
Potential Vegetation and Carbon Redistribution in Northern North America from Climate Change

 , ,
, ,
Abstract
:1. Introduction
2. Methods
2.1. Model
2.2. Climate Data
2.3. Remote Sensing Data
3. Results
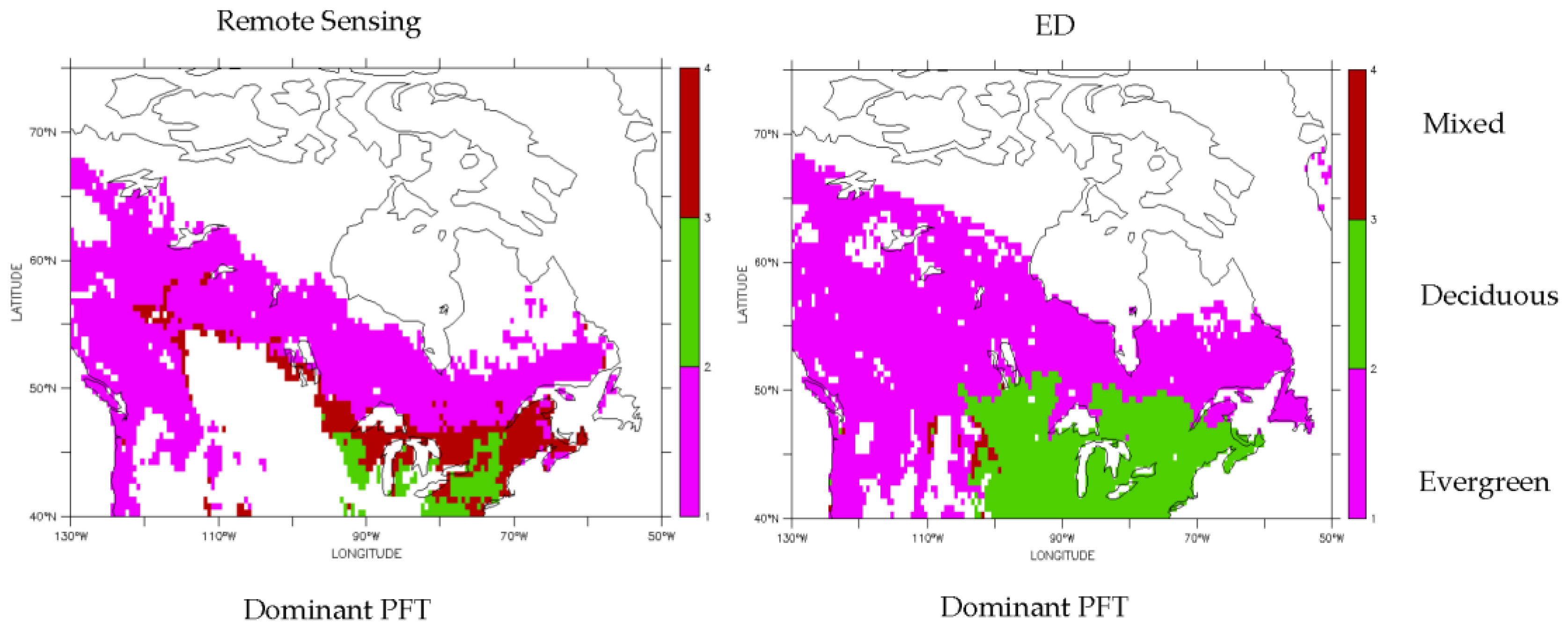
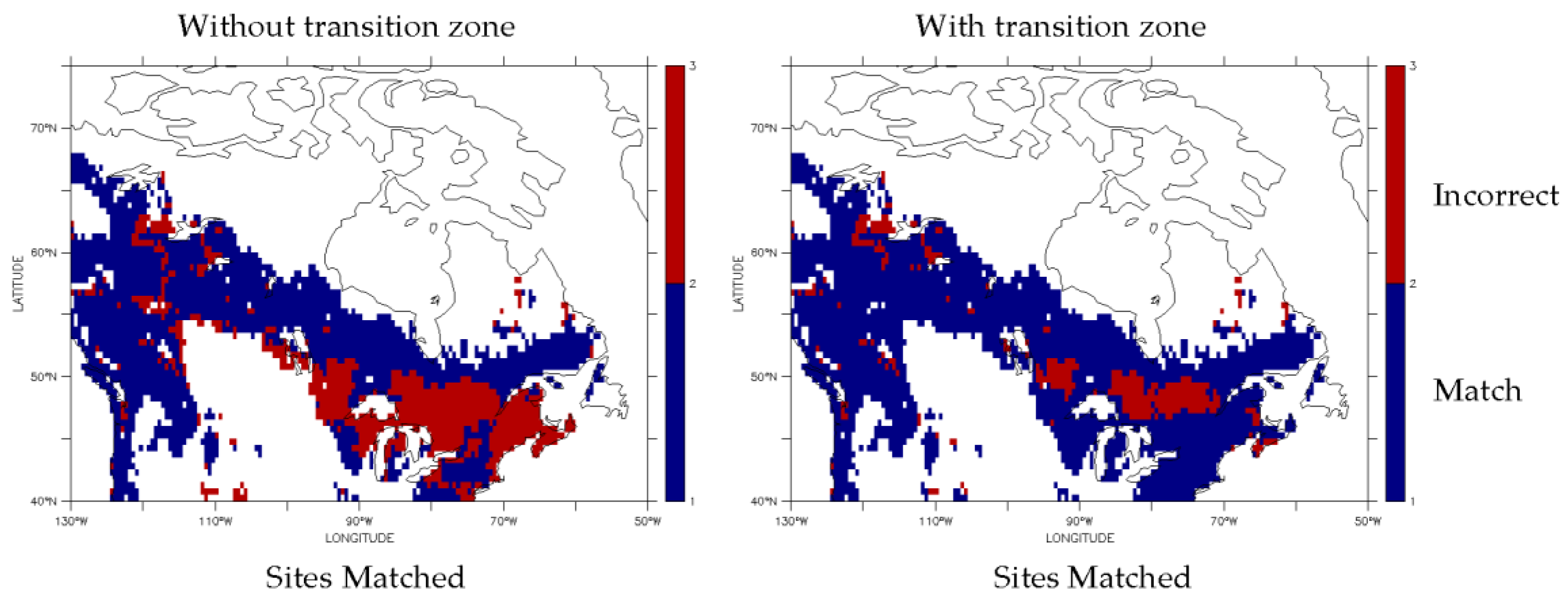
3.1. Dominant Plant Functional Type Distribution
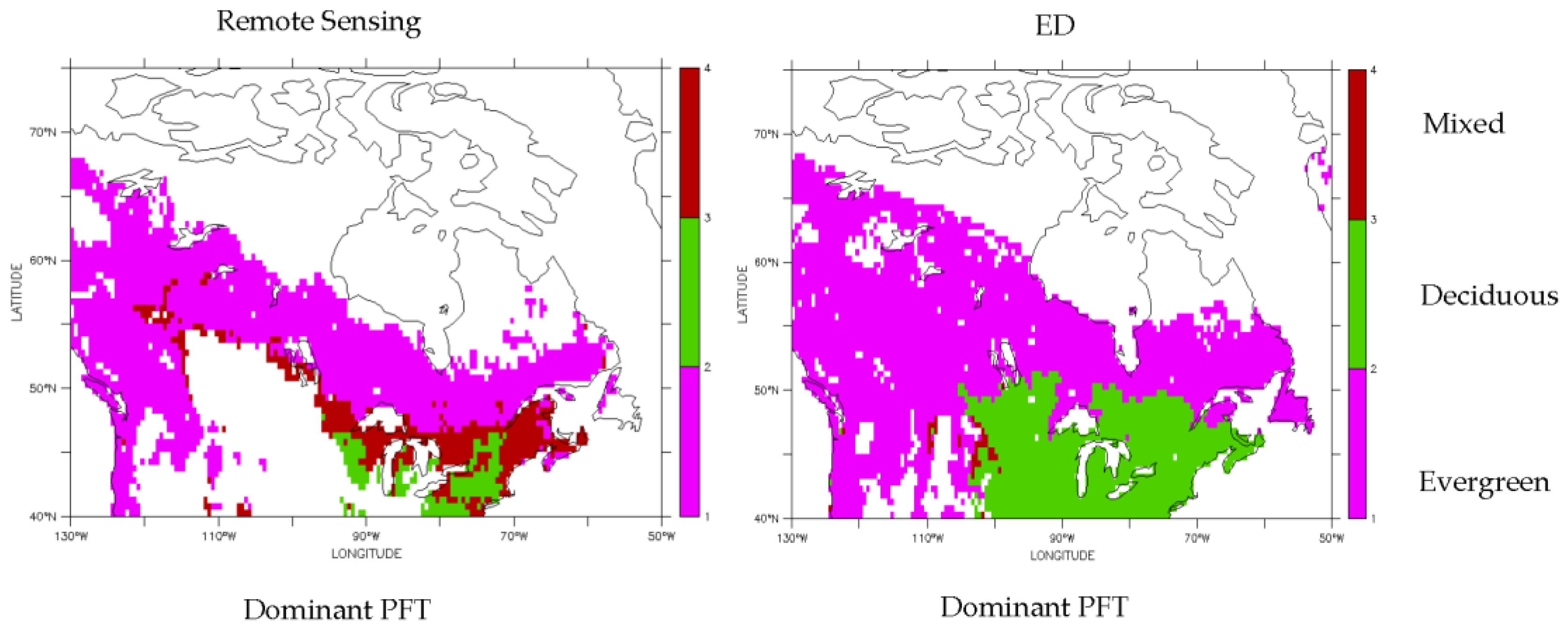
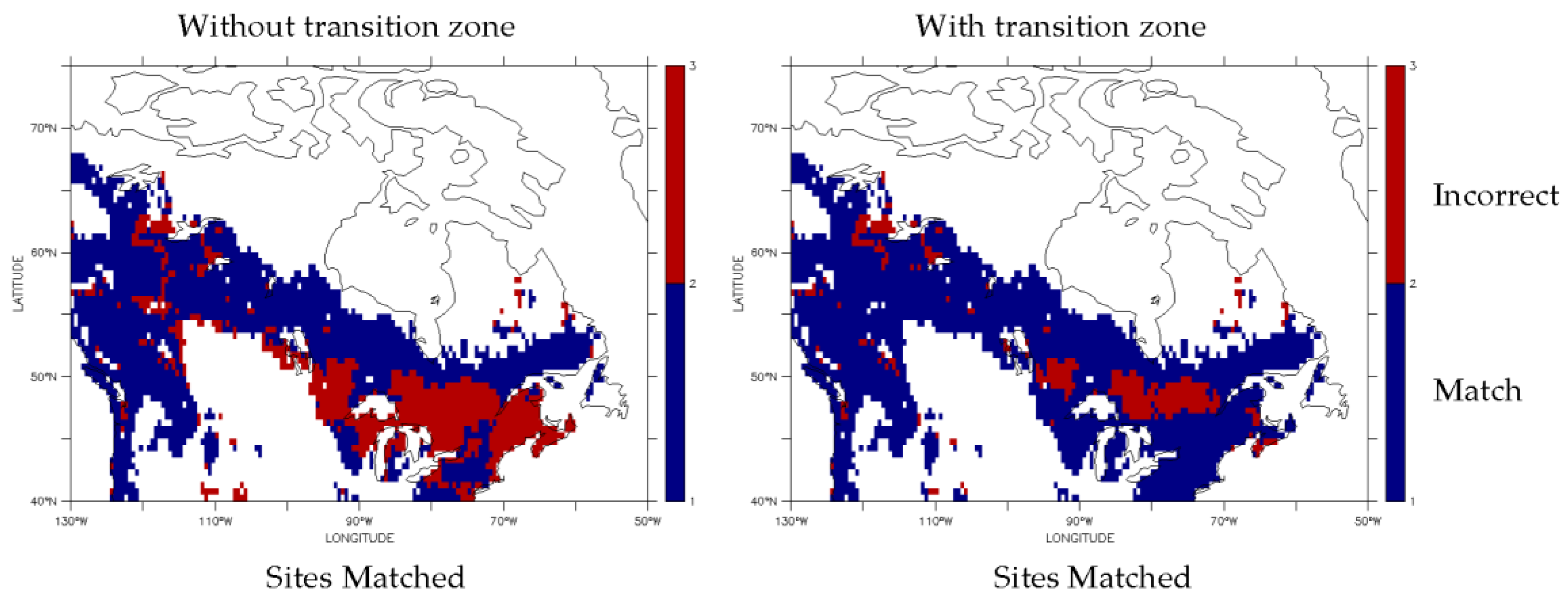
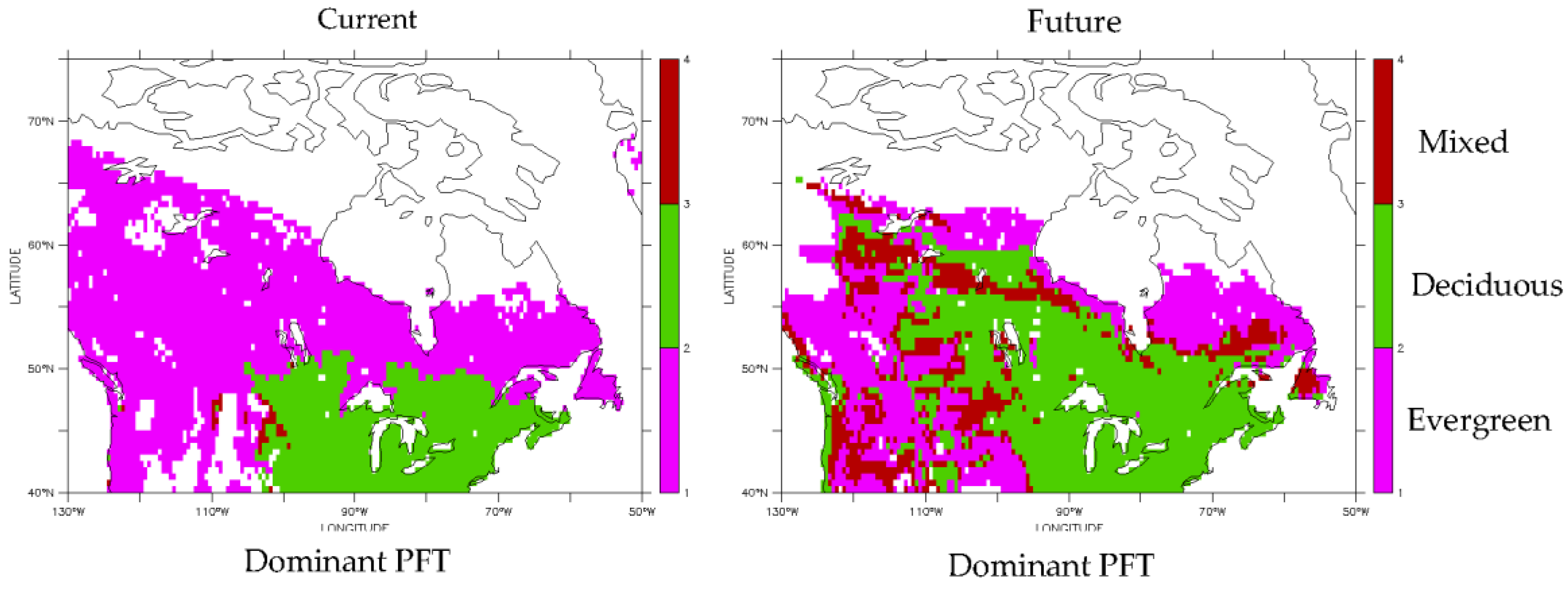
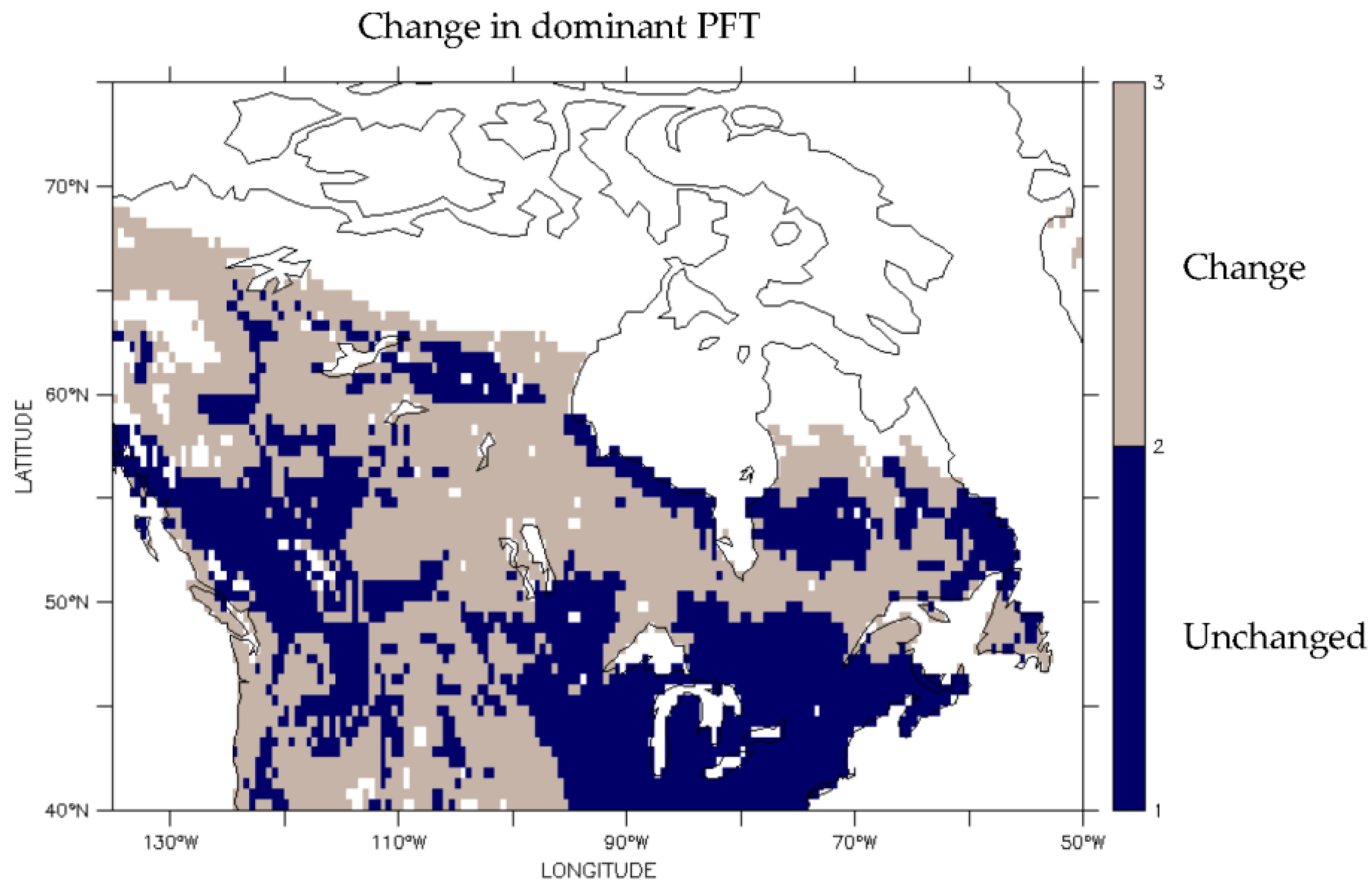
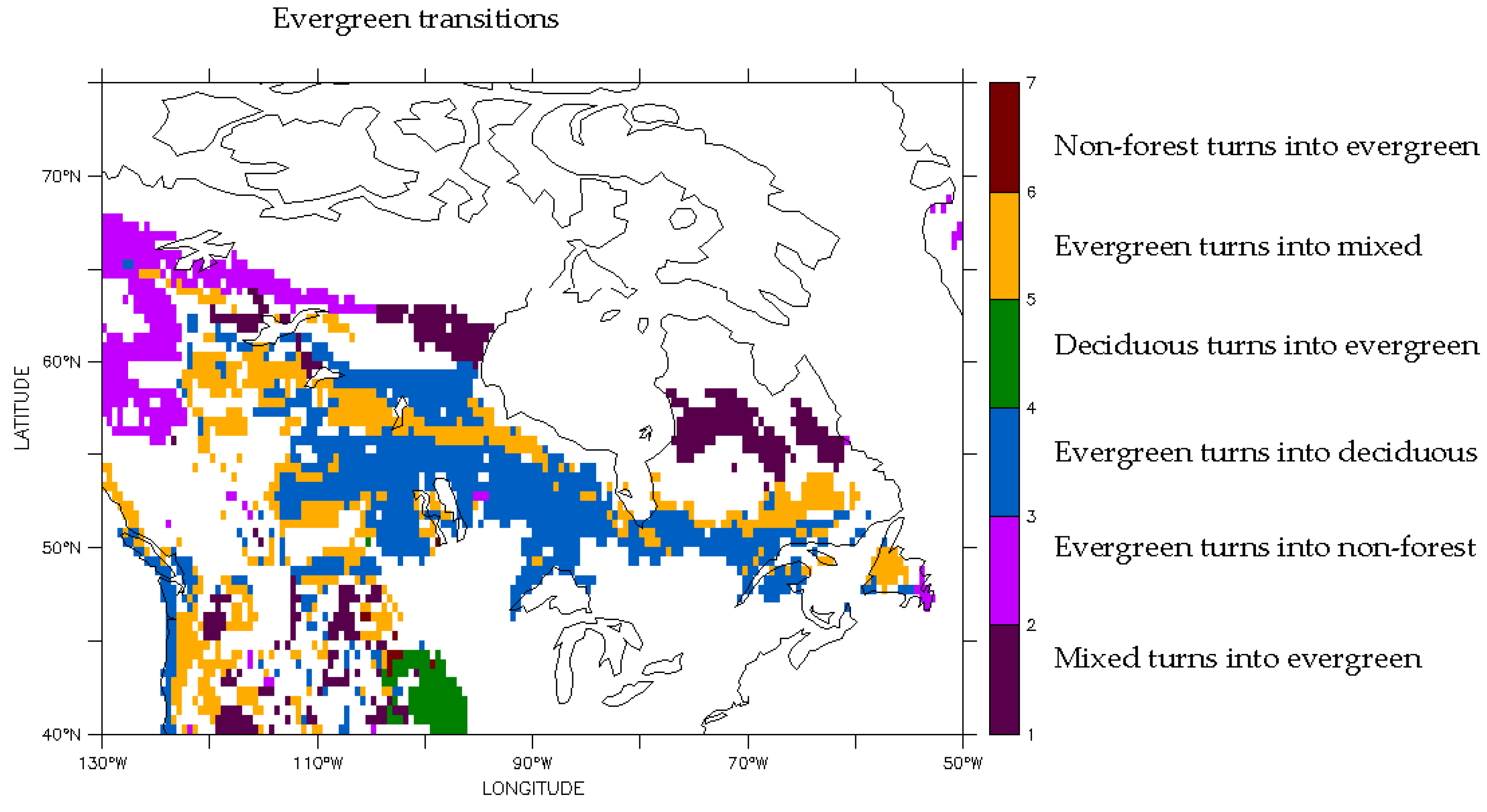
