Regulation of Pollen Tube Growth by Transglutaminase


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. The Pollen and Its Germination
2. Structure and Function of TGases
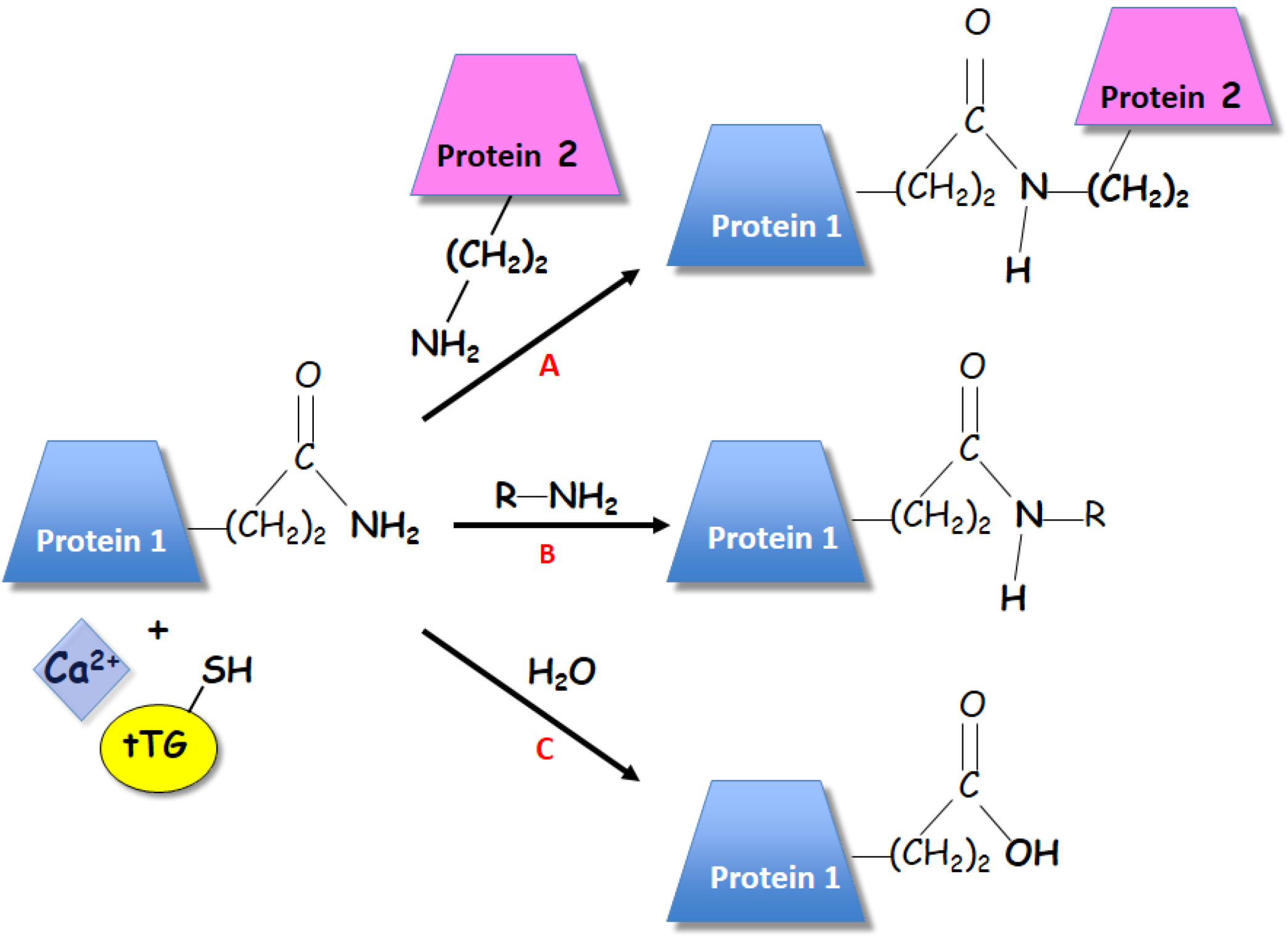
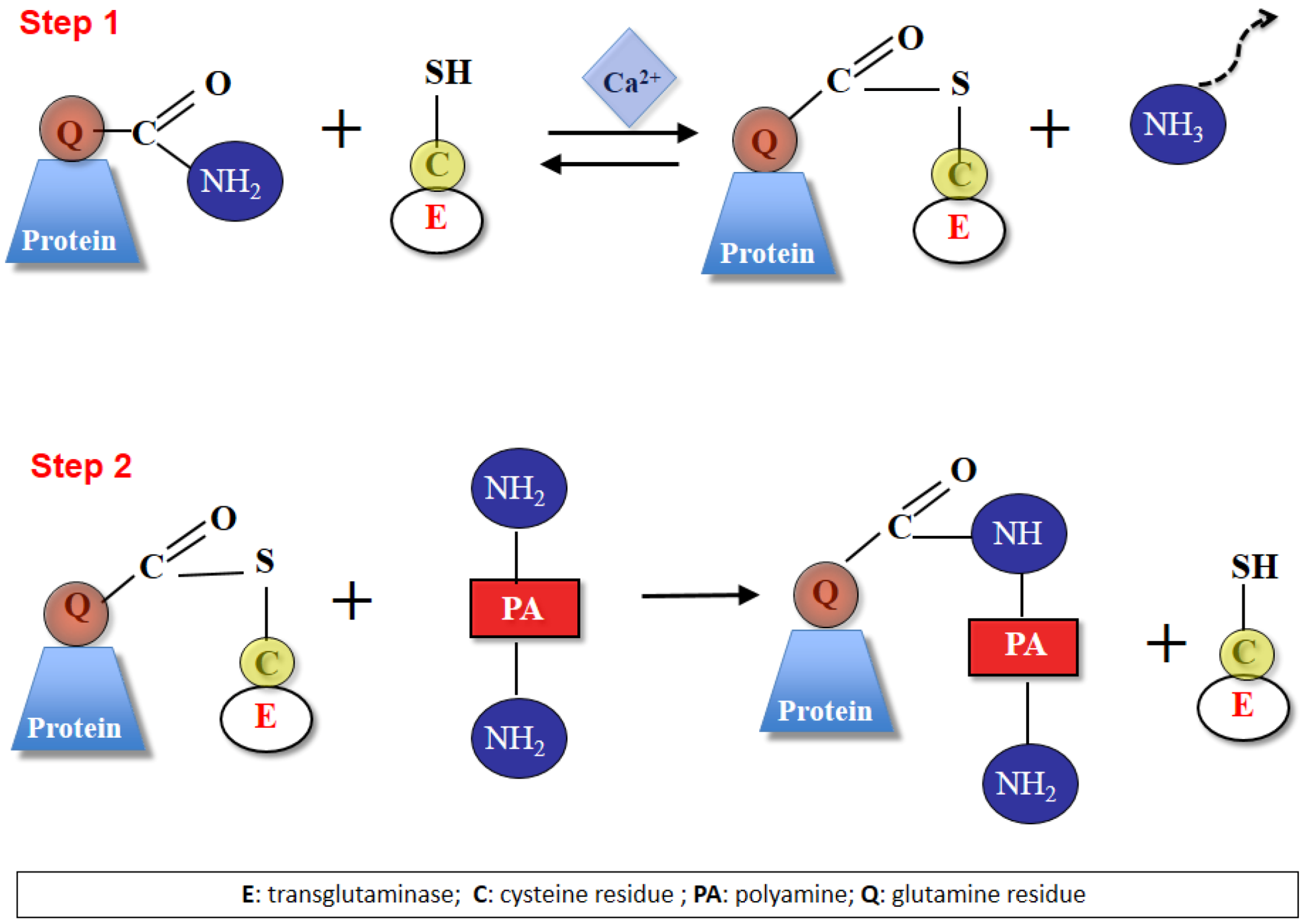
3. Polyamines and Transglutaminases in Plants
4. Transglutaminase in Pollen
4.1. Intracellular Targets of TGase Activity: The Cytoskeleton
4.2. Secretion and Targeting of Extracellular TGase
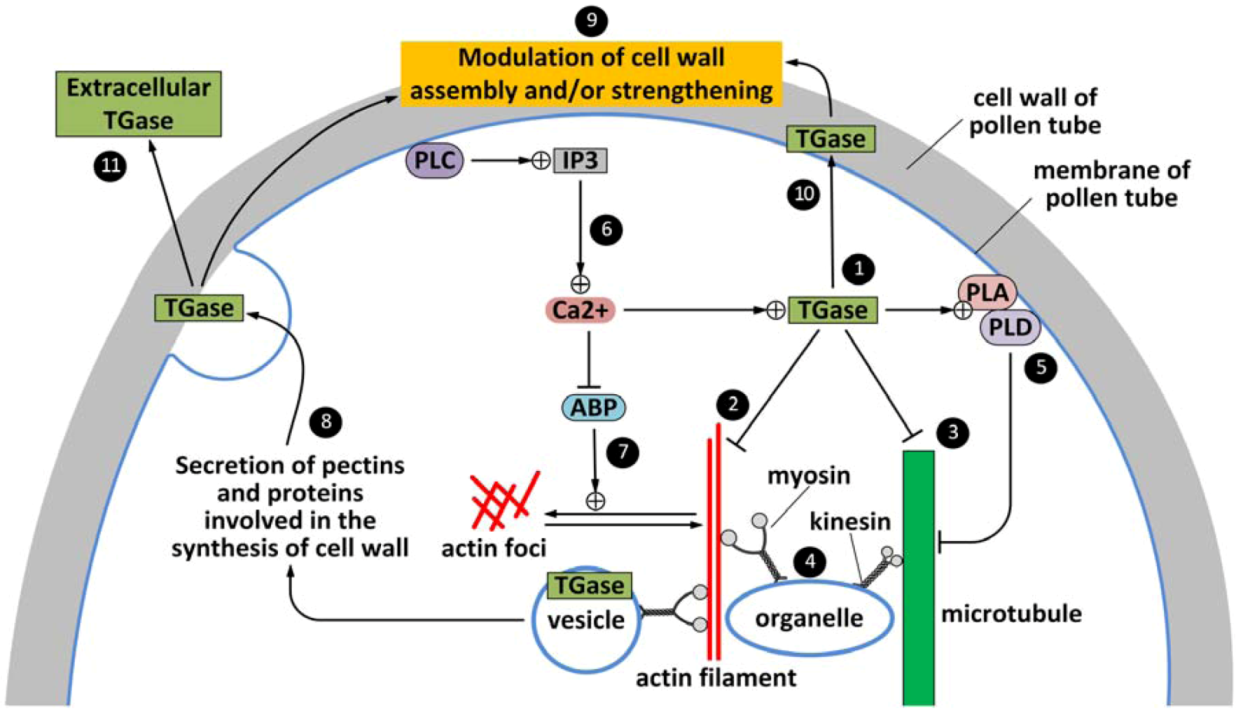
5. Extracellular TGase Released in the Pollen Germination Medium
6. Conclusions and Future Perspectives
Acknowledgements
References
- Palanivelu, R.; Preuss, D. Pollen tube targeting and axon guidance: Parallels in tip growth mechanisms. Trends Cell Biol. 2000, 10, 517–524. [Google Scholar] [CrossRef]
- Kroeger, J.H.; Zerzour, R.; Geitmann, A. Regulator or driving force? The role of turgor pressure in oscillatory plant cell growth. PLoS One 2011, 6, e18549. [Google Scholar] [CrossRef]
- Parre, E.; Geitmann, A. More than a leak sealant. The mechanical properties of callose in pollen tubes. Plant Physiol. 2005, 137, 274–286. [Google Scholar] [CrossRef]
- Aouar, L.; Chebli, Y.; Geitmann, A. Morphogenesis of complex plant cell shapes: The mechanical role of crystalline cellulose in growing pollen tubes. Sex. Plant Reprod. 2010, 23, 15–27. [Google Scholar] [CrossRef]
- Cai, G.; Faleri, C.; Del Casino, C.; Emons, A.M.C.; Cresti, M. Distribution of callose synthase, cellulose synthase and sucrose synthase in tobacco pollen tube is controlled in dissimilar ways by actin filaments and microtubules. Plant Physiol. 2011, 155, 1169–1190. [Google Scholar] [CrossRef]
- Cheung, A.Y.; Wu, H.M. Structural and signaling networks for the polar cell growth machinery in pollen tubes. Ann. Rev. Plant Biol. 2008, 59, 547–572. [Google Scholar] [CrossRef]
- Folk, J.E. Transglutaminases. Annu. Rev. Biochem. 1980, 49, 517–531. [Google Scholar] [CrossRef]
- Beninati, S.; Bergamini, C.M.; Piacentini, M. An overview of the first 50 years of transglutaminase research. Amino Acids 2009, 36, 591–598. [Google Scholar] [CrossRef]
- Lorand, L.; Graham, R.M. Transglutaminases: Crosslinking enzymes with pleiotropic functions. Nat. Rev. Mol. Cell Biol. 2003, 4, 140–156. [Google Scholar] [CrossRef]
- Icekson, I.; Apelbaum, A. Evidence for transglutaminase activity in plant tissue. Plant Physiol. 1987, 84, 972–974. [Google Scholar] [CrossRef]
- Serafini-Fracassini, D.; Del Duca, S.; D’Orazi, D. First evidence for polyamine conjugation mediated by an enzymic activity in plants. Plant Physiol. 1988, 87, 757–761. [Google Scholar] [CrossRef]
