Regulation Systems of Bacteria such as Escherichia coli in Response to Nutrient Limitation and Environmental Stresses

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
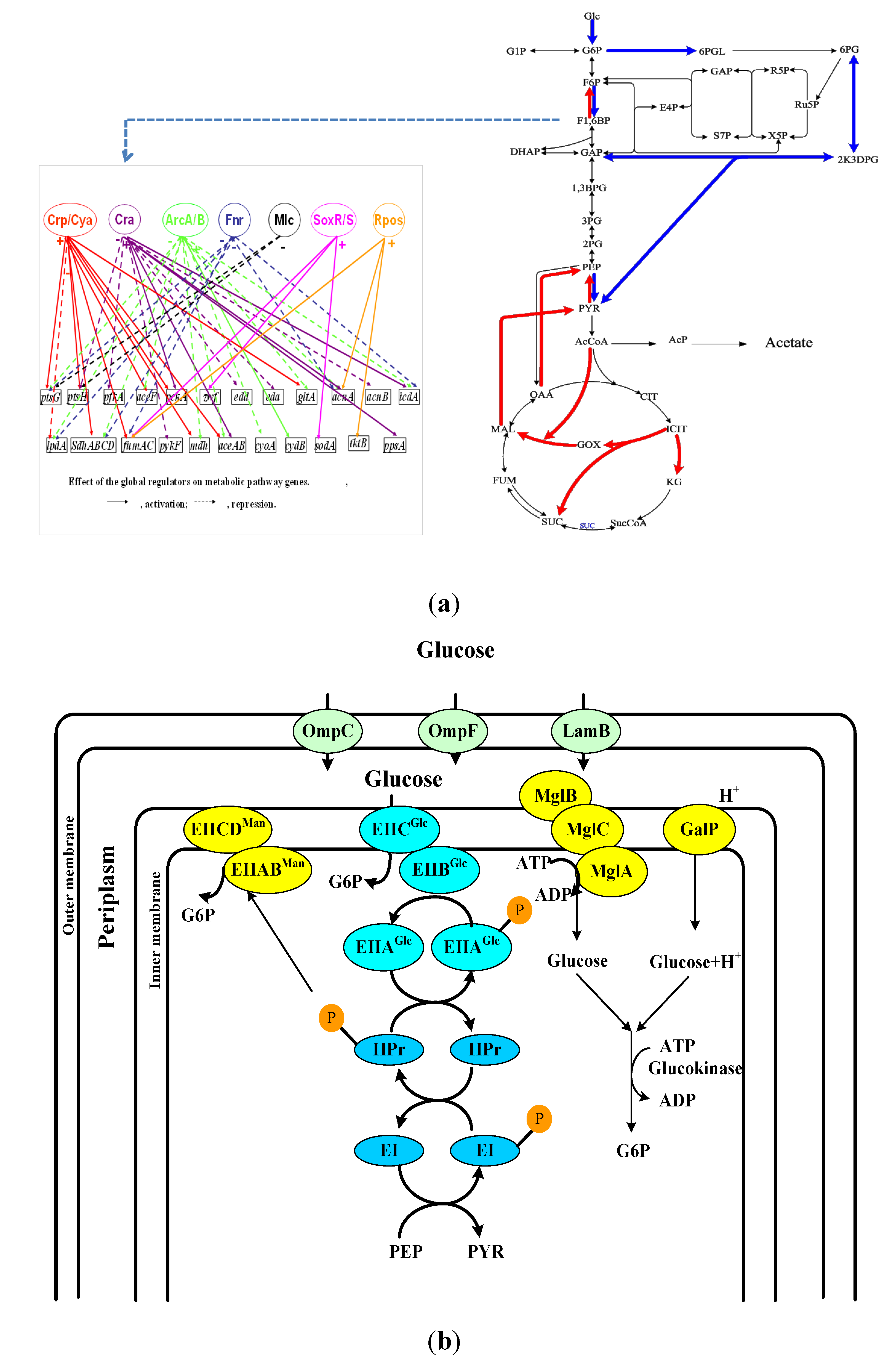
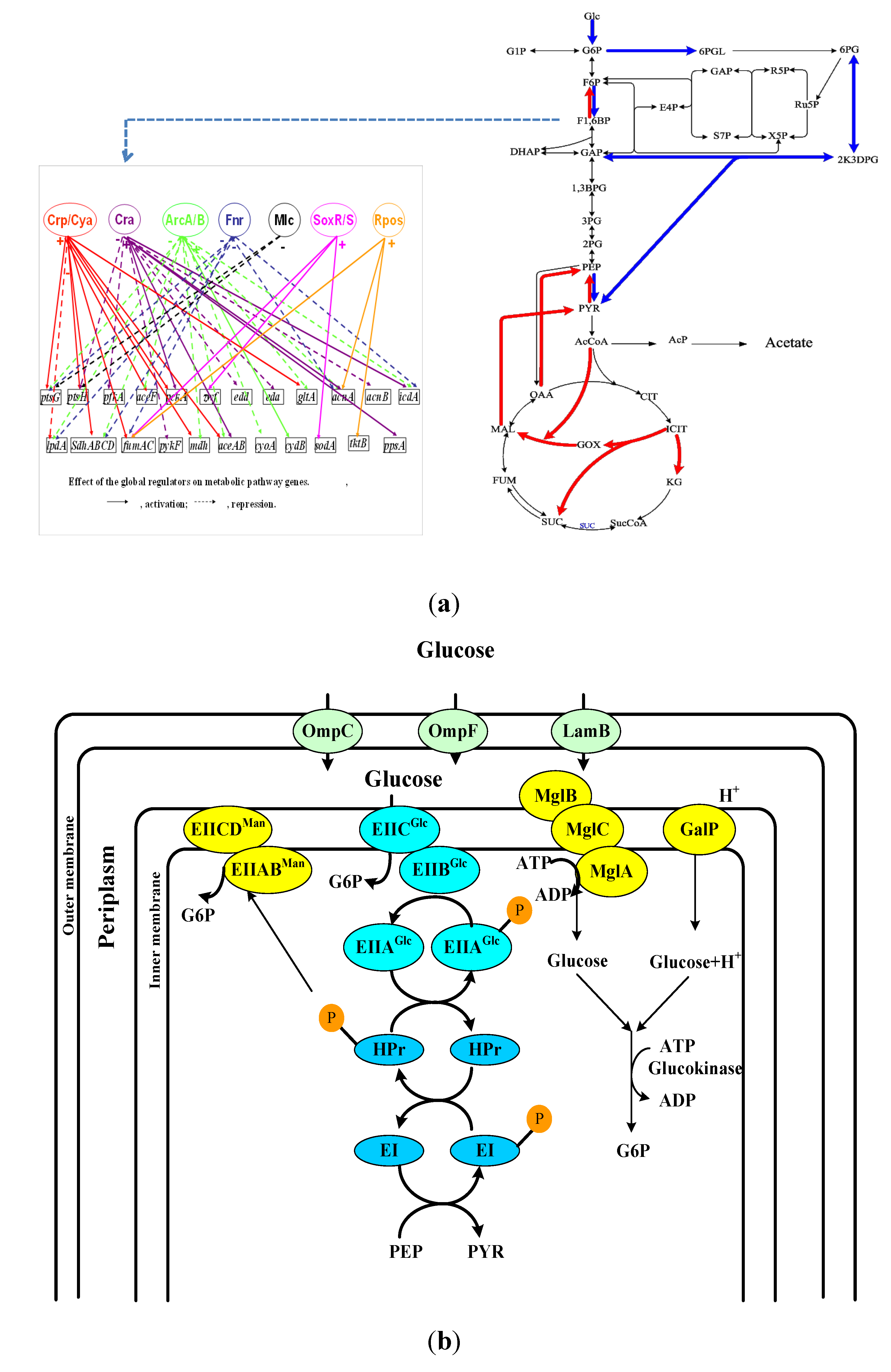
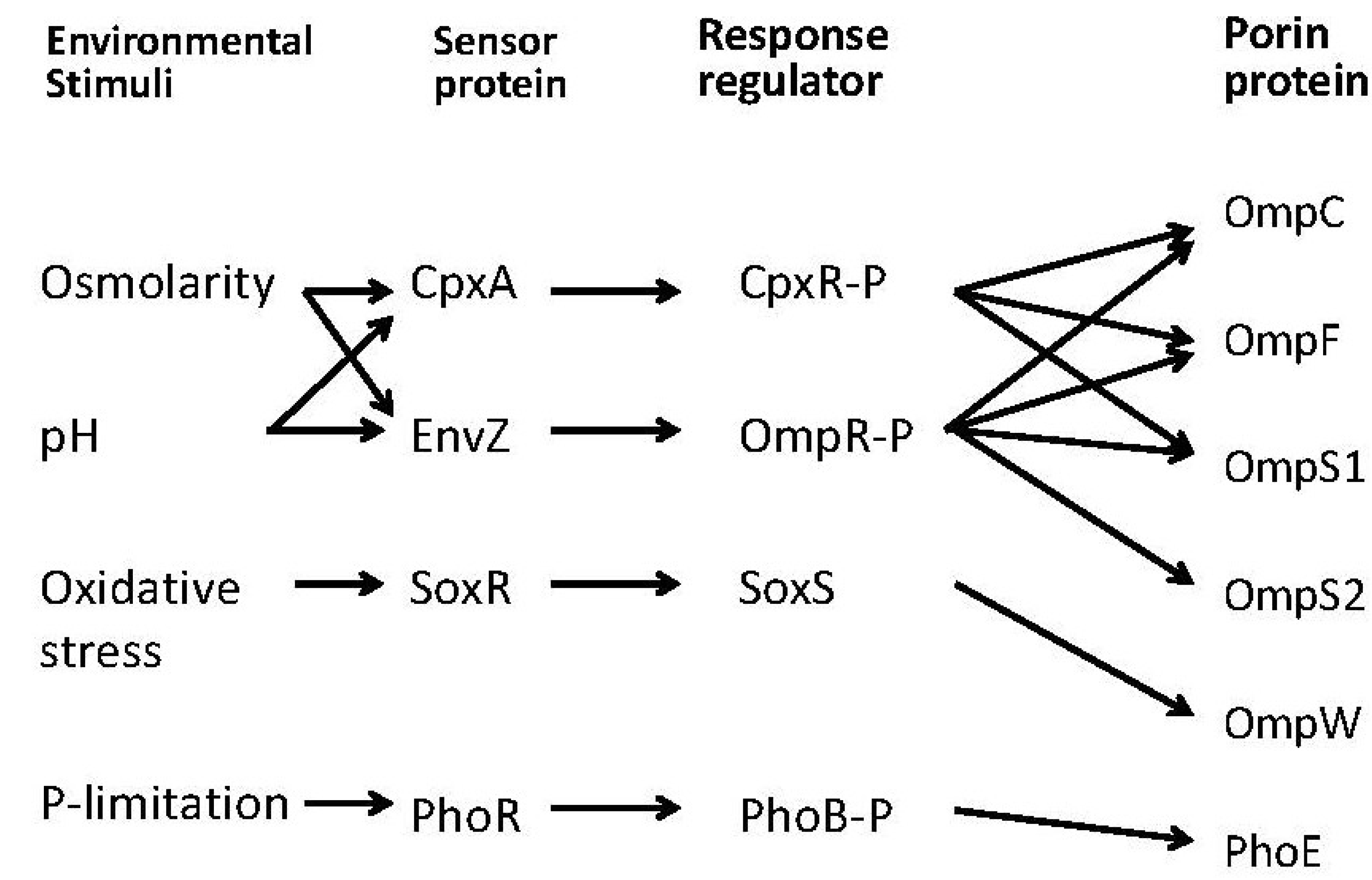
2. Transport of Nutrient Molecules and PTS

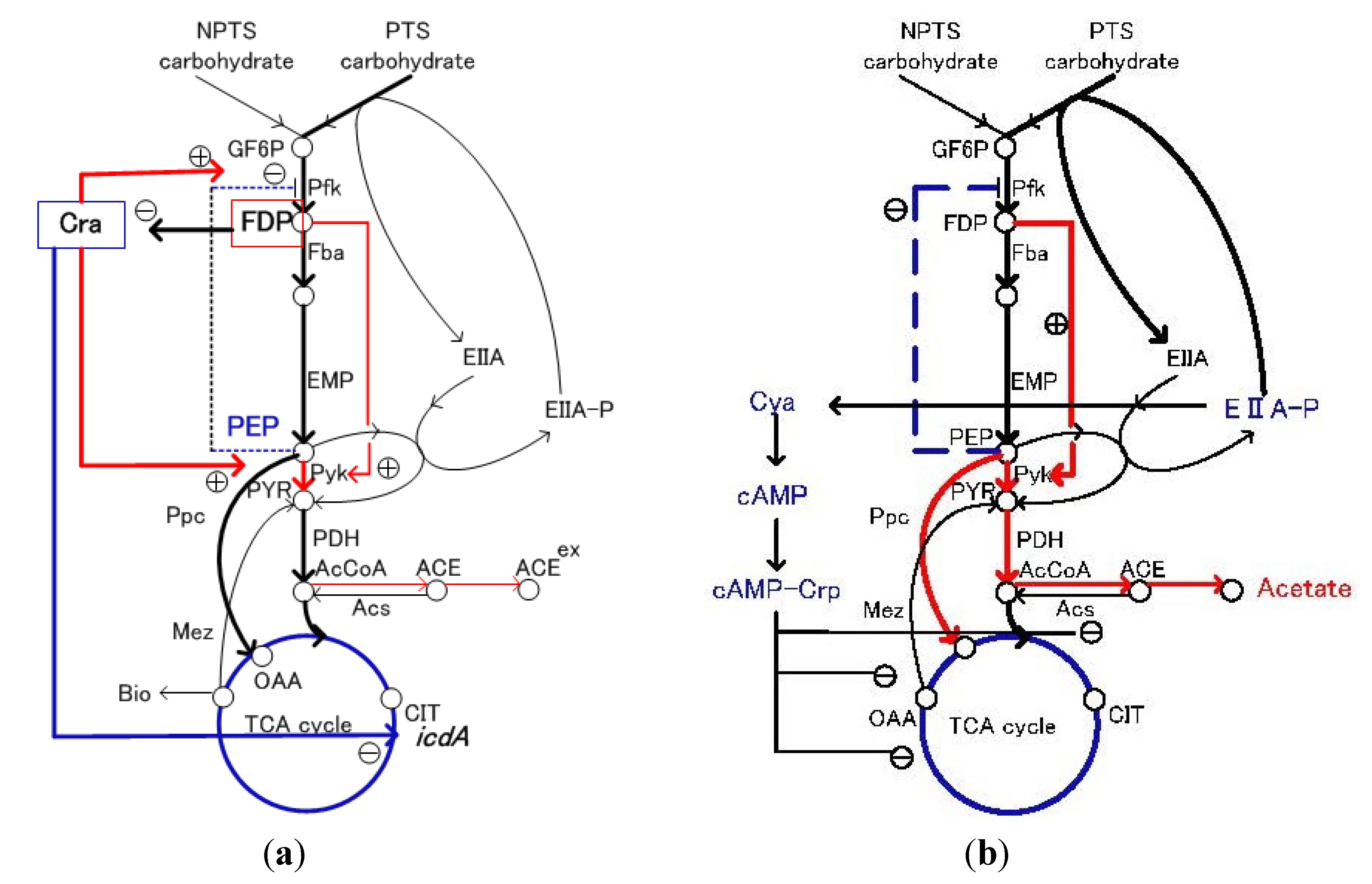
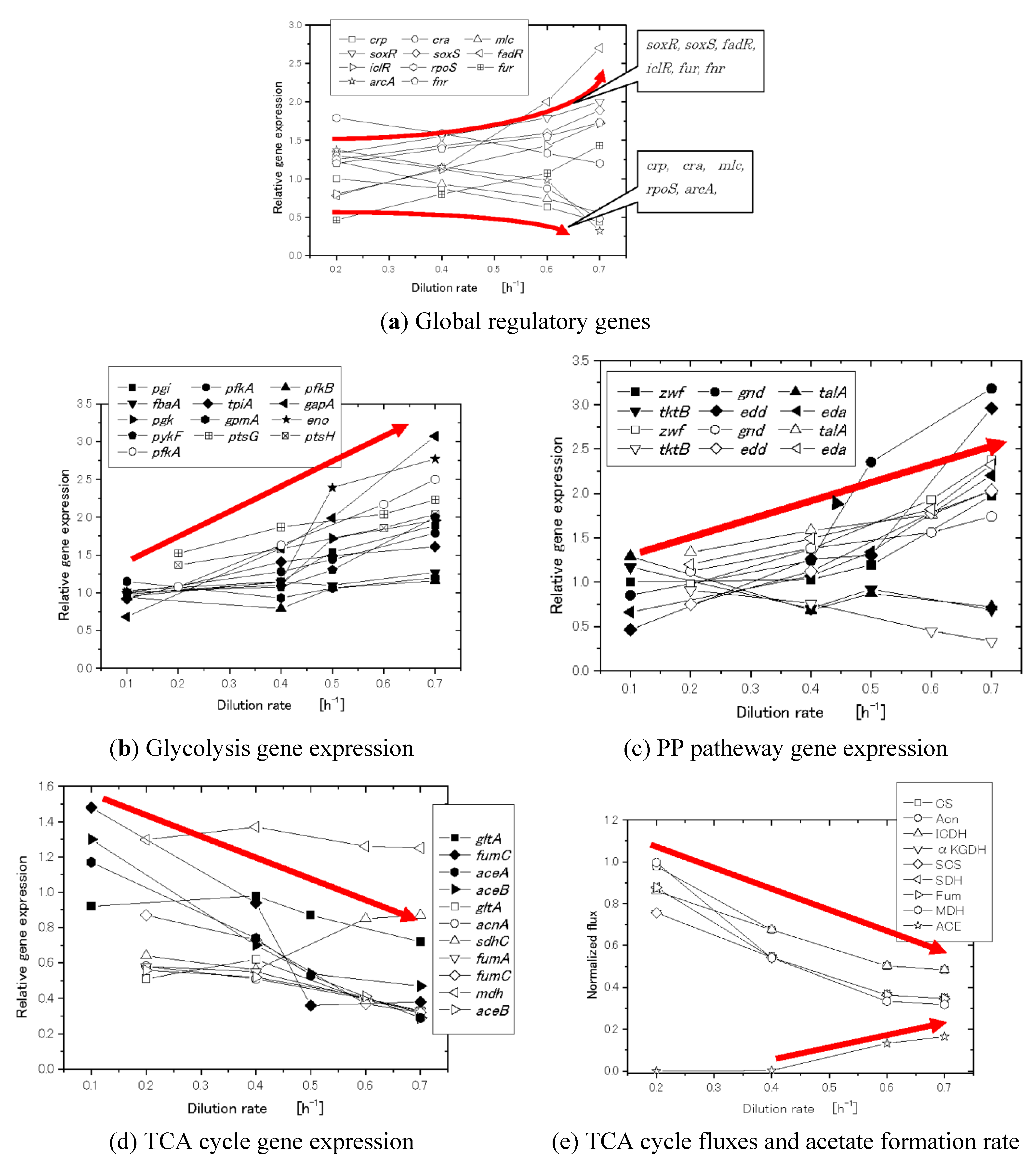
3. Flux Sensor


4. Carbon Storage Regulation
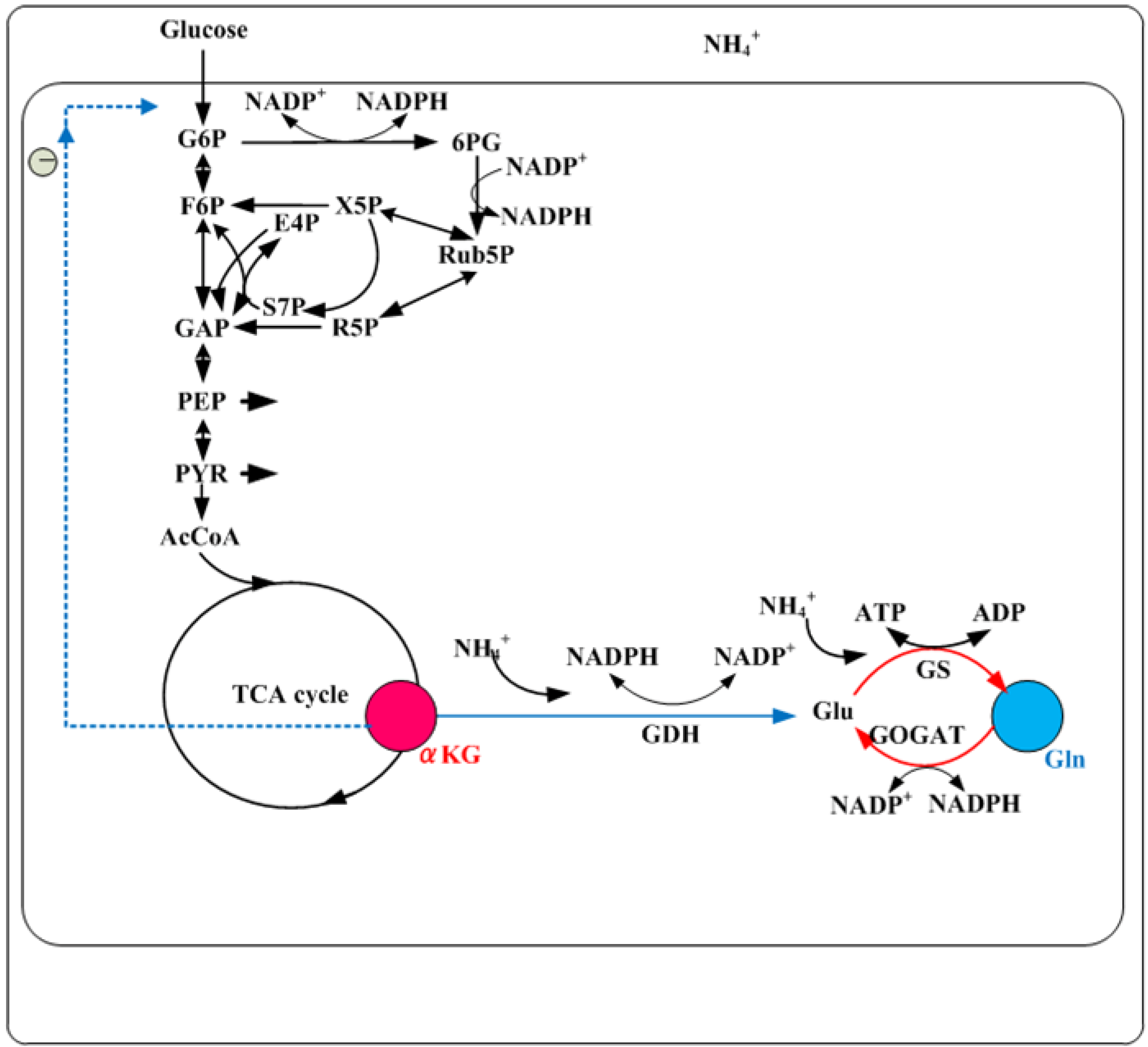
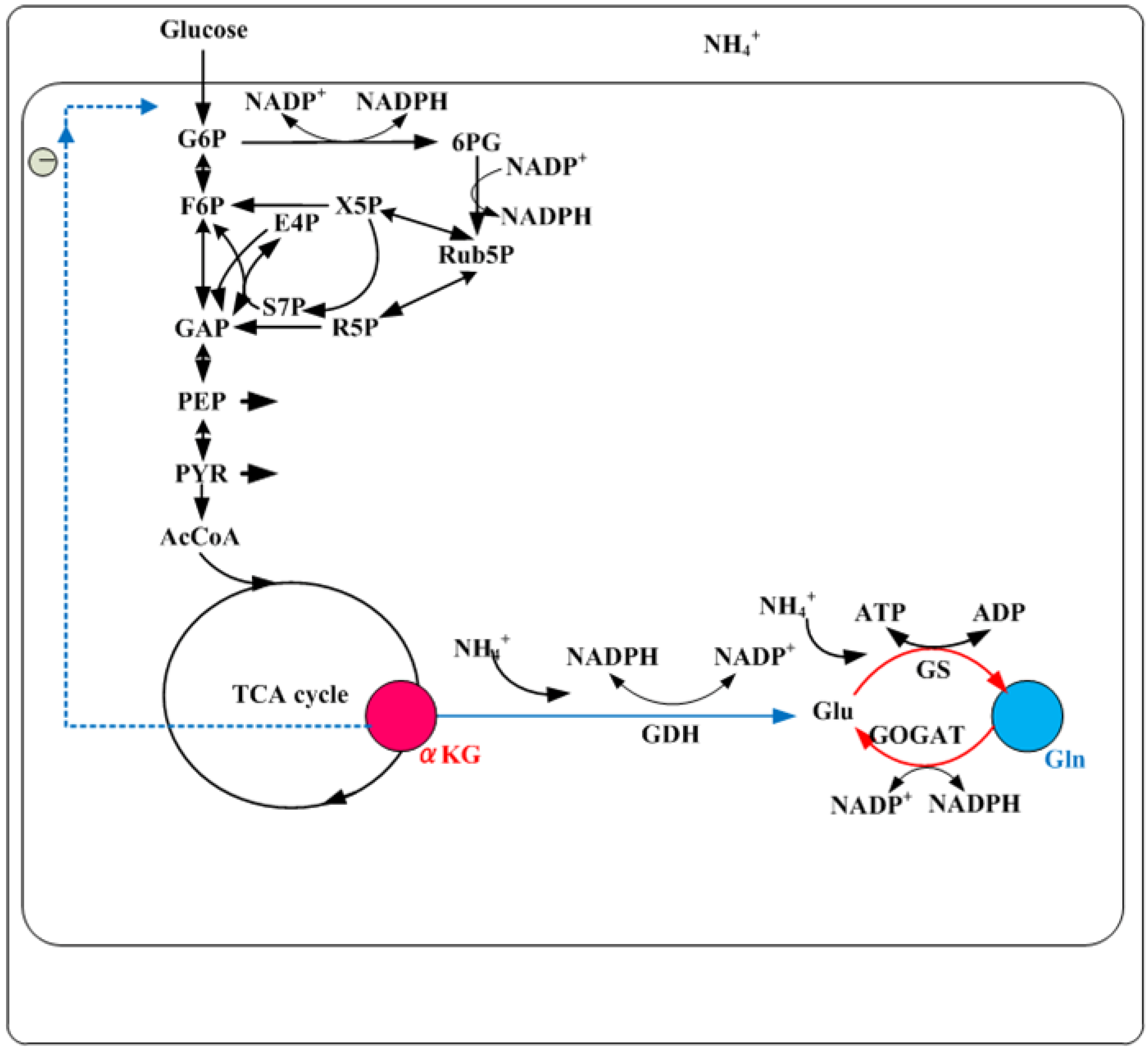
5. Nitrogen Regulation
6. The Role of PTSNtr
7. Regulation for S-limitation
8. Phosphate Regulation
9. Regulation for Ion Uptake
10. Growth-Phase Dependent Fis Expression
11. Stringent Response to Nutrient Starvation
12. Stationary Phase Regulation by RpoS
13. Biofilm, Motility by Flagella, and Quorum Sensing
14. Death Phase and Long-Term Stationary Phase Metabolism
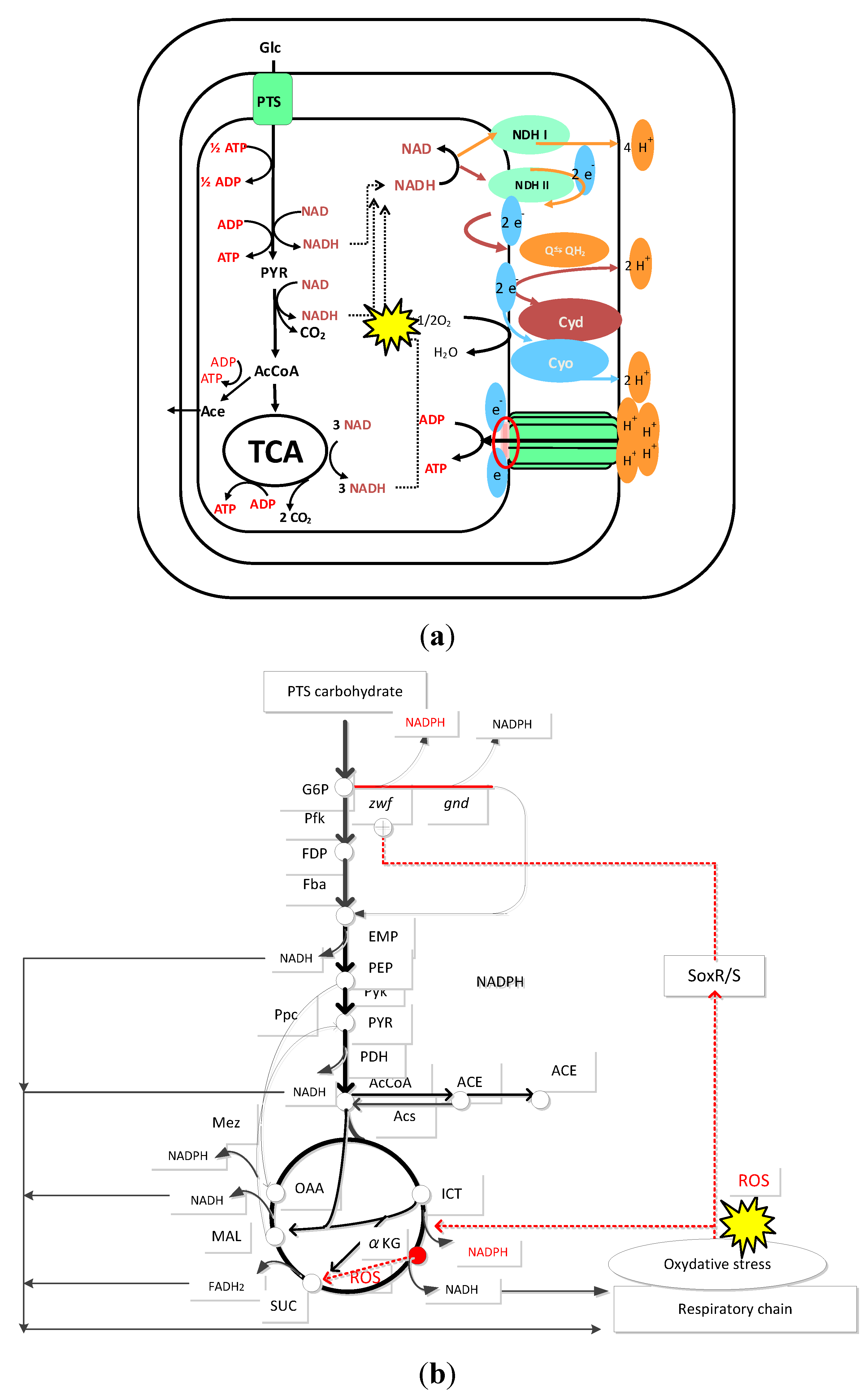
15. Effect of Oxidative Stress on the Metabolism
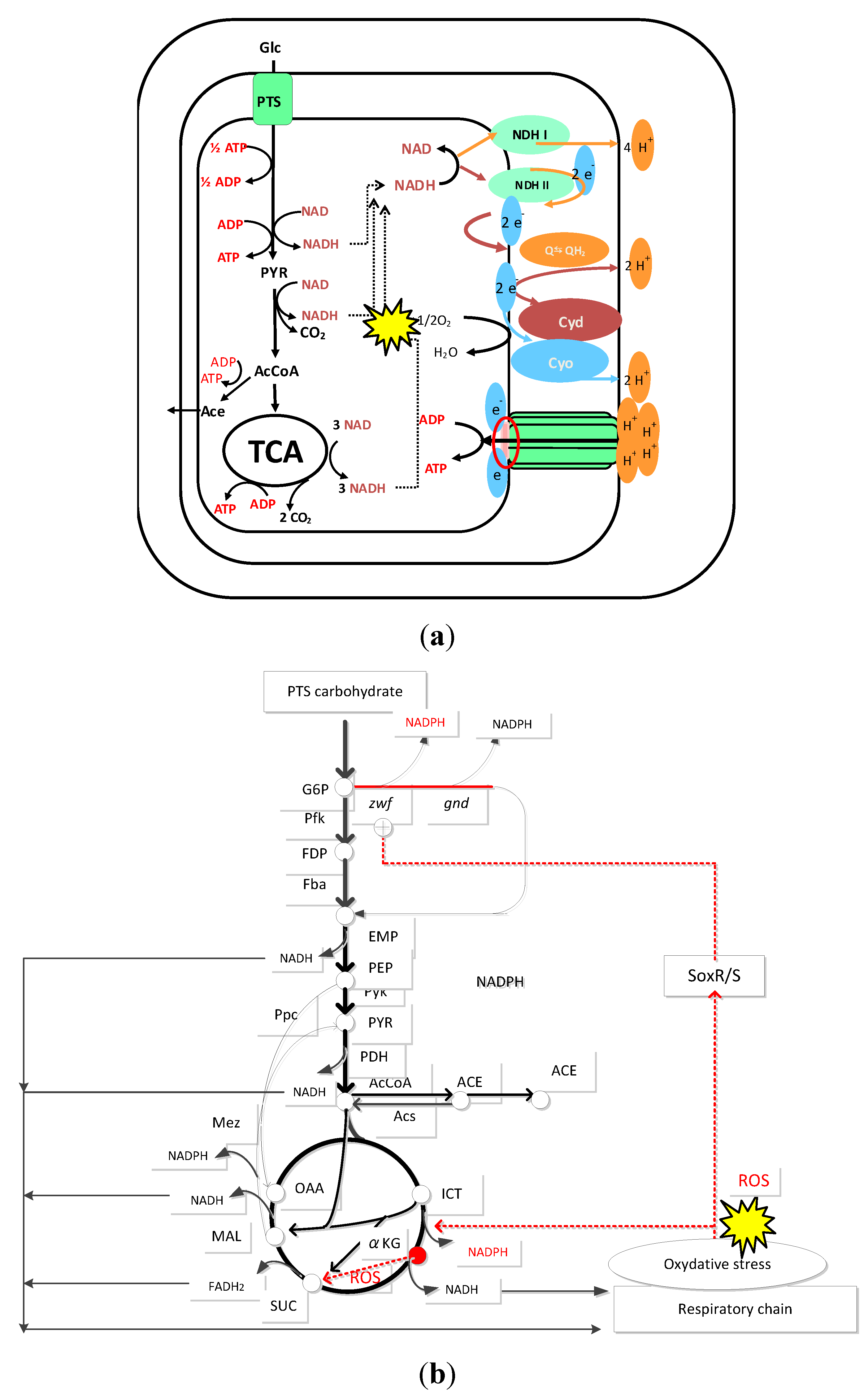
16. Acid Shock Response
17. Heat Shock Response
18. Tolerance to Various Stresses in Biofuels Production by Microorganisms
19. Concluding Remarks
| Abbreviations | |
|---|---|
| PP pathway | pentose phosphate pathway |
| TCA cycle | tricarboxylic acid cycle |
| PTS | phosphotransferase system |
| EI | enzyme I |
| EII | enzyme II |
| HPr | histidine-phosphorylatable protein |
| Cya | adenylate cyclase |
| CIT | citrate |
| E4P | erythrose-4-phosphate |
| FDP | fructose-1,6-bisphosphate |
| F6P | fructose-6-phosphate |
| G6P | glucose-6-phosphate |
| GAP | glycelaldehyde-3-phosphate |
| GLC | glucose |
| GOX | glyoxylate |
| ICI | isocitrate |
| KDPG | 2-keto-3-deoxy-6-phosphogluconate |
| αKG | α-ketoglutarate |
| MAL | malate |
| OAA | oxaloacetate |
| PEP | phosphoenol pyruvate |
| 6PG | 6-phosphogluconate |
| PYR | pyruvate |
| R5P | ribose-5-phosphate |
| RU5P | ribulose-5-phosphate |
| S7P | sedoheptulose-7-phosphate |
| SUC | succinate |
| X5P | xylulose-5-phosphate |
| Ack | acetate kinase |
| Acs | acetyl coenzyme A synthetase |
| Adk | adenylate kinase |
| Ald | aldolase |
| CS | citrate synthase |
| Eno | enolase |
| Fum | fumarase |
| G6PDH | glucose-6-phosphate dehydrogenase |
| GAPDH | glyceraldehyde-3-phosphate dehydrogenase |
| ICDH | isocitrate dehydrogenase |
| Icl | isocitrate lyase |
| LDH | lactate dehydrogenase |
| MDH | malate dehydrogenase |
| Mez | malic enzyme |
| MS | malate synthase |
| Pck | phosphoenolpyruvate carboxykinase |
| PDH | pyruvate dehydrogenase |
| Pfk | phosphofructokinase |
| PGDH | 6-phosphogluconate dehydrogenase |
| Pgi | phosphoglucose isomerase |
| Pgk | phosphoglycerate kinase |
| Ppc | phosphoenolpyruvate carboxylase |
| Pps | phosphoenolpyruvate synthase |
| Pta | phosphotransacetylase |
| Pyk | pyruvate kinase |
| Ru5P | ribulose phosphate epimerase |
| R5PI | ribose phosphate isomerase |
| SDH | succinate dehydrogenase |
| SOD | superoxide dismutase |
| Tal | transaldolase |
| Tkt | transketolase |
Supplementary Files
Conflicts of Interest
References
- Seshasayee, A.S.; Bertone, P.; Fraser, G.M.; Luscombe, N.M. Transcriptional regulatory networks in bacteria: From input signals to output responses. Curr. Opin. Microbiol. 2006, 9, 511–519. [Google Scholar] [CrossRef]
- Kitano, H. Systems biology: A brief overview. Science 2002, 295, 1662–1664. [Google Scholar] [CrossRef]
- Kitano, H. Computational systems biology. Nature 2002, 420, 206–210. [Google Scholar] [CrossRef]
- Kotte, O.; Zaugg, J.B.; Heinemann, M. Bacterial adaptation through distributed sensing of metabolic fluxes. Mol. Syst. Biol. 2010, 6, 355. [Google Scholar]
- Kanehisa, M.; Goto, S.; Hattori, M.; Aoki-Kinoshita, K.F.; Itoh, M.; Kawashima, S.; Katayama, T.; Araki, M.; Hirakawa, M. From genomics to chemical genomics: New developments in KEGG. Nucleic Acids Res. 2006, 34, D354–D357. [Google Scholar] [CrossRef]
- Yamamoto, K.; Hirao, K.; Oshima, T.; Aiba, H.; Utsumi, R.; Ishihama, A. Functional characterization in vitro of all two component signal transduction systems from Escherichia coli. J. Biol. Chem. 2005, 280, 1448–1456. [Google Scholar]
- Maeda, H.; Fujita, N.; Ishihama, A. Competition among seven Escherichia coli sigma subunits: Relative binding affi nities to the core RNA polymerase. Nucleic Acids Res. 2000, 28, 3497–3503. [Google Scholar] [CrossRef]
- Timmermans, J.; van Melderen, L. Post-transcriptional global regulation by CsrA in bacteria. Cell. Mol. Life Sci. 2010, 67, 2897–2908. [Google Scholar] [CrossRef]
