceptible varieties [
46], so it is plausible that it may also have the opposite effect. Resistance to one strain of Fusarium is not always associated with resistance to another, and it is therefore important to evaluate against them independently. GMSs 144, 521, 544, and 903 were recently planted on two R1-infected farms on the Atherton Tablelands, Queensland, with their disease susceptibility soon to be determined. The outcome of the screening trial will be of major significance to growers in parts of Australia, and the world, where R1 is present but which are not yet impacted by TR4. Making up around 4% of Australian banana production are niche varieties, like Lady Finger and Ducasse (ABB, Pisang Awak), which readily succumb to R1 [
2,
67]. As the disease continues to spread through the regions where these varieties are grown, farmers are seeking to adopt resistant alternatives to continue production and help secure their livelihoods. Screening the improved selections against TR4 and SR4 is yet to be performed but is a top priority.
5. Conclusions
This is the first banana mutagenesis study that has focused on improving the eating characteristics of a variety, rather than agronomic features or disease resistance. Assessing the consumer acceptability of over 450 mutant plants was a significant challenge, but 4 selections preferred by consumers over the Goldfinger control were able to be identified during this project. Instrumental high-throughput screening is still limited to detecting banana variants which appeal to consumer preferences, leaving taste-testing as an important part of the selection process for now.
The implications of this study reach further than the development of a commercially acceptable Fusarium wilt-resistant banana variety. It has also demonstrated that a huge amount of diversity can be created in both appearance and taste within a relatively short timeframe, highlighting the potential mutagenesis has for other disease-resistant tetraploid banana varieties which have their commercial prospects limited by inferior taste or agronomic qualities.
The relatively rapid development of new plant varieties through biotechnology-facilitated improvements, like mutagenesis, will only become more important in improving crop resilience in the face of a changing climate [
68]. Worldwide, more than 3400 mutation-derived plant varieties have contributed to global agriculture production by increasing the yield and quality of the crop [
7,
69]. One, or all, of the best-performing Goldfinger selections could eventually join the four other registered banana varieties developed through mutagenesis to become a Fusarium wilt-resistant variety with commercially acceptable agronomic and taste qualities.
The next phase of this work is currently underway to confirm that the four selected variants have retained their Fusarium wilt resistance and to further assess their commercial performance across various production regions in Australia. Developing an increased understanding of the underlying genetic changes which have arisen from the mutagenesis treatment is also being pursued through an upcoming collaboration with Murdoch University in Western Australia. This knowledge aims to further assist banana plant improvement endeavours.


 ,
, 

 .
.

 .
.







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}