- Griffin, M.; Casadio, R.; Bergamini, C.M. Transglutaminases: Nature’s biological glues. Biochem. J. 2002, 368, 377–396. [Google Scholar] [CrossRef]
- Coussons, P.J.; Price, N.C.; Kelly, S.M.; Smith, B.; Sawyer, L. Factors that govern the specificity of transglutaminase-catalysed modification of proteins and peptides. Biochem. J. 1992, 282, 929930. [Google Scholar]
- Folk, J.E.; Park, M.H.; Chung, S.I.; Schrode, J.; Lester, E.P.; Cooper, H.L. Polyamines as physiological substrates for transglutaminases. J. Biol. Chem. 1980, 255, 3695–3700. [Google Scholar]
- Beninati, S.; Folk, J.E. Covalent polyamine-protein conjugates: Analysis and distribution. Adv. Exp. Med. Biol. 1988, 250, 411–422. [Google Scholar] [CrossRef]
- Della Mea, M.; Caparros-Ruiz, D.; Claparols, I.; Serafini-Fracassini, D.; Rigau, J. AtPng1p. The first plant transglutaminase. Plant Physiol. 2004, 135, 2046–2054. [Google Scholar] [CrossRef]
- Nicholas, B.; Smethurst, P.; Verderio, E.; Jones, R.; Griffin, M. Cross-linking of cellular proteins by tissue transglutaminase during necrotic cell death: A mechanism for maintaining tissue integrity. Biochem. J. 2003, 371, 413–422. [Google Scholar] [CrossRef]
- Mukherjee, B.B.; Nemir, M.; Beninati, S.; Cordella-Miele, E.; Singh, K.; Chackalaparampil, I.; Shanmugam, V.; DeVouge, M.W.; Mukherjee, A.B. Interaction of osteopontin with fibronectin and other extracellular matrix molecules. Ann. NY Acad. Sci. 1995, 760, 201–212. [Google Scholar] [CrossRef]
- Lentini, A.; Abbruzzese, A.; Caraglia, M.; Marra, M.; Beninati, S. Protein-polyamine conjugation by transglutaminase in cancer cell differentiation: Review article. Amino Acids 2004, 26, 331–337. [Google Scholar]
- Bagni, N.; Tassoni, A. Biosynthesis, oxidation and conjugation of aliphatic polyamines in higher plants. Amino Acids 2001, 20, 301–317. [Google Scholar] [CrossRef]
- Special Issue: Polyamines—Dedicated to the memory of Nello Bagni. Plant Physiol. Biochem. 2010, 48, 487–634.
- Bertossi, F.; Bagni, N.; Moruzzi, G.; Caldarera, C.M. Spermine as a new growth-promoting substance for Helianthus tuberosus (Jerusalem Artichoke) in vitro. Experientia 1965, 21, 80–81. [Google Scholar] [CrossRef]
- Tasco, G.; Della Mea, M.; Serafini-Fracassini, D.; Casadio, R. Building a low resolution model of a transglutaminase domain of an hypothetical N-glycanase from Arabidopsis thaliana. Amino Acids 2003, 25, 197. [Google Scholar]
- Masahara-Negishi, Y.; Hosomi, A.; Della Mea, M.; Serafini-Fracassini, D.; Suzuki, T. A plant peptide: N-glycanase orthologue facilitates glycoprotein ER-associated degradation in yeast. Biochim. Biophys. Acta 2012, 1820, 1457–1462. [Google Scholar] [CrossRef]
