Homeotic Genes and the ABCDE Model for Floral Organ Formation in Wheat

Abstract
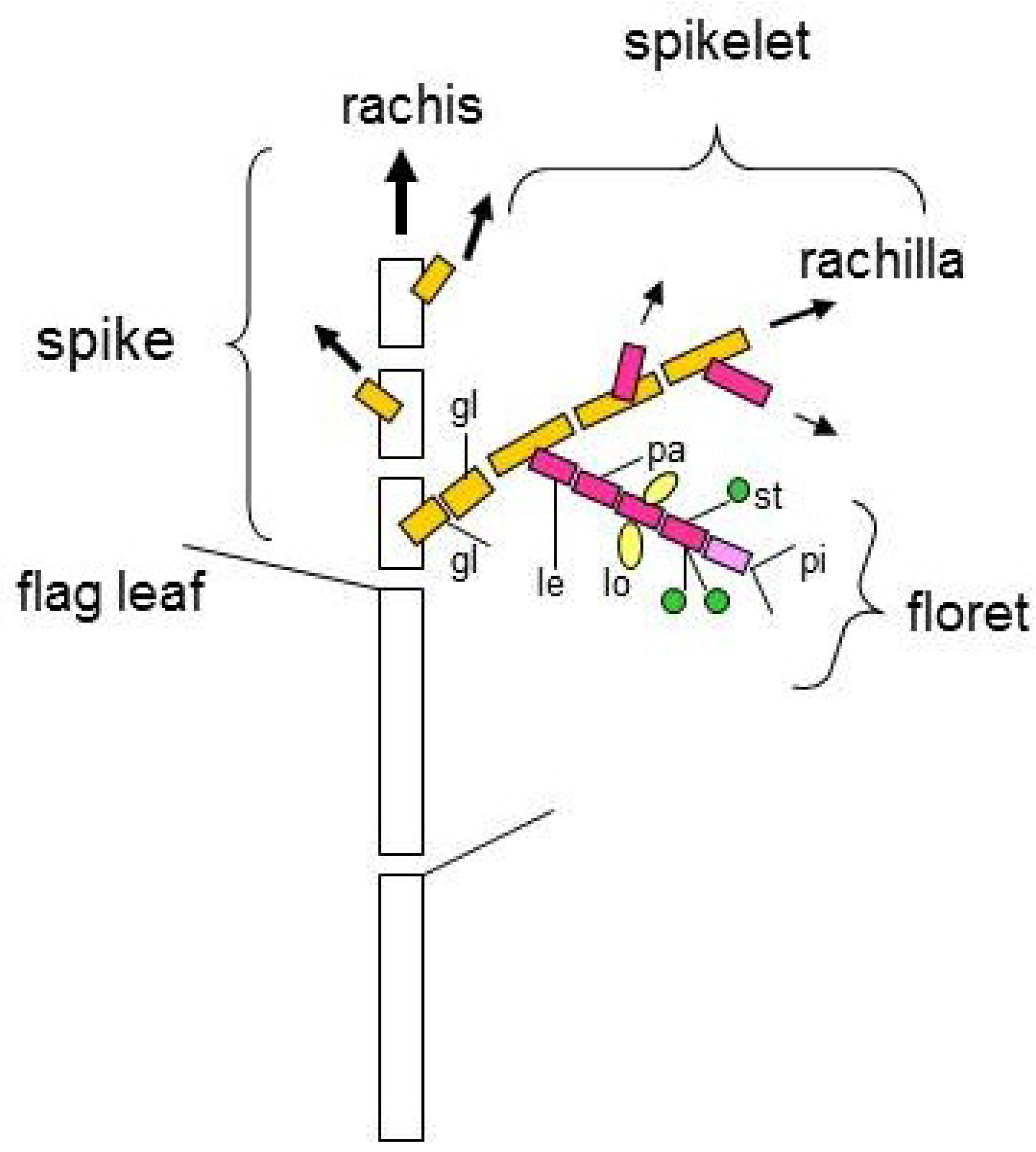
:1. Introduction


2. Summary of ABCDE Model in Rice
2.1. Rice Class A Genes
2.2. Rice Class B Genes
2.3. Rice Class C Genes
2.4. Rice Class D Genes
2.5. Rice Class E Genes
3. Pistillody, Homeotic Transformation of Stamens into Pistil-like Structures, in the Alloplasmic Wheat Line
4. Pistillody Reveals the Function of Class BCD MADS-Box Genes in Wheat
4.1. Wheat Class B Genes
4.2. Wheat Class C Genes
4.3. Wheat Class D Genes
4.4. Wheat DLOOPING LEAF Gene, TaDL
5. Other Homeotic Genes in Wheat
5.1. Wheat Class E Genes
5.2. Wheat Class A Genes
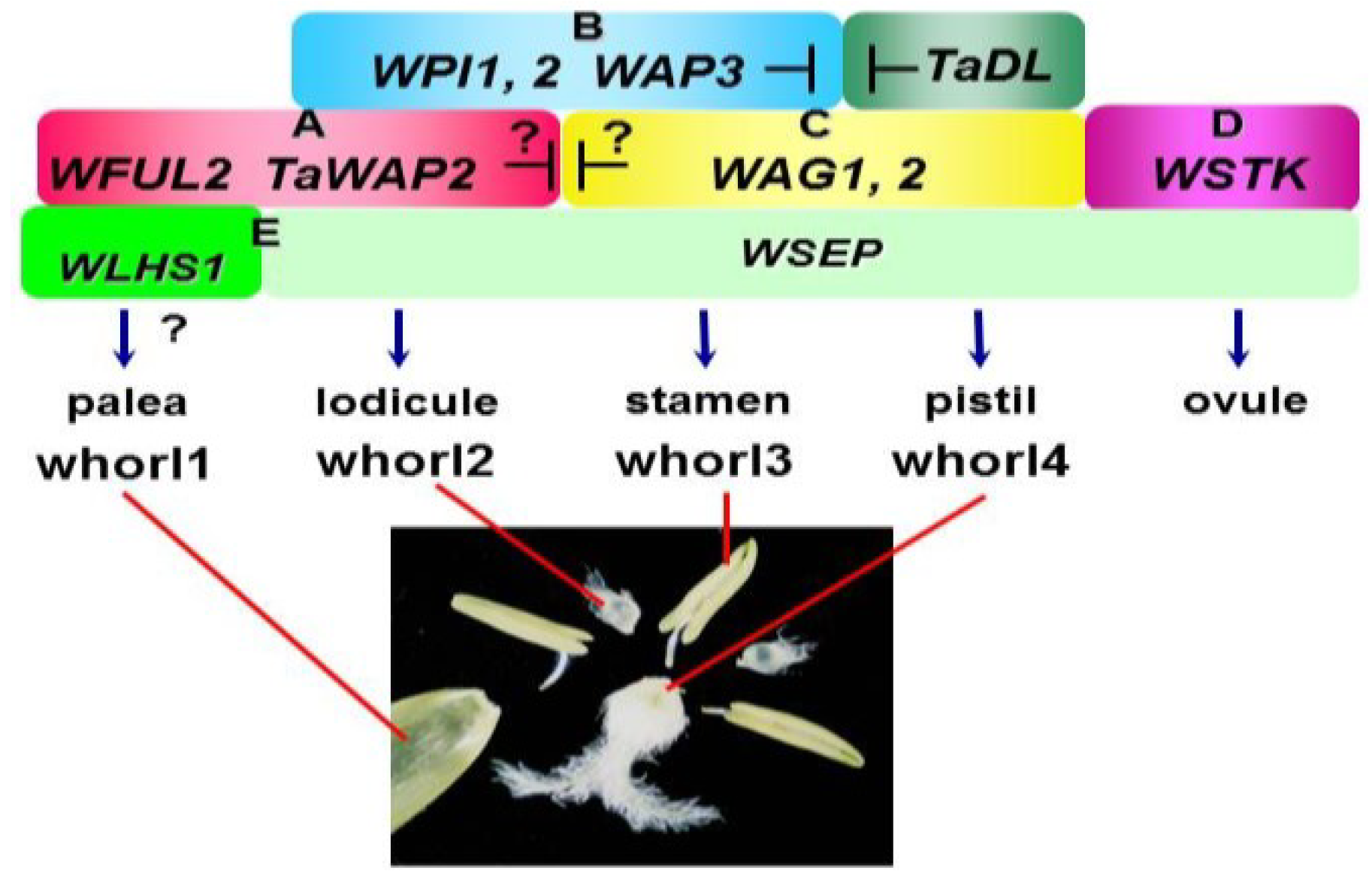
6. Wheat ABCDE Model, Complicated Homoeologous Gene Interaction


{kind=link}
{kind=link}
{kind=link}
| Class | Clade | Arabidopsis | Rice | Wheat |
|---|---|---|---|---|
| class A | AP1 | OsMADS14/RAP1B | WFUL1/VRN1 | |
| OsMADS15/RAP1A | WFUL2 | |||
| OsMADS18 | WFUL3 | |||
| AP2 | SNB | TaAP2 | ||
| OsIDS1 | Q | |||
| MFS1 | ||||
| class B | AP3 | OsMADS16/SPW1 | WAP3/TaAP3 * | |
| PI | OsMADS2 | WPI2/TaPI-2/TaAGL26 | ||
| OsMADS4 | WPI1/TaPI-1 | |||
| class C | AG | OsMADS3 | WAG2/TaAG-2/TaAGL39 | |
| OsMADS58 | WAG1/TaAG-1 | |||
| class D | STK | OsMADS13 | WSTK ** | |
| SHP1, 2 | ||||
| class E | SEP | SEP1, 2, 3, 4 | OsMADS7/OsMADS45 | WSEP |
| OsMADS8/OsMADS24 | TaMADS1 | |||
| LOFSEP | OsMADS1/LHS1 | WLHS1 | ||
| OsMADS5/OsM5 | ||||
| OsMADS34/PAP2 | ||||
| AGL6 | (AGL6) | OsMADS6/MFO1 | TaAGL6 *** | |
| OsMADS17 | ||||
| other | (CRC) | DL | TaDL |
Acknowledgments
Abbreviations
| AG | AGAMOUS |
| AGL | AGAMOUS LIKE |
| AP | APETALA |
| CS | Chinese Spring |
| DL | DROOPING LEAF |
| FUL | FRUITFULL |
| LHS | LEAFY HULL STERILE |
| MFO | MOSAIC FLORAL ORGAN |
| MFS | MULTI-FLORET SPIKELET |
| N26 | Norin 26 |
| OsIDS | Os INDETERMINATE SPIKELET |
| PAP | PANICLE PHYTOMER |
| PI | PISTILLATA |
| SHP | SHATTERPROOF |
| SEP | SEPALLATA |
| SNB | SUPERNUMERARY BRACT |
| SPW | SUPERWOMAN |
| STK | SEEDSTICK |
| VRN | VERNARIZATION |
Conflict of Interest
References
- Rijpkema, A.S.; Vandenbussche, M.; Koes, R.; Heijmans, K.; Gerats, T. Variations on a theme: Changes in the floral ABCs in angiosperms. Semin. Cell Dev. Biol. 2010, 21, 100–107. [Google Scholar] [CrossRef]
- Smaczniak, C.; Immink, R.G.H.; Angenent, G.C.; Kaufmann, K. Developmental and evolutionary diversity of plant MADS-domain factors: Insights from recent studies. Development 2012, 139, 3081–3098. [Google Scholar] [CrossRef]
- Jofuku, K.D.; den Boer, B.G.; Montagu, M.V.; Okamuro, J.K. Control of Arabidopsis flower and seed development by the homeotic gene APETALA2. Plant Cell 1994, 6, 1211–1225. [Google Scholar]