- Bruckner, R.; Titgemeyer, F. Carbon catabolite repression in bacteria: Choice of the carbon source and autoregulatory limitation of sugar utilization. FEMS Microbiol. Lett. 2002, 209, 141–148. [Google Scholar] [CrossRef]
- Gutierrez-RiosR, M.; Freyre-Gonzalez, J.A.; Resendis, O.; Collado-Vides, J.; Saier, M.; Gosset, G. Identification of regulatory network topological units coordinating the genome-wide transcriptional response to glucose in Escherichia coli. BMC Microbiol. 2007, 7, 53. [Google Scholar] [CrossRef]
- De Lay, N.; Gottesman, S. The crp-activated small noncoding regulatory RNA CyaR (RyeE) links nutritional status to group behavior. J. Bacteriol. 2009, 191, 461–476. [Google Scholar] [CrossRef]
- Muller, C.M.; Aberg, A.; Straseviciene, J.; Emody, L.; Uhlin, B.E.; Balsalobre, C. Type 1 fimbriae, a colonization factor of uropathogenic Escherichia coli, are controlled by the metabolic sensor CRP-cAMP. PLoS Pathog. 2009, 5, e1000303. [Google Scholar] [CrossRef]
- Zheng, D.; Constantinidou, C.; Hobman, J.L.; Minchin, S.D. Identification of the CRP regulon using in vitro and in vivo transcriptional profiling. Nucleic Acids Res. 2004, 32, 5874–5893. [Google Scholar] [CrossRef]
- Gottesman, S. Micros for microbes: Non-coding regulatory RNAs in bacteria. Trends Genet. 2005, 21, 399–404. [Google Scholar] [CrossRef]
- Vanderpool, C.K.; Gottesman, S. Involvement of a novel transcriptional activator and smallRNA in post-transcriptional regulation of the glucose phosphoenolpyruvate phosphotransferase system. Mol. Microbiol. 2004, 54, 1076–1089. [Google Scholar] [CrossRef]
- Babitzke, P.; Romeo, T. CsrB sRNA family: Sequestration of RNA-binding regulatory proteins. Curr. Opin. Microbiol. 2007, 10, 156–163. [Google Scholar] [CrossRef]
- Sauer, E.; Weichenrieder, O. Structural basis for RNA 3'-end recognition by Hfq. Proc. Natl. Acad. Sci. USA 2011, 108, 13065–13070. [Google Scholar] [CrossRef]
- Sauer, E.; Schmidt, S.; Weichenrieder, O. Small RNA binding to the lateral surface of Hfq hexamers and structural rearrangements upon mRNA target recognition. Proc. Natl. Acad. Sci. USA 2012, 109, 9396–9401. [Google Scholar] [CrossRef]
- Voge, J.; Luis, B.F. Hfq and its constellation of RNA. Nat. Rev. Microbiol. 2011, 9, 578–589. [Google Scholar] [CrossRef]
- Shimizu, K. Bacterial Cellular Metabolic Systems; Woodhead Publ. Co.: Oxford, UK, 2013. [Google Scholar]
- Shimizu, K. Metabolic regulation of a bacterial cell system with emphasis on Escherichia coli metabolism. ISRN Biochem. 2013, 645983:1–645983:47. [Google Scholar]
- Simon, S.A.; Gutknecht, J. Solubility of carbon dioxide in lipid bilayer membranes and organic solvents. Biochim. Biophys. Acta 1980, 596, 352–358. [Google Scholar] [CrossRef]
- Walter, A.; Gutknecht, J. Permeability of small nonelectrolytes through lipid bilayer membranes. J. Membr. Biol. 1986, 90, 207–217. [Google Scholar] [CrossRef]
- Nikaido, H.; Nakae, T. The outer membrane of gram-negative bacteria. Adv. Microb. Physiol. 1980, 20, 163–250. [Google Scholar] [CrossRef]
- De la Cruz, M.A.; Fernandez-Mora, M.; Guadarrama, C.; Flores-Valdez, M.A.; Bustamante, V.H.; Vázquez, A.; Calva, E. LeuO antagonizes H-NS and StpA-dependent repression in Salmonella entericaompS1. Mol. Microbiol. 2007, 66, 727–743. [Google Scholar] [CrossRef]
- Nikaido, H. Molecular basis of bacterial outer membrane permeability revisited. Microbiol. Mol. Biol. Rev. 2003, 67, 593–656. [Google Scholar] [CrossRef]
- Nikaido, H. Outer Membrane; Neidhardt, F.C., Ed.; ASM Press: Washington, DC, USA, 1996. [Google Scholar]
- Death, A.; Ferenci, T. Between feast and famine: Endogenous inducer synthesis in the adaptation of Escherichia coli to growth with limiting carbohydrates. J. Bacteriol. 1994, 176, 5101–5107. [Google Scholar]
- Von Meyenburg, K.; Nikaido, H. Outer membrane of gramnegative bacteria. XVII. Specificity of transport process catalyzed by the λ-receptor protein in Escherichia coli. Biochem. Biophys. Res. Commun. 1977, 78, 1100–1107. [Google Scholar] [CrossRef]
- Brzostek, K.; Brzostkowska, M.; Bukowska, I.; Karwicka, E.; Raczkowska, A. OmpR negatively regulates expression of invasion in Yersinia enterocolitica. Microbiology 2007, 153, 2416–2425. [Google Scholar] [CrossRef]
- Gunnewijk, M.G.W.; van den Bogaard, P.T.C.; Veenhoff, L.M.; Heuberger, E.H.; de Vos, W.M.; Kleerebezem, M.; Kuipers, O.P.; Poolman, B. Hierarchical control versus autoregulation of carbohydrate utilization in bacteria. J. Mol. Microbiol. Biotechnol. 2001, 3, 401–413. [Google Scholar]