- Beninati, S.; Iorio, R.A.; Tasco, G.; Serafini-Fracassini, D.; Casadio, R.; Del Duca, S. Expression of different forms of transglutaminases by immature cells of Helianthus tuberosus sprout apices. Amino Acids 2012, 44, 271–283. [Google Scholar]
- Serafini-Fracassini, D.; Del Duca, S. Transglutaminases: Widespread cross-linking enzymes in plants. Ann. Bot. 2008, 102, 145–152. [Google Scholar] [CrossRef]
- Villalobos, E.; Santos, M.; Talavera, D.; Rodriguez-Falcon, M.; Torne, J.M. Molecular cloning and characterization of a maize transglutaminase complementary DNA. Gene 2004, 336, 93–104. [Google Scholar] [CrossRef]
- Del Duca, S.; Tidu, V.; Bassi, R.; Esposito, C.; Serafini-Fracassini, D. Identification of chlorophyll-a/b proteins as substrates of transglutaminase activity in isolated chloroplasts of Helianthus tuberosus L. Planta 1994, 193, 283–289. [Google Scholar]
- Della Mea, M.; de Filippis, F.; Genovesi, V.; Serafini Fracassini, D.; Del Duca, S. The acropetal wave of developmental cell death of tobacco corolla is preceded by activation of transglutaminase in different cell compartments. Plant Physiol. 2007, 144, 1211–1222. [Google Scholar] [CrossRef]
- Del Duca, S.; Serafini-Fracassini, D. Transglutaminases of higher, lower plants and fungi. Prog. Exp. Tumor Res. 2005, 38, 223–247. [Google Scholar] [CrossRef]
- Serafini-Fracassini, D.; Della Mea, M.; Tasco, G.; Casadio, R.; Del Duca, S. Plant and animal transglutaminases: Do similar functions imply similar structures? Amino Acids 2009, 36, 643657. [Google Scholar]
- Della Mea, M.; di Sandro, A.; Dondini, L.; Del Duca, S.; Vantini, F.; Bergamini, C.; Bassi, R.; SerafiniFracassini, D. A Zea mays 39-kDa thylakoid transglutaminase catalyses the modification by polyamines of light-harvesting complex II in a light-dependent way. Planta 2004, 219, 754–764. [Google Scholar]
- Del Duca, S.; Faleri, C.; Iorio, R.A.; Cresti, M.; Serafini-Fracassini, D.; Cai, G. Distribution of transglutaminase in pear pollen tubes in relation to cytoskeleton and membrane dynamics. Plant Physiol. 2013. [Google Scholar] [CrossRef]
- Di Sandro, A.; Serafini-Fracassini, D.; Del Duca, S.; Della Mea, M.; de Franceschi, P.; Dondini, L.; Faleri, C.; Cai, G.; Sansavini, S. Pollen transglutaminase in pear self incompatibility and relationship with S-RNases and S-allele variability. Acta Hort. 2008, 800, 423–429. [Google Scholar]
- Di Sandro, A.; Del Duca, S.; Verderio, E.; Hargreaves, A.J.; Scarpellini, A.; Cai, G.; Cresti, M.; Faleri, C.; Iorio, R.A.; Shigehisa, H.; et al. An extracellular transglutaminase is required for apple pollen tube growth. Biochem. J. 2010, 429, 261–271. [Google Scholar] [CrossRef]