- Mandel, M.A.; Gustafson-Brown, C.; Savidge, B.; Yanofsky, M.F. Molecular characterization of the Arabidopsis floral homeotic gene APETALA1. Nature 1992, 360, 273–277. [Google Scholar] [CrossRef]
- Jack, T.; Brockman, L.L.; Meyerowitz, E.M. The homeotic gene APETALA3 of Arabidopsis thaliana encodes a MADS box and is expressed in petals and stamens. Cell 1992, 68, 683–697. [Google Scholar] [CrossRef]
- Goto, K.; Meyerowitz, E.M. Function and regulation of the Arabidopsis floral homeotic gene PISTILLATA. Genes Dev. 1994, 8, 1548–1560. [Google Scholar] [CrossRef]
- Yanofsky, M.F.; Ma, H.; Bowman, J.L.; Drews, G.N.; Feldmann, K.A.; Meyerowitz, E.M. The protein encoded by the Arabidopsis homeotic gene agamous resembles transcription factors. Nature 1990, 346, 35–39. [Google Scholar] [CrossRef]
- Favaro, R.; Pinyopich, A.; Battaglia, R.; Kooiker, M.; Borghi, L.; Ditta, G.; Yanofsky, M.F.; Kater, M.M.; Colombo, L. MADS-box protein complexes control carpel and ovule development in Arabidopsis. Plant Cell 2003, 15, 2603–2611. [Google Scholar] [CrossRef]
- Pinyopich, A.; Ditta, G.S.; Savidge, B.; Liljegren, S.J.; Baumann, E.; Wisman, E.; Yanofsky, M.F. Assessing the redundancy of MADS-box genes during carpel and ovule development. Nature 2003, 424, 85–88. [Google Scholar]
- Pelaz, S.; Ditta, G.S.; Baumann, E.; Wisman, E.; Yanofsky, M.F. B and C floral organ identity functions require SEPSLLATA MADS-box genes. Nature 2000, 405, 200–203. [Google Scholar] [CrossRef]
- Ditta, G.; Pinyopich, A.; Robles, P.; Pelaz, S.; Yanofsky, M.F. The SEP4 gene of Arabidopsis thaliana functions in floral organ and meristem identity. Curr. Biol. 2004, 14, 1935–1940. [Google Scholar] [CrossRef]
- Litt, A.; Kramer, E.M. The ABC model and the diversification of floral organ identity. Semin. Cell Dev. Biol. 2010, 21, 129–137. [Google Scholar] [CrossRef]
- Schauer, S.E.; Schluter, P.M.; Baskar, R.; Gheyselinck, J.; Bolanos, A.; Curtis, M.D.; Grossniklaus, U. Intronic regulatory elements determine the divergent expression patterns of AGAMOUS-LIKE6 subfamily members in Arabidopsis. Plant J. 2009, 59, 987–1000. [Google Scholar] [CrossRef]
- Schmidt, R.J.; Ambrose, B.A. The blooming of grass flower development. Curr. Opin. Plant Biol. 1998, 1, 60–67. [Google Scholar] [CrossRef]
- Bommert, P.; Satoh-Nagasawa, N.; Jackson, D.; Hirano, H.-Y. Genetics and evolution of inflorescence and flower development in grasses. Plant Cell Physiol. 2005, 46, 69–78. [Google Scholar] [CrossRef]
- Murai, K.; Takumi, S.; Koga, H.; Ogihara, Y. Pistillody, homeotic transformation of stamens into pistil-like structures, caused by nuclear-cytoplasm interaction in wheat. Plant J. 2002, 29, 169–181. [Google Scholar] [CrossRef]
