


Phytoene Desaturase (PDS) Gene-Derived Markers Identify “A” and “B” Genomes in Banana (Musa spp.)

 , , ,
, , ,  , , and
, , and
Abstract
:1. Introduction
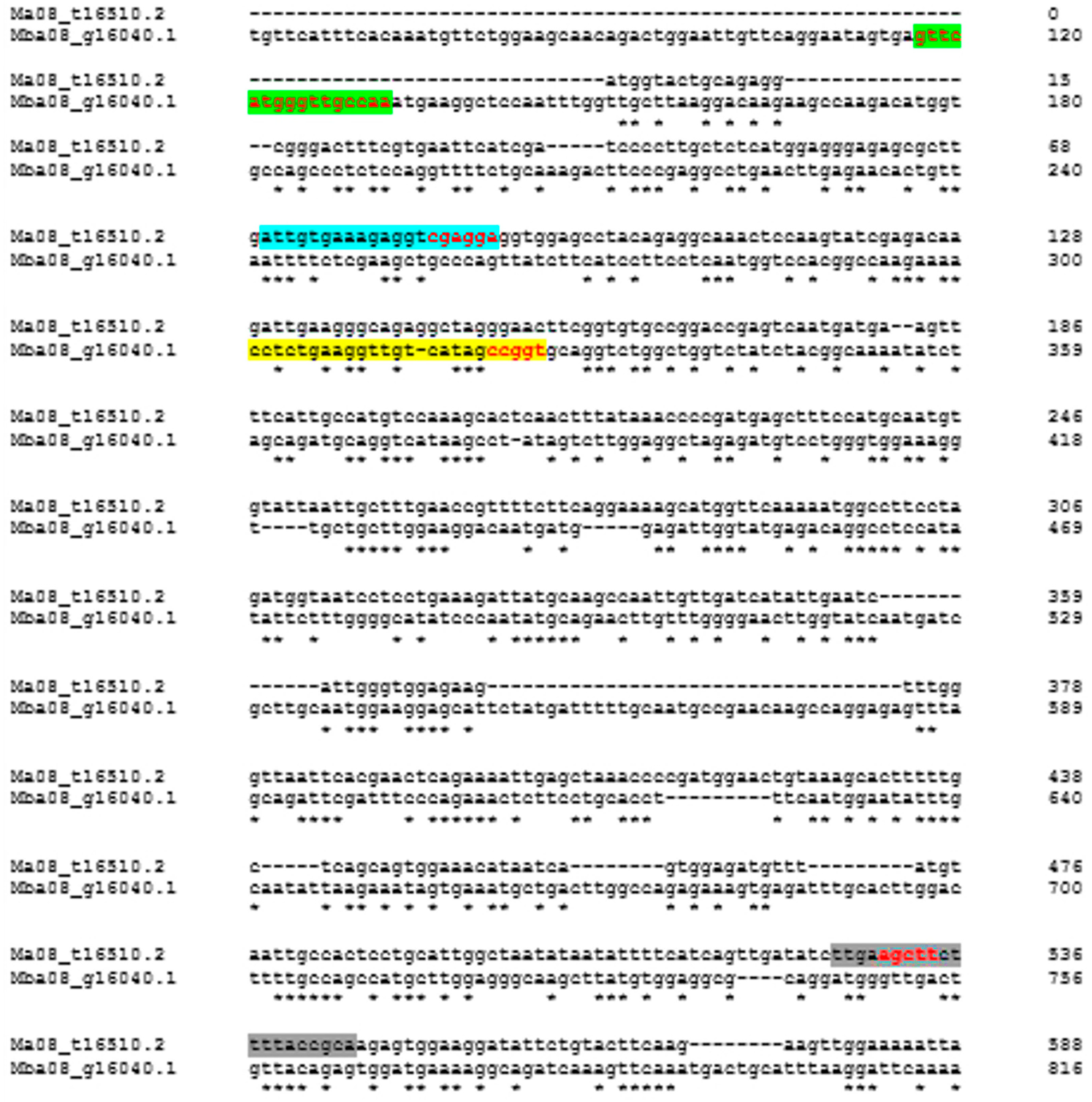
2. Materials and Methods
2.1. Plant Material
2.2. Primer Construction
2.3. DNA Extraction and PCR Conditions
2.4. Data Analysis
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAOSTAT. Food and Agriculture Organization of the United Nations. Available online: http://www.fao.org/faostat/en/#home (accessed on 10 August 2023).