- Postma, P.W.; Lengeler, J.W.; Jacobson, G.R. Phosphoenolpyruvate: Carbohydrate Phosphotransferase Systems. In Escherichia coli and Salmonella: Cellular and Molecular Biology; Neidhardt, F.C., Ed.; ASM Press: Washington, DC, USA, 1996; pp. 1149–1174. [Google Scholar]
- Tchieu, J.H.; Norris, V.; Edwards, J.S.; Saier, M.H., Jr. The complete phosphotransferase system in Escherichia coli. J. Mol. Microbiol. Biotechnol. 2001, 3, 329–346. [Google Scholar]
- Chou, C.H.; Bennett, G.N.; San, K.Y. Effect ofmodulated glucose uptake on high-level recombinant protein production in a dense Escherichia coli culture. Biotechnol. Prog. 1994, 10, 644–647. [Google Scholar] [CrossRef]
- Gosset, G. Improvement of Escherichia coli production strains by modification of the phosphoenolpyruvate: Sugar phosphotransferase system. Microb. Cell Fact. 2005, 4, 14. [Google Scholar] [CrossRef] [Green Version]
- Lunin, V.V.; Li, Y.; Schrag, J.D.; Iannuzzi, P.; Cygler, M.; Matte, A. Crystal structures of Escherichia coli ATP-dependent glucokinase and its complex with glucose. J. Bacteriol. 2004, 186, 6915–6927. [Google Scholar] [CrossRef]
- Huberts, D.H.; Niebel, B.; Heinemann, M. A flux-sensing mechanism could regulate the switch between respiration and fermentation. FEMS Yeast Res. 2012, 12, 118–128. [Google Scholar] [CrossRef]
- Kochanowski, K.; Volkmer, B.; Gerosa, L.; van Rijsewijk, H.B.R.; Schmidt, A.; Heinemann, M. Functioning of a metabolic flux sensor in Escherichia coli. Proc. Natl. Acad. Sci. USA 2013, 110, 1130–1135. [Google Scholar] [CrossRef]
- Valgepea, K.; Adamberg, K.; Nahku, R.; Lahtvee, P.J.; Arike, L.; Vilu, R. Systems biology approach reveals that overflow metabolism of acetate in Escherichia coli is triggered by carbon catabolite repression of acetyl-CoA synthetase. BMC Syst. Biol. 2010, 4, 166. [Google Scholar] [CrossRef]
- Valgepea, K.; Adamberg, K.; Vilu, R. Decrease of energy spilling in Escherichia coli continuous cultures with rising specific growth rate and carbon wasting. BMC Syst. Biol. 2011, 5, 106. [Google Scholar] [CrossRef]
- Kayser, A.; Weber, J.; Hecht, V.; Rinas, U. Metabolic flux analysis of Escherichia coli in glucose-limited continuous culture. I. Growth-rate dependent metabolic efficiency at steady state. Microbiology 2005, 151, 693–706. [Google Scholar]
- Nachen, A.; Schicker, A.; Revelles, O.; Sauer, U. Cyclic AMP-dependent catabolite repression is the dominant control mechanism of metabolic fluxes under glucose limitation in Escherichia coli. J. Biotechnol. 2008, 190, 2323–2330. [Google Scholar]
- Hua, Q.; Yang, C.; Baba, T.; Mori, H.; Shimizu, K. Responses of the central carbon metabolism in Escherichia coli to phosphoglucose isomerase and glucose-6-phosphate dehydrogenase knockouts. J. Bacteriol. 2003, 185, 7053–7067. [Google Scholar] [CrossRef]
- Bettenbrock, K.; Sauter, T.; Jahreis, K.; Klemling, A.; Lengeler, J.W.; Gilles, E.D. Correlation be-tween growth rates, EIIACrr phosphorylation, and intra-cellular cyclic AMP levels in Escherichia coli K-12. J. Bacteriol. 2007, 189, 6891–6900. [Google Scholar] [CrossRef]
- Hogema, B.M.; Arents, J.C.; Bader, R.; Eijkemans, K.; Yoshida, H.; Takahashi, H.; Aiba, H.; Postma, P.W. Inducer exclusion in Escherichia coli by non-PTS substrates: The role of the PEP to pyruvate ratio in deter-mining the phosphorylation state of enzyme IIAGlc. Mol. Microbiol. 1998, 30, 487–498. [Google Scholar] [CrossRef]
- Matsuoka, Y.; Shimizu, K. Catabolite regulation analysis of Escherichia coli for acetate overflow mechanism and co-consumption of multiple sugars based on systems biology approach using computer simulation. J. Biotechnol. 2013, 168, 155–173. [Google Scholar] [CrossRef]
- Yao, R.; Hirose, Y.; Sarkar, D.; Nakahigashi, K.; Ye, Q.; Shimizu, K. Catabolic regulation analysis of Escherichia coli and its crp, mlc, mgsA, pgi and ptsG mutants. Microb. Cell Fact. 2011, 10, 67. [Google Scholar] [CrossRef]
- Ishii, N.; Nakahigashi, K.; Baba, T.; Robert, M.; Soga, T.; Kanai, A.; Hirasawa, T.; Naba, M.; Hirai, K.; Hoque, A.; et al. Multiple high-throughput analyses monitor the response of E. coli to perturbations. Science 2007, 316, 593–597. [Google Scholar] [CrossRef]
- Brauer, M.J.; Yuan, J.; Bennett, B.D.; Lu, W.; Kimball, E.; Botstein, D.; Rabinowitz, J.D. Conservation of the metabolomic response to starvation across two divergent microbes. Proc. Natl. Acad. Sci. USA 2006, 103, 19302–19307. [Google Scholar] [CrossRef]
- Zampar, G.G.; Kümmel, A.; Ewald, J.; Jol, S.; Niebel, B.; Picotti, P.; Aebersold, R.; Sauer, U.; Zamboni, N.; Heinemann, M. Temporal system-level organization of the switch from glycolytic to gluconeogenic operation in yeast. Mol. Syst.Biol. 2013, 9, 651. [Google Scholar]