- Shitsukawa, N.; Takagishi, A.; Ikari, C.; Takumi, S.; Murai, K. WFL, a wheat FLORICAULA/LEAFY ortholog, is associated with spikelet formation as lateral branch of the inflorescence meristem. Genes Genet. Syst. 2006, 81, 13–20. [Google Scholar] [CrossRef]
- Ciaffi, M.; Paolacci, A.R.; Tanzarella, O.A.; Porceddu, E. Molecular aspects of flower development in grasses. Sex. Plant Reprod. 2011, 24, 247–282. [Google Scholar] [CrossRef]
- Kellogg, E.A. Evolutionary history of the grasses. Plant Physiol. 2001, 125, 1198–1205. [Google Scholar] [CrossRef]
- Yoshida, H. Is the lodicule a petal: Molecular evidence? Plant Sci. 2012, 184, 121–128. [Google Scholar]
- Yoshida, H.; Nagato, Y. Flower development in rice. J. Exp. Bot. 2011, 62, 4719–4730. [Google Scholar] [CrossRef]
- Litt, A.; Irish, V.F. Duplication and diversification in the APETALA1/FRUITFULL floral homeotic gene lineage: Implications for the evolution of floral development. Genetics 2003, 165, 821–833. [Google Scholar]
- Jeon, J.-S.; Lee, S.; Jung, K.-H.; Yang, W.-S.; Yi, G.-H.; Oh, B.-G.; An, G. Production of transgenic rice plants showing reduced heading date and plant height by ectopic expression of rice MADS-box genes. Mol. Breed. 2000, 6, 581–592. [Google Scholar] [CrossRef]
- Wang, K.; Tang, D.; Hong, L.; Xu, W.; Huang, J.; Li, M.; Gu, M.; Xue, Y.; Cheng, Z. DEP and AFO regulate reproductive habit in rice. PLoS Genet. 2010, 6, e1000818. [Google Scholar] [CrossRef]
- Zhu, Q.-H.; Upadhyaya, N.M.; Gubler, F.; Helliwell, C.A. Over-expression of miR172 causes loss of spikelet determinacy and floral organ abnormalities in rice (Oryza sativa). BMC Plant Biol. 2009, 9, 149. [Google Scholar] [CrossRef]
- Lee, D.-Y.; An, G. Two AP2 family genes, SUPERNUMERARY BRACT (SNB) and OsINDETERMINATE SPIKELET 1 (OsIDS1), synergistically control inflorescence architecture and floral meristem establishment in rice. Plant J. 2012, 69, 445–461. [Google Scholar] [CrossRef]
- Ren, D.; Li, Y.; Zhao, F.; Sang, X.; Shi, J.; Wang, N.; Guo, S.; Ling, Y.; Zhang, C.; Yang, Z.; et al. MULTI-FLORET SPIKELET1, which encodes and AP2/ERF protein, determines spikelet meristem gate and sterile lemma identity in rice. Plant Physiol. 2013. [Google Scholar] [CrossRef]
- Chung, Y.-Y.; Kim, S.-R.; Kang, H.-G.; Noh, Y.-S.; Park, M.C.; Finkel, D.; An, G. Characterization of two rice MADS box genes homologous to GLOBOSA. Plant Sci. 1995, 109, 45–56. [Google Scholar] [CrossRef]
- Prasad, K.; Vijayraghavan, U. Double-stranded RNA interference of a rice PI/GLO paralog, OsMADS2, uncovers its second-whorl-specific function in floral organ patterning. Genetics 2003, 165, 2301–2305. [Google Scholar]
