Molecular Typing of Campylobacter jejuni and Campylobacter coli Isolated from Various Retail Meats by MLST and PFGE


Abstract
:1. Introduction
2. Experimental Section
2.1. Bacterial Isolates

{kind=link}
{kind=link}
| Source | C. jejuni | C. coli |
|---|---|---|
| Chicken | 7 | 3 |
| Chicken Livers | 4 | 5 |
| Chicken Gizzards | 7 | 2 |
| Turkey | 5 | 2 |
| Beef Livers | 5 | 5 |
| Pork | 0 | 2 |
| Total | 28 | 19 |
2.2. Pulsed-Field Gel Electrophoresis Typing
2.3. Multilocus Sequence Typing
| Genes | Primer Sequences | Use | References |
|---|---|---|---|
| aspA | 5′-AGTACTAATGATGCTTATCC-3′ 5′-ATTTCATCAATTTGTTCTTTGC-3′ | C. jejuni PCR | [14] |
| glnA | 5′-TAGGAACTTGGCATCATATTACC-3′ 5′-TTGGACGAGCTTCTACTGGC-3′ | C. jejuni PCR | [14] |
| gltA | 5′-GGGCTTGACTTCTACAGCTACTTG-3′ 5′-CCAAATAAAGTTGTCTTGGACGG-3′ | C. jejuni PCR | [14] |
| glyA | 5′-GAGTTAGAGCGTCAATGTGAAGG-3′ 5′-AAACCTCTGGCAGTAAGGGC-3′ | C. jejuni PCR | [14] |
| pgm | 5′-TACTAATAATATCTTAGTAGG-3′ 5′-CACAACATTTTTCATTTCTTTTTC-3′ | C. jejuni PCR | [14] |
| tkt | 5′-GCAAACTCAGGACACCCAGG-3′ 5′-AAAGCATTGTTAATGGCTGC-3′ | C. jejuni PCR | [14] |
| uncA | 5′-ATGGACTTAAGAATATTATGGC-3′ 5′-GCTAAGCGGAGAATAAGGTGG-3′ | C. jejuni PCR | [14] |
| aspA | 5′-AGTACTAATGATGCTTATCC-3′ 5′-ATTTCATCAATTTGTTCTTTGC-3′ | C. jejuni sequencing | [14] |
| glnA | 5′-TAGGAACTTGGCATCATATTACC-3′ 5′-TTGGACGAGCTTCTACTGGC-3′ | C. jejuni sequencing | [14] |
| gltA | 5′-GGGCTTGACTTCTACAGCTACTTG-3′ 5′-CCAAATAAAGTTGTCTTGGACGG-3′ | C. jejuni sequencing | [14] |
| glyA | 5′-GAGTTAGAGCGTCAATGTGAAGG-3′ 5′-AAACCTCTGGCAGTAAGGGC-3′ | C. jejuni sequencing | [14] |
| pgm | 5′-TACTAATAATATCTTAGTAGG-3′ 5′-CACAACATTTTTCATTTCTTTTTC-3′ | C. jejuni sequencing | [14] |
| tkt | 5′-GCAAACTCAGGACACCCAGG-3′ 5′-AAAGCATTGTTAATGGCTGC-3′ | C. jejuni sequencing | [14] |
| uncA | 5′-ATGGACTTAAGAATATTATGGC-3′ 5′-GCTAAGCGGAGAATAAGGTGG-3′ | C. jejuni sequencing | [14] |
| aspA | 5′-CCAACTGCAAGATGCTGTACC-3′ 5′-TTCATTTGCGGTAATACCATC-3′ | C. coli PCR and sequencing | [27] |
| glnA | 5′-CATGCAATCAATGAAGAAAC-3′ 5′-TTCCATAAGCTCATATGAAC-3′ | C. coli PCR and sequencing | [27] |