- Gentile, A.; Antognoni, F.; Iorio, R.A.; Distefano, G.; Las, C.G.; La, M.S.; Serafini-Fracassini, D.; Del Duca, S. Polyamines and transglutaminase activity are involved in compatible and selfincompatible pollination of Citrus grandis. Amino Acids 2012, 42, 1025–1035. [Google Scholar] [CrossRef]
- Iorio, R.A.; di Sandro, A.; Paris, R.; Pagliarani, G.; Tartarini, S.; Ricci, G.; Serafini-Fracassini, D.; Verderio, E.; Del Duca, S. Simulated environmental criticalities affect transglutaminase of Malus and Corylus pollens having different allergenic potential. Amino Acids 2012, 42, 1007–1024. [Google Scholar] [CrossRef]
- Hegyi, G.; Michel, H.; Shabanowitz, J.; Hunt, D.F.; Chatterjie, N.; Healy-Louie, G.; Elzinga, M. Gln-41 is intermolecularly cross-linked to Lys-113 in F-actin by N-(4-azidobenzoyl)-putrescine. Protein Sci. 1992, 1, 132–144. [Google Scholar]
- Nemes, Z., Jr.; Adany, R.; Balazs, M.; Boross, P.; Fesus, L. Identification of cytoplasmic actin as an abundant glutaminyl substrate for tissue transglutaminase in HL-60 and U937 cells undergoing apoptosis. J. Biol. Chem. 1997, 272, 20577–20583. [Google Scholar]
- Eligula, L.; Chuang, L.; Phillips, M.L.; Motoki, M.; Seguro, K.; Muhlrad, A. Transglutaminase-induced cross-linking between subdomain 2 of G-actin and the 636–642 lysine-rich loop of myosin subfragment 1. Biophys. J. 1998, 74, 953–963. [Google Scholar] [CrossRef]
- Chowdhury, Z.A.; Barsigian, C.; Chalupowicz, G.D.; Bach, T.L.; Garcia-Manero, G.; Martinez, J. Colocalization of tissue transglutaminase and stress fibers in human vascular smooth muscle cells and human umbilical vein endothelial cells. Exp. Cell Res. 1997, 231, 38–49. [Google Scholar] [CrossRef]
- Miller, C.C.; Anderton, B.H. Transglutaminase and the neuronal cytoskeleton in Alzheimer’s disease. J. Neurochem. 1986, 46, 1912–1922. [Google Scholar] [CrossRef]
- Esposito, C.; Mariniello, L.; Cozzolino, A.; Amoresano, A.; Orru, S.; Porta, R. Rat coagulating gland secretion contains a kinesin heavy chain-like protein acting as a type IV transglutaminase substrate. Biochemistry 2001, 40, 4966–4971. [Google Scholar] [CrossRef]
- Cai, G.; Cresti, M. Organelle motility in the pollen tube: A tale of 20 years. J. Exp. Bot. 2009, 60, 495–508. [Google Scholar] [CrossRef]
- Daher, F.B.; Geitmann, A. Actin is involved in pollen tube tropism through redefining the spatial targeting of secretory vesicles. Traffic 2011, 12, 1537–1551. [Google Scholar] [CrossRef]