- Simmonds, N.W.; Shepherd, K. The taxonomy and origins of the cultivated bananas. J. Linean Soc. Lond. 1955, 55, 302–312. [Google Scholar] [CrossRef]
- D’hont, A.; Denoeud, F.; Aury, J.M.; Baurens, F.C.; Carreel, F.; Garsmeur, O.; Noel, B.; Bocs, S.; Droc, G.; Rouard, M.; et al. The banana (Musa acuminata) genome and the evolution of monocotyledonous plants. Nature 2012, 488, 213–217. [Google Scholar] [CrossRef] [PubMed]
- Martin, G.; Baurens, F.C.; Droc, G.; Rouard, M.; Cenci, A.; Kilian, A.; Hastie, A.; Doležel, J.; Aury, J.M.; Alberti, A.; et al. Improvement of the banana “Musa acuminata” reference sequence using NGS data and semi-automated bioinformatics methods. BMC Genom. 2016, 17, 243. [Google Scholar] [CrossRef]
- Davey, M.W.; Gudimella, R.; Harikrishna, J.A.; Sin, L.W.; Khalid, N.; Keulemans, J. A draft Musa balbisiana genome sequence for molecular genetics in polyploid, inter-and intra-specific Musa hybrids. BMC Genom. 2013, 14, 683. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Miao, H.; Liu, J.; Xu, B.; Yao, X.; Xu, C.; Zhang, J. Musa balbisiana genome reveals subgenome evolution and functional divergence. Nat. Plants 2019, 5, 810–821. [Google Scholar] [CrossRef]
- Geering, A.D.W.; Mcmichael, L.A.; Dietzgen, R.G.; Thomas, J.E. Genetic diversity among Banana streak virus isolates from Australia. Phytopathology 2000, 90, 921–927. [Google Scholar] [CrossRef]
- Gayral, P.; Noa-Carrazana, J.C.; Lescot, M.; Lheureux, F.; Lockhart, B.E.L.; Matsumoto, T.; Piffanelli, P.; Iskra-Caruana, M.L. A Single Banana Streak Virus Integration Event in the Banana Genome as the Origin of Infectious Endogenous Pararetrovirus. J. Virol. 2008, 82, 6697–6710. [Google Scholar] [CrossRef]
- Chabannes, M.; Baurens, F.C.; Duroy, P.O.; Bocs, S.; Vernerey, M.S.; Goud, M.R.; Barbe, V.; Gayral, P.; Iskra-Caruana, M.L. Three infectious viral species lying in wait in the Banana Genome. J. Virol. 2013, 87, 862–863. [Google Scholar] [CrossRef]
- Dallot, S.; Acuna, P.; Rivera, C.; Ramírez, P.; Cōte, F.; Lockhart, B.E.; Caruana, M.L. Evidence that the proliferation stage of micropropagation procedure is determinant in the expression of Banana streak virus integrated into the genome of the FHIA 21 hybrid (Musa AAAB). Arch. Virol. 2001, 146, 2179–2190. [Google Scholar] [CrossRef]
- Côte, F.X.; Galzi, S.; Folliot, M.; Lamagnère, Y.; Teycheney, P.Y.; Iskra-Caruana, M.L. Micropropagation by tissue culture triggers differential expression of infectious endogenous Banana streak virus sequences (eBSV) present in the B genome of natural and synthetic interspecific banana plantains. Mol. Plant Pathol. 2010, 11, 137–144. [Google Scholar] [CrossRef]
- Perrier, X.; Bakry, F.; Carreel, F.; Jenny, C.; Horry, J.P.; Lebot, V.; Hippolyte, I. Combining biological approaches to shed light on evolution of edible bananas. Ethnobot. Res. Appl. 2009, 7, 199–216. [Google Scholar] [CrossRef]
- Jesus, O.N.; Amorim, E.P.; Ferreira, C.F.; Campos, J.M.S.; Silva, G.D.G.; Figueira, A. Genetic diversity and population structure of Musa accessions in ex situ conservation. BMC Plant Biol. 2013, 13, 41. [Google Scholar] [CrossRef]
- Santos-Serejo, J.A.; Amorim, E.P.; Jesus, O.N.; Silva, S.O. Germoplasma de Musa, Conservação, Caracterização e Uso. In O Agronegócio da Banana; Ferreira, C.F., Silva, S.O., Amorim, E.P., Santos-Serejo, J.A., Eds.; Embrapa: Brasília, Brazil, 2016; pp. 113–136. [Google Scholar]
- Nwakanma, D.C.; Pillay, M.; Okoli, B.E.; Tenkouano, A. PCR-RFLP of the ribosomal DNA internal transcribed spacers (ITS) provides markers for the A and B genomes in Musa L. Theor. Appl. Genet. 2003, 108, 154–159. [Google Scholar] [CrossRef]