- Romeo, T. Global regulation by the small RNA-binding protein CsrA and the non- coding RNA molecule CsrB. Mol. Microbiol. 1998, 29, 1321–1330. [Google Scholar] [CrossRef]
- Jonas, K.; Edwards, A.N.; Ahmad, I.; Romeo, T.; Romling, U.; Melefors, O. Complex regulatory network encompassing the Csr, c-di-GMP and motility systems of Salmonella typhimurium. Environ. Microbiol. 2010, 12, 524–540. [Google Scholar] [CrossRef]
- Yakhnin, H.; Pandit, P.; Petty, T.J.; Baker, C.S.; Romeo, T.; Babitzke, P. CsrA of Bacillus subtilis regulates translation initiation of the gene encoding the flagellin protein (hag) by blocking ribosome binding. Mol. Microbiol. 2007, 64, 1605–1620. [Google Scholar] [CrossRef]
- Romeo, T.; Gong, M. Genetic and physical mapping of the regulatory gene csrA on the Escherichia coli K-12 chromosome. J. Bacteriol. 1993, 175, 5740–5741. [Google Scholar]
- Romeo, T.; Gong, M.; Liu, M.Y.; Brun-Zinkernagel, A.M. Identification and molecular characterization of csrA, a pleiotropic gene from Escherichia coli that affects glycogen biosynthesis, gluconeogenesis, cell size, and surface properties. J. Bacteriol. 1993, 175, 4744–4755. [Google Scholar]
- Romeo, T. Post-transcriptional regulation of bacterial carbohydratemetabolism: Evidence that the gene productCsrA is global mRNA decay factor. Res. Microbiol. 1996, 147, 505–512. [Google Scholar] [CrossRef]
- Dubey, A.K.; Baker, C.S.; Romeo, T.; Babitzke, P. RNA sequence and secondary structure participate in high-affinity CsrA-RNA interaction. RNA 2005, 11, 1579–1587. [Google Scholar] [CrossRef]
- Suzuki, K.; Wang, X.; Weilbacher, T.; Pernestig, A.K.; Melefors, O.; Georgellis, D.; Babitzke, P.; Romeo, T. Regulatory circuitry of the CsrA/CsrB and BarA/UvrY systems of Escherichia coli. J. Bacteriol. 2002, 184, 5130–5140. [Google Scholar] [CrossRef]
- Weilbacher, T.; Suzuki, K.; Dubey, A.K.; Wang, X.; Gudapaty, S.; Morozov, I.; Baker, C.S.; Georgellis, D.; Babitzke, P.; Romeo, T. A novel sRNA component of the carbon storage regulatory system of Escherichia coli. Mol. Microbiol. 2003, 48, 657–670. [Google Scholar] [CrossRef]
- Baker, C.S.; Morozov, I.; Suzuki, K.; Romeo, T.; Babitzke, P. CsrA regulates glycogen biosynthesis by preventing translation of glgC in Escherichia coli. Mol. Microbiol. 2002, 44, 1599–1610. [Google Scholar] [CrossRef]
- McKee, A.E.; Rutherford, B.J.; Chivian, D.C.; Baidoo, E.K.; Juminaga, D.; Kuo, D.; Benke, P.I.; Dietrich, J.A.; Ma, S.M.; Arkin, A.P.; et al. Manipulation of the carbon storage regulator system for metabolite remodeling and biofuel production in Escherichia coli. Microb. Cell Fact. 2012, 11, 79. [Google Scholar] [CrossRef]
- Tatarko, M.; Romeo, T. Disruption of a global regulatory gene to enhance CEntral carbon flux into Phenylalanine biosynthesis in Escherichia coli. Curr. Microbiol. 2001, 43, 26–32. [Google Scholar] [CrossRef]
- Edwards, A.N.; Patterson-Fortin, L.M.; Vakulskas, C.A.; Mercante, J.W.; Potrykus, K.; Vinella, D.; Camacho, M.I.; Fields, J.A.; Thompson, S.A.; Georgellis, D.; et al. Circuitry linking the Csr and stringent response global regulatory systems. Mol. Microbiol. 2011, 80, 1561–1580. [Google Scholar] [CrossRef]
- Yan, D. Protection of the glutamate pool concentration in enteric bacteria. Proc. Natl. Acad. Sci. USA 2007, 104, 9475–9480. [Google Scholar] [CrossRef]
- Ninfa, A.J.; Jiang, P.; Atkinson, M.R.; Peliska, J.A. Integration of antagonistic signals in the regulation of nitrogen assimilation in Escherichia coli. Curr. Topics Cell. Regul. 2000, 36, 31–75. [Google Scholar]
- Gruswitz, F.; O’Connell, J.; Stroud, R.M. Inhibitory complex of the transmembrane ammonia channel, AmtB, and the cytosolic regulatory protein, GlnK, at 1.96 A. Proc. Natl. Acad. Sci. USA 2007, 104, 42–47. [Google Scholar] [CrossRef]
- Radchenko, M.V.; Thornton, J.; Merrick, M. Control of AmtB-GlnK complex formation by intracellular levels of ATP, ADP, and 2-oxoglutarate. J. Biol. Chem. 2010, 285, 31037–31045. [Google Scholar] [CrossRef]
- Truan, D.; Huergo, L.F.; Chubatsu, L.S.; Merrick, M.; Li, X.D.; Winkler, F.K. A new P(II) protein structure identifies the 2-oxoglutarate binding site. J. Mol. Biol. 2010, 400, 531–539. [Google Scholar] [CrossRef]
- Kim, M.; Zhang, Z.; Okano, H.; Yan, D.; Groisman, A.; Hwa, T. Need-based activation of ammonium uptake in Escherichia coli. Mol. Syst. Biol. 2012, 8, 616. [Google Scholar]
- Yuan, J.; Doucette, C.D.; Fowler, W.U.; Feng, X.-J.; Piazza, M.; Rabitz, H.A.; Wingreen, N.S.; Rabinowitz, J.D. Metabolomics-driven quantitative analysis of ammonia assimilation in E. coli. Mol. Syst. Biol. 2009, 5, 302. [Google Scholar]