- Yao, S.-G.; Ohmori, S.; Kimizu, M.; Yoshida, H. Unequal genetic redundancy of rice PISTILLATA orthologs, OsMADS2 and OsMADS4, in lodicule and stamen development. Plant Cell Physiol. 2008, 49, 853–857. [Google Scholar] [CrossRef]
- Nagasawa, N.; Miyoshi, M.; Sano, Y.; Satoh, H.; Hirano, H.-Y.; Sakai, H.; Nagato, Y. SUPERWOMAN1 and DROOPING LEAF genes control floral organ identity in rice. Development 2003, 130, 705–718. [Google Scholar] [CrossRef]
- Yoshida, H.; Itoh, J.-I.; Ohmori, S.; Miyoshi, K.; Horigome, A.; Uchida, E.; Kimizu, M.; Matsumura, Y.; Kusaba, M.; Satoh, H.; et al. Superwoman1-cleistogamy, a hopeful allele for gene containment in GM rice. Plant Biotechnol. J. 2007, 5, 835–846. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Lee, D.Y.; Miyao, A.; Hirochika, H.; An, G.; Hirano, H.-Y. Functional diversification of the two C-class MADS box genes OSMADS3 and OSMADS58 in Oryza sativa. Plant Cell 2006, 18, 15–28. [Google Scholar] [CrossRef]
- Dreni, L.; Pilatone, A.; Yun, D.; Erreni, S.; Pajoro, A.; Caporali, E.; Zhang, D.; Kater, M.M. Functional analysis of all AGAMOUS subfamily members in rice reveals their roles in reproductive organ identity determination and meristem determinacy. Plant Cell 2011, 23, 2850–2863. [Google Scholar] [CrossRef]
- Yun, D.; Liang, W.; Dreni, L.; Yin, C.; Zhou, Z.; Kater, M.M.; Zhang, D. OsMADS16 interacts with OsMADS3 and OsMADS58 in specifying floral patterning in rice. Mol. Plant 2013. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Nagasawa, N.; Kawasaki, S.; Matsuoka, M.; Nagato, Y.; Hirano, H.-Y. The YABBY gene DROOPING LEAF regulates carpel specification and midrib development in Oryza sativa. Plant Cell 2004, 16, 500–509. [Google Scholar] [CrossRef]
- Lopez-Dee, Z.P.; Wittich, P.; Pé, M.E.; Rigora, D.; del Buono, I.; Sari Gorla, M.; Kater, M.M.; Colombo, L. OsMADS13, a novel rice MADS-box gene expressed during ovule development. Dev. Genet. 1999, 25, 237–244. [Google Scholar] [CrossRef]
- Favaro, R.; Immink, R.G.H.; Ferioli, V.; Bernasconi, B.; Byzova, M.; Angenent, G.C.; Kater, M.M.; Colombo, L. Ovule-specific MADS-box proteins have conserved protein-protein interactions in monocot and dicot plants. Mol. Genet. Genomics 2002, 268, 152–159. [Google Scholar] [CrossRef]
- Dreni, L.; Jacchia, S.; Fornara, F.; Fornari, M.; Ouwerkerk, P.B.E.; An, G.; Colombo, L.; Kater, M.M. The D-lineage MADS-box gene OsMADS13 controls ovule identity in rice. Plant J. 2007, 52, 690–699. [Google Scholar] [CrossRef]