- Perrier, X.; Du Montcel, H.T. Musaid: A Computerized Determination System. In Proceedings of Identification of Genetic Diversity in the Genus Musa; Los Banos (PHL); Jarret, R., Ed.; INIBAP: Montpellier, France, 1990; pp. 76–91. [Google Scholar]
- Pillay, M.; Nwakanma, D.C.; Tenkouano, A. Identification de RAPD markers linked to A and B genome sequences in Musa L. Genome 2000, 43, 763–767. [Google Scholar] [CrossRef]
- Bhat, J.A.; Ali, S.; Salgotra, R.K.; Mir, Z.A.; Dutta, S.; Jadon, V.; Tyagi, A.; Mushtaq, M.; Jain, N.; Singh, G.P.; et al. Genomic selection in the era of next generation sequencing for complex traits in plant breeding. Front. Genet. 2016, 7, 221. [Google Scholar] [CrossRef]
- Molinari, H.B.C.; Vieira, L.R.; Silva, N.; Prado, G.S.; Lopes Filho, J.H. Tecnologia CRISPR na Genômica de Plantas: Biotecnologia Aplicada à Agricultura; Embrapa: Brasília, Brazil, 2020; p. 207. [Google Scholar]
- Naim, F.; Dugdale, B.; Kleidon, J.; Brinin, A.; Shand, K.; Waterhouse, P.; Dale, J. Gene editing the phytoene desaturase alleles of Cavendish banana using CRISPR/Cas9. Transgenic Res. 2018, 27, 451–460. [Google Scholar] [CrossRef] [PubMed]
- Kaur, N.; Alok, A.; Kaur, N.; Pandey, P.; Awasthi, P.; Tiwari, S. CRISPR/Cas9-mediated efficient editing in phytoene desaturase (PDS) demonstrates precise manipulation in banana cv. Rasthali genome. Funct. Integr. Genom. 2018, 18, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, L.; Ntui, V.O.; Tripathi, J.N. Application of genetic modification and genome editing for developing climate-smart banana. Food Energy Secur. 2019, 8, e00168. [Google Scholar] [CrossRef]
- Bai, C.; Capell, T.; Berman, J.; Medina, V.; Sandmann, G.; Christou, P.; Zhu, C. Bottlenecks in carotenoid biosynthesis and accumulation in rice endosperm are influenced by the precursor product balance. Plant Biotechnol. J. 2016, 14, 195–205. [Google Scholar] [CrossRef]
- Wang, M.; Wang, G.; Ji, J.; Wang, J. The effect of pds gene silencing on chloroplast pigment composition, thylakoid membrane structure and photosynthesis efficiency in tobacco plants. Plant Sci. 2009, 177, 222–226. [Google Scholar] [CrossRef]
- Koschmieder, J.; Fehling-Kaschek, M.; Schaub, P.; Ghisla, S.; Brausemann, A.; Timmer, J.; Beyer, P. Plant-type phytoene desaturase: Functional evaluation of structural implications. PLoS ONE 2017, 12, e0187628. [Google Scholar] [CrossRef]
- Ntui, V.O.; Tripathi, J.N.; Tripathi, L. Robust CRISPR/Cas9 mediated genome editing tool for banana and plantain (Musa spp.). Curr. Plant Biol. 2020, 21, 100128. [Google Scholar] [CrossRef]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef] [PubMed]
- Doyle, J.J.; Doyle, J.L. Isolation of plant DNA from fresh tissue. Focus 1990, 12, 13–15. [Google Scholar]
- Ferreira, C.F.; Gutierrez, D.; Kreuze, J.; Iskra Caruana, M.L.; Chabannes, M.; Barbosa, A.C.O.; Santos, T.A.; Silva, A.G.S.; Santos, R.M.F.; Amorim, E.P.; et al. Rapid plant DNA and RNA extraction protocol using a bench drill. GMR 2019, 18, gmr18394. [Google Scholar] [CrossRef]
- Podevin, N.; Krauss, A.; Henry, I.; Swennen, R.; Remy, S. Selection and validation of reference genes for quantitative RT-PCR expression studies of the non-model crop Musa. Mol. Breed. 2012, 30, 1237–1252. [Google Scholar] [CrossRef] [PubMed]
- MedCalc Software Ltd. Diagnostic Test Evaluation Calculator. Version 22.009. Available online: https://www.medcalc.org/calc/diagnostic_test.php (accessed on 4 August 2023).