| gltA | 5′-CTTATATTGATGGAGAAAATGG-3′ 5′-CCAAAGCGCACCAATACCTG-3′ | C. coli PCR and sequencing | [27] |
| glyA | 5′-AGCTAATCAAGGTGTTTATGCGG-3′ 5′-AGGTGATTATCCGTTCCATCGC-3′ | C. coli PCR and sequencing | [27] |
| pgm | 5′-GGTTTTAGATGTGGCTCATG-3′ 5′-TCCAGAATAGCGAAATAAGG-3′ | C. coli PCR and sequencing | [27] |
| tkt | 5′-GCTTAGCAGATATTTTAAGTG-3′ 5′-AAGCCTGCTTGTTCTTTGGC-3′ | C. coli PCR and sequencing | [27] |
| uncA | 5′-AAAGTACAGTGGCACAAGTGG-3′ 5′-TGCCTCATCTAAATCACTAGC-3′ | C. coli PCR and sequencing | [27] |
3. Results and Discussion
3.1. Pulsed-Field Gel Electrophoresis

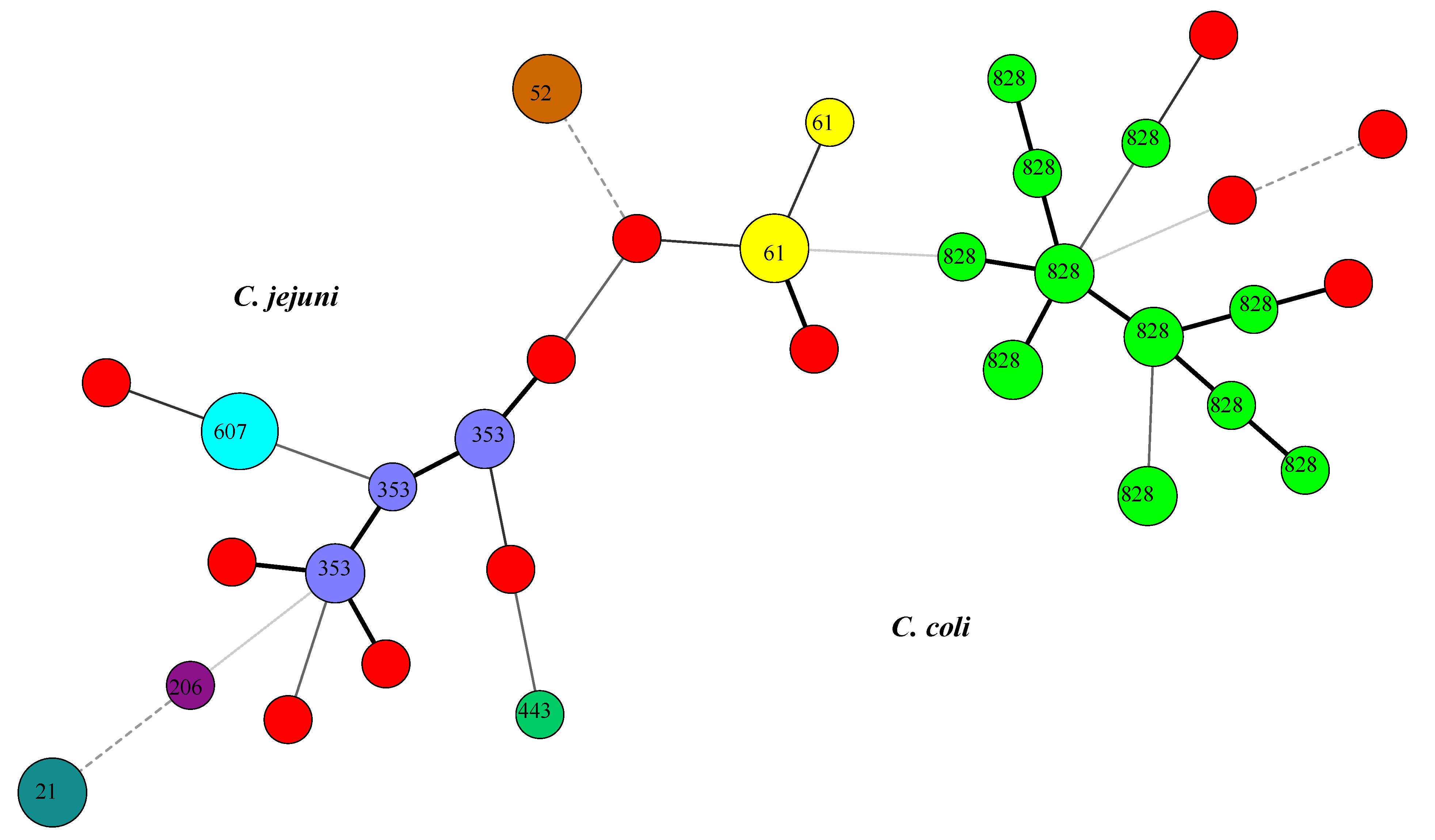
3.2. Multilocus Sequence Typing
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Centers for Disease Control (CDC). CDC Estimates of Foodborne Illness in the United States. 2011. Available online: http://www.cdc.gov/foodborneburden/2011-foodborne-estimates.html (accessed on 16 May 2013). [Google Scholar]
- Debruyne, L.; Gevers, D.; Vandamme, P. Taxonomy of the Family Campylobacteraceae. In Campylobacter, 3rd ed.; Nachamkin, I., Szymanski, C.M., Blaser, M.J., Eds.; American Society for Microbiology: Washington, DC, USA, 2008; pp. 3–26. [Google Scholar]
- Lastovica, A.J.; Allos, B.M. Clinical Significance of Campylobacter and Related Species Other than Campylobacter jejuni and Campylobacter coli. In Campylobacter, 3rd ed.; Szymanski, C.M., Blaser, M.J., Eds.; American Society for Microbiology: Washington, DC, USA, 2008; pp. 123–149. [Google Scholar]
- Zhao, S.; Young, S.R.; Tong, E.; Abbott, J.W.; Womack, N.; Friedman, S.L.; McDermott, P.F. Antimicrobial resistance of Campylobacter isolates from retail meat in the United States: 2002–2007. Appl. Environ. Microbiol. 2010, 76, 7949–7956. [Google Scholar] [CrossRef]
- Behringer, M.; Miller, W.G.; Oyarzabal, O.A. Typing of Campylobacter jejuni and Campylobacter coli isolated from live broilers and retail broiler meat by flaA-RFLP, MLST, PFGE and REP-PCR. J. Microbiol. Methods 2011, 84, 194–201. [Google Scholar] [CrossRef]