- Commichau, F.M.; Forchhammer, K.; Stulke, J. Regulatory links between carbon and nitrogen metabolism. Curr. Opin. Microbiol. 2006, 9, 167–172. [Google Scholar] [CrossRef]
- Mao, X.J.; Huo, Y.X.; Buck, M.; Kolb, A.; Wang, Y.P. Interplay between CRP-cAMP and PII-Ntr systems forms novel regulatory network between carbon metabolism and nitrogen assimilation in Escherichia coli. Nucleic Acids Res. 2007, 35, 1432–1440. [Google Scholar] [CrossRef]
- Jiang, P.; Ninfa, A.J. Escherichia coli PII signal transduction protein controlling nitrogen assimilation acts as a sensor of adenylate energy charge in vitro. Biochemistry 2007, 46, 12979–12996. [Google Scholar]
- Ninfa, J.; Jiang, P. PII signal transduction proteins: Sensors of α-ketoglutarate that regulate nitrogenmetabolism. Curr. Opin. Microbiol. 2005, 8, 168–173. [Google Scholar] [CrossRef]
- Hart, Y.; Madar, D.; Yuan, J.; Bren, A; Mayo, A.E.; Rabinowitz, J.D.; Alon, U. Robust control of nitrogen assimilation by a bifunctional enzyme in E. coli. Mol. Cell 2011, 41, 117–127. [Google Scholar] [CrossRef]
- Doucette, C.D.; Schwab, D.J.; Wingreen, N.S.; Rabinowitz, J.D. Alpha-ketoglutarate coordinates carbon and nitrogen utilization via enzyme I inhibition. Nat. Chem. Biol. 2011, 7, 894–901. [Google Scholar] [CrossRef]
- Powell, B.S.; Court, D.L.; Inada, T.; Nakamura, Y.; Michotey, V.; Cui, X.; Reizer, A.; Saier, M.H., Jr.; Reizer, J. Novel proteins of the phosphotransferase system encoded within the rpoN operon of Escherichia coli. Enzyme IIANtr affects growth on organic nitrogen and the conditional lethality of an erats mutant. J. Biol. Chem. 1995, 270, 4822–4839. [Google Scholar] [CrossRef]
- Peterkofski, A.; Wang, G.; Seok, Y.-J. Parallel PTS systems. Arch. Biochem. Biophys. 2006, 453, 101–107. [Google Scholar] [CrossRef]
- Pflüger-Grau, K.; Görke, B. Regulatory roles of the bacterial nitrogen-related phosphotransferase system. Trend Microbiol. 2010, 18, 205–214. [Google Scholar] [CrossRef]
- Lee, C.R.; Cho, S.H.; Yoon, M.J.; Peterkofsky, A.; Seok, Y.J. Escherichia coli enzyme IIANtr regulates the K+ transporter TrkA. Proc. Natl. Acad. Sci. USA 2007, 104, 4124–4129. [Google Scholar] [CrossRef]
- Lüttmann, D.; Heermann, R.; Zimmer, B.; Hillmann, A.; Rampp, I.S.; Jung, K.; Görke, B. Stimulation of the potassium sensor KdpD kinase activity by interaction with the phosphotransferase protein IIANtr in Escherichia coli. Mol. Micobiol. 2009, 72, 978–994. [Google Scholar] [CrossRef]
- Lee, C.-R.; Cho, S.-H.; Kim, H.-J.; Kim, M.; Peterkofsky, A.; Seok, Y.-J. Potassium mediates Escherichia coli enzyme IIANtr-dependent regulation of sigma factor selectivity. Mol. Microbiol. 2010, 78, 1468–1483. [Google Scholar] [CrossRef]
- Lüttmann, D.; Göpel, Y.; Görke, B. The phosphotransferase protein EIIANtr modulates the phosphate starvation response through interaction with histidine kinase PhoR in Escherichia coli. Mol. Microbiol. 2012, 86, 96–110. [Google Scholar] [CrossRef]
- Kim, H.-J.; Lee, C.-R.; Kim, M.; Peterkofsky, A.; Seok, Y.-J. Dephosphorylated NPr of the nitrogen PTS regulates lipid A biosynthesis by direct interaction with LpxD. Biochem. Biophys. Res. Commun. 2011, 409, 556–561. [Google Scholar] [CrossRef]
- Lee, C.-R.; Park, Y.-H.; Kim, M.; Kim, Y.-R.; Park, S.; Peterkofsky, A.; Seok, Y.-J. Reciprocal regulation of the autophosphorylation of enzyme INtr by glutamine and α-ketoglutarate in Escherichia coli. Mol. Microbiol. 2013, 88, 473–485. [Google Scholar] [CrossRef]
- Bykowski, T.; van der Ploeg, J.R.; Iwanicka-Nowicka, R.; Hryniewicz, M.M. The switch from inorganic to organic sulphur assimilation in Escherichia coli: Adenosine 5'-phosphosulphate (APS) as a signalling molecule for sulphate excess. Mol. Microbiol. 2002, 43, 1347–1358. [Google Scholar] [CrossRef]
- Kredich, N.M. Biosynthesis of Cysteine. In Escherichia coli and Salmonella: Cellular and Molecular Biology, 2nd ed.; Neidhardt, F.C., Curtiss, R., III, Ingraham, J.L., Lin, E.C.C., Low, K.B., Magasanik, B., Reznikoff, W.S., Riley, M., Schaechter, M., Umbarger, H.H., Eds.; ASM Press: Washington, DC, USA, 1996; Volume 1, pp. 514–527. [Google Scholar]
- Iwanicka-Nowicka, R.; Hryniewicz, M.M. A new gene, cbl, encoding a member of the LysR family of transcriptional regulators belongs to Escherichia coli cys regulon. Gene 1995, 166, 11–17. [Google Scholar] [CrossRef]
- Zimmer, D.P.; Soupene, H.L.E.; Lee, V.F.; Wendisch, A.B.; Khodursky, B.J.; Peter, R.; Bender, A.; Kustu, S. Nitrogen regulatory protein Ccontrolled genes of Escherichia coli: Scavenging as a defense against nitrogen limitation. Proc. Natl. Acad. Sci. USA 2000, 97, 14674–14679. [Google Scholar] [CrossRef]