- Matthews, P.D.; Luo, R.; Wurtzel, E.T. Maize phytoene desaturase and ζ-carotene desaturase catalyse a poly-Z desaturation pathway: Implications for genetic engineering of carotenoid content among cereal crops. J. Exp. Bot. 2003, 54, 2215–2230. [Google Scholar] [CrossRef] [PubMed]
- Qin, G.; Gu, H.; Ma, L.; Peng, Y.; Deng, X.W.; Chen, Z.; Qu, L.J. Disruption of phytoene desaturase gene results in albino and dwarf phenotypes in Arabidopsis by impairing chlorophyll, carotenoid, and gibberellin biosynthesis. Cell Res. 2007, 17, 471–482. [Google Scholar] [CrossRef] [PubMed]
- Naing, A.H.; Kyu, S.Y.; Pe, P.P.W.; Park, K.I.; Lee, J.M.; Lim, K.B.; Kim, C.K. Silencing of the phytoene desaturase (PDS) gene affects the expression of fruit-ripening genes in tomatoes. Plant Methods 2019, 15, 110. [Google Scholar] [CrossRef]
- Banakar, R.; Schubert, M.; Collingwood, M.; Vakulskas, C.; Eggenberger, A.L.; Wang, K. Comparison of CRISPR-Cas9/Cas12a ribonucleoprotein complexes for genome editing efficiency in the rice phytoene desaturase (OsPDS) gene. Rice 2020, 13, 4. [Google Scholar] [CrossRef]
- Silva, S.O.; Santos-Serejo, J.A.; Amorim, E.P. Pré-Melhoramento da Banana. In Pré-Melhoramento de Plantas: Estado da Arte e Experiências de Sucesso; Lopes, M.A., Fávero, A.P., Ferreira, M.A.J.F., Faleiro, F.G., Folle, S.M., Guimarães, E.P., Eds.; Embrapa Informação Tecnológica: Brasília, Brazil, 2011; pp. 317–350. [Google Scholar]
- Amorim, E.P.; Santos-Serejo, J.A.; Amorim, V.B.O.; Silva, S.O. Melhoramento Genético. In O agronegócio da Banana; Ferreira, C.F., Silva, S.O., Amorim, E.P., Santos-Serejo, J.A., Eds.; Embrapa: Brasília, Brazil, 2016; pp. 171–200. [Google Scholar]
- Roux, N.; Dolezel, J.; Swennen, R.; Zapata-Arias, F.J. Effectiveness of three micropropagation techniques to dissociate cytochimeras in Musa spp. Plant Cell Tissue Organ Cult. 2001, 66, 189–197. [Google Scholar] [CrossRef]
- De Langhe, E.; Hřibová, E.; Carpentier, S.; Doležel, J.; Swennen, R. Did backcrossing contribute to the origin of hybrid edible bananas? Ann. Bot. 2010, 106, 849–857. [Google Scholar] [CrossRef] [PubMed]
- Cenci, A.; Sardos, J.; Hueber, Y.; Martin, G.; Breton, C.; Roux, N.; Swennen, R.; Carpentier, S.C.; Rouard, M. Unravelling the complex story of intergenomic recombination in ABB allotriploid bananas. Ann. Bot. 2021, 127, 7–20. [Google Scholar] [CrossRef]
- Dhivya, S.; Ashutosh, S.; Gowtham, I.; Baskar, V.; Harini, A.B.; Mukunthakumar, S.; Sathishkumar, R. Molecular identification and evolutionary relationships between the subspecies of Musa by DNA barcodes. BMC Genom. 2020, 21, 659. [Google Scholar] [CrossRef]
- Hollingsworth, P.M. Refining the DNA barcode for land plants. Proc. Natl. Acad. Sci. USA 2011, 108, 19451–19452. [Google Scholar] [CrossRef]
- Mabonga, L.; Pillay, M. SCAR Marker for the A Genome of Bananas (Musa spp. L.) Supports Lack of Differentiation between the A and B Genomes. J. Agric. Sci. 2017, 9, 64. [Google Scholar] [CrossRef]
