Molecular Mapping of D1, D2 and ms5 Revealed Linkage between the Cotyledon Color Locus D2 and the Male-Sterile Locus ms5 in Soybean


Abstract
:1. Introduction
2. Materials and Methods
2.1. Microscopy: ms5
2.2. Mapping Population: ms5

{kind=link}
{kind=link}
{kind=link}
| Population | No. F2 plants | No. F2:3 families | ||||||
|---|---|---|---|---|---|---|---|---|
| Fertile | Sterile | χ2 (3:1) | P | All fertile | Segregating | χ2 (1:2) | P | |
| A12-g-8A | 76 | 27 | 0.08 | 0.78 | 25 | 51 | 0.01 | 0.94 |
2.3. Progeny Testing: ms5
2.4. Mapping Population: d1d2
| Population | No. F2 plants | χ2 (15:1) | P | No. F2 families | χ2 (7:4:4:1) | P | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| Yellow | Green | All yellow | 15:1 | 3:1 | All green | |||||
| A09-308 | 118 | 5 | 1 | 0.32 | 61 | 31 | 26 | 5 | 2.64 | 0.45 |
| A09-309 | 118 | 7 | 0.01 | 0.76 | 48 | 26 | 44 | 7 | 6.99 | 0.07 |
| A09-310 | 117 | 9 | 0.17 | 0.68 | 55 | 29 | 33 | 9 | 0.43 | 0.94 |
| Total | 353 | 21 | 1.18 | 0.28 | 164 | 86 | 103 | 21 | 1.81 | 0.61 |
2.5. Progeny Testing: d1d2
2.6. Molecular Analysis
2.7. Data Analysis
3. Results
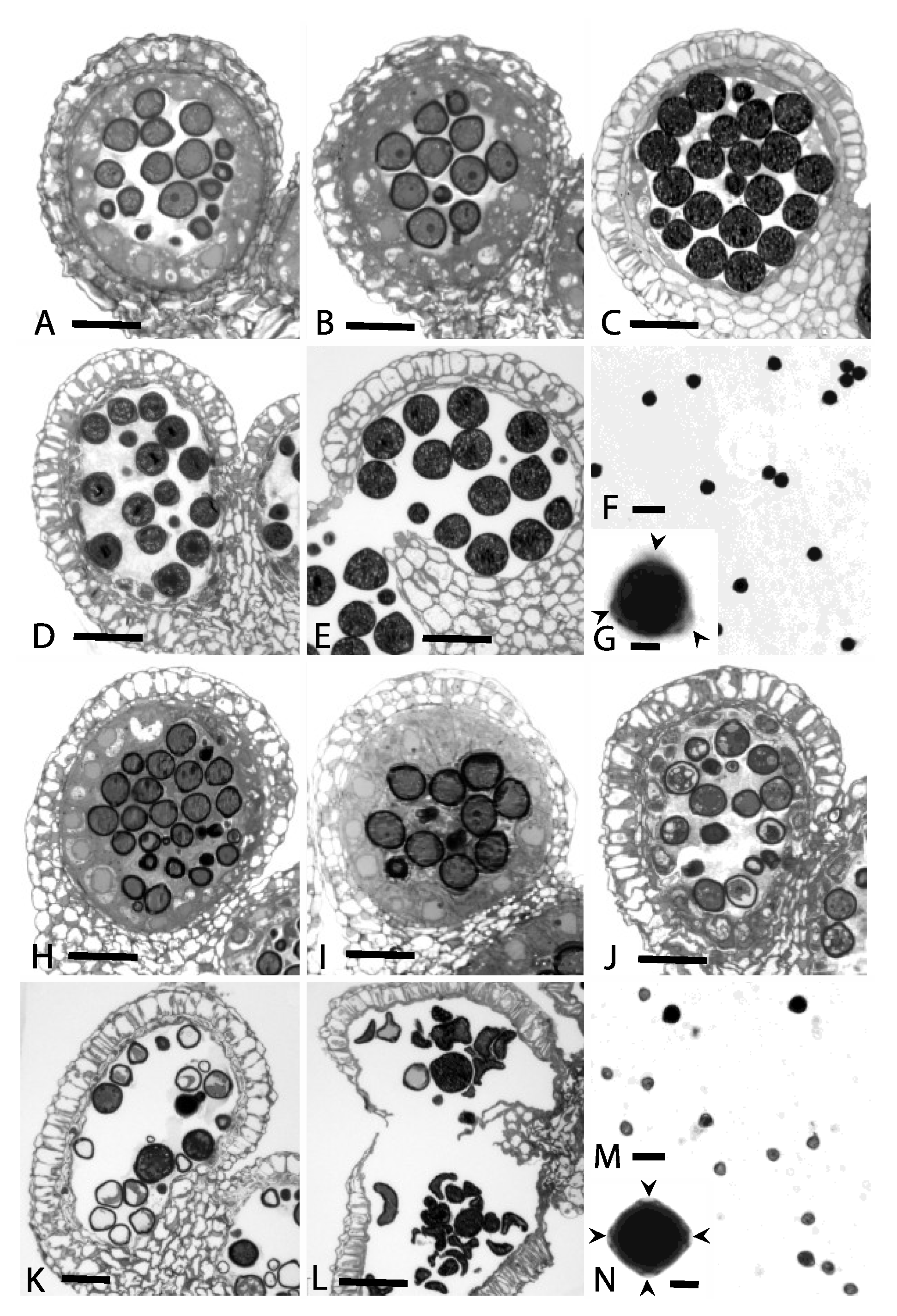
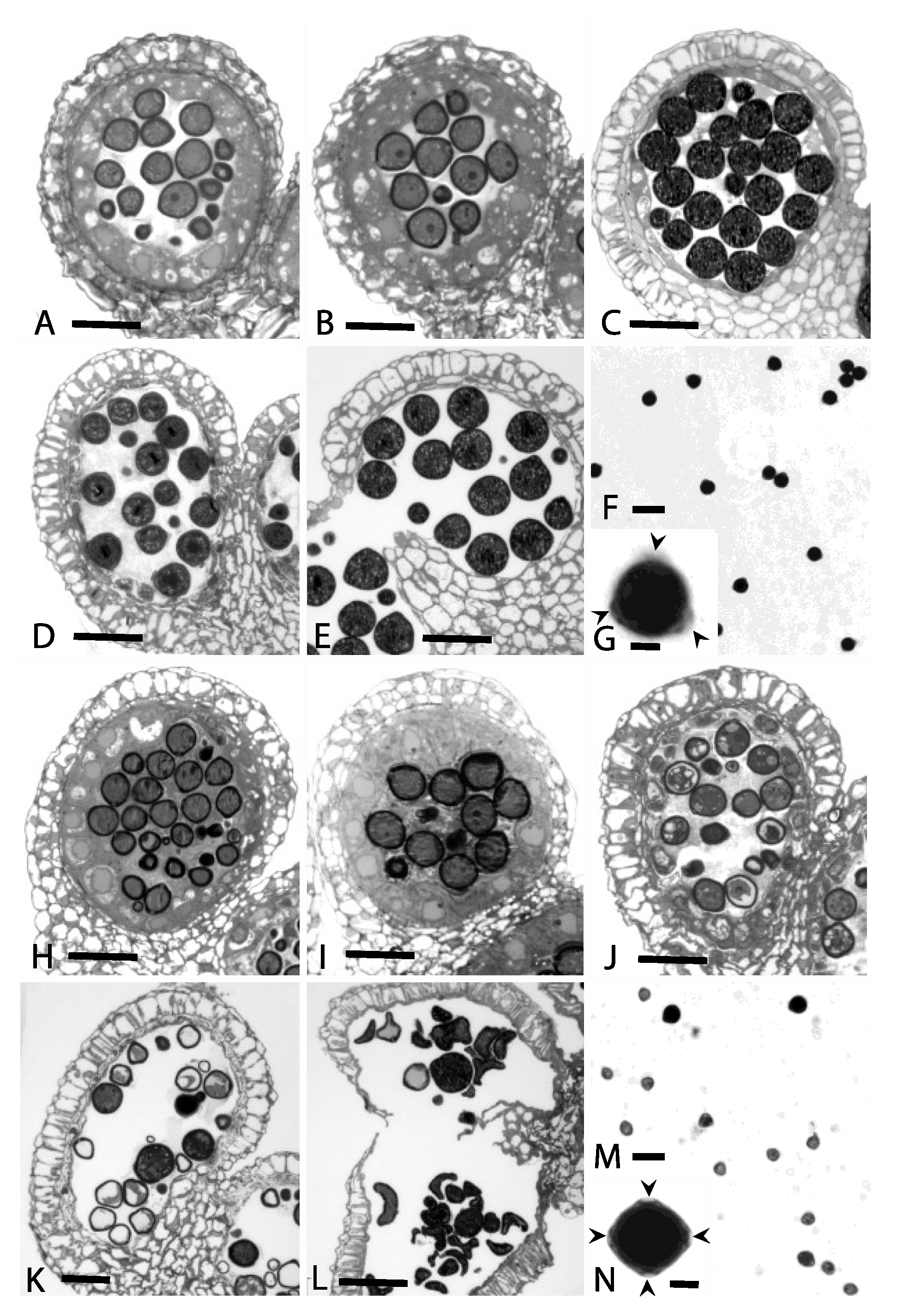
3.1. Cytological Analyses
3.2. Progeny Testing: ms5
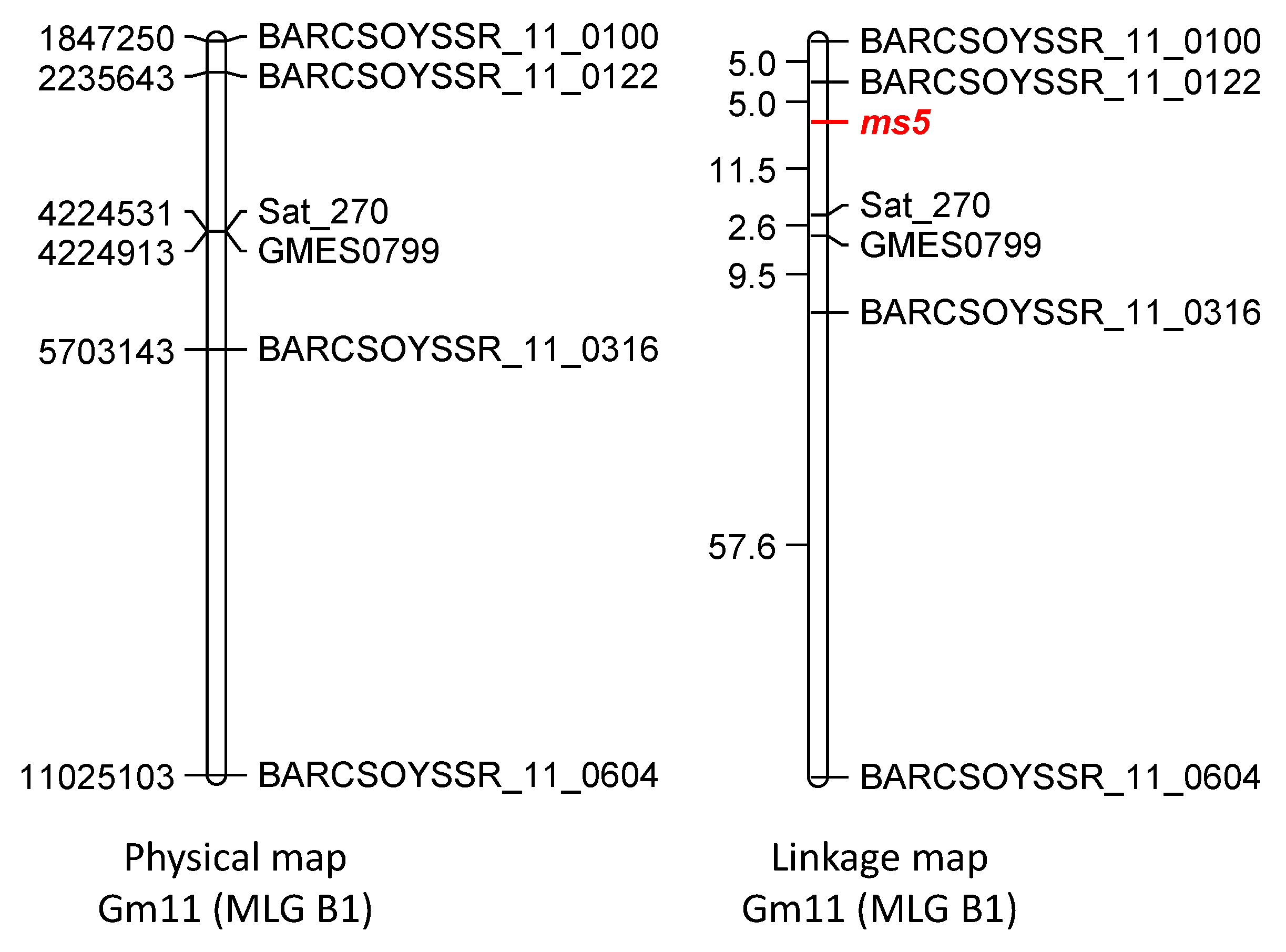
3.3. Molecular Mapping of ms5
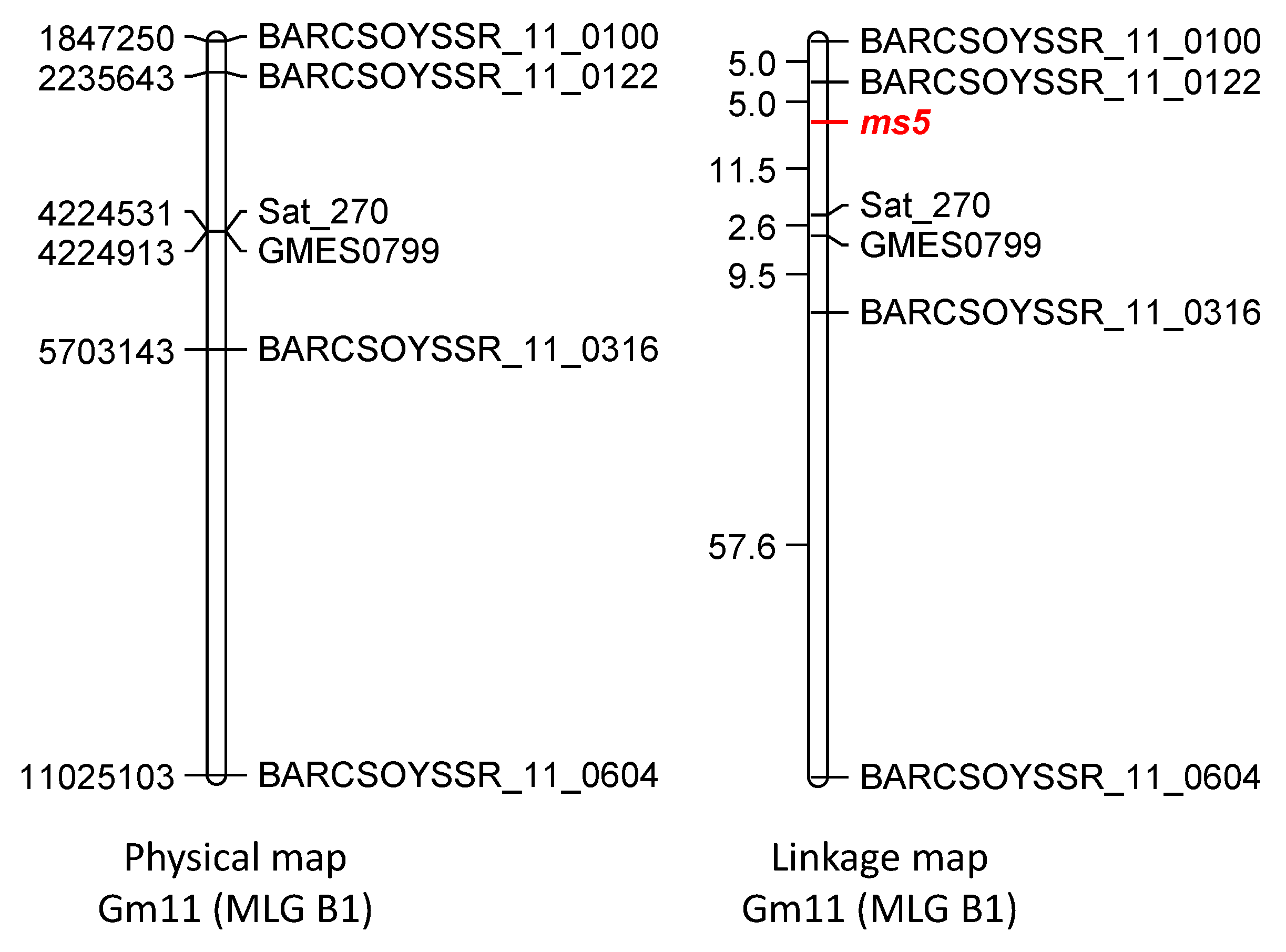
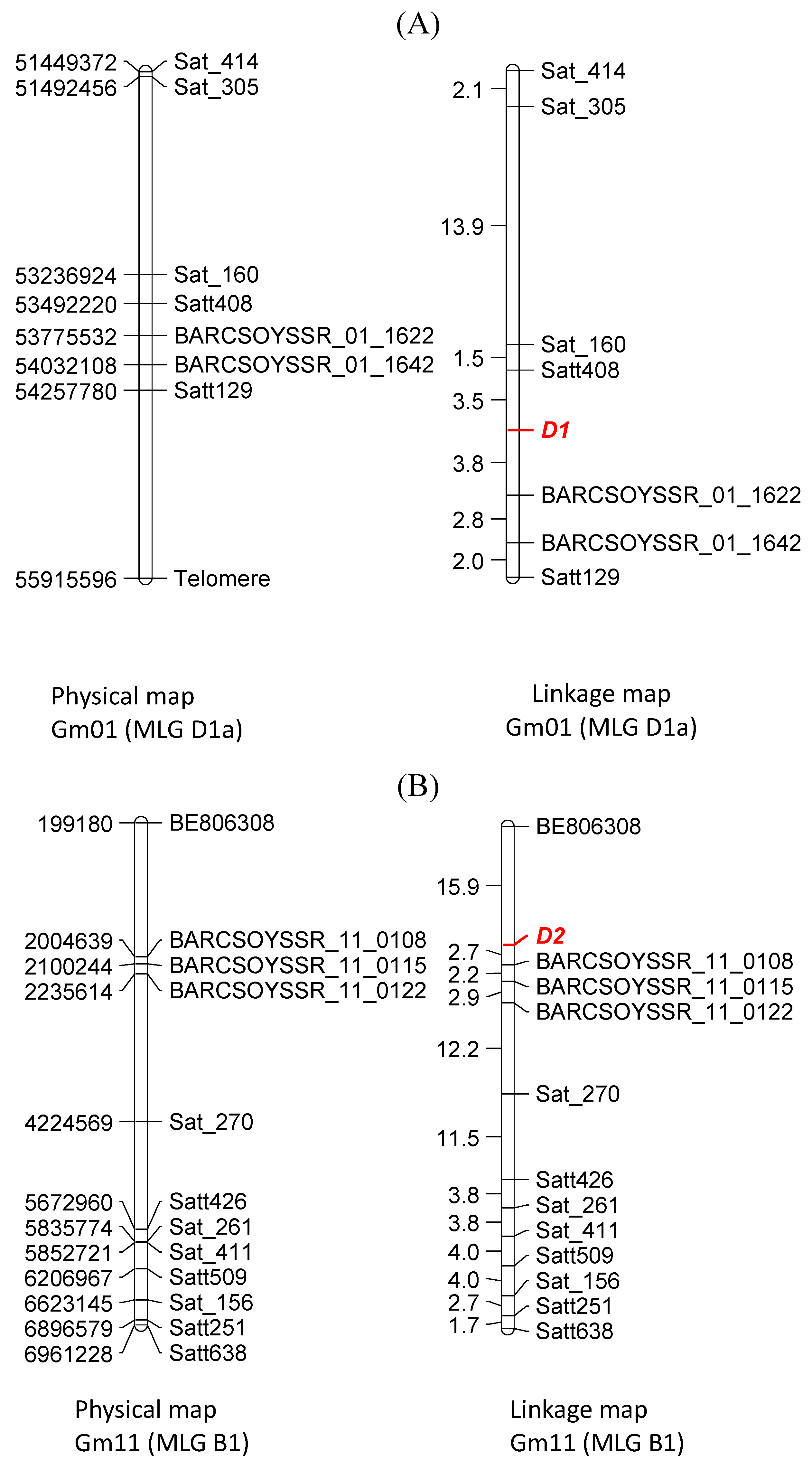
3.4. Molecular Mapping of D1 and D2
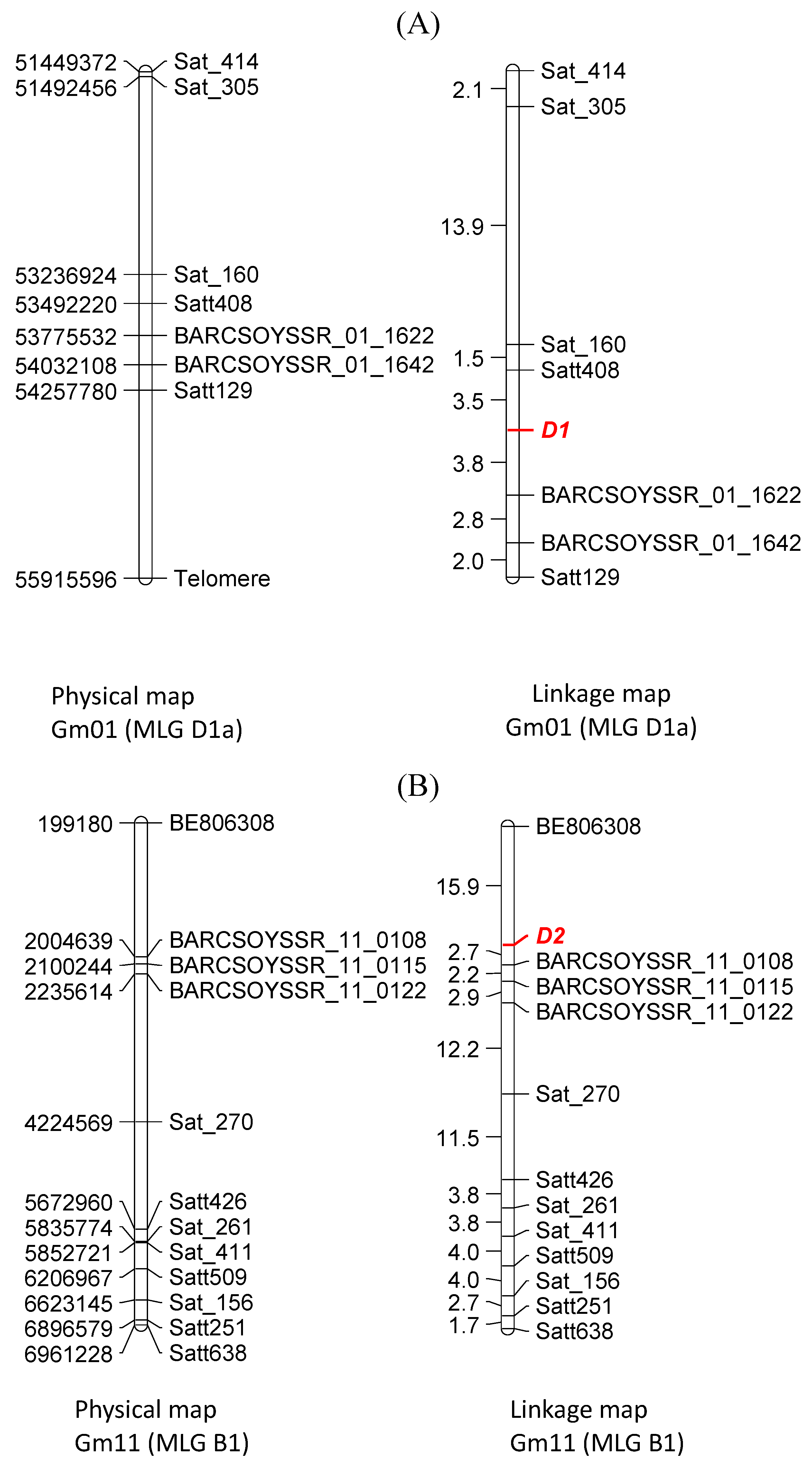
4. Discussion
4.1. Cytological Analyses
5. Conclusions
Acknowledgements
Abbreviations
| SSR | Simple Sequence Repeat |
| PCR | Polymerase Chain Reaction |
| BSA | Bulked Segregant Analysis |
| cM | centiMorgan |
| MLG | Molecular Linkage Group |
Conflict of Interest
References
- Kaul, M.L.H. Male Sterility in Higher Plants; Springer-Verlag: New York, NY, USA, 1988. [Google Scholar]
- Palmer, R.G.; Pfeiffer, T.W.; Buss, G.R.; Kilen, T.C. Qualitative genetics. In Soybeans: Improvement, Production, and Uses, Agronomy Monograph 16, 3rd ed.; American Society of Agrnomy, Inc.: Madison, WI, USA, 2004; pp. 137–233. [Google Scholar]
- Bai, Y.N.; Gai, J.Y. Development of a new cytoplasmic-nuclear male-sterility line of soybean and inheritance of its male-fertility restorability. Plant Breed. 2006, 125, 85–88. [Google Scholar] [CrossRef]
- Davis, W.H. Route to hybrid soybean production. US 4545146 A, 8 October 1985. [Google Scholar]
- Ding, D.; Gai, J.; Cui, Z.; Qiu, J. Development of a cytoplasmic-nuclear male-sterile line of soybean. Euphytica 2002, 124, 85–91. [Google Scholar] [CrossRef]
- Sun, H.; Zhao, L.; Huang, M. Cytoplasmic-Nuclear Male Sterile Soybean Line from Interspecific Crosses between G. max and G. soja. In Proceedings World Soybean Research Conference V: Soybean Feeds the World, Chiang Mai, Thailand, 1994; Chainuvat, C., Sarobol, N., Eds.; Kasetsart University Press: Chiang Mai, Thailand, 1997; pp. 99–102. [Google Scholar]
- Zhao, T.J.; Gai, J.Y. Discovery of new male-sterile cytoplasm sources and development of a new cytoplasmic-nuclear male-sterile line NJCMS3a in soybean. Euphytica 2006, 152, 387–396. [Google Scholar] [CrossRef]
- Horner, H.T.; Palmer, R.G. Mechanisms of genic male sterility. Crop Sci. 1995, 35, 1527–1535. [Google Scholar] [CrossRef]
- Rao, M.K.; Devi, K.U.; Arundhati, A. Applications of genic male sterility in plant breeding. Plant Breed. 1990, 105, 1–25. [Google Scholar] [CrossRef]
- Specht, J.E.; Graef, G.L. Registration of soybean germplasm SG1E6. Crop Sci. 1992, 32, 1080–1082. [Google Scholar] [CrossRef]
- Specht, J.E.; Williams, J.H.; Kenworthy, W.J.; Orf, J.H.; Helsel, D.G.; St. Martin, S.K. Registration of SG1 soybean germplasm. Crop Sci. 1985, 25, 717–718. [Google Scholar] [CrossRef]
- St. Martin, S.K.; Ehounou, N.E. Randomness of intermating in soybean populations containing male-sterile plants. Crop Sci. 1989, 29, 69–71. [Google Scholar] [CrossRef]
- Lewers, K.S.; Palmer, R.G. Recurrent selection in soybean. Plant Breed. Rev. 1997, 15, 275–314. [Google Scholar]
- Ortiz-Perez, E.; Wiley, H.; Horner, H.T.; Davis, W.H.; Palmer, R.G. Insect-mediated cross-pollination in soybean [Glycine max (L.) Merrill]: II. Phenotypic recurrent selection. Euphytica 2008, 162, 269–280. [Google Scholar]
- Jin, W.; Palmer, R.G.; Horner, H.T.; Shoemaker, R.C. Molecular mapping of a male-sterile gene in soybean. Crop Sci. 1998, 38, 1681–1685. [Google Scholar] [CrossRef]
- Cervantes-Martinez, I.; Xu, M.; Zhang, L.; Huang, Z.; Kato, K.K.; Horner, H.T.; Palmer, R.G. Molecular mapping of male-sterility loci ms2 and ms9 in soybean. Crop Sci. 2007, 47, 374–379. [Google Scholar] [CrossRef]
- Kato, K.K.; Palmer, R.G. Genetic identification of a female partial-sterile mutant in soybean. Genome 2003, 46, 128–134. [Google Scholar] [CrossRef]
- Frasch, R.M.; Weigand, C.; Perez, P.T.; Palmer, R.G.; Sandhu, D. Molecular mapping of 2 environmentally sensitive male-sterile mutants in soybean. J. Hered. 2010, 102, 11–16. [Google Scholar]
- Burton, J.W.; Brownie, C. Heterosis and inbreeding depression in two soybean single crosses. Crop Sci. 2006, 46, 2643–2648. [Google Scholar] [CrossRef]
- Perez, P.T.; Cianzio, S.R.; Ortiz-Perez, E.; Palmer, R.G. Agronomic performance of soybean hybrids from single, three-way, four-way, and five-way crosses, and backcross populations. J. Crop Improv. 2009, 23, 95–118. [Google Scholar] [CrossRef]
- Perez, P.T.; Cianzio, S.R.; Palmer, R.G. Evaluation of soybean [Glycine max (L.) Merr.] F1 hybrids. J. Crop Improv. 2009, 23, 1–18. [Google Scholar]
- Li, J.; Zhang, L.; Huang, Z.; Zhang, L.; Zhu, L. Yield heterosis and its utilization of F1 and F2 generations of CMS-type hybrid soybean. In International Conference on Utilization of Heterosis in Crops, Xi’an, China, 19–22 August 2012.
- Sun, H. Progress and problems of hybrid soybean development. In International Conference on Utilization of Heterosis in Crops, Xi’an, China, 19–22 August 2012.
- Yang, S.; Zhao, T.; Gai, J. Studies on the utilization of heterosis at the National Center for Soybean Improvement. In International Conference on Utilization of Heterosis in Crops, Xi’an, China, 19–22 August 2012.
- Zong, R.; Li, H.; Li, F.; Li, H.; Xu, M. Research progress on three-line hybrid soybean and prospects of soybean heterosis application. In International Conference on Utilization of Heterosis in Crops, Xi’an, China, 19–22 August 2012.
- Horner, H.T.; Healy, R.A.; Palmer, R.G. Floral nectary fine structure and development in Glycine max L. (Fabaceae). Int. J. Plant Sci. 2003, 164, 675–690. [Google Scholar]
- Palmer, R.G.; Perez, P.T.; Ortiz-Perez, E.; Maalouf, F.; Suso, M.J. The role of crop-pollinator relationship in breeding for pollinator-friendly legumes: From a breeding perspective. Euphytica 2009, 170, 35–52. [Google Scholar] [CrossRef]
- Buss, G.R. Inheritance of a male-sterile mutant from irradiated Essex soybeans. Soybean Genet. Newsl. 1983, 10, 104–108. [Google Scholar]
- Woodworth, C.M. Inheritance of cotyledon, seed-coat, hilum and pubescence colors in soy-beans. Genetics 1921, 6, 487. [Google Scholar]
- Lohnes, D.G.; Specht, J.E.; Cregan, P.B. Evidence for homoeologous linkage groups in soybean. Crop Sci. 1997, 37, 254–257. [Google Scholar] [CrossRef]
- Carter, T.E., Jr.; Burton, J.W. A Tight Linkage between the ms5 Male-Sterility Gene and the Green Cotyledon Trait in Soybean. In Agron. Abst.; American Society of Agronomy, Inc.: Madison, WI, USA, 1992; p. 91. [Google Scholar]
- Burton, J.W.; Carter, T.E.J. A method for production of experimental quantities of hybrid soybean seed. Crop Sci. 1983, 23, 388–390. [Google Scholar] [CrossRef]
- Cooper, R.L.; Tew, J. Use of leaf cutter bees in the production of hybrid soybean seed. In Annual Meeting Abstracts; [CD-ROM]; ASA, CSSA, and SSSA: Madison, WI, USA, 2001. [Google Scholar]
- Ruzin, S.E. Plant Microtechnique and Microscopy; Oxford University Press: New York, NY, USA, 1999. [Google Scholar]
- Horner, H.T.; Healy, R.A.; Ren, G.; Fritz, D.; Klyne, A.; Seames, C.; Thornburg, R.W. Amyloplast to chromoplast conversion in developing ornamental tobacco floral nectaries provides sugar for nectar and antioxidants for protection. Am. J. Bot. 2007, 94, 12–24. [Google Scholar] [CrossRef]
- Spurr, A.R. A low-viscosity epoxy resin embedding medium for electron microscopy. J. Ultrastruct. Res. 1969, 26, 31–34. [Google Scholar] [CrossRef]
- Fehr, W.R.; Caviness, C.E. Stages of soybean development. In Special Report 80; Iowa State University, Coop. Ext. Serv.; Iowa State University of Science and Technology: Ames, IA, USA, 1977. [Google Scholar]
- Sandhu, D.; Gao, H.; Cianzio, S.; Bhattacharyya, M.K. Deletion of a disease resistance nucleotide-binding-site leucine-rich-repeat-like sequence is associated with the loss of the Phytophthora resistance gene Rps4 in soybean. Genetics 2004, 168, 2157–2167. [Google Scholar] [CrossRef]
- Michelmore, R.W.; Paran, I.; Kesseli, R.V. Identification of markers linked to disease-resistance genes by bulked segregant analysis: a rapid method to detect markers in specific genomic regions by using segregating populations. Proc. Natl. Acad. Sci. USA 1991, 88, 9828–9832. [Google Scholar] [CrossRef]
- Song, Q.J.; Marek, L.F.; Shoemaker, R.C.; Lark, K.G.; Concibido, V.C.; Delannay, X.; Specht, J.E.; Cregan, P.B. A new integrated genetic linkage map of the soybean. Theor. Appl. Genet. 2004, 109, 122–128. [Google Scholar] [CrossRef]
- Grant, D.; Nelson, R.T.; Cannon, S.B.; Shoemaker, R.C. SoyBase, the USDA-ARS soybean genetics and genomics database. Nucleic Acids Res. 2010, 38, D843–D846. [Google Scholar] [CrossRef]
- Lander, E.S.; Green, P.; Abrahamson, J.; Barlow, A.; Daly, M.J.; Lincoln, S.E.; Newburg, L. MAPMAKER: An interactive computer package for constructing primary genetic linkage maps of experimental and natural populations. Genomics 1987, 1, 174–181. [Google Scholar] [CrossRef]
- Kosambi, D.D. The estimation of map distances from recombination values. Ann. Eugen. 1943, 12, 172–175. [Google Scholar] [CrossRef]
- Schmutz, J.; Cannon, S.B.; Schlueter, J.; Ma, J.; Mitros, T.; Nelson, W.; Hyten, D.L.; Song, Q.; Thelen, J.J.; Cheng, J.; et al. Genome sequence of the palaeopolyploid soybean. Nature 2010, 463, 178–183. [Google Scholar] [CrossRef]
- Palmer, R.G. A desynaptic mutant in the soybean. J. Hered. 1974, 65, 280–286. [Google Scholar]
- Guiamet, J.J.; Schwartz, E.; Pichersky, E.; Nooden, L.D. Characterization of cytoplasmic and nuclear mutations affecting chlorophyll and chlorophyll-binding proteins during senescence in soybean. Plant Physiol. 1991, 96, 227–231. [Google Scholar] [CrossRef]
- Chao, W.S.; Liu, V.; Thomson, W.W.; Platt, K.; Walling, L.L. The impact of chlorophyll-retention mutations, d1d2 and cyt-G1, during embryogeny in soybean. Plant Physiol. 1995, 107, 253–262. [Google Scholar]
- Eagles, H.A.; Bariana, H.S.; Ogbonnaya, F.C.; Rebetzke, G.J.; Hollamby, G.J.; Henry, R.J.; Henschke, P.H.; Carter, M. Implementation of markers in Australian wheat breeding. Crop Pasture Sci. 2001, 52, 1349–1356. [Google Scholar] [CrossRef]
- Rawat, N.; Neelam, K.; Tiwari, V.K.; Randhawa, G.S.; Friebe, B.; Gill, B.S.; Dhaliwal, H.S.; Somers, D. Development and molecular characterization of wheat, Aegilops kotschyi addition and substitution lines with high grain protein, iron, and zinc. Genome 2011, 54, 943–953. [Google Scholar] [CrossRef]
- Andersen, J.R.; Lubberstedt, T. Functional markers in plants. Trends Plant Sci. 2003, 8, 554–560. [Google Scholar] [CrossRef]
Supplementary Material
| Molecular Linkage Group | Chromosome Number | Number of markers |
|---|---|---|
| A1 | 5 | 35 |
| A2 | 8 | 35 |
| B1 | 11 | 25 |
| B2 | 14 | 28 |
| C1 | 4 | 20 |
| C2 | 6 | 23 |
| D1a | 1 | 46 |
| D1b | 2 | 54 |
| D2 | 17 | 21 |
| E | 15 | 23 |
| F | 13 | 39 |
| G | 18 | 45 |
| H | 12 | 22 |
| I | 20 | 28 |
| J | 16 | 20 |
| K | 9 | 25 |
| L | 19 | 24 |
| M | 7 | 37 |
| N | 3 | 25 |
| O | 10 | 25 |
| TOTAL | 600 |
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ott, A.; Yang, Y.; Bhattacharyya, M.; Horner, H.T.; Palmer, R.G.; Sandhu, D. Molecular Mapping of D1, D2 and ms5 Revealed Linkage between the Cotyledon Color Locus D2 and the Male-Sterile Locus ms5 in Soybean. Plants 2013, 2, 441-454. https://doi.org/10.3390/plants2030441
Ott A, Yang Y, Bhattacharyya M, Horner HT, Palmer RG, Sandhu D. Molecular Mapping of D1, D2 and ms5 Revealed Linkage between the Cotyledon Color Locus D2 and the Male-Sterile Locus ms5 in Soybean. Plants. 2013; 2(3):441-454. https://doi.org/10.3390/plants2030441
Chicago/Turabian StyleOtt, Alina, Yang Yang, Madan Bhattacharyya, Harry T. Horner, Reid G. Palmer, and Devinder Sandhu. 2013. "Molecular Mapping of D1, D2 and ms5 Revealed Linkage between the Cotyledon Color Locus D2 and the Male-Sterile Locus ms5 in Soybean" Plants 2, no. 3: 441-454. https://doi.org/10.3390/plants2030441