- Lukinmaa, S.; Nakari, U.M.; Eklund, M.; Siitonen, A. Application of molecular genetic methods in diagnostics and epidemiology of food-borne bacterial pathogens. APMIS 2004, 112, 908–929. [Google Scholar] [CrossRef]
- Van Belkum, A.; Tassios, P.T.; Dijkshoorn, L.; Haeggman, S.; Cookson, B.; Fry, N.K.; Fussing, V.; Green, J.; Feil, E.; Gerner-Smidt, P.; et al. Guidelines for the validation and application of typing methods for use in bacterial epidemiology. Clin. Microbiol. Infect. 2007, 13, 1–46. [Google Scholar]
- Maslow, J.N.; Slutsky, A.M.; Arbeit, R.D. The Application of Pulsed-Field Gel Electrophoresis to Molecular Epidemiology. In Diagnostic Molecular Microbiology; Persing, D.H., Tenover, F.C., Smith, T.F., White, T.J., Eds.; ASM Press: Washington, DC, USA, 1993; pp. 563–572. [Google Scholar]
- Olive, D.M.; Bean, P. Principles and applications of methods for DNA-based typing of microbial organisms. J. Clin. Microbiol. 1999, 37, 1661–1669. [Google Scholar]
- Wassenaar, T.M.; Newell, D.G. Genotyping of Campylobacter spp. Appl. Environ. Microbiol. 2000, 66, 1–9. [Google Scholar] [CrossRef]
- Thakur, S.; White, D.G.; McDermott, P.F.; Zhao, S.; Kroft, B.; Gebreyes, W.; Abbott, J.; Cullen, P.; English, L.; Carter, P.; et al. Genotyping of Campylobacter coli isolated from humans and retail meats using multilocus sequence typing and pulsed-field gel electrophoresis. J. Appl. Microbiol. 2009, 106, 1722–1733. [Google Scholar] [CrossRef]
- On, S.L.; Nielsen, E.M.; Engberg, J.; Madsen, M. Validity of SmaI-defined genotypes of Campylobacter jejuni examined by SalI, KpnI, and BamHI polymorphisms: Evidence of identical clones infecting humans, poultry, and cattle. Epidemiol. Infect. 1998, 120, 231–237. [Google Scholar] [CrossRef]
- Oyarzabal, O.A.; Backert, S.; Williams, L.L.; Lastovica, A.J.; Miller, R.S.; Pierce, S.J.; Vieira, S.L.; Rebollo-Carrato, F. Molecular typing of Campylobacter jejuni strains isolated from commercial broilers in Puerto Rico. J. Appl. Microbiol. 2008, 105, 800–812. [Google Scholar] [CrossRef]