- Nadeem, M.A.; Nawaz, M.A.; Shahid, M.Q.; Doğan, Y.; Comertpay, G.; Yıldız, M.; Hatipoglu, R.; Ahmadg, F.; Alsalehh, A.; Labhane, N.; et al. DNA molecular markers in plant breeding: Current status and recent advancements in genomic selection and genome editing. Biotechnol. Biotechnol. Equip. 2018, 32, 261–285. [Google Scholar] [CrossRef]
- Iskra-Caruana, M.L.; Baurens, F.C.; Gayral, P.; Chabannes, M. A four-partner plant–virus interaction: Enemies can also come from within. Mol. Plant Microbe Interact. 2010, 23, 1394–1402. [Google Scholar] [CrossRef]
- Noumbissié, G.B.; Chabannes, M.; Bakry, F.; Ricci, S.; Cardi, C.; Njembele, J.C.; Yohoume, D.; Tomekpe, K.; Iskra-Caruana, M.-L.; D’Hont, A.; et al. Chromosome segregation in an allotetraploid banana hybrid (AAAB) suggests a translocation between the A and B genomes and results in eBSV-free offsprings. Mol. Breed. 2016, 36, 38. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Sequence | |||||
|---|---|---|---|---|---|
| Primers | F (5′-3′) | R (5′-3′) | At | pb | Reference |
| β-tubulin | ACATTGTCAGGT GGG GAGTT | CCTTTTGTTCCACACGAGATT | 60 °C | 110 | [30] |
| PDS_AB | CAGCTAACTGAGATCAGTTT | AGATGGCTATATTTCGGTAC | 55 °C | 994 | [26] |
| PDSMa | ATTGTGAAAGAGGTCGAGGA | TGCGGTAAAAGAAGCTTCAA | 60 °C | 2166 | |
| PDSMb | GTGAGTTCATGGGTTGCCAA | ACCGGCTATGACAACCTTCA | 65 °C | 332 | |
| Statistic | Value-PDSMa | Value-PDSMb | 95% CI-PDSMa | 95% CI-PDSMb |
|---|---|---|---|---|
| Sensitivity | 99.32% | 100.00% | 96.24% to 99.98% | 96.07% to 100.00% |
| Specificity | 100.00% | 100.00% | 39.76% to 100.00% | 93.84% to 100.00% |
| Positive likelihood ratio | ||||
| Negative likelihood ratio | 0.01 | 0.00 | 0.00 to 0.05 | |
| Disease prevalence | 97.33% | 61.33% | 93.31% to 99.27% | 53.05% to 69.16% |
| Positive predictive value | 100.00% | 100.00% | 96.07% to 100.00% | |
| Negative predictive value | 80.00% | 100.00% | 36.19% to 96.58% | 93.84% to 100.00% |
| Accuracy | 99.33% | 100.00% | 96.34% to 99.98% | 97.57% to 100.00% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nascimento, F.d.S.; Mascarenhas, M.S.; Boaventura, S.C.; de Souza, C.C.H.; de Souza Ramos, A.P.; de Jesus Rocha, A.; da Silva Soares, J.M.; Diniz, L.E.C.; de Oliveira Mendes, T.A.; Ferreira, C.F.; et al. Phytoene Desaturase (PDS) Gene-Derived Markers Identify “A” and “B” Genomes in Banana (Musa spp.). Horticulturae 2024, 10, 294. https://doi.org/10.3390/horticulturae10030294
Nascimento FdS, Mascarenhas MS, Boaventura SC, de Souza CCH, de Souza Ramos AP, de Jesus Rocha A, da Silva Soares JM, Diniz LEC, de Oliveira Mendes TA, Ferreira CF, et al. Phytoene Desaturase (PDS) Gene-Derived Markers Identify “A” and “B” Genomes in Banana (Musa spp.). Horticulturae. 2024; 10(3):294. https://doi.org/10.3390/horticulturae10030294
Chicago/Turabian StyleNascimento, Fernanda dos Santos, Marcelly Santana Mascarenhas, Samantha Costa Boaventura, Carla Catharina Hora de Souza, Andresa Priscila de Souza Ramos, Anelita de Jesus Rocha, Julianna Matos da Silva Soares, Leandro Eugenio Cardamone Diniz, Tiago Antônio de Oliveira Mendes, Claudia Fortes Ferreira, and et al. 2024. "Phytoene Desaturase (PDS) Gene-Derived Markers Identify “A” and “B” Genomes in Banana (Musa spp.)" Horticulturae 10, no. 3: 294. https://doi.org/10.3390/horticulturae10030294