- Cardenas, L.; Lovy-Wheeler, A.; Wilsen, K.L.; Hepler, P.K. Actin polymerization promotes the reversal of streaming in the apex of pollen tubes. Cell Motil. Cytoskeleton 2005, 61, 112–127. [Google Scholar] [CrossRef]
- Staiger, C.J.; Poulter, N.S.; Henty, J.L.; Franklin-Tong, V.E.; Blanchoin, L. Regulation of actin dynamics by actin-binding proteins in pollen. J. Exp. Bot. 2010, 61, 1969–1986. [Google Scholar] [CrossRef]
- Kost, B. Spatial control of Rho (Rac-Rop) signaling in tip-growing plant cells. Trends Cell Biol. 2008, 18, 119–127. [Google Scholar] [CrossRef]
- Cardenas, L.; Lovy-Wheeler, A.; Kunkel, J.G.; Hepler, P.K. Pollen tube growth oscillations and intracellular calcium levels are reversibly modulated by actin polymerization. Plant Physiol. 2008, 146, 1611–1621. [Google Scholar] [CrossRef]
- Del Casino, C.; Li, Y.; Moscatelli, A.; Scali, M.; Tiezzi, A.; Cresti, M. Distribution of microtubules during the growth of tobacco pollen tubes. Biol. Cell 1993, 79, 125–132. [Google Scholar]
- Cheung, A.Y.; Duan, Q.H.; Costa, S.S.; de Graaf, B.H.J.; di Stilio, V.S.; Feijo, J.; Wu, H.M. The dynamic pollen tube cytoskeleton: Live cell studies using actin-binding and microtubule-binding reporter proteins. Mol. Plant 2008, 1, 686–702. [Google Scholar] [CrossRef]
- Laitiainen, E.; Nieminen, K.M.; Vihinen, H.; Raudaskoski, M. Movement of generative cell and vegetative nucleus in tobacco pollen tubes is dependent on microtubule cytoskeleton but independent of the synthesis of callose plugs. Sex. Plant Reprod. 2002, 15, 195–204. [Google Scholar] [CrossRef]
- Astrom, H.; Sorri, O.; Raudaskoski, M. Role of microtubules in the movement of the vegetative nucleus and generative cell in tobacco pollen tubes. Sex. Plant Reprod. 1995, 8, 61–69. [Google Scholar]
- Del Duca, S.; Bregoli, A.M.; Bergamini, C.; Serafini-Fracassini, D. Transglutaminase-catalyzed modification of cytoskeletal proteins by polyamines during the germination of Malus domestica pollen. Sex. Plant Reprod. 1997, 10, 89–95. [Google Scholar] [CrossRef]
- Del Duca, S.; Serafini-Fracassini, D.; Bonner, P.L.; Cresti, M.; Cai, G. Effects of post-translational modifications catalyzed by pollen transglutaminase on the functional properties of microtubules and actin filaments. Biochem. J. 2009, 418, 651–664. [Google Scholar] [CrossRef]
- Xue, X.; Du, F.; Zhu, J.; Ren, H. Actin organization and regulation during pollen tube growth. Front. Biol. 2011, 6, 40–51. [Google Scholar] [CrossRef]
- Reed, N.A.; Cai, D.; Blasius, T.L.; Jih, G.T.; Meyhofer, E.; Gaertig, J.; Verhey, K.J. Microtubule acetylation promotes kinesin-1 binding and transport. Curr. Biol. 2006, 16, 2166–2172. [Google Scholar]