- Dingle, K.E.; Colles, F.M.; Wareing, D.R.A.; Ure, R.; Fox, A.J.; Bolton, F.J.; Bootsma, R.J.L.; Willems, R.; Urwin, R.; Maiden, M.C.J. Multilocus sequence typing system for Campylobacter jejuni. J. Clin. Microbiol. 2001, 39, 14–23. [Google Scholar] [CrossRef]
- Dingle, K.E.; Colles, F.M.; Ure, R.; Wagenaar, J.A.; Duim, B.; Bolton, F.J.; Fox, A.J.; Wareing, D.R.; Maiden, M.C. Molecular characterization of Campylobacter jejuni clones: A basis for epidemiologic investigation. Emerg. Infect. Dis. 2002, 8, 949–955. [Google Scholar]
- Clark, C.G.; Taboada, E.; Grant, C.C.R.; Blakeston, C.; Pollari, F.; Marshall, B.; Rahn, K.; Mackinnon, J.; Daignault, D.; Pillai, D.; et al. Comparison of molecular typing methods useful for detecting clusters of Campylobacter jejuni and C. coli isolates through routine surveillance. J. Clin. Microbiol. 2012, 50, 798–809. [Google Scholar] [CrossRef]
- Colles, F.M.; Maiden, M.C.J. Campylobacter sequence typing databases: Applications and future prospects. Microbiology 2012, 158, 2695–2709. [Google Scholar] [CrossRef]
- Kotetishvili, M.; Stine, O.C.; Chen, Y.; Kreger, A.; Sulakvelidze, A.; Sozhamannan, S.; Morris, J.G., Jr. Multilocus sequence typing has better discriminatory ability for typing Vibrio cholerae than does pulsed-field gel electrophoresis and provides a measure of phylogenetic relatedness. J. Clin. Microbiol. 2003, 41, 2191–2196. [Google Scholar] [CrossRef]
- Alcaine, S.D.; Soyer, Y.; Warnick, L.D.; Su, W.L.; Sukhnanand, S.; Richards, J.; Fortes, E.D.; McDonough, P.; Root, T.P.; Dumas, N.B.; et al. Multilocus sequence typing supports the hypothesis that cow- and human-associated Salmonella isolates represent distinct and overlapping populations. Appl. Environ. Microbiol. 2006, 72, 7575–7585. [Google Scholar] [CrossRef]
- Miller, W.G.; On, S.L.; Wang, G.; Fontanoz, S.; Lastovica, A.J.; Mandrell, R.E. Extended multilocus sequence typing system for Campylobacter coli, C. lari, C. upsaliensis, and C. helveticus. J. Clin. Microbiol. 2005, 43, 2315–2329. [Google Scholar]
- Campylobacter MLST Home Page. Available online: www.pubmlst.org/campylobacter/ (accessed on 15 March 2012).
- Manning, G.; Dowson, C.G.; Bagnall, M.C.; Ahmed, I.H.; West, M.; Newell, D.G. Multilocus sequence typing for comparison of veterinary and human isolates of Campylobacter jejuni. Appl. Environ. Microbiol. 2003, 69, 6370–6379. [Google Scholar] [CrossRef]
- Suerbaum, S.; Lohrengel, M.; Sonnevend, A.; Ruberg, F.; Kist, M. Allelic diversity and recombination in Campylobacter jejuni. J. Bacteriol. 2001, 183, 2553–2559. [Google Scholar] [CrossRef]
- Noormohamed, A.; Fakhr, M.K. Incidence and antimicrobial resistance profiling of Campylobacter in retail chicken livers and gizzards. Foodborne Pathog. Dis. 2012, 9, 617–624. [Google Scholar] [CrossRef]
- Noormohamed, A.; Fakhr, M.K. A higher prevalence rate of Campylobacter in retail beef livers compared to other beef and pork meat cuts. Int. J. Environ. Res. Public Health 2013, 10, 2058–2068. [Google Scholar] [CrossRef]
- Centers for Disease Control. Standard Operating Procedure for PulseNet PFGE of Campylobacter jejuni. 2011. Available online: http://www.cdc.gov/pulsenet/PDF/campylobacter-pfge-protocol-508c.pdf (accessed on 15 August 2011). [Google Scholar]
- Dingle, K.E.; Colles, F.M.; Falush, D.; Maiden, M.C. Sequence typing and comparison of population biology of Campylobacter coli and Campylobacter jejuni. J. Clin. Microbiol. 2005, 43, 340–347. [Google Scholar] [CrossRef]
- Marmur, J. A procedure for the isolation of deoxyribonucleic acid from microorganisms. J. Mol. Biol. 1961, 3, 208–218. [Google Scholar] [CrossRef]
- Jolley, K.A.; Maiden, M.C. BIGSdb: Scalable analysis of bacterial genome variation at the population level. BMC Bioinformatics 2010, 11, 595. [Google Scholar] [CrossRef]
- Primers Used for MLST of Campylobacter. Available online: pubmlst.org/campylobacter/info/primers.shtml (accessed on 15 March 2012).
- Miller, W.G.; Englen, M.D.; Kathariou, S.; Wesley, I.V.; Wang, G.; Pittenger-Alley, L.; Siletz, R.M.; Muraoka, W.; Fedorka-Cray, P.J.; Mandrell, R.E. Identification of host-associated alleles by multilocus sequence typing of Campylobacter coli strains from food animals. Microbiology 2006, 152, 245–255. [Google Scholar] [CrossRef]
- Thakur, S.; Morrow, W.E.; Funk, J.A.; Bahnson, P.B.; Gebreyes, W.A. Molecular epidemiologic investigation of Campylobacter coli in swine production systems, using multilocus sequence typing. Appl. Environ. Microbiol. 2006, 72, 5666–5669. [Google Scholar] [CrossRef]
- Abley, M.J.; Wittum, T.E.; Funk, J.A.; Gebreyes, W.A. Antimicrobial susceptibility, pulsed-field gel electrophoresis, and multi-locus sequence typing of Campylobacter coli in swine before, during, and after the slaughter process. Foodborne Pathog. Dis. 2012, 9, 506–512. [Google Scholar] [CrossRef]
- Carrillo, C.D.; Kruczkiewicz, P.; Mutschall, S.; Tudor, A.; Clark, C.; Taboada, E.N. A framework for assessing the concordance of molecular typing methods and the true strain phylogeny of Campylobacter jejuni and C. coli using draft genome sequence data. Front. Cell. Infect. Microbiol. 2012, 2, 57. [Google Scholar]