- Yamaki, S.; Nagato, Y.; Kurata, N.; Nonomura, K. Ovule is a lateral organ finally differentiated from the terminating floral meristem in rice. Dev. Biol. 2011, 351, 208–216. [Google Scholar]
- Malcomber, S.T.; Kellogg, E.A. SEPALLATA gene diversification: Brave new whorls. Trends Plant Sci. 2005, 10, 427–435. [Google Scholar] [CrossRef]
- Cui, R.; Han, J.; Zhao, S.; Su, K.; Wu, F.; Du, X.; Xu, Q.; Chong, K.; Theissen, G.; Meng, Z. Functional conservation and diversification of class E floral homeotic genes in rice (Oryza sativa). Plant J. 2010, 61, 767–781. [Google Scholar] [CrossRef]
- Christensen, A.R.; Malcomber, S.T. Duplication and diversification of the LEAFY HULL STERILE1 and Oryza sativa MADS5 SEPALLATA lineages in graminoid Poales. EvoDevo 2012, 3, 4. [Google Scholar] [CrossRef]
- Jeon, J.-S.; Jang, S.; Lee, S.; Nam, J.; Kim, C.; Lee, S.-H.; Chung, Y.-Y.; Kim, S.-R.; Lee, Y.H.; Cho, Y.-G.; et al. Leafy hull sterile1 is a homeotic mutation in a rice MADS box gene affecting rice flower development. Plant Cell 2000, 12, 871–884. [Google Scholar]
- Prasad, K.; Parameswaran, S.; Vijayraghavan, U. OsMADS1, a rice MADS-box factor, controls differentiation of specific cell types in the lemma and palea and is an early-acting regulator of inner floral organs. Plant J. 2005, 43, 915–928. [Google Scholar] [CrossRef]
- Gao, X.; Liang, W.; Yin, C.; Ji, S.; Wang, H.; Su, X.; Guo, C.; Kong, H.; Xue, H.; Zhang, D. The SEPALLATA-like gene OsMADS34 is required for rice inflorescence and spikelet development. Plant Physiol. 2010, 153, 728–740. [Google Scholar] [CrossRef]
- Kobayashi, K.; Yasuno, N.; Sato, Y.; Yoda, M.; Yamazaki, R.; Kimizu, M.; Yoshida, H.; Nagamura, Y.; Kyozuka, J. Inflorescence meristem identity in rice is specified by overlapping functions of three AP1/FUL-like MADS box genes and PAP2, a SEPALLATA MADS box gene. Plant Cell 2012, 24, 1848–1859. [Google Scholar] [CrossRef]
- Ohmori, S.; Kimizu, M.; Sugita, M.; Miyao, A.; Hirochika, H.; Uchida, E.; Nagato, Y.; Yoshida, H. MOSAIC FLORAL ORGANS1, an AGL6-like MADS box gene, regulates floral organ identity and meristem fate in rice. Plant Cell 2009, 21, 3008–3025. [Google Scholar] [CrossRef]
- Li, H.; Liang, W.; Jia, R.; Yin, C.; Zong, J.; Kong, H.; Zhang, D. The AGL6-like gene OsMADS6 regulates floral organ and meristem identities in rice. Cell Res. 2010, 20, 299–313. [Google Scholar] [CrossRef]
- Li, H.; Liang, W.; Hu, Y.; Zhu, L.; Yin, C.; Xu, J.; Dreni, L.; Kater, M.M.; Zhang, D. Rice MADS6 interacts with the floral homeotic genes SUPERWOMAN1, MADS3, MADS58, MADS13, and DROOPING LEAF in specifying floral organ identities and meristem fate. Plant Cell 2011, 23, 2536–2552. [Google Scholar] [CrossRef]
- Duan, Y.; Xing, Z.; Diao, Z.; Xu, W.; Li, S.; Du, X.; Wu, G.; Wang, C.; Lan, T.; Meng, Z.; et al. Characterization of Osmads6–5, a null allele, reveals that OsMADS6 is acritical regulator for early flower development in rice (Oryza sativa L.). Plant Mol. Biol. 2012, 80, 429–442. [Google Scholar]
- Tsunewaki, K.; Wang, G.-Z.; Matsuoka, Y. Plasmon analysis of Triticum (wheat) and Aegilops. 1. Production of alloplasmic common wheats and their fertilities. Genes Genet. Syst. 1996, 71, 293–311. [Google Scholar] [CrossRef]
- Tsunewaki, K.; Wang, G.-Z.; Matsuoka, Y. Plasmon analysis of Triticum (wheat) and Aegilops. 2. Characterization and classification of 47 plasmons based on their effects on common wheat phenotype. Genes Genet. Syst. 2002, 77, 409–427. [Google Scholar]
- Murai, K.; Tsunewaki, K. Photoperiod-sensitive cytoplasmic male sterility in wheat with Aegilops crassa cytoplasm. Euphytica 1993, 67, 41–48. [Google Scholar]
- Murai, K. Genetic effects of an alien cytoplasm on male and female fertility in wheat. Recent Res. Dev. Genet. 2001, 1, 47–54. [Google Scholar]
- Murai, K.; Tsunewaki, K. Genetic analysis on the fertility restration by Triticum aestivum cv. Chinese Spring against photoperiod-sensitive cytoplasmic male sterility. Jpn. J. Genet.Genes Genet. Syst. 1994, 69, 195–202. [Google Scholar]
- Murai, K.; Takumi, S.; Koga, H.; Ogihara, Y. Pistillody, homeotic transformation of stamens into pistil-like structures, caused by nuclear-cytoplasm interaction in wheat. Plant J. 2002, 29, 169–182. [Google Scholar] [CrossRef]
- Zhu, Y.; Saraike, T.; Yamamoto, Y.; Hagita, H.; Takumi, S.; Murai, K. orf260cra, a novel mitochondrial gene, is associated with the homeotic transformation of stamen into pistil-like structures (pistillody) in alloplasmic wheat. Plant Cell Physiol. 2008, 49, 1723–1733. [Google Scholar] [CrossRef]
- Saraike, T.; Shitsukawa, N.; Yamamoto, Y.; Hagita, H.; Iwasaki, Y.; Takumi, S.; Murai, K. Identification of a protein kinase gene associated with pistillody, homeotic transformation of stamens into pistil-like structures, in alloplasmic wheat. Planta 2007, 227, 211–221. [Google Scholar] [CrossRef]
- Yamamoto, M.; Shitsukawa, N.; Yamada, M.; Kato, K.; Takumi, S.; Kawaura, K.; Ogihara, Y.; Murai, K. Identification of a novel homolog for a calmodulin-binding protein that is upregulated in alloplasmic wheat showing pistillody. Planta 2013, 237, 1001–1013. [Google Scholar] [CrossRef]
- Murai, K.; Murai, R.; Takumi, S.; Ogihara, Y. Cloning and Characterization of cDNAs Corresponding to the Wheat MADS Box Genes. In Proceedings of 9th International Wheat Genetics Symposium, Saskatchewan, Canada, 2–7 August 1998; Volume 1, pp. 89–94.