- Cai, G.; Cresti, M. Microtubule motors and pollen tube growth—Still an open question. Protoplasma 2010, 247, 131–143. [Google Scholar]
- Bosch, M.; Franklin-Tong, V.E. Self-incompatibility in Papaver: Signalling to trigger PCD in incompatible pollen. J. Exp. Bot. 2008, 59, 481–490. [Google Scholar]
- Poulter, N.S.; Vatovec, S.; Franklin-Tong, V.E. Microtubules are a target for self-incompatibility signaling in Papaver pollen. Plant Physiol. 2008, 146, 1358–1367. [Google Scholar]
- McClure, B.; Cruz-Garcia, F.; Romero, C. Compatibility and incompatibility in S-RNase-based systems. Ann. Bot. 2011, 108, 647–658. [Google Scholar]
- Cruz-Garcia, F.; Nathan, H.C.; Kim, D.; McClure, B. Stylar glycoproteins bind to S-RNase in vitro. Plant J. 2005, 42, 295–304. [Google Scholar]
- Liu, Z.Q.; Xu, G.H.; Zhang, S.L. Pyrus pyrifolia stylar S-RNase induces alterations in the actin cytoskeleton in self-pollen and tubes in vitro. Protoplasma 2007, 232, 61–67. [Google Scholar]
- Cai, G.; Della Mea, M.; Faleri, C.; Serafini-Fracassini, D.; Del Duca, S. Spermine regulates the development and programmed cell death of Nicotiana tabacum corolla. Personal communication, University of Bologna: Bologna, Italy, 2013. [Google Scholar]
- Del Duca, S.; Cai, G.; di Sandro, A.; Serafini-Fracassini, D. Compatible and self-incompatible pollination in Pyrus communis displays different polyamine levels and transglutaminase activity. Amino Acids 2010, 38, 659–667. [Google Scholar]
- Dhonukshe, P.; Laxalt, A.M.; Goedhart, J.; Gadella, T.W.J.; Munnik, T. Phospholipase D activation correlates with microtubule reorganization in living plant cells. Plant Cell 2003, 15, 2666–2679. [Google Scholar]
- Gardiner, J.; Andreeva, Z.; Barton, D.; Ritchie, A.; Overall, R.; Marc, J. The phospholipase A inhibitor, aristolochic acid, disrupts cortical microtubule arrays and root growth in Arabidopsis. Plant Biol. 2008, 10, 725–731. [Google Scholar]
- Kim, H.J.; Ok, S.H.; Bahn, S.C.; Jang, J.; Oh, S.A.; Park, S.K.; Twell, D.; Ryu, S.B.; Shin, J.S. Endoplasmic reticulum- and Golgi-localized phospholipase A2 plays critical roles in Arabidopsis pollen development and germination. Plant Cell 2011, 23, 94–110. [Google Scholar]
- Zemskov, E.A.; Mikhailenko, I.; Hsia, R.C.; Zaritskaya, L.; Belkin, A.M. Unconventional secretion of tissue transglutaminase involves phospholipid-dependent delivery into recycling endosomes. PLoS One 2011, 6, e19414. [Google Scholar]
- Parton, R.M.; Fischer-Parton, S.; Watahiki, M.K.; Trewavas, A.J. Dynamics of the apical vesicle accumulation and the rate of growth are related in individual pollen tubes. J. Cell Sci. 2001, 114, 2685–2695. [Google Scholar]
- Certal, A.C.; Almeida, R.B.; Carvalho, L.M.; Wong, E.; Moreno, N.; Michard, E.; Carneiro, J.; RodriguezLeon, J.; Wu, H.M.; Cheung, A.Y.; et al. Exclusion of a proton ATPase from the apical membrane is associated with cell polarity and tip growth in Nicotiana tabacum pollen tubes. Plant Cell 2008, 20, 614–634. [Google Scholar]
- Ding, Y.; Wang, J.; Wang, J.; Stierhof, Y.D.; Robinson, D.G.; Jiang, L. Unconventional protein secretion. Trends Plant Sci. 2012, 17, 606–615. [Google Scholar]
- Toyooka, K.; Goto, Y.; Asatsuma, S.; Koizumi, M.; Mitsui, T.; Matsuoka, K. A mobile secretory vesicle cluster involved in mass transport from the Golgi to the plant cell exterior. Plant Cell 2009, 21, 1212–1229. [Google Scholar]
- Nakaoka, H.; Perez, D.M.; Baek, K.J.; Das, T.; Husain, A.; Misono, K.; Im, M.J.; Graham, R.M. Gh: A GTPbinding protein with transglutaminase activity and receptor signaling function. Science 1994, 264, 1593–1596. [Google Scholar]
- Antonyak, M.A.; McNeill, C.J.; Wakshlag, J.J.; Boehm, J.E.; Cerione, R.A. Activation of the Ras-ERK pathway inhibits retinoic acid-induced stimulation of tissue transglutaminase expression in NIH3T3 cells. J. Biol. Chem. 2003, 278, 15859–15866. [Google Scholar]
- Baek, K.J.; Kang, S.; Damron, D.; Im, M. Phospholipase Cdelta1 is a guanine nucleotide exchanging factor for transglutaminase II (Galpha h) and promotes alpha 1B-adrenoreceptor-mediated GTP binding and intracellular calcium release. J. Biol. Chem. 2001, 276, 5591–5597. [Google Scholar]
- Dowd, P.E.; Coursol, S.; Skirpan, A.L.; Kao, T.H.; Gilroy, S. Petunia phospholipase c1 is involved in pollen tube growth. Plant Cell 2006, 18, 1438–1453. [Google Scholar]
- Helling, D.; Possart, A.; Cottier, S.; Klahre, U.; Kost, B. Pollen tube tip growth depends on plasma membrane polarization mediated by tobacco PLC3 activity and endocytic membrane recycling. Plant Cell 2006, 18, 3519–3534. [Google Scholar]
- Dinnella, C.; Serafini-Fracassini, D.; Grandi, B.; Del Duca, S. The cell cycle in Helianthus tuberosus: Analysis of polyamine-endogenous protein conjugates by transglutaminase-like activity. Plant Physiol. Biochem. 1992, 30, 531–539. [Google Scholar]
- Waffenschmidt, S.; Kusch, T.; Woessner, J.P. A transglutaminase immunologically related to tissue transglutaminase catalyzes cross-linking of cell wall proteins in Chlamydomonas reinhardtii. Plant Physiol. 1999, 121, 1003–1015. [Google Scholar]
- Brunner, F.; Rosahl, S.; Lee, J.; Rudd, J.J.; Geiler, C.; Kauppinen, S.; Rasmussen, G.; Scheel, D.; Nurnberger, T. Pep-13, a plant defense-inducing pathogen-associated pattern from Phytophthora transglutaminases. EMBO J. 2002, 21, 6681–6688. [Google Scholar]