- Magnússon, S.H.; Guðmundsdóttir, S.; Reynisson, E.; Rúnarsson, A.R.; Harðardóttir, H.; Gunnarson, E.; Georgsson, F.; Reiersen, J.; Marteinsson, V.T. Comparison of Campylobacter jejuni isolates from human, food, veterinary and environmental sources in Iceland using PFGE, MLST and fla-SVR sequencing. J. Appl. Microbiol. 2011, 111, 971–981. [Google Scholar] [CrossRef]
- Griekspoor, P.; Engvall, E.O.; Olsen, B.; Waldenstrom, J. Multilocus sequence typing of Campylobacter jejuni from broilers. Vet. Microbiol. 2010, 140, 180–185. [Google Scholar] [CrossRef]
- Cornelius, A.J.; Gilpin, B.; Carter, P.; Nicol, C.; On, S.L. Comparison of PCR binary typing (P-BIT), a new approach to epidemiological subtyping of Campylobacter jejuni, with serotyping, pulsed-field gel electrophoresis, and multilocus sequence typing methods. Appl. Environ. Microbiol. 2010, 76, 1533–1544. [Google Scholar] [CrossRef]
- Nielsen, L.N.; Sheppard, S.K.; McCarthy, N.D.; Maiden, M.C.; Ingmer, H.; Krogfelt, K.A. MLST clustering of Campylobacter jejuni isolates from patients with gastroenteritis, reactive arthritis and Guillain-Barré syndrome. J. Appl. Microbiol. 2010, 108, 591–599. [Google Scholar] [CrossRef]
- Colles, F.M.; Jones, K.; Harding, R.M.; Maiden, M.C. Genetic diversity of Campylobacter jejuni isolates from farm animals and the farm environment. Appl. Environ. Microbiol. 2003, 69, 7409–7413. [Google Scholar] [CrossRef]
- Duim, B.; Wassenaar, T.M.; Rigter, A.; Wagenaar, J. High-resolution genotyping of Campylobacter strains isolated from poultry and humans with amplified fragment length polymorphism fingerprinting. Appl. Environ. Microbiol. 1999, 65, 2369–2375. [Google Scholar]
- Strachan, N.J.C.; MacRae, M.; Thomson, A.; Rotariu, O.; Ogden, I.D.; Forbes, K.J. Source attribution, prevalence and enumeration of Campylobacter spp. from retail liver. Int. J. Food Microbiol. 2012, 153, 234–236. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Noormohamed, A.; Fakhr, M.K. Molecular Typing of Campylobacter jejuni and Campylobacter coli Isolated from Various Retail Meats by MLST and PFGE. Foods 2014, 3, 82-93. https://doi.org/10.3390/foods3010082
Noormohamed A, Fakhr MK. Molecular Typing of Campylobacter jejuni and Campylobacter coli Isolated from Various Retail Meats by MLST and PFGE. Foods. 2014; 3(1):82-93. https://doi.org/10.3390/foods3010082
Chicago/Turabian StyleNoormohamed, Aneesa, and Mohamed K. Fakhr. 2014. "Molecular Typing of Campylobacter jejuni and Campylobacter coli Isolated from Various Retail Meats by MLST and PFGE" Foods 3, no. 1: 82-93. https://doi.org/10.3390/foods3010082
APA StyleNoormohamed, A., & Fakhr, M. K. (2014). Molecular Typing of Campylobacter jejuni and Campylobacter coli Isolated from Various Retail Meats by MLST and PFGE. Foods, 3(1), 82-93. https://doi.org/10.3390/foods3010082