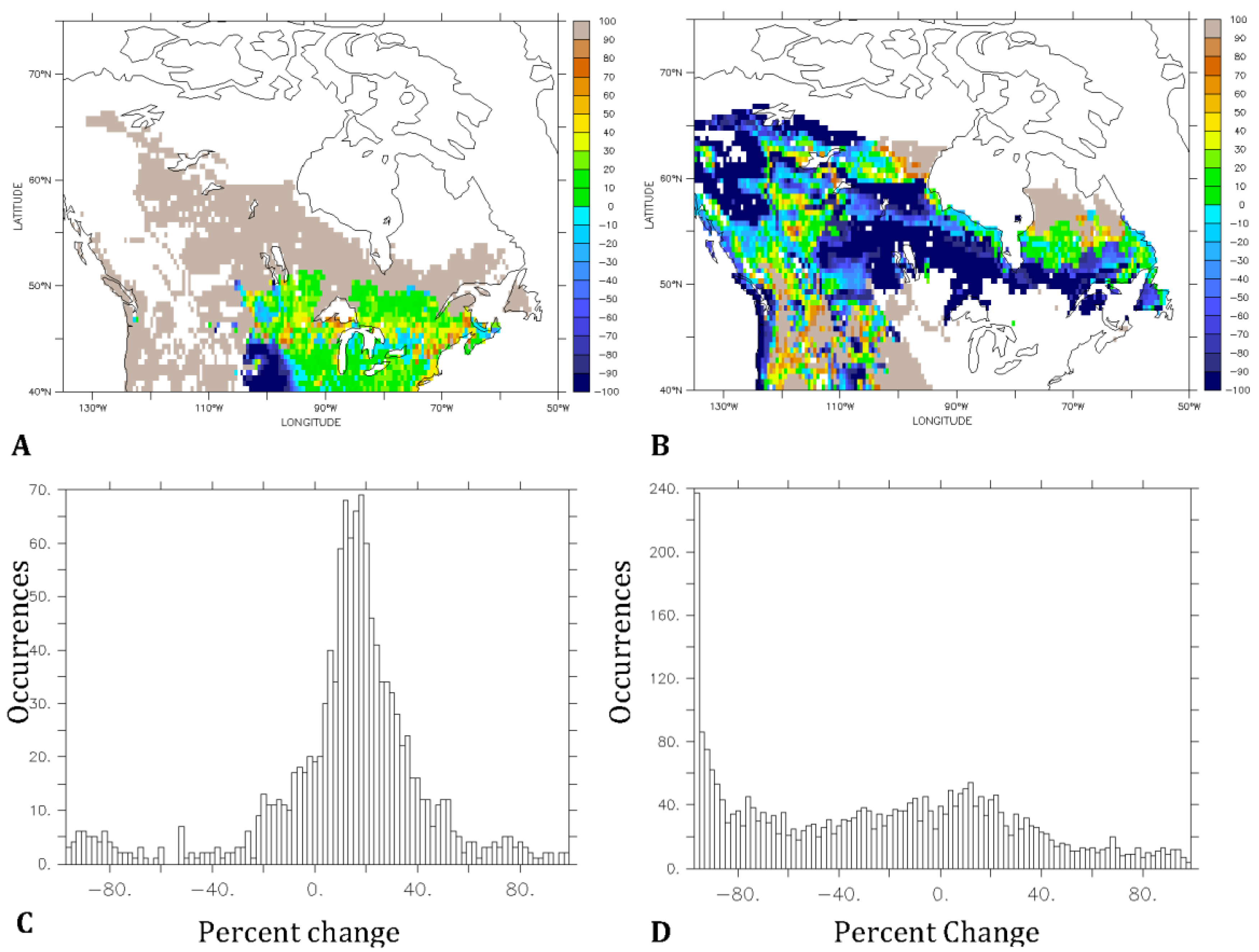
3.2. Predicted Dominant Plant Functional Type Redistribution from Climate Change

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dominant Plant Functional Type Change | Percentage of Sites in Domain |
|---|---|
| Evergreen turns into deciduous | 21 |
| Evergreen turns into mixed | 15 |
| Non-forest turns into evergreen | 8 |
| Evergreen turns into non-forest | 7 |
| Deciduous turns into evergreen | 2 |
| Mixed turns into evergreen | >1 |


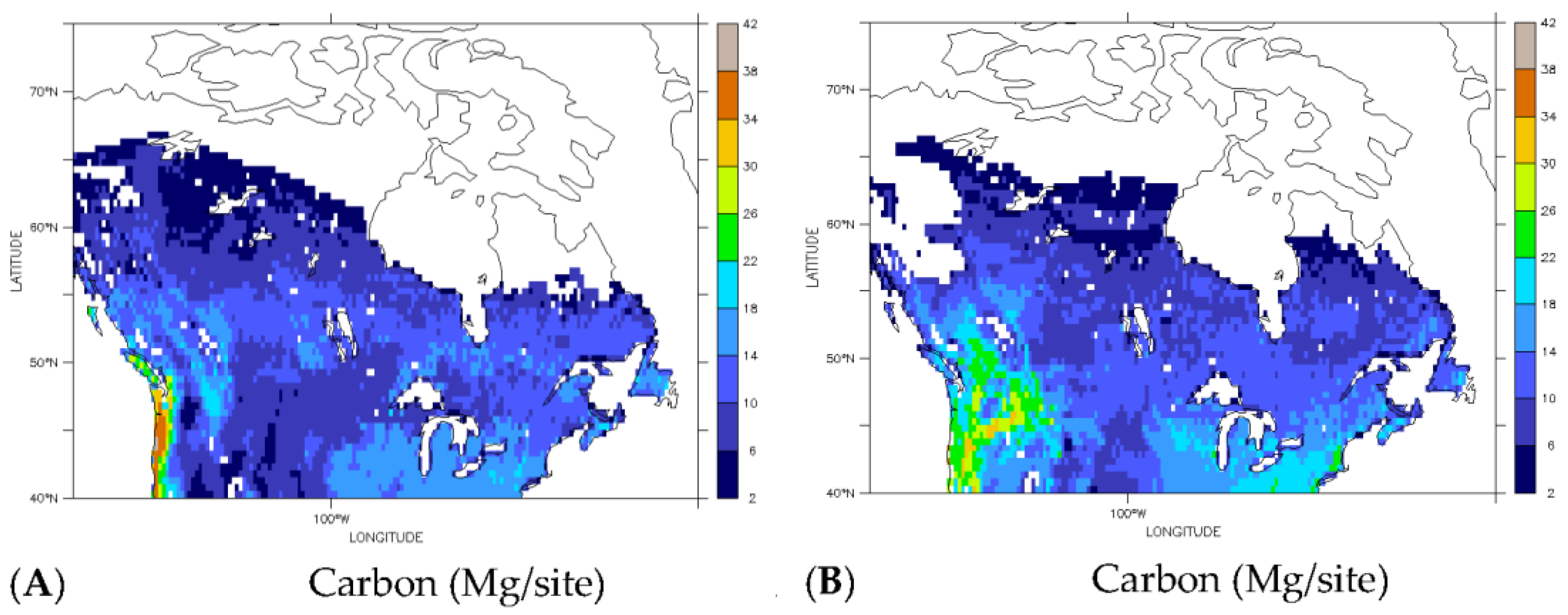