- Gyaneshwar, P.; Paliy, O.; McAuliffe, J.; Popham, D.L.; Jordan, M.I.; Kustu, S. Sulfur and nitrogen limitation in Escherichia coli K-12: Specific homeostatic responses. J. Bacteriol. 2005, 187, 1074–1090. [Google Scholar] [CrossRef]
- Wanner, B.L. Phosphorus Assimilation and Control of the Phosphate Regulon. In Escherichia Coli and Salmonella: Cellular and Molecular Biology; Neidhardt, F.C., Curtiss, R., III, Ingraham, J.L., Eds.; ASM Press: Washington, DC, USA, 1996; pp. 1357–1381. [Google Scholar]
- Wanner, B.L. Gene regulation by phosphate in enteric bacteria. J. Cell. Biochem. 1993, 51, 47–54. [Google Scholar] [CrossRef]
- Baek, J.H.; Lee, S.Y. Transcriptome analysis of phosphate starvation response in Escherichia coli. J. Microbiol. Biotechnol. 2007, 17, 244–252. [Google Scholar]
- Ruiz, N.; Silhavy, T.J. Constitutive activation of the Escherichia coli Pho regulon upregulates rpoS translation in an Hfq dependent fashion. J. Bacteriol. 2003, 185, 5984–5992. [Google Scholar] [CrossRef]
- Taschner, N.P.; Yagil, E.; Spira, B. A differential effect of σs on the expression of the PHO regulon genes of Escherichia coli. Microbiology 2004, 150, 2985–2992. [Google Scholar] [CrossRef]
- Spira, B.; Silberstein, N.; Yagil, E. Guanosine 3',5'-bispyrophosphate (ppGpp) synthesis in cells of Escherichia coli starved for Pi. J. Bacteriol. 1995, 177, 4053–4058. [Google Scholar]
- Spira, B.; Yagil, E. The integration host factor (IHF) affects the expression of the phosphate-binding protein and of alkaline phosphatase in Escherichia coli. Curr. Microbiol. 1999, 38, 80–85. [Google Scholar] [CrossRef]
- Taschner, N.P.; Yagil, E.; Spira, B. The effect of IHF on σs selectivity of the phoA and pst promoters of Escherichia coli. Arch. Microbiol. 2006, 185, 234–237. [Google Scholar] [CrossRef]
- Schurdell, M.S.; Woodbury, G.M.; McCleary, W.R. Genetic evidence suggests that the intergenic region between pstA and pstB plays a role in the regulation of rpoS translation during phosphate limitation. J. Bacteriol. 2007, 189, 1150–1153. [Google Scholar] [CrossRef]
- Wanner, B.L.; Wilmes, M.R.; Young, D.C. Control of bacterial alkaline phosphatase synthesis and variation in an Escherichia coli K-12 phoR mutant by adenyl cyclase, the cyclic AMP receptor protein, and the phoM operon. J. Bacteriol. 1988, 170, 1092–1102. [Google Scholar]
- Kang, Y.; Weber, K.D.; Qiu, Y.; Kiley, P.J.; Blattner, F.R. Genome-wide expression analysis indicates that FNR of Escherichia coli K-12 regulates a large number of genes of unknown function. J. Bacteriol. 2005, 187, 1135–1160. [Google Scholar] [CrossRef]
- Dann, C.E., III; Wakeman, C.A.; Sieling, C.L.; Baker, S.C.; Irnov, I.; Winker, W.C. Structure and mechanism of a metal-sensing regulatory RNA. Cell 2007, 130, 878–892. [Google Scholar] [CrossRef]
- Helmann, J.D. Measuring metals with RNA. Mol. Cell. 2007, 27, 859–860. [Google Scholar] [CrossRef]
- Zheng, M.; Doan, B.; Schneider, T.D.; Storz, G. OxyR and SoxRS regulation of fur. J. Biotechnol. 1999, 181, 4639–4643. [Google Scholar]
- McHugh, J.P.; Rodríguez-Quiñones, F.; Abdul-Tehrani, H.; Svistunenko, D.A.; Poole, R.K.; Cooper, C.E.; Andrews, S.C. Global iron-dependent gene regulation in Escherichia coli. J. Biol. Chem. 2003, 278, 29478–29486. [Google Scholar] [CrossRef]
- Storz, G.; Imlay, J.A. Oxidative stress. Curr. Opin. Microbiol. 1999, 2, 188–194. [Google Scholar] [CrossRef]
- Gennis, R.B.; Stewart, V. Respiration in Escherichia coli and Salmonella: Cellular and Molecular Biology; Bock, A., Curtiss, R., III, Kaper, J.B., Neidhardt, F.C., Nystrom, T., Rudd, K.E., Squires, C.L., Eds.; ASM Press: Washington, DC, USA, 1996. [Google Scholar]
- Jovanovic, G.; Lloyde, L.J.; Stumpf, M.P.; Mayhew, A.J.; Buck, M. Induction and function of the phase shock protein extracytoplasmic stress response in Escherichia coli. J. Biol. Chem. 2006, 281, 21147–21161. [Google Scholar]
- Blanchard, J.R.; Wholey, W.Y.; Conlon, E.M.; Pomposiello, P.J. Rapid changes in gene expression dynamics in response to superoxide reveal SoxRS dependent and independent transcriptional network. PLoS One 2007, 2, e1186. [Google Scholar] [CrossRef]
- Brynildsen, M.P.; Liao, J.C. An integrated network approach indentifies the isobutanol response network of Escherichia coli. Mol. Syst. Biol. 2009, 5, 277. [Google Scholar]
- Braun, V. Regulation of iron uptake minimizes iron-mediated oxidative stress. J. Biosci. 1998, 23, 483–489. [Google Scholar] [CrossRef]