- Feldman, M. The Origin of Cultivated Wheat. In The World Wheat Book; Bonjean, A.P., Angus, W.J., Eds.; Lavoisier Publisher: Paris, France, 2001; pp. 1–56. [Google Scholar]
- Paolacci, A.R.; Tanzarella, O.A.; Porceddu, E.; Varotto, S.; Ciaffi, M. Molecular and phylogenetic analysis of MADS-box genes of MIKC type and chromosome location of SEP-like gene in wheat (Triticum aestivum L.). Mol. Genet. Genomics 2007, 278, 689–708. [Google Scholar] [CrossRef]
- Hama, E.; Takumi, S.; Ogihara, Y.; Murai, K. Pistillody is caused by alterations to the class-B MADS-box gene expression pattern in alloplasmic wheats. Planta 2004, 218, 712–720. [Google Scholar] [CrossRef]
- Zhao, T.; Ni, Z.; Dai, Y.; Yao, Y.; Nie, X.; Sun, Q. Characterization and expression of 42 MADS-box genes in wheat (Triticum aestivum L.). Mol. Genet. Genomics 2006, 276, 334–350. [Google Scholar]
- Meguro, A.; Takumi, S.; Ogihara, Y.; Murai, K. WAG, a wheat AGAMOUS homolog, is associated with development of pistil-like stamens in alloplasmic wheats. Sex. Plant Reprod. 2003, 15, 221–230. [Google Scholar]
- Hirabayashi, C.; Murai, K. Class C MADS-box gene AGAMOUS was duplicated in the wheat genome. Wheat Inf. Serv. 2009, 107, 13–16. [Google Scholar]
- Yamada, K.; Saraike, T.; Shitsukawa, N.; Hirabayashi, C.; Takumi, S.; Murai, K. Class D and Bsister MADS-box genes are associated with ectopic ovule formation in the pistil-like stamens of alloplasmic wheat (Triticum aestivum L.). Plant Mol. Biol. 2009, 71, 1–14. [Google Scholar] [CrossRef]
- Wei, S.; Peng, Z.; Zhou, Y.; Yang, Z.; Wu, K.; Ouyang, Z. Nucloetide diversity and molecular evolution of the WAG-2 gene in common wheat (Triticum aestivum L.) and its relatives. Genet. Mol. Biol. 2011, 34, 606–615. [Google Scholar]
- Ishikawa, M.; Ohmori, Y.; Tanaka, W.; Hirabayashi, C.; Murai, K.; Ogihara, Y.; Yamaguchi, T.; Hirano, H.-Y. The spatial expression patterns of DROOPING LEAF orthologs suggest a conserved function in grasses. Genes Genet. Syst. 2009, 84, 137–146. [Google Scholar] [CrossRef]
- Shitsukawa, N.; Tahira, C.; Kassai, K.-I.; Hirabayashi, C.; Shimizu, T.; Takumi, S.; Mochida, K.; Kawaura, K.; Ogihara, Y.; Murai, K. Genetic and epigenetic alteration among three homoeologous genes of a class E MADS box gene in hexaploid wheat. Plant Cell 2007, 19, 1723–1737. [Google Scholar] [CrossRef]
- Reinheimer, R.; Kellogg, E.A. Evolution of AGL6-like MADS box genes in grasses (Poaceae): Ovule expression is ancient and palea expression is new. Plant Cell 2009, 21, 2591–2605. [Google Scholar] [CrossRef]
- Smaczniak, C.; Immink, R.G.H.; Muino, J.M.; Blanvillain, R.; Busscher, M.; Busscher-Lange, J.; Dinh, Q.D.P.; Liu, S.; Westphal, A.H.; Boeren, S.; et al. Characterization of MADS-domain transcription factor complexes in Arabidopsis flower development. Proc. Natl. Acad. Sci. USA 2012, 109, 1560–1565. [Google Scholar]
- Zhao, X.Y.; Cheng, Z.J.; Zhang, X.S. Overexpression of TaMADS1, a SEPALLATA-like gene in wheat, causes early flowering and the abnormal development of floral organs in Arabidopsis. Planta 2006, 223, 698–707. [Google Scholar] [CrossRef]
- Malcomber, S.T.; Kellogg, E.A. Heterogeneous expression patterns and separate roles of the SEPALLATA gene LEAFY HULL STERILE1 in grasses. Plant Cell 2004, 16, 1692–1706. [Google Scholar] [CrossRef]
- Ferrandiz, C.; Gu, Q.; Martienssen, R.; Yanofsky, M.F. Redundant regulation of meristem identity and plant architecture by FRUITFULL, APETALA1 and CAULIFLOWER. Development 2000, 127, 725–734. [Google Scholar]