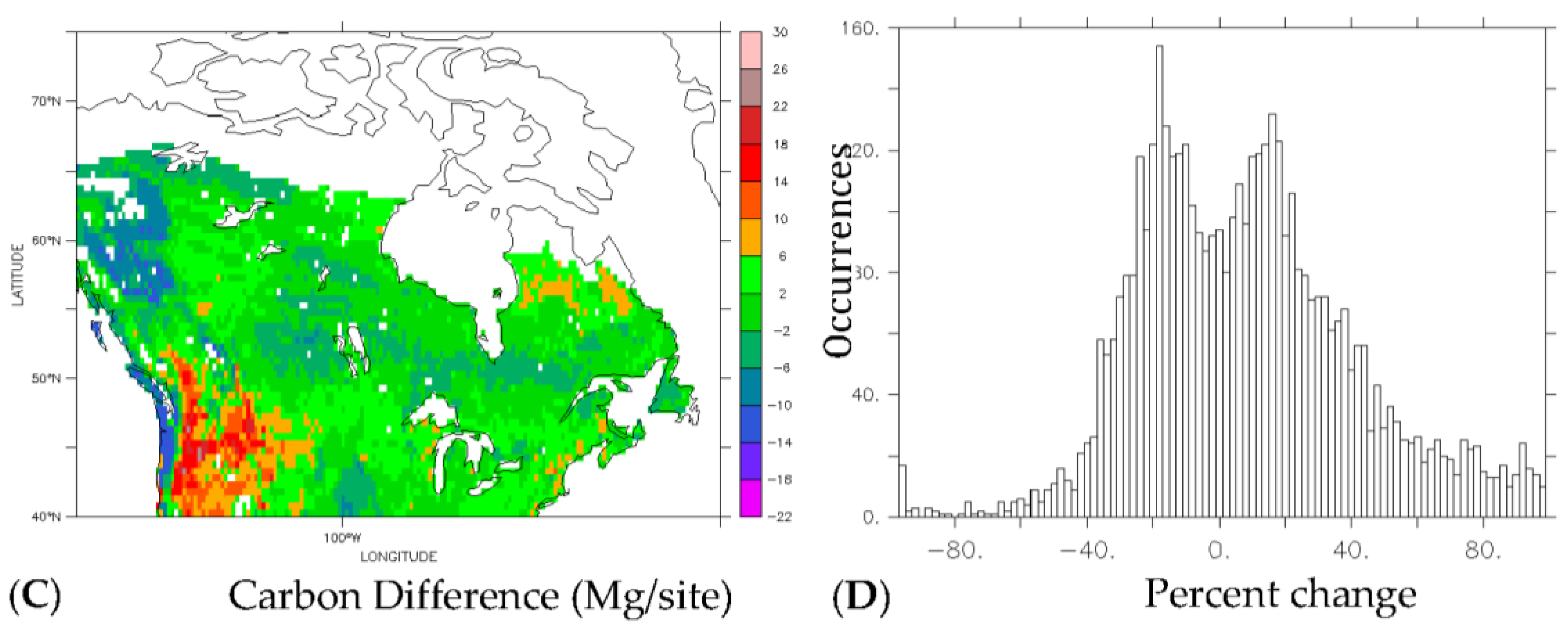
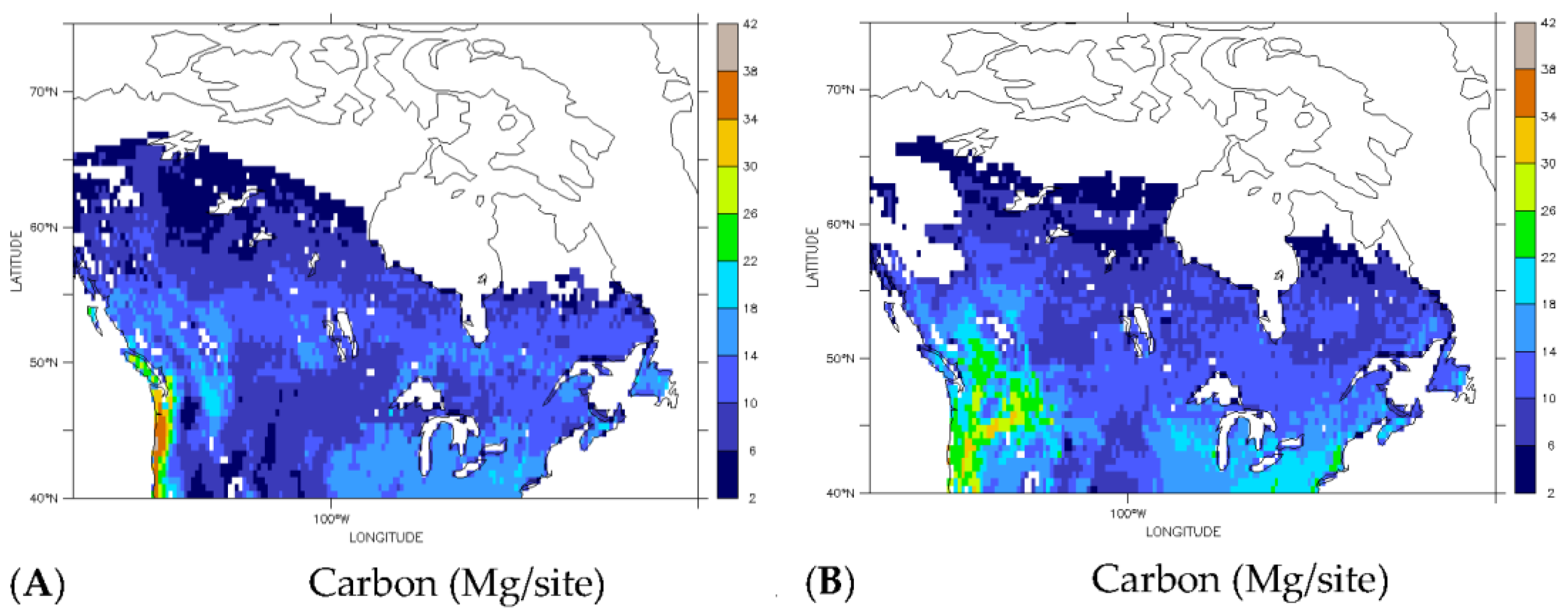
3.3. Implications for Carbon Redistribution and Change
| Carbon (Tg) | ||||
|---|---|---|---|---|
| Carbon Type | Current | Future | Difference | Percent Change |
| Total | 54 | 58 | 4 | 8 |
| Deciduous | 15 | 31 | 16 | 107 |
| Evergreen | 39 | 27 | −12 | −31 |
| Percentage of Total Carbon | ||
|---|---|---|
| Scenario | Deciduous | Evergreen |
| Current | 28 | 72 |
| Future | 54 | 46 |

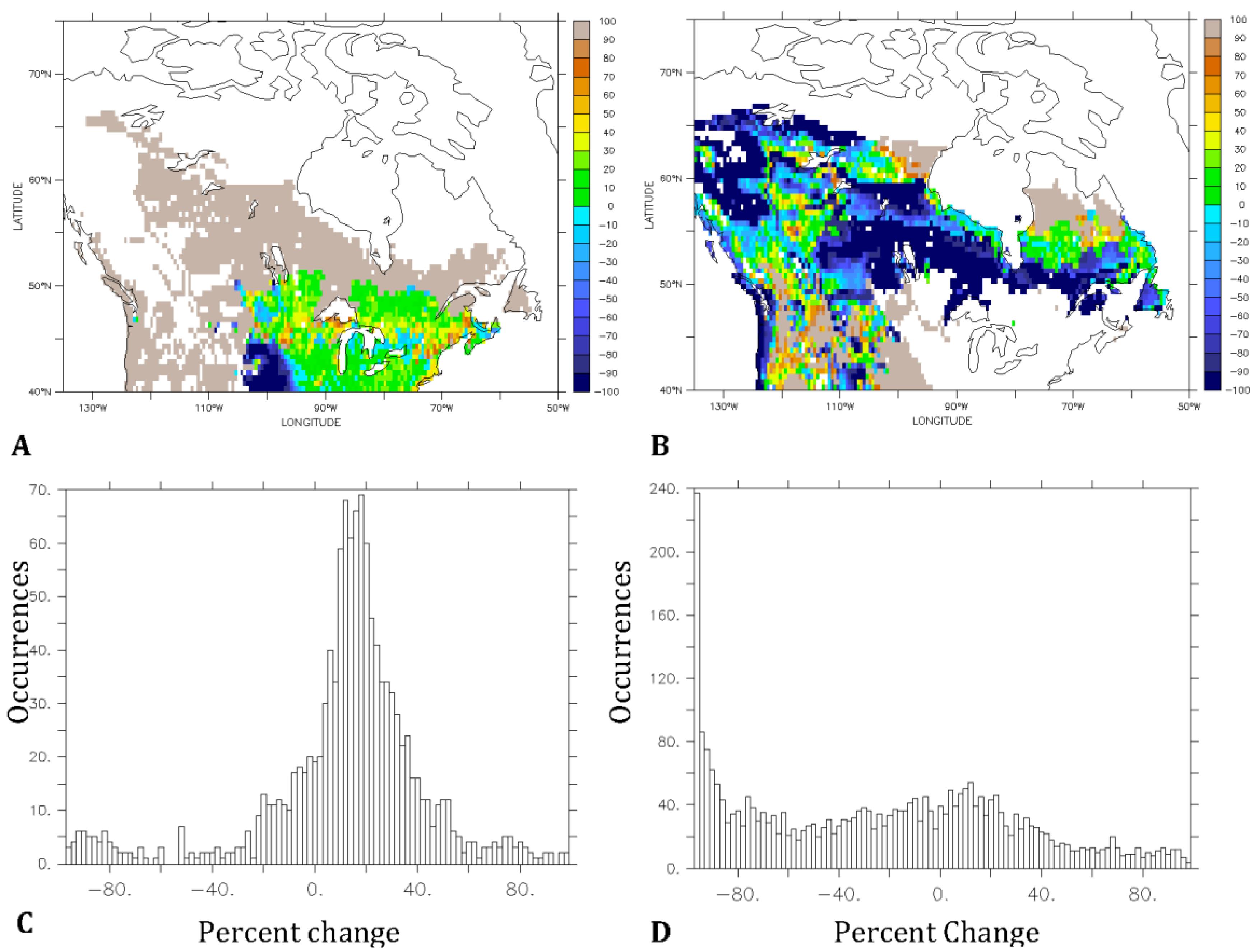
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Köppen, W. Versuch einer Klassifikation der Klimate, vorzugsweise nach ihren Beziehungen zur Pflanzenwelt. Geogr. Z. 1900, 11, 593–611. [Google Scholar]
- Thornthwaite, C.W. The climates of North America: According to a new classification. Geogr. Rev. 1931, 21, 633–655. [Google Scholar] [CrossRef]
- Thornthwaite, C.W. An approach toward a rational classification of climate. Geogr. Rev. 1948, 38, 55–94. [Google Scholar] [CrossRef]
- Holdrige, L.R. Determination of world plant formation from simple climate data. Science 1947, 105, 367–368. [Google Scholar] [CrossRef] [PubMed]
- Dale, V.H.; Joyce, L.A.; McNulty, S.; Neilson, R.P.; Ayres, M.P.; Flannigan, M.D.; Hasnon, P.J.; Irland, L.C.; Lugo, A.E.; Peterson, C.J.; et al. Climate change and forest disturbances. BioScience 2001, 51, 723–734. [Google Scholar] [CrossRef]
- Houghton, D.D. Global Climate Change—Challenges and Impacts. Available online: http://www.ssec.wisc.edu/library/ft/global_climate_change_houghton_2003.pdf (accessed on 1 September 2015).
- Vitousek, P.M.; D’Antonio, C.M.; Loope, L.L.; Westbrooks, R. Biological invasions as global environmental change. Am. Sci. 1996, 84, 468–478. [Google Scholar]
- Gruber, N.; Galloway, J.N. An Earth-system perspective of the global nitrogen cycle. Nature 2008, 451, 293–296. [Google Scholar] [CrossRef] [PubMed]
- Matear, R.J.; Wang, Y.-P.; Lenton, A. Land and ocean nutrient and carbon cycle interactions. Curr. Opin. Environ. Sustain. 2010, 2, 258–263. [Google Scholar] [CrossRef]
- Bonan, G.B. Forests and climate change: Forcings, feedbacks, and the climate benefits of forests. Science 2008, 320, 1444–1449. [Google Scholar] [CrossRef] [PubMed]
- Goulden, M.L.; Munger, J.W.; Fan, S.; Daube, B.C.; Wofsy, S.C. Measurements of carbon sequestration by long-term eddy covariance: Methods and a critical evaluation of accuracy. Glob. Chang. Biol. 1996, 2, 169–182. [Google Scholar] [CrossRef]
- Goetz, S.; Dubayah, R. Advances in remote sensing technology and implications for measuring and monitoring forest carbon stocks and change. Carbon Manag. 2011, 2, 231–244. [Google Scholar] [CrossRef]
- Prentice, K.C.; Fung, I.Y. The sensitivity of terrestrial carbon storage to climate change. Nature 1990, 346, 48–51. [Google Scholar] [CrossRef]
- Neilson, R.P.; Pitelka, L.F.; Solomon, A.M.; Nathan, R.; Midgley, G.F.; Fragoso, J.M.V.; Lischke, H.; Thompson, K. Forecasting regional to global plant migration in response to climate change. BioScience 2005, 55, 749–759. [Google Scholar] [CrossRef]
- Hurtt, G.C.; Dubayah, R.; Drake, J.; Moorcroft, P.; Pacala, S. Beyond potential vegetation: Combining lidar remote sensing and a height-structured ecosystem model for improved estimates of carbon stocks and fluxes. Ecol. Appl. 2004, 14, 873–883. [Google Scholar] [CrossRef]
- Medvigy, D.; Moorcroft, P.R. Predicting ecosystem dynamics at regional scales: An evaluation of a terrestrial biosphere model for the forests of northeastern North America. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 222–235. [Google Scholar] [CrossRef] [PubMed]
- Prentice, K.C. Bioclimatic distribution of vegetation for general-circulation model studies. J. Geophys. Res. Atmos. 1990, 95, 11811–11830. [Google Scholar] [CrossRef]
- Clark, J.S.; Fastie, C.; Hurtt, G.; Jackson, S.T.; Johnson, C.; King, G.A.; Lewis, M.; Lynch, J.; Pacala, S.; Prentice, C.; et al. Reid’s paradox of rapid plant migration. BioScience 1998, 48, 13–24. [Google Scholar] [CrossRef]
- Pacala, S.W.; Hurtt, G.C. Terrestrial vegetation and climate change: Integrating models and experiments. In Biotic Interactions and Global Change; Sinauer Associates Incorporated: Sunderland, MA, USA, 1993. [Google Scholar]
- Lischke, H.; Zimmermann, N.E.; Bolliger, J.; Rickebusch, S.; Loeffler, T.J. TreeMig: A forest-landscape model for simulating spatio-temporal patterns from stand to landscape scale. Ecol. Model. 2006, 199, 409–420. [Google Scholar] [CrossRef]
- Clark, D.B.; Mercado, L.M.; Sitch, S.; Jones, C.D.; Gedney, N.; Best, M.J.; Pryor, M.; Rooney, G.G.; Essery, R.L.H.; Blyth, E.; et al. The Joint UK Land Environment Simulator (JULES), model description—Part 2: Carbon fluxes and vegetation dynamics. Geosci. Model Dev. 2011, 4, 701–722. [Google Scholar] [CrossRef]
- Cox, P.M. Description of the TRIFFID Dynamic Global Vegetation Model; Hadley Cebtre: Exeter, UK, 2001. [Google Scholar]
- Sato, H.; Itoh, A.; Kohyama, T. SEIB-DGVM: A new dynamic global vegetation model using a spatially explicit individual-based approach. Ecol. Model. 2007, 200, 279–307. [Google Scholar] [CrossRef]
- Sato, H.; Ise, T. Effect of plant dynamic processes on African vegetation responses to climate change: Analysis using the spatially explicit individual-based dynamic global vegetation model (SEIB-DGVM). J. Geophys. Res. 2012, 117. [Google Scholar] [CrossRef]
- Hurtt, G.C.; Moorcroft, P.R.; Pacala, S.W.; Levin, S.A. Terrestrial models and global change: Challenges for the future. Glob. Chang. Biol. 1998, 4, 581–590. [Google Scholar] [CrossRef]
- Moorcroft, P.R.; Hurtt, G.C.; Pacala, S.W. A method for scaling vegetation dynamics: The ecosystem demography model (ED). Ecol. Monogr. 2001, 71, 557–586. [Google Scholar] [CrossRef]
- Hurtt, G.; Wickland, D.; Jucks, K.; Bowman, K.; Brown, M.E.; Duren, R.M.; Hagen, S.; Verdy, A. NASA Carbon Monitoring System: Prototype Monitoring, Reporting, and Verification. Available online: http://carbon.nasa.gov/pdfs/2014_CarbonMonitoringSystem_ProgressReport.pdf (accessed on 1 November 2015).
- Prentice, I.C.; Cramer, W.; Harrison, S.P.; Leemans, R.; Monserud, R.A.; Solomon, A.M. A global biome model based on plant physiology and dominance, soil properties and climate. J. Biogeogr. 1992, 19, 117–134. [Google Scholar] [CrossRef]
- Haxeltine, A.; Prentice, I.C. BIOME3: An equilibrium terrestrial biosphere model based on ecophysiological constraints, resource availability, and competition among plant functional types. Glob. Biogeochem. Cycles 1996, 10, 693–709. [Google Scholar] [CrossRef]
- Foley, J.A.; Prentice, I.C.; Ramankutty, N.; Levis, S.; Pollard, D.; Sitch, S.; Haxeltine, A. An integrated biosphere model of land surface processes, terrestrial carbon balance, and vegetation dynamics. Glob. Biogeochem. Cycles 1996, 10, 603–628. [Google Scholar] [CrossRef]
- Duckworth, J.C.; Kent, M.; Ramsay, P.M. Plant functional types: An alternative to taxonomic plant community description in biogeography? Prog. Phys. Geogr. 2000, 24, 515–542. [Google Scholar] [CrossRef]
- Hurtt, G.C.; Pacala, S.W.; Moorcroft, P.R.; Caspersen, J.; Shevliakova, E.; Houghton, R.A.; Moore, B. Projecting the future of the U.S. carbon sink. Proc. Natl. Acad. Sci. USA 2002, 99, 1389–1394. [Google Scholar] [CrossRef] [PubMed]
- Thomas, R.Q.; Hurtt, G.C.; Dubayah, R.; Schilz, M.H. Using lidar data and a height-structured ecosystem model to estimate forest carbon stocks and fluxes over mountainous terrain. Can. J. Remote Sens. 2008, 34, S351–S363. [Google Scholar] [CrossRef]
- Von Caemmerer, S.; Farquhar, G.D. Some relationships between the biochemistry of photosynthesis and the gas exchange of leaves. Planta 1981, 153, 376–387. [Google Scholar] [CrossRef] [PubMed]
- Farquhar, G.D.; Sharkey, T.D. Stomatal conductance and photosynthesis. Annu. Rev. Plant Physiol. 1982, 33, 317–345. [Google Scholar] [CrossRef]
- Vogelmann, J.E.; Howard, S.M.; Yang, L.; Larson, C.R.; Wylie, B.K.; van Driel, J.N. Completion of the 1990’s National Land Cover Data Set for the conterminous United States from Landsat Thematic Mapper data and ancillary data sources. Photogramm. Eng. Remote Sens. 2001, 67, 650–662. [Google Scholar]
- Hurtt, G.C.; Fisk, J.; Thomas, R.Q.; Dubayah, R.; Moorcroft, P.R.; Shugart, H.H. Linking models and data on vegetation structure. J. Geophys. Res. 2010, 115, G00E10. [Google Scholar] [CrossRef]
- Wei, Y.; Liu, S.; Huntzinger, D.N.; Michalak, A.M.; Viovy, N.; Post, W.M.; Schwalm, C.R.; Schaefer, K.; Jacobson, A.R.; Lu, C.; et al. NACP MsTMIP: Global and North American Driver Data for Multi-Model Intercomparison; Data Set; Oak Ridge National Laboratory Distributed Active Archive Center: Oak Ridge, TN, USA, 1994. [Google Scholar]
- Wei, Y.; Liu, S.; Huntzinger, D.N.; Michalak, A.M.; Viovy, N.; Post, W.M.; Schwalm, C.R.; Schaefer, K.; Jacobson, A.R.; Lu, C.; et al. The North American carbon program multi-scale synthesis and terrestrial model intercomparison project—Part 2: Environmental driver data. Geosci. Model Dev. 2013, 6, 5375–5422. [Google Scholar] [CrossRef]
- Mearns, L.O.; Gutowski, W.J.; Jones, R.; Leung, L.-Y.; McGinnis, S.; Nunes, A.M.B.; Qian, Y. A regional climate change assessment program for North America. EOS 2009, 90, 311–312. [Google Scholar] [CrossRef]
- DeFries, R.; Hansen, M.; Townshend, J.R.G.; Janetos, A.C.; Loveland, T.R. 1 Kilometer Tree Cover Continuous Fields, 1.0; Department of Geography, University of Maryland: College Park, MD, USA, 1992–1993. [Google Scholar]
- Schaphoff, S.; Lucht, W.; Gerten, D.; Sitch, S.; Cramer, W.; Prentice, I.C. Terrestrial biosphere carbon storage under alternative climate projections. Clim. Chang. 2006, 74, 97–122. [Google Scholar] [CrossRef]
- Solomon, A.M.; Kirilenko, A.P. Climate change and terrestrial biomass: What if trees do not migrate? Glob. Ecol. Biogeogr. Lett. 1997, 6, 139–148. [Google Scholar] [CrossRef]
- Rehfeldt, G.E.; Crookston, N.L.; Warwell, M.V.; Evans, J.S. Empirical analyses of plant-climate relationships for the Western United States. Int. J. Plant Sci. 2006, 167, 1123–1150. [Google Scholar] [CrossRef]
- Rehfeldt, G.E.; Ying, C.C.; Spittlehouse, D.L.; Hamilton, D.A., Jr. Genetic responses to climate in pinus contorta: Niche breadth, climate change, and reforestation. Ecol. Monogr. 1999, 69, 375–407. [Google Scholar] [CrossRef]
- Rehfeldt, G.E.; Tchebakova, N.M. Intraspecific responses to climate in Pinus sylvestris. Glob. Chang. Biol. 2002, 8, 912–929. [Google Scholar] [CrossRef]
- Sykes, M.T.; Prentice, I.C. Boreal forest futures—Modeling the controls on tree species range limits and transient responses to climate-change. Water Air Soil Pollut. 1995, 82, 415–428. [Google Scholar] [CrossRef]
- Emanuel, W.R.; Shugart, H.H.; Stevenson, M.P. Climatic change and the broad-scale distribution of terrestrial ecosystem complexes. Clim. Chang. 1985, 7, 29–43. [Google Scholar] [CrossRef]
- Ranson, K.J.; Sun, G.; Kharuk, V.I.; Kovacs, K. Assessing tundra-taiga boundary with multi-sensor satellite data. Remote Sens. Environ. 2004, 93, 283–295. [Google Scholar] [CrossRef]
- Montesano, P.M.; Nelson, R.; Sun, G.; Margolis, H. MODIS tree cover validation for the circumpolar taiga-tundra transition zone. Remote Sens. Environ. 2009, 113, 2130–2141. [Google Scholar] [CrossRef]
- Chapin, F.S.; McGuire, A.D.; Ruess, R.W.; Hollingsworth, T.N.; Mack, M.C.; Johnstone, J.F.; Kasischke, E.S.; Euskirchen, E.S.; Jones, J.B.; Jorgenson, M.T.; et al. Resilience of Alaska’s boreal forest to climatic change. Can. J. For. Res. 2010, 40, 1360–1370. [Google Scholar] [CrossRef]
- Soja, A.J.; Tchebakova, N.M.; French, N.H.F.; Flannigan, M.D.; Shugart, H.H.; Stocks, B.J.; Sukhinin, A.I.; Parfenova, E.I.; Chapin, F.S., III; Stackhouse, P.W., Jr. Climate-induced boreal forest change: Predictions versus current observations. Glob. Planet. Chang. 2007, 56, 274–296. [Google Scholar] [CrossRef]
- Grace, J. Impacts of climate change on the tree line. Ann. Bot. 2002, 90, 537–544. [Google Scholar] [CrossRef] [PubMed]
- Goldblum, D.; Rigg, L.S. Tree growth response to climate change at the deciduous boreal forest ecotone, Ontario, Canada. Can. J. For. Res. 2005, 35, 2709–2718. [Google Scholar] [CrossRef]
- Bachelet, D.; Neilson, R.P.; Lenihan, J.M.; Drapek, R.J. Climate change effects on vegetation distribution and carbon budget in the United States. Ecosystems 2001, 4, 164–185. [Google Scholar] [CrossRef]
- Iverson, L.R.; Schwartz, M.W.; Prasad, A.M. Potential colonization of newly available tree-species habitat under climate change: An analysis for five eastern US species. Landsc. Ecol. 2004, 19, 787–799. [Google Scholar] [CrossRef]
- Küchler, A.W. Problems in classifying and mapping vegetation for ecological regionalization. Ecology 1973, 54, 512–523. [Google Scholar] [CrossRef]
- Denslow, J.S. Patterns of plant-species diversity during succession under different disturbance regimes. Oecologia 1980, 46, 18–21. [Google Scholar] [CrossRef]
- Bailey, R.G. Delineation of ecosystem regions. Environ. Manag. 1983, 7, 365–373. [Google Scholar] [CrossRef]
- Pitelka, L.F. Plant migration and climate change: A more realistic portrait of plant migration is essential to predicting biological responses to global warming in a world drastically altered by human activity. Am. Sci. 1997, 85, 464–473. [Google Scholar]
- Collingham, Y.C.; Huntley, B. Impacts of habitat fragmentation and patch size upon migration rates. Ecol. Appl. 2000, 10, 131–144. [Google Scholar] [CrossRef]
- Plotnick, R.E.; Gardner, R.H. A general model for simulating the effects of landscape heterogeneity and disturbance on community patterns. Ecol. Model. 2002, 147, 171–197. [Google Scholar] [CrossRef]
- Lavorel, S.; McIntyre, S.; Landsberg, J.; Forbes, T. Plant functional classifications: From general groups to specific groups based on response to disturbance. Trends Ecol. Evol. 1997, 12, 474–478. [Google Scholar] [CrossRef]
- Svenning, J.C.; Skov, F. Limited filling of the potential range in European tree species. Ecol. Lett. 2004, 7, 565–573. [Google Scholar] [CrossRef]
- Fahrig, L. Effect of habitat fragmentation on the extinction threshold: A synthesis. Ecol. Appl. 2002, 12, 346–353. [Google Scholar] [CrossRef]
- Kot, M.; Lewis, M.A.; van den Driessche, P. Dispersal data and the spread of invading organisms. Ecology 1996, 77, 2027–2042. [Google Scholar] [CrossRef]
- Peel, M.C.; Finlayson, B.L.; McMahon, T.A. Updated world map of the Koppen-Geiger climate classification. Hydrol. Earth Syst. Sci. 2007, 11, 1633–1644. [Google Scholar] [CrossRef]
- Midgley, G.F.; Thuiller, W.; Higgins, S.I. Plant species migration as a key uncertainty in predicting future impacts of climate change on ecosystems: Progress and challenges. In Global Change—The IGBP Series; Springer: Berling/Heidelberg, Germany; New York, NY, USA, 2007; pp. 129–137. [Google Scholar]
- Peñuelas, J.; Ogaya, R.; Boada, M.; Jump, A.S. Migration, invasion and decline: Changes in recruitment and forest structure in a warming-linked shift of European beech forest in Catalonia (NE Spain). Ecography 2007, 30, 829–837. [Google Scholar] [CrossRef]
- Lenoir, J.; Gegout, J.C.; Marquet, P.A.; de Ruffray, P.; Brisse, H. A significant upward shift in plant species optimum elevation during the 20th Century. Science 2008, 320, 1768–1771. [Google Scholar] [CrossRef] [PubMed]
- Iverson, L.; Prasad, A.; Matthews, S. Modeling potential climate change impacts on the trees of the northeastern United States. Mitig. Adapt. Strateg. Glob. Chang. 2007, 13, 487–516. [Google Scholar] [CrossRef]
- Iverson, L.R.; Prasad, A.M.; Matthews, S.N.; Peters, M.P. Lessons learned while integrating habitat, dispersal, disturbance, and life-history traits into species habitat models under climate change. Ecosystems 2011, 14, 1005–1020. [Google Scholar] [CrossRef]
- Matthews, S.N.; Iverson, L.R.; Peters, M.P.; Prasad, A.M.; Subburayalu, S. Assessing and comparing risk to climate changes among forested locations: Implications for ecosystem services. Landsc. Ecol. 2014, 29, 213–228. [Google Scholar] [CrossRef]
- Duveneck, M.J.; Scheller, R.M.; White, M.A.; Handler, S.D.; Ravenscroft, C. Climate change effects on northern Great Lake (USA) forests: A case for preserving diversity. Ecosphere 2014, 5, 23. [Google Scholar] [CrossRef]
- Zolkos, S.G.; Jantz, P.; Cormier, T.; Iverson, L.R.; McKenney, D.W.; GOETZ, S.J. Projected tree species redistribution under climate change: Implications for ecosystem vulnerability across protected areas in the Eastern United States. Ecosystems 2015, 18, 202–220. [Google Scholar] [CrossRef]
- Brandt, L.; He, H.; Iverson, L.; Thompson, F.R., III; Butler, P.; Handler, S.; Janowiak, M.; Shannon, P.D.; Swanston, C.; Albrecht, M.; et al. Central Hardwoods Ecosystem Vulnerability Assessment and Synthesis; USDA Forest Service: Newton Square, MA, USA, 2014. [Google Scholar]
- Kasischke, E.S.; Verbyla, D.L.; Rupp, T.S.; McGuire, A.D.; Murphy, K.A.; Jandt, R.; Barnes, J.L.; Hoy, E.E.; Duffy, P.A.; Calef, M.; et al. Alaska’s changing fire regime—Implications for the vulnerability of its boreal forests. Can. J. For. Res. 2010, 40, 1313–1324. [Google Scholar] [CrossRef]
- McGuire, A.D.; Macdonald, R.W.; Schuur, E.A.; Harden, J.W.; Kuhry, P.; Hayes, D.J.; Christensen, T.R.; Heimann, M. The carbon budget of the northern cryosphere region. Curr. Opin. Environ. Sustain. 2010, 2, 231–236. [Google Scholar] [CrossRef]
- Beck, P.S.A.; Goetz, S.J.; Mack, M.C.; Alexander, H.D.; Jin, Y.; Randerson, J.T.; Loranity, M.M. The impacts and implications of an intensifying fire regime on Alaskan boreal forest composition and albedo. Glob. Chang. Biol. 2011, 17, 2853–2866. [Google Scholar] [CrossRef]
- Turetsky, M.R.; Kane, E.S.; Harden, J.W.; Ottmar, R.D.; Manies, K.L.; Hoy, E.; Kasischke, E.S. Recent acceleration of biomass burning and carbon losses in Alaskan forests and peatlands. Nat. Geosci. 2010, 4, 27–31. [Google Scholar] [CrossRef]
- Ciais, P.; Canadell, J.G.; Luyssaert, S.; Chevallier, F.; Shvidenko, A.; Poussi, Z.; Jonas, M.; Peylin, P.; King, A.W.; Schulze, E.-D.; et al. Can we reconcile atmospheric estimates of the Northern terrestrial carbon sink with land-based accounting? Curr. Opin. Environ. Sustain. 2010, 2, 225–230. [Google Scholar] [CrossRef]
- Dietze, M.C.; Vargas, R.; Richardson, A.D.; Stoy, P.C.; Barr, A.G.; Anderson, R.S.; Arain, M.A.; Baker, I.T.; Black, T.A.; Chen, J.M.; et al. Characterizing the performance of ecosystem models across time scales: A spectral analysis of the North American Carbon Program site-level synthesis. J. Geophys. Res. 2011, 116, G04029. [Google Scholar] [CrossRef]
- Fisher, R.; McDowell, N.; Purves, D.; Moorcroft, P.; Sitch, S.; Cox, P.; Huntingford, C.; Meir, P.; Ian Woodward, F. Assessing uncertainties in a second-generation dynamic vegetation model caused by ecological scale limitations. New Phytol. 2010, 187, 666–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Flanagan, S.A.; Hurtt, G.C.; Fisk, J.P.; Sahajpal, R.; Hansen, M.C.; Dolan, K.A.; Sullivan, J.H.; Zhao, M. Potential Vegetation and Carbon Redistribution in Northern North America from Climate Change. Climate 2016, 4, 2. https://doi.org/10.3390/cli4010002
Flanagan SA, Hurtt GC, Fisk JP, Sahajpal R, Hansen MC, Dolan KA, Sullivan JH, Zhao M. Potential Vegetation and Carbon Redistribution in Northern North America from Climate Change. Climate. 2016; 4(1):2. https://doi.org/10.3390/cli4010002
Chicago/Turabian StyleFlanagan, Steven A., George C. Hurtt, Justin P. Fisk, Ritvik Sahajpal, Matthew C. Hansen, Katelyn A. Dolan, Joe H. Sullivan, and Maosheng Zhao. 2016. "Potential Vegetation and Carbon Redistribution in Northern North America from Climate Change" Climate 4, no. 1: 2. https://doi.org/10.3390/cli4010002
APA StyleFlanagan, S. A., Hurtt, G. C., Fisk, J. P., Sahajpal, R., Hansen, M. C., Dolan, K. A., Sullivan, J. H., & Zhao, M. (2016). Potential Vegetation and Carbon Redistribution in Northern North America from Climate Change. Climate, 4(1), 2. https://doi.org/10.3390/cli4010002