- Azpiroz, M.F.; Lavińa, M. Involvement of enterobactin synthesis pathway in production of Microcin H47. Antimicrob. Agents Chemother. 2004, 48, 1235–1241. [Google Scholar] [CrossRef]
- Semsey, S.; Andersson, A.M.; Krishna, S.; Jensen, M.H.; Masse, E.; Sneppen, K. Genetic regulation of fluxes: Iron homeostasis of Escherichia coli. Nucleic Acids Res. 2006, 34, 4960–4967. [Google Scholar] [CrossRef]
- Hantash, F.M.; Ammerlaan, M.; Earhart, C.F. Enterobactin synthase polypeptides of Escherichia coli are present in an osmotic-shocksensitive cytoplasmic locality. Microbiology 1997, 143, 147–156. [Google Scholar] [CrossRef]
- Kumar, R.; Shimizu, K. Transcriptional regulation of main metabolic pathways of cyoA, cydB, fnr, and fur gene knockout Escherichia coli in C-limited and N-limited aerobic continuous cultures. Microb. Cell Fact. 2011, 10, 3. [Google Scholar] [CrossRef]
- Zhang, Z.; Gosset, G.; Barabote, R.; Gonzalez, C.S.; Cuevas, W.A.; Saier, M.H., Jr. Functional interactions between the carbon and iron utilization regulators, Crp and Fur, in Escherichia coli. J. Bacteriol. 2005, 187, 980–990. [Google Scholar] [CrossRef]
- Ali Azam, T.; Iwata, A.; Nishimura, A.; Ueda, S.; Ishihama, A. Growth phase-dependent variation in protein composition of the Escherichia coli nucleoid. J. Bacteriol. 1999, 181, 6361–6370. [Google Scholar]
- Bradley, M.D.; Beach, M.B.; de Koning, J.A.P.; Pratt, T.S.; Osuna, R. Effects of Fis on Escherichia coli gene expression during different growth stages. Microbiology 2007, 153, 2922–2940. [Google Scholar] [CrossRef]
- Mallik, P.; Pratt, T.S.; Beach, M.B.; Bradley, M.D.; Undamatla, J.; Osuna, R. Growth phase-dependent regulation and stringent control of fis are conserved processes in enteric bacteria and involve a single promoter (fis P) in Escherichia coli. J. Bacteriol. 2004, 186, 122–135. [Google Scholar] [CrossRef]
- Mallik, P.; Paul, B.J.; Rutherford, S.T.; Gourse, R.L.; Osuna, R. DksA is required for growth phase-dependent regulation, growth rate-dependent control, and stringent control of fis expression in Escherichia coli. J. Bacteriol. 2006, 188, 5775–5782. [Google Scholar] [CrossRef]
- Paul, B.J.; Berkmen, M.B.; Gourse, R.L. DksA potentiates direct activation of amino acid promoters by ppGpp. Proc. Natl. Acad. Sci. USA 2005, 102, 7823–7828. [Google Scholar] [CrossRef]
- Ferenci, T. Hungry bacteria—definition and properties of a nutritional state. Environ. Microbiol. 2001, 3, 605–611. [Google Scholar] [CrossRef]
- Hardiman, T.; Lemuth, K.; Keller, M.A.; Reuss, M.; Siemann-Herzberg, M. Topology of the global regulatory network of carbon limitation in Escherichia coli. J. Biotechnol. 2007, 132, 359–374. [Google Scholar] [CrossRef]
- Braeken, K.; Moris, M.; Daniels, R.; Vanderleyden, J.; Michiels, J. New horizons for (p)ppGpp in bacterial and plant physiology. Trends Microbiol. 2006, 14, 45–54. [Google Scholar] [CrossRef]
- Artsimovitch, I.; Patlan, V.; Sekine, S.; Vassylyeva, M.N.; Hosaka, T.; Ochi, K.; Yokoyama, S. Structural basis for transcription regulation by alarmone ppGpp. Cell 2004, 117, 299–310. [Google Scholar] [CrossRef]
- Kanjee, U.; Ogata, K.; Houry, W.A. Direct binding targets of the stringent response alarmone(p)ppGpp. Mol. Micobiol. 2012, 85, 1029–1043. [Google Scholar] [CrossRef]
- Hengge-Aronis, R. Signal transduction and regulatory mechanisms involved in control of the σs (RpoS) subunit of RNA polymerase. Microbiol. Mol. Biol. Rev. 2002, 66, 373–395. [Google Scholar] [CrossRef]
- Patten, C.L.; Kirchhof, M.G.; Schertzberg, M.R.; Morton, R.A.; Schellhorn, H.E. Microarray analysis of RpoS-mediated gene expression in Escherichia coli K-12. Mol. Genet. Genomics 2004, 272, 580–591. [Google Scholar] [CrossRef]
- Jishage, M.; Kvint, K.; Shingler, V.; Nystrom, T. Regulation of σfactor competition by the alarmone ppGpp. Genes Dev. 2002, 16, 1260–1270. [Google Scholar] [CrossRef]
- Aronis, H.R. Regulation of Gene Expression during Entry into Stationary Phase. In Escherichia Coli and Salmonella: Cellular and Molecular Biology, 2nd ed.; Neidhardt, F.C., Curtiss, R., III, Ingram, J.L., Eds.; ASM Press: Washington, DC, USA, 1996; pp. 1497–1512. [Google Scholar]
- Aronis, H.R. Stationary phase gene regulation: What makes an Escherichia coli promoter σs-selective? Curr. Opin. Microbiol. 2002, 5, 591–595. [Google Scholar] [CrossRef]
- Lacour, S.; Landini, P. σs-dependent gene expression at the onset of stationary phase in Escherichia coli: Function of σs-dependent genes and identification of their promoter sequences. J. Bacteriol. 2004, 186, 7186–7195. [Google Scholar] [CrossRef]
- Vijayakumar, S.R.V.; Kirchhof, M.G.; Patten, C.L.; Schellhorn, H.E. RpoS-regulated genes of Escherichia coli identified by random lacZ fusion mutagenesis. J. Bacteriol. 2004, 186, 8499–8507. [Google Scholar] [CrossRef]
- Wei, B.; Shin, S.; LaPorte, D.; Wolfe, A.J.; Romeo, T. Global regulatory mutations in csrA and rpoS cause severe central carbon stress in Escherichia coli in the presence of acetate. J. Bacteriol. 2000, 182, 1632–1640. [Google Scholar] [CrossRef]
- Rahman, M.; Hasan, M.R.; Oba, T.; Shimizu, K. Effect of rpoS gene knockout on the metabolism of Escherichia coli during exponential growth phase and early stationary phase based on gene expressions, enzyme activities and intracellular metabolite concentrations. Biotech. Bioeng. 2006, 94, 585–595. [Google Scholar] [CrossRef]
- Weber, H.; Polen, T.; Heuveling, J.; Wendisch, V.F.; Hengge, R. Genome-wide analysis of the general stress response network in Escherichia coli: σs-Dependent genes, promoters, and sigma factor selectivity. J. Bacteriol. 2005, 187, 1591–1603. [Google Scholar] [CrossRef]
- Wang, X.; Dubey, A.K.; Suzuki, K.; Baker, C.S.; Babitzke, P.; Romeo, T. CsrA post-transcriptionally represses pgaABCD, responsible for synthesis of a biofilm polysaccharide adhesin of Escherichia coli. Mol. Microbiol. 2005, 56, 1648–1663. [Google Scholar] [CrossRef]
- Hengge, R. Principles of c-di-GMP signalling in bacteria. Nat. Rev. Microbiol. 2009, 7, 263–273. [Google Scholar] [CrossRef]
- Barnhart, M.M.; Chapman, M.R. Curli biogenesis and function. Annu. Rev. Microbiol. 2006, 60, 131–147. [Google Scholar] [CrossRef]
- Thomason, M.K.; FFontaine, F.; de Lay, N.; Storz1, G. A small RNA that regulates motility and biofilm formation in response to changes in nutrient availability in Escherichia coli. Mol. Microbiol. 2012, 84, 17–35. [Google Scholar] [CrossRef]
- Dong, T.; Schellhorn, H.E. Control of RpoS in global gene expression of Escherichia coli in minimal media. Mol. Genet. Genomics 2009, 281, 19–33. [Google Scholar] [CrossRef]
- Jackson, D.W.; Simecka, J.W.; Romeo, T. Catabolite repression of Escherichia coli biofilm formation. J. Bacteriol. 2002, 184, 3406–3410. [Google Scholar] [CrossRef]
- Waters, C.M.; Bassler, B.L. Quorum sensing: Cell-to-cell communication in bacteria. Annu. Rev. Cell Dev. Biol. 2005, 21, 319–346. [Google Scholar] [CrossRef]
- Llorens, N.J.M.; Tormo, A.; Martínez-García, E. Stationary phase in gram-negative bacteria. FEMS Microbiol. Rev. 2010, 34, 476–495. [Google Scholar] [CrossRef]
- Desnues, B.; Cuny, C.; Gregori, G.; Dukan, S.; Aguilaniu, H.; Nyström, T. Differential oxidative damage and expression of stress defence regulons in culturable and non-culturable Escherichia coli cells. EMBO Rep. 2003, 4, 400–404. [Google Scholar] [CrossRef]
- Nagamitsu, H.; Murata, M.; Kosaka, T.; Kawaguchi, J.; Mori, H.; Yamada, M. Crucial roles of MicA and RybB as vital factors for σE-dependent cell lysis in Escherichia coli long-term stationary phase. J. Mol. Microbiol. Biotechnol. 2013, 23, 227–232. [Google Scholar] [CrossRef]
- Murata, M.; Noor, R.; Nagamitsu, H.; Tanaka, S.; Yamada, M. Novel pathway directed by sigma E to cause cell lysis in Escherichia coli. Genes Cells 2012, 17, 234–247. [Google Scholar] [CrossRef]
- Noor, R.; Murata, M.; Yamada, M. Oxidative stress as a trigger for growth phase-specific sigmaE-dependent cell lysis in Escherichia coli. J. Mol. Microbiol. Biotechnol. 2009, 17, 177–187. [Google Scholar] [CrossRef]
- Pomposiello, P.J.; Demple, B. Redox-operated genetic switches: The SoxR and OxyR transcription factors. Trends Biotechnol. 2001, 19, 109–114. [Google Scholar] [CrossRef]
- Gaudu, P.; Weiss, B. SoxR, a [2Fe-2S] transcription factor, is active only in its oxidized form. Proc. Natl. Acad. Sci. USA 1996, 93, 10094–10098. [Google Scholar] [CrossRef]
- Liochev, S.I.; Fridovich, I. The role of O2. -in the production of HO.: In vitro and in vivo. Free Radic. Biol. Med. 1994, 16, 29–33. [Google Scholar] [CrossRef]
- Greenberg, J.T.; Monach, P.; Chou, J.H.; Josephy, P.D.; Demple, B. Positive control of a global antioxidant defense regulon activated by superoxide-generating agents in Escherichia coli. Proc. Natl. Acad. Sci. USA 1990, 87, 6181–6185. [Google Scholar] [CrossRef]
- Kabir, M.M.; Shimizu, K. Investigation into the effect of soxR and soxS genes deletion on the central metabolism of Escherichia coli based on gene expressions and enzyme activities. Biochem. Eng. J. 2006, 30, 39–47. [Google Scholar] [CrossRef]
- Wolf, R.E.; Prather, D.M.; Shea, F.M. Growth-rate dependent alteration of 6-phosphogluconate dehydrