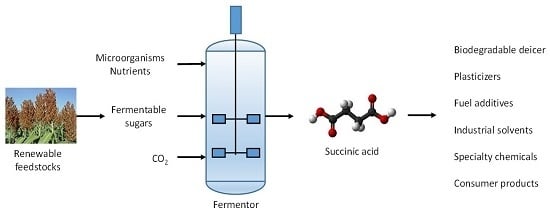
2. Succinic Acid-Producing Microorganisms
Succinic acid lends itself to biological production, because it is part of every organism’s central metabolism. This was recognized decades ago, when the US DOE ranked succinic acid, along with other dicarboxylic acids, on its top value-added chemicals list [
5], and sparked the isolation and development of multiple microorganisms for succinic acid production. The various routes to succinic acid are summarized in
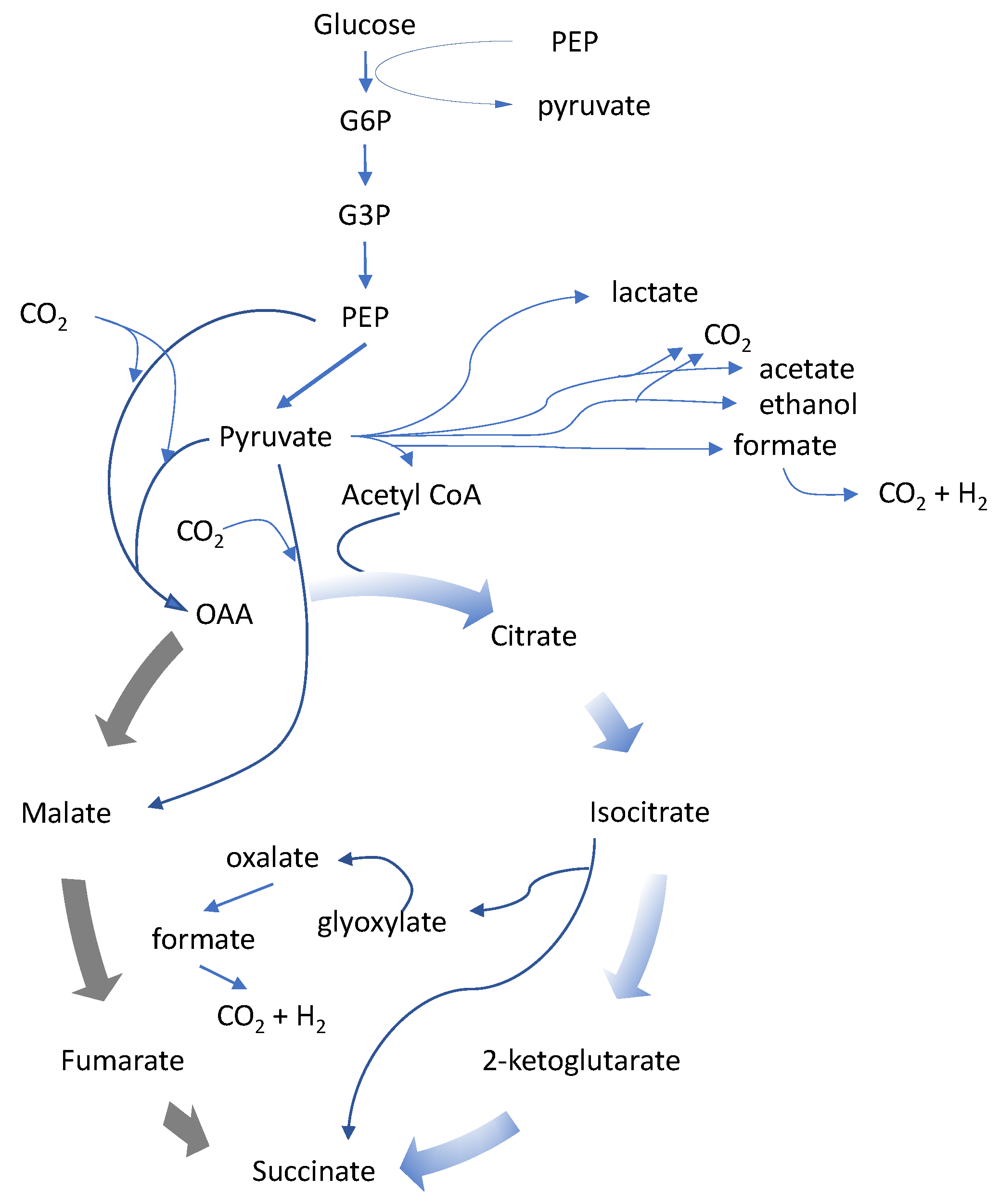
Figure 1.
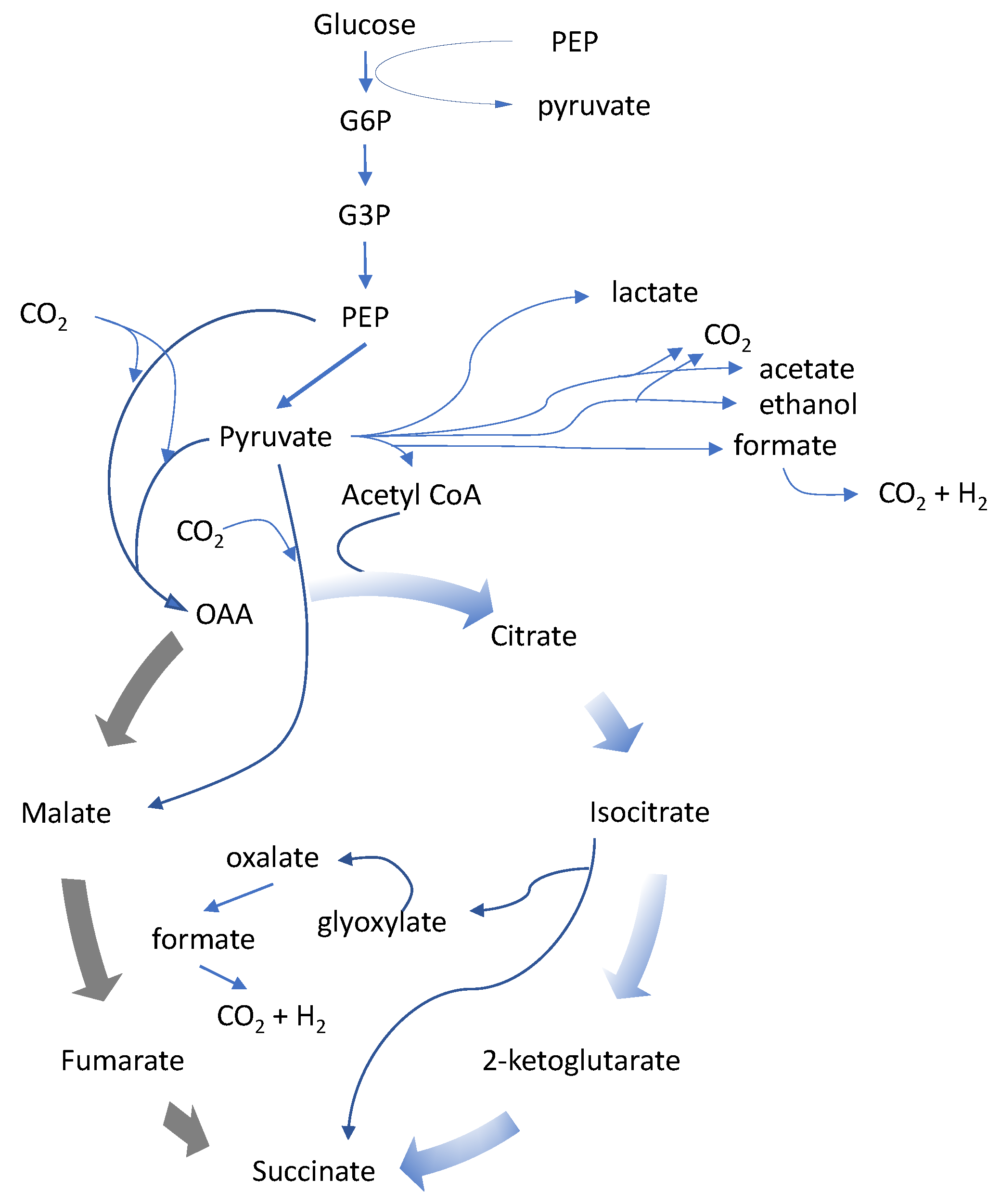
Succinic acid is a metabolite of the tricarboxylic acid (TCA) cycle. The use of the reductive direction of the TCA cycle for succinic acid production is appealing, and can theoretically produce two molecules of succinic from each molecule of a 6-carbon sugar with incorporation of 2 carbon dioxide molecules in the conversion of phosphoenolpyruvate (PEP) to oxaloacetate (OAA), offering the potential for an excellent fermentation yield. The required carbon dioxide can be provided by using an alkali or alkali-earth carbonate solution for pH control or sparging carbon dioxide directly into the fermentor while controlling the pH with other bases such as NaOH, KOH and NH
4OH or alkali-earth hydroxides [
6]. Succinic acid is a more reduced molecule than its sugar fermentation feedstocks. This implies that in conversion to succinic acid, some of the sugar substrate must also be used for the production of reducing power. The theoretical yield of succinic acid from 6-carbon sugars such as glucose (C
6H
12O
6) and CO
2 via the reductive TCA route is 1.71 mole/mole sugar, or 1.12 g/g sugar, which is summarized in the following reaction [
7]:
Many independent and parallel efforts have been made to fermentatively produce succinic acid, with many choices for which organism was most suitable as a host. Regardless of which organism was chosen, each needed corrections that were often similar among the various organisms. The reduction or elimination of fermentation by-products other than succinic acid was the first step for improvement of most host organisms. For many of them the by-products were other organic acids such as lactic, acetic, or formic acid, and ethanol. The control and regulation of carbon flow surrounding pyruvate as the central node was also a common target for genetic manipulation.
A second challenge in the enhancement of succinic acid-producing hosts was the alleviation of an electron imbalance in the host organism. Succinic acid is a more reduced, or electron rich molecule, than its commonly used carbon feedstock, i.e., sugar, and many different approaches were taken to harness reducing equivalents or to restore co-factors. This is the underlying reason that it is not possible to produce two molecules of succinic from one C6-sugar molecule.
An important difference among the various microorganisms involves the optimum pH of the succinic acid fermentation. The accumulation of succinic acid in the fermentation broth inevitably leads to lowering of the pH in the fermentation medium. The pKa values for succinic acid, pKa1 = 4.61 and pKa2 = 5.61, lie below the optimal growth and production pH of many organisms. Maintaining the pH of the fermentation in a range suitable for the microorganism, and the choice of base to neutralize the acid, has a significant impact on the overall succinic acid production costs. Furthermore, the choice of base determines what succinate salt is produced, which can influence the target market for succinic acid. For example, the use of ammonium hydroxide as base, leading to the production of di-ammonium succinate, lends itself to post-fermentative production of pyrrolidinones. Several inventions surrounding succinic and other organic acids targeted the recovery and recycling of the neutralizing agent. The recognition that neutralization and base recycling contributed significantly to succinic acid production costs prompted the engineering of host organisms that can tolerate a lower pH and have reduced base requirements.
Natural succinic acid producers are organisms which produce succinic acid as the main fermentation product, and many of them are capable of fumarate respiration. All natural succinic acid-producing microorganisms were isolated from rumen fluids, an environment rich in microorganisms that produce volatile fatty acids (VFAs), which are directly and readily absorbed into the bloodstream of the ruminant host animal and used for energy production. These microorganisms are anaerobes, thrive at a near neutral pH, are capnophilic, and naturally use the reductive TCA cycle for succinic acid production. All natural succinic acid-producing organisms isolated from the rumen require pH control for a fast and efficient fermentation.
Anaerobiospirillum succiniciproducens (ANS) was one of the first bacteria isolated and shown to produce succinic acid [
3,
8]. Fermentations with this strict anaerobe reached titers of 50 g/L succinic acid. A classical selection for reduction of by-products, which included formic and acetic acid, was successful. However, the organism experienced instability and deterioration, was sensitive to high glucose concentrations and exposure to air, which made it unsuitable for industrial application.
The same isolation scheme used for ANS was also used to identify
Actinobacillus succinogenes, a more robust organism, and a facultative anaerobe [
9]. The pathway to succinic via the reductive TCA cycle in this gram-negative bacterium was biochemically characterized [
10], and the fermentation by-products, acetic and formic acid, were nearly eliminated using a classical selection method [
3]. Variants capable of reaching titers above 100 g/L succinate were reported. However, the organism did not thrive at pH values below pH 6.0, necessitating the neutralization of the produced acid, and the best performances were reported when using magnesium Mg
++ as a counterion. Consistent with its rumen origin, the organism was versatile in its use of carbon substrates. Most notable was its simultaneous utilization of 5- and 6-carbon sugars, as well as polyols such as sorbitol and glycerol. Fermentations using cellulosic biomass sugars, specifically corn stover hydrolysates, reached titers approaching 50 g/L in simultaneous saccharification fermentations (SSF) [
11]. In recent years, metabolic flux analyses and genome analysis confounded the understanding of the organism [
12] and metabolic engineering to achieve better electron balance enhanced the yield by channeling carbon through the pentose phosphate pathway [
13]. Fermentation performance reaching a titer of 109 g/L with productivity of 2.0 g/L.h and yield of 0.96 g succinic acid/g glucose was achieved in medium containing small amounts of complex nutrient sources, i.e., less than 0.5 g/L yeast extract or corn steep liquor (Guettler and Kleff, personal communications). Growth in defined minimal medium was possible but the low titer of 9 g/L did not meet industrial performance standards under these conditions [
14].
Genome analysis of Actinobacillus revealed many shared features with other natural succinic acid producers, especially its closest relative Mannheimia, and for these organisms phosphoenolpyruvate (PEP) is a key branch point. PEP carboxykinase, encoded by the pckA gene, is responsible for fixing CO2 to PEP and generating oxaloacetate, while harnessing the energy in form of GTP. A. succinogenes lacks a complete TCA cycle, missing key enzymes such as isocitrate dehydrogenase and α-ketoglutarate dehydrogenase, indicating a metabolism geared towards succinic acid production.
Mannheimia succiniciproducens was isolated from the rumen of a Korean cow, and initially referenced as a mixed acid producer. The organism uses the same reductive TCA production pathway to succinic acid as
A. succinogenes. By-products such as formate and acetate were eliminated through gene disruption of pyruvate formate lyase (
pflB), acetate kinase (
ackA), and phosphotransacetylase (
pta) [
15]. Additional paths to acetic acid are absent in the
M. succiniciproducens genome. The original
Mannheimia isolate also produced lactate, when grown in CO
2-poor conditions, which was precluded through disruption of the
ldhA gene, thereby generating a homosuccinate producing daughter strain, LPK7. A thorough genome analysis of
Mannheimia and comparison to engineered organisms elucidated predilection of
Mannheimia for succinic acid production, e.g., the reliance on PEP-ckA as a CO
2-fixing enzyme. This is possible in
Mannheimia, because glucose import is accomplished by a glucokinase activity that transfers phosphate from ATP to glucose [
16]. The organism lacks an NAD
+-dependent malic enzyme, pyruvate kinase, and pyruvate oxidase, thereby reducing the need for a large pyruvate pool.
Mannheimia has a complete TCA cycle, which may indicate that its metabolism may have different and more complex controls for channeling carbon towards succinic acid.
Mannheimia has been shown to demonstrate high rates of succinate production, although often in medium containing complex nitrogen sources such as yeast extract [
17]. A chemically defined medium has been developed [
18].
Isolation of
Basfia succiniciproducens followed the knowledge gained from the isolations of
Mannheimia and
Actinobacillus and was geared to isolate a strain belonging to the family of the
Pasteurellaceae. The genome of the original
Basfia isolate (DD1) was compared to the genome of
Mannheimia, and showed high genetic identity [
19], yet it was classified as a distinct organism. Similar to
Mannheimia,
Basfia also produces lactic acid as a major by-product under fermentative conditions. Lactic and formic acid production were eliminated through targeted deletions [
20].
Basfia can utilize a variety of carbon feedstocks, and continuous fermentations using crude glycerol from biodiesel production were successful [
21].
With regard to genetically engineered succinic acid producers, many research teams developed
Escherichia coli for succinic acid production, utilizing the genetic tools and knowledge available for extensive metabolic engineering approaches for this organism. The efforts at the Argonne National Laboratory (ANL) resulted in development of several promising
Escherichia coli strains for succinic acid production. Two of the most efficient strains were designated AFP111 and AFP184. Subsequently, a two-stage fermentation process was developed at the Oak Ridge National Laboratory (ORNL) for production of succinic acid using these
E. coli strains [
22,
23]. The developed technology was licensed to Applied Carbochemicals, Inc. (ACC), a start-up company in the United States. ACC became the first company that attempted to develop a commercially feasible fermentation process for the manufacture of succinic acid. Earlier efforts built on
E. coli’s mixed acid fermentation pathways to strengthen the reductive route to succinic acid, and built on the above mentioned starting strains AFP111 and AFP184 [
24]. The first improvement steps eliminated the production of lactic acid and formic acid, the latter through deletions in pyruvate formate lyase.
E. coli can use its PEP-carboxylase to enter the reductive TCA cycle, but the energy from PEP is lost as inorganic phosphate in the carboxylation step. This energy loss is undesirable under anaerobic, ATP limited conditions. Furthermore, the import of glucose through the phosphoenolpyruvate:carbohydrate phosphotransferase system (PTS) system, where PEP is the source of phosphate for the generation of glucose-6-phosphate, leads to a large pool of pyruvate, making it a key biological branch point in the organism, where multiple enzymatic activities compete for the pyruvate substrate [
25]. The control of the pyruvate node in
E. coli included changes in the glucose import mechanism, the introduction of heterologous genes with more favorable enzymatic characteristics, which either supported or replaced the corresponding endogenous genes, including the overexpression of malic enzyme [
26]. In contrast to natural anaerobic succinic producers,
E. coli was amenable to the separation of growth and succinic production, by combining fast aerobic growth, followed by a switch to anaerobic conditions for succinic production [
27], and the use of microaerobic conditions.
In an alternative approach,
E. coli was engineered for succinic production under fully aerobic conditions, thereby harnessing the higher production of cell-biomass, and faster carbon throughput to succinic production to favor a high productivity fermentation [
28]. However, this reduced the theoretical yield to one mole succinic per mole glucose. These innovative strain improvements channeled the carbon flow in
E. coli through either the glyoxylate shunt or a two-pronged route using the glyoxylate shunt and the oxidative TCA cycle [
29].
In contrast to many natural succinic producers
E. coli can grow to high cell densities in defined mineral media, while maintaining high succinic production capabilities [
30]. This broadened the technology deployment options, eliminated the potential need for costly complex nutrients and facilitated product recovery.
E. coli is capable of using a variety of sugars, and several engineered strains were designed to use sucrose or molasses as feedstock. However, the organism naturally shows a preference for glucose, which is consumed first or preferentially, when multiple sugars are present. Even among the 5-carbon sugars
E. coli demonstrates a preference for arabinose over xylose [
31]. The organism is also sensitive to high acetate concentrations, a commonly found inhibitor in cellulosic sugar streams.
E. coli may not be the best choice as host for the use of cellulosic mixed sugar streams, but this is a lesser concern currently, when other carbon sources are available for the bio-based production of succinic acid.
Corynebacterium glutamicum is an established industrial organism for the production of amino acids, and many tools have been developed for its genetic manipulation.
C. glutamicum can grow aerobically and anaerobically, and high succinic acid titers have been reached with an engineered strain of this bacterium under fed-batch conditions [
32]. Interestingly, the fed-batch process used glucose as carbon source and formic acid as a source of reducing equivalents to support succinic acid production under anaerobic conditions. The reported production yields consider only the succinic production phase, and do not account for the aerobic production of cell biomass. The small amounts of by-products seen in this two-phase system included α-ketoglutarate, pyruvate, and acetic acid. The process was run at a near neutral pH, and the succinic acid produced was neutralized with potassium hydroxide.
All bacterial succinic acid producers described above require the neutralization of the fermentation product. The final production process may include a base recycle, or recover the base-salt as a fermentation by-product. It is worth noting that the salt by-product in pH neutral succinic acid fermentations is generated at an equivalent scale to the amount of succinic acid produced. Such a high volume market is available for ammonium hydroxide neutralized fermentations, in which the diammonium succinate salt (DAS) is acidified with sulfuric acid to form free succinic acid and ammonium sulfate. The salt by-product, ammonium sulfate, can serve the fertilizer market.
The cost and effort associated with the use of a neutralizing base fostered the development of succinic acid production at a low pH by using a low-pH-tolerant host.
Saccharomyces cerevisiae, or other yeasts such as
Yarrowia, can thrive under slightly acidic conditions and methods for their metabolic engineering are well established [
33].
S. cerevisiae is an industrial organism capable of fermentative production and tolerance of high sugar concentrations. Similar to
E. coli, the organism has the potential to produce succinic acid both aerobically and anaerobically [
34]. However, fermentative conditions favor ethanol as well as glycerol production in yeast, and the elimination of these natural fermentation products was more complicated than in prokaryotic systems [
35]. This is due to the presence of multiple genes encoding alcohol dehydrogenases, and the deletion of genes leading to natural products had unfavorable side effects, such as low osmotolerance. Furthermore, the genes involved in anaerobic succinate production, using the reductive TCA cycle starting from oxaloacetate, underlie glucose repression, and the route is thermodynamically unfavorable.
Succinic acid production under aerobic conditions via the glyoxylate shunt is possible in yeast. As described for E. coli, this route has the benefit of faster growth, faster carbon metabolism, and better ATP balance, but also leads to a lower theoretical yield. Dual production pathways can be implemented and recent publications show improvements, but the performance of all three fermentation parameters, titer, productivity and yield, remain unclear. It is also worth noting that the fermentative production of succinic acid at a pH of 4.6, the pKa1 of succinic acid, will reduce the amount of base needed for neutralization, but will not eliminate the need for base entirely. At this pH 25% of the acid produced will remain in the mono-salt form, which needs to be considered in the recovery. Nevertheless, low pH succinic production enjoys the benefits of reduced base and reduced salt formation.
Many succinic acid-producing microorganisms have been developed and much effort was put into characterizing and improving the organisms. However, a direct performance comparison between the strains is difficult because the culture conditions were different and in many cases were insufficiently described. In addition, the fermentation process used for several organisms included a growth phase, during which there was none or negligible succinic acid synthesis, followed by a production phase, whereas the growth phase was not separated from the production phase for others. Calculations of succinic acid yield and productivity, therefore, could not be performed on the same basis. Despite these problems, one publication has provided a thorough summary of many succinic acid-producing bacterial strains [
36].
3. Succinic Acid Recovery
The recovery and purification of succinic acid from the fermentation broth can be a complex, multi-step, and expensive process. Knowledge of purification procedures, and the required purity of the final product, also guided the selection of inputs for the production of succinic acid. Due to the expense that the recovery and purification process can impart on the final product and its uses, each of the current succinic acid producers developed and patented their own recovery and purification processes. These processes vary depending on the organism used for production, feedstock used, nutrients supplied, acids and bases used to maintain pH, solubility of the intermediates, final product titer achievable, and intended applications of the recovered succinic acid products. The first step, which is common in all the described and patented processes, is removal of cells and insoluble solids through the use of standard equipment such as filtration or centrifugation. Subsequent recovery and purification steps described in the recent patents and patent applications are highlighted.
BASF describes a recovery in patents and publications surrounding claims for its proprietary microorganism. The first step describes the concentration of clarified broth through multistage evaporation to reduce processing volumes, followed by a cation exchange chromatography, in which the succinic acid salt is reacted with a strong acid cation exchange resin at a temperature of 46 to 60 °C. This achieves the conversion of the succinate salt to succinic acid, which can be crystallized to form the final product. Alternatively, if the fermentation is neutralized by a calcium base, the calcium succinate product has low solubility and can be separated from the broth by filtration. The precipitate is treated with sulfuric acid to form soluble succinic acid and calcium sulfate (gypsum). The latter has very low solubility and can be separated by filtration. The succinic acid solution may be further purified by the same cation exchange chromatography as described previously [
37,
38].
For BioAmber, the starting material for the recovery is diammonium succinate (DAS), which is produced by neutralizing the succinic acid in the fermentation broth with NH
3. The filtered broth undergoes a reactive evaporation, in which it is heated to 135 °C and 50 psig, converting the DAS to monoammonium succinate (MAS) and resulting in roughly a two-fold concentration. The MAS solution is sent to a three stage evaporative crystallization system, which cools the solution to the point when MAS is no longer soluble enabling up to 95 wt % recovery. The crystalline MAS can be dissolved in reverse osmosis water, and the solution may be converted to succinic acid by either bipolar membrane electrodialysis, or through the use of ion exchange chromatography. The succinic acid solution undergoes another evaporative crystallization step in which 95 wt % is recovered as a solid, with the remaining succinic acid in the mother liquor recycled back to the reactive evaporation step [
39,
40].
The DSM/Roquette process begins with an evaporation phase carried out between 65 and 80 °C followed by a crystallization phase in which the concentrated solution is cooled to between 1 and 25 °C to produce intermediate crystals and a mother liquor. The intermediate crystals are separated from the mother liquor by centrifugation and the mother liquor is subjected to microfiltration and nanofiltration. The small pore (100–300 Da) membrane allows succinic acid to pass through, while higher molecular weight soluble materials are retained. The filtered mother liquor is recycled to the evaporation step in the beginning of the process to produce additional intermediate crystals. The intermediate crystals are dissolved into a minimum volume of 40–90 °C water. The solution is passed through a series of purification steps including activated carbon column, cation exchange, and anion exchange. The purified solution undergoes a final crystallization to produce a succinic acid product of high purity [
41].
Mitsubishi’s recovery method employs crystallization, which is applicable to solutions of succinic acid of either petrochemical or biological origin. The solution containing succinic acid at near saturated concentrations is fed to a crystallization tank where the overhead pressure is reduced to below atmospheric to cause a drop in temperature, which subsequently causes precipitation of succinic acid. Stirring is carried out at pre-determined rates to ensure uniform size of the crystals. The crystals are removed from the tank and pulverized to form a product having particle size between 100 and 300 mm [
42].
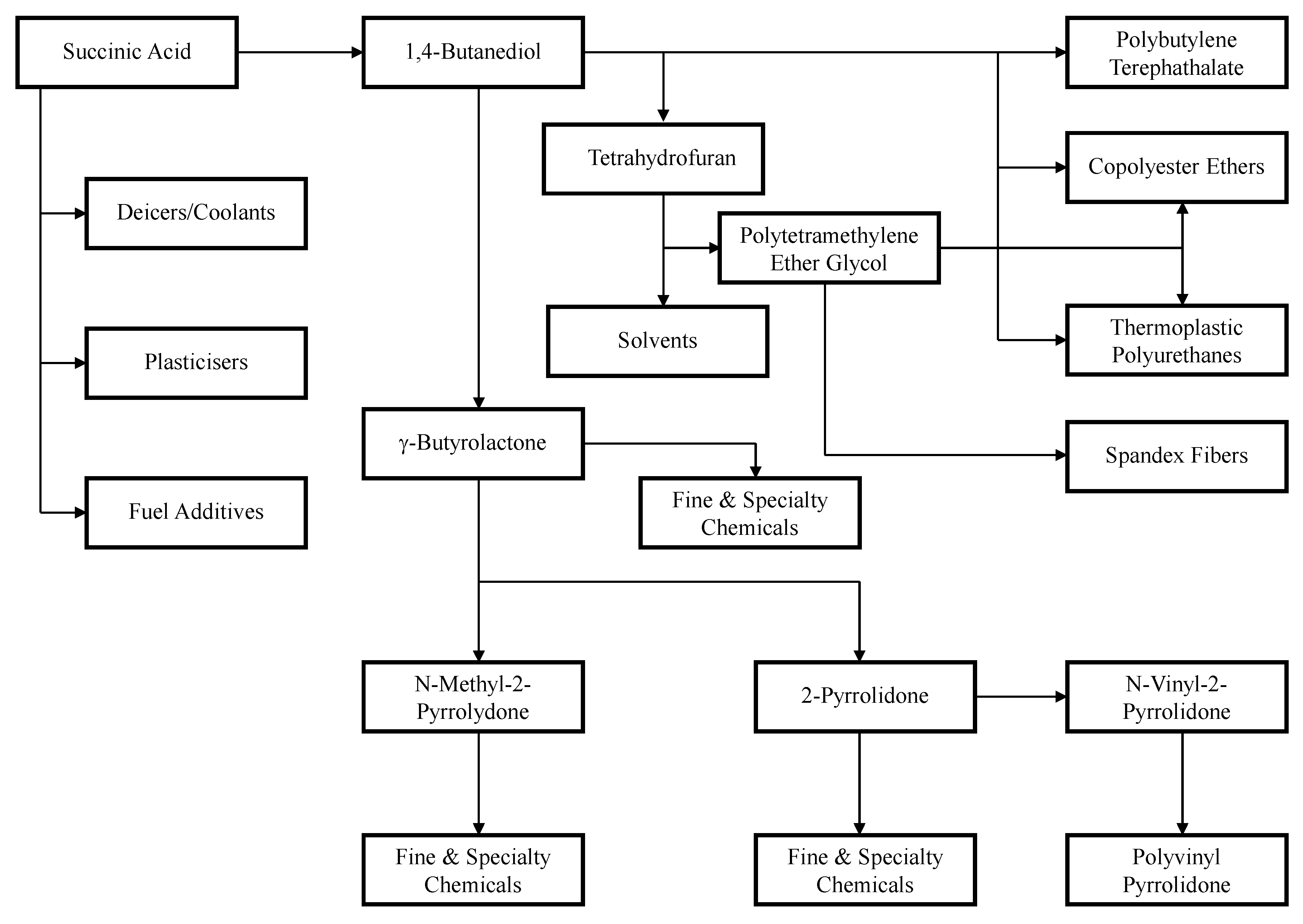
For Myriant, the purification process begins with a fermentation broth that has been neutralized with NaOH to produce diammonium succinate. The clarified broth is concentrated by evaporation under vacuum and subsequently acidified with H
2SO
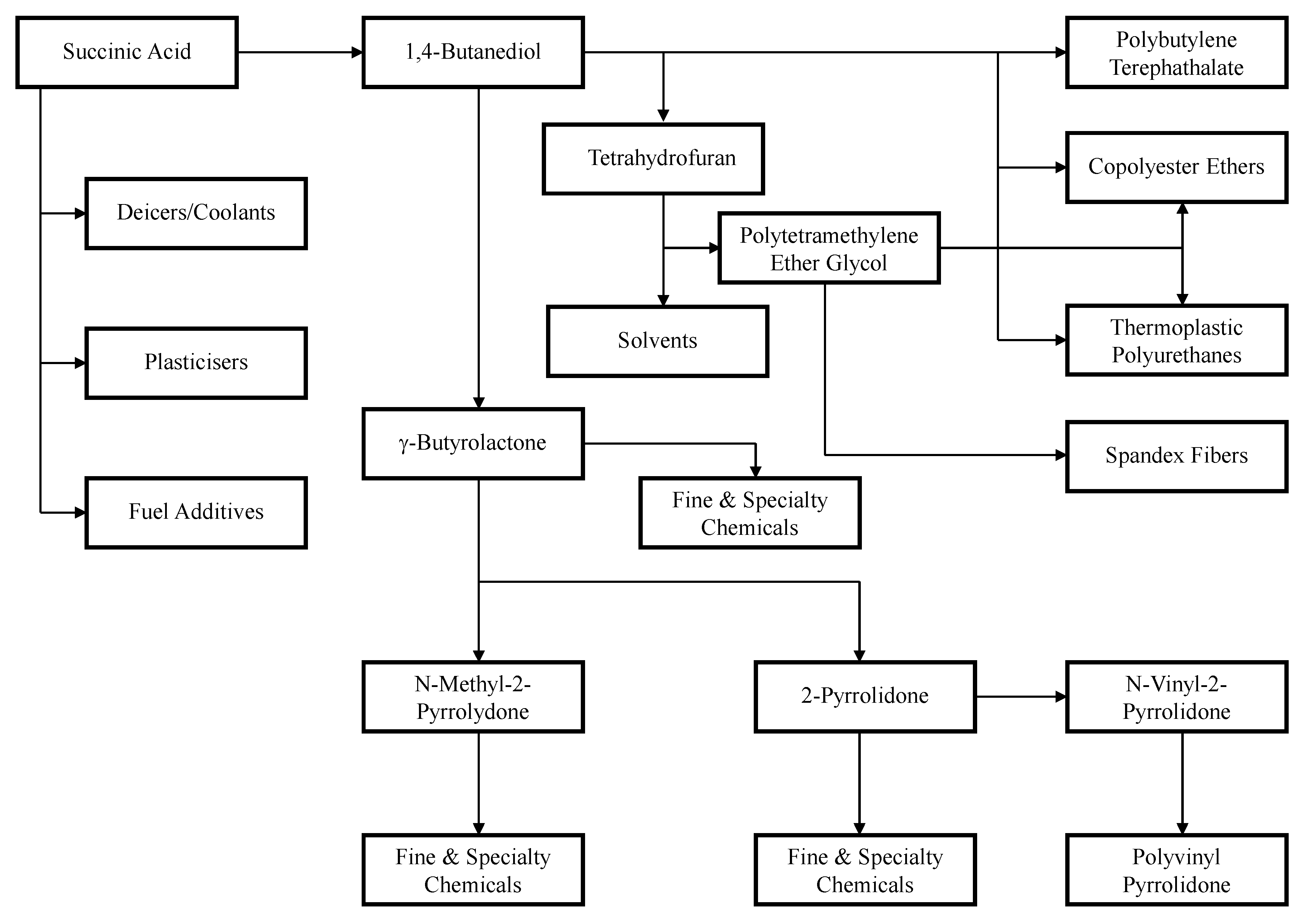
4 to produce succinic acid and ammonium sulfate. The temperature of the acidified broth is further lowered to cause the succinic acid to crystallize and precipitate, while the ammonium sulfate remains soluble. The succinic acid crystals are harvested by centrifugation. The crystals stream may either be dissolved again and recrystallized to improve purity, or be sent to an esterification process to produce a stream that is suitable for the further production of 1,4-butanediol, γ-butyrolactone, or tetrahydrofuran. The mother liquor, which contains primarily ammonium sulfate and the remaining succinic acid, is separated into two streams by simulated moving bed chromatography. The ammonium sulfate stream can be sold as fertilizer in liquid or crystallized solid form, and the succinic acid stream can be polished by nanofiltration before being crystallized and dried to form the final product [
43].
The process patented by Purac focuses on recovery of magnesium or calcium succinate. Following broth clarification, a monovalent base is added to convert the divalent succinate to the monovalent succinate salt. The best conversion is obtained using sodium hydroxide in the case of magnesium succinate, and sodium carbonate for calcium succinate, which yielded a 99.8% and 99.3% conversion, respectively. While these are the preferred monovalent bases, other monovalent bases may be used for economic reasons or to produce a different end product. The monovalent succinate salt and the magnesium or calcium hydroxide/carbonate can be separated by filtration. The divalent base may be washed, to reduce product losses, before being recycled back into the fermentation process. The succinate salt is further purified via ion exchange to reduce the calcium/magnesium ion content to levels that would allow the use of bipolar electrodialysis. This would yield a very high-purity succinic acid stream ready for crystallization, or for conversion to succinate esters and alternate product synthesis [
44].
5. Commercialization of Bio-Based Succinic Acid
Bio-based succinic acid currently is produced commercially by four companies, which include BioAmber, Myriant, Reverdia and Succinity. Each of these manufacturers of bio-based succinic acid has collaborations with a host of other companies on development of new technologies for production of succinic acid and its derivatives.
As discussed previously, ACC was the first company that was formed to commercialize bio-based succinic acid using the
E. coli strains and fermentation process developed by the US DOE. ACC became Diversified Natural Products (DNP) Green Technology, which subsequently formed a collaboration with the France-based Agro-Industrie Recherches et Développements (ARD) to develop and commercialize bio-based succinic acid. In 2010, DNP Green Technology acquired 100 percent of the joint venture from ARD and changed its corporate name to BioAmber, Inc. (Plymouth, MN, USA). BioAmber (and its predecessors) developed succinic acid technology based on the US DOE’s
E. coli strains. The company recently collaborated with Cargill to develop yeast strains for succinic acid production [
54]. BioAmber also has collaborated with Mitsui to build a 30,000 MT/year succinic acid production plant in Sarnia, ON, Canada, which is now in operation. In addition to the Sarnia plant, BioAmber is planning to build a second plant in North America that will produce 1,4-butanediol (BDO), tetrahydrofuran (THF) and succinic acid. The nameplate capacity is expected to be 91,000 MT of BDO/THF and 63,500 MT of succinic acid per year. A third succinic acid plant is being planned for in Thailand in a partnership with PTT-MCC Biochem, which is a joint venture between PTT PLC and Mitsubishi Chemical. The succinic acid produced at this plant is intended for use exclusively for production of PBS by PTT-MCC Biochem [
55].
Myriant is another bio-based succinic acid producer in the United States. The company initially licensed the
E. coli strains developed at the University of Florida [
56,
57] then continued their development to obtain
E. coli strains capable of utilizing sugars derived from lignocellulosic feedstocks for succinic acid production [
58]. Myriant currently is operating a 13.6 MT succinic acid production plant in Lake Providence, Louisiana. The intended feedstocks for use in this plant include glucose derived from grain sorghum and sugars derived from lignocellulosic biomass [
59]. A second succinic acid production plant with initial annual output of 500 MT and plan for expansion to 5000 MT is located in Leuna, Germany and is operated by Myriant’s partner ThyssenKrupp Uhde [
46,
59]. There have been reports on potential partnership with China National BlueStar to build a third succinic acid production plant in Nanjing, China with annual output of 100,000 MT. The succinic acid produced is intended for use as feedstock for BDO production [
46,
60]. In addition to succinic acid, Myriant also attempts to develop technologies for production of other organic acids, which include lactic, acrylic, muconic and fumaric acid [
59].
Reverdia is a joint venture between Dutch chemical company DSM and French starch derivatives producer Roquette. Reverdia successfully operated a demonstration plant with annual capacity of 500 MT in Lestrem, France from 2010 to 2012. In December 2012, commercial succinic acid production began at the 10,000 MT/year plant in Cassano Spinola, AL, Italy [
61]. The fermentation process is based on a recombinant
S. cerevisiae strain developed by DSM [
62]. DSM also developed a recombinant
S. cerevisiae strain for co-production of ethanol and succinic acid. In this strain, the energy generated by ethanol production in the form of ATP is sufficient to support succinic acid synthesis [
63]. There has been no indication that this strain would be used in a commercial process.
Succinity is a joint venture between Germany-based BASF and Dutch company Corbion Purac, which was formed to produce succinic acid using a proprietary strain of
Basfia succiniciproducens. Succinity successfully operated a 500 MT/year demonstration plant in Barcelona, Spain [
64]. Commercial production of succinic acid started in 2014 at a 10,000 MT/year plant in Montmeló, Spain. In addition to this plant, planning for a second production plant was reported [
46,
65].
There have been only one LCA study and one sustainability study performed on the commercial processes for succinic acid production. In the LCA study of the Myriant’s process, real data from the plant in Louisiana were used [
66]. In the base case in this study, sorghum grains were used as feedstock. Comparing to the base case, the use of glucose as feedstock would increase the global warming potentials (GWP) and the non-renewable fossil cumulative energy demand (non-ren CED) by 1.72 times and 1.86 times, respectively. The GWP and non-ren CED in the manufacture of succinic acid by the petrochemical route using maleic anhydride as feedstock were found to be 3.85 and 10.44 times higher than in the base case. A sustainability study was performed on the Myriant’s process using sorghum grains and sugar beet as feedstocks and the Reverdia’s process for co-production of ethanol and succinic acid [
67]. The material efficiency ratios for bio-based succinic acid produced from sorghum and sugar beet were calculated to be 13% in both cases, compared to 76% calculated for petrochemical-based succinic acid. The corresponding energy efficiency ratios were calculated to be 30%, 31%, and 23%, respectively. The calculated energy efficiency ratios for the Reverdia’s process using sorghum grains and sugar beet were 51% and 54%, respectively. The calculated production cost in all cases of succinic acid manufactured by either bio-based process was significantly lower than the calculated cost for petrochemical-based succinic acid. Even in the worst-case scenario, the production cost of bio-based succinic acid was only 41% of the production cost of petrochemical-based succinic acid (
$1.17/kg vs.
$2.86/kg). The results of the two aforementioned studies clearly indicated the advantages of bio-based succinic acid.




{kind=link}
{kind=link}
{kind=link}