- Yan, L.; Loukoianov, A.; Tranquilli, G.; Helguera, M.; Fahima, T.; Dubcovsky, J. Positional cloning of the wheat vernalization gene VRN1. Proc. Natl. Acad. Sci. USA 2003, 100, 6263–6268. [Google Scholar] [CrossRef]
- Murai, K.; Miyamae, M.; Kato, H.; Takumi, S.; Ogihara, Y. WAP1, a wheat APETALA1 homolog, plays a central role in the phase transition from vegetative to reproductive growth. Plant Cell Physiol. 2003, 44, 1255–1265. [Google Scholar] [CrossRef]
- Trevaskis, B.; Bagnall, D.J.; Ellis, M.H.; Peacock, W.J.; Dennis, E.S. MADS box genes control vernalization-induced flowering in cereals. Proc. Natl. Acad. Sci. USA 2003, 100, 13099–13104. [Google Scholar]
- Danyluk, J.; Kane, N.A.; Breton, G.; Limin, A.E.; Fowler, D.B.; Sarhan, F. TaVRT-1, a putative transcription factor associated with vegetative to reproductive transition in cereals. Plant Physiol. 2003, 132, 1849–1860. [Google Scholar] [CrossRef]
- Kinjo, H.; Shitsukawa, N.; Takumi, S.; Murai, K. Diversification of three APETALA1/FRUITFULL-like genes in wheat. Mol. Genet. Genomics 2012, 287, 283–294. [Google Scholar] [CrossRef]
- Lim, J.; Moon, Y.-H.; An, G.; Jang, S.K. Two rice MADS domain proteins interact with OsMADS1. Plant Mol. Biol. 2000, 44, 513–527. [Google Scholar] [CrossRef]
- Zhang, Z.; Belcram, H.; Gornicki, P.; Charles, M.; Just, J.; Huneau, C.; Magdelenat, G.; Couloux, A.; Samain, S.; Gill, B.S.; et al. Duplication and partitioning in evolution and function of homoeologous Q loci governing domestication characters in polyploidy wheat. Proc. Natl. Acad. Sci. USA 2011, 108, 18737–18742. [Google Scholar] [CrossRef]
- Ning, S.; Wang, N.; Sakuma, S.; Pourkheirandish, M.; Wu, J.; Matsumoto, T.; Koba, T.; Komatsuda, T. Structure, transcription and post-transcriptional regulation of the bread wheat orthologs of the barley cleistogamy gene Cly1. Theor. Appl. Genet. 2013. [Google Scholar] [CrossRef]
- Nair, S.K.; Wang, N.; Turuspekov, Y.; Pourkheirandish, M.; Sinsuwongwat, S.; Chen, G.; Sameri, M.; Tagiri, A.; Honda, I.; Watanabe, Y.; et al. Cleistogamous flowering in barley arises from the suppression of microRNA-guided HvAP2 mRNA cleavage. Proc. Natl. Acad. Sci. USA 2010, 107, 490–495. [Google Scholar] [CrossRef] [Green Version]
- Wendel, J.F. Genome evolution in polyploids. Plant Mol. Biol. 2000, 42, 225–249. [Google Scholar] [CrossRef]
- Te Beest, M.; Le Roux, J.J.; Richardson, D.M.; Brysting, A.K.; Suda, J.; Kubesova, M.; Pysek, P. The more the better? The role of polyploidy in facilitating plant invasions. Ann. Bot. 2012, 109, 19–45. [Google Scholar] [CrossRef]
- Shitsukawa, N.; Kinjo, H.; Takumi, S.; Murai, K. Heterochronic development of the floret meristem determines grain number per spikelet in diploid, tetraploid and hexaploid wheats. Ann. Bot. 2009, 104, 243–251. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Murai, K. Homeotic Genes and the ABCDE Model for Floral Organ Formation in Wheat. Plants 2013, 2, 379-395. https://doi.org/10.3390/plants2030379
Murai K. Homeotic Genes and the ABCDE Model for Floral Organ Formation in Wheat. Plants. 2013; 2(3):379-395. https://doi.org/10.3390/plants2030379
Chicago/Turabian StyleMurai, Koji. 2013. "Homeotic Genes and the ABCDE Model for Floral Organ Formation in Wheat" Plants 2, no. 3: 379-395. https://doi.org/10.3390/plants2030379