- Iorio, R.A.; di Sandro, A.; Scarpellini, A.; Del Duca, S.; Serafini-Fracassini, D.; Verderio, E. Visualisation of transglutaminase-mediated cross-linking activity in germinating pollen by laser confocal microscopy. Plant Biosyst. 2008, 142, 360–365. [Google Scholar]
- Furutani, Y.; Kato, A.; Notoya, M.; Ghoneim, M.A.; Hirose, S. A simple assay and histochemical localization of transglutaminase activity using a derivative of green fluorescent protein as substrate. J. Histochem. Cytochem. 2001, 49, 247–258. [Google Scholar]
- Verderio, E.; Nicholas, B.; Gross, S.; Griffin, M. Regulated expression of tissue transglutaminase in Swiss 3T3 fibroblasts: Effects on the processing of fibronectin, cell attachment, and cell death. Exp. Cell Res. 1998, 239, 119–138. [Google Scholar]
- Chae, K.; Lord, E.M. Pollen tube growth and guidance: Roles of small, secreted proteins. Ann. Bot. 2011, 108, 627–636. [Google Scholar]
- Sun, Y.; Qian, H.; Xu, X.-D.; Han, Y.; Yen, L.-F.; Sun, D.-Y. Integrin-like proteins in the pollen tube: Detection, localization and function. Plant Cell Physiol. 2000, 41, 1136–1142. [Google Scholar]
- D’Orazi, D.; Bagni, N. In vitro interactions between polyamines and pectic substances. Biochem. Biophys. Res. Commun. 1987, 148, 1259–1263. [Google Scholar]
- Zonia, L.; Munnik, T. Understanding pollen tube growth: The hydrodynamic model versus the cell wall model. Trends Plant Sci. 2011, 16, 347–352. [Google Scholar]
- Majewska-Sawka, A.; Nothnagel, E.A. The multiple roles of arabinogalactan proteins in plant development. Plant Physiol. 2000, 122, 3–10. [Google Scholar]
- Lee, C.B.; Swatek, K.N.; McClure, B. Pollen proteins bind to the C-terminal domain of Nicotiana alata pistil arabinogalactan proteins. J. Biol. Chem. 2008, 283, 26965–26973. [Google Scholar]
- Mollet, J.C.; Kim, S.; Jauh, G.Y.; Lord, E.M. Arabinogalactan proteins, pollen tube growth, and the reversible effects of Yariv phenylglycoside. Protoplasma 2002, 219, 89–98. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Cai, G.; Serafini-Fracassini, D.; Del Duca, S. Regulation of Pollen Tube Growth by Transglutaminase. Plants 2013, 2, 87-106. https://doi.org/10.3390/plants2010087
Cai G, Serafini-Fracassini D, Del Duca S. Regulation of Pollen Tube Growth by Transglutaminase. Plants. 2013; 2(1):87-106. https://doi.org/10.3390/plants2010087
Chicago/Turabian StyleCai, Giampiero, Donatella Serafini-Fracassini, and Stefano Del Duca. 2013. "Regulation of Pollen Tube Growth by Transglutaminase" Plants 2, no. 1: 87-106. https://doi.org/10.3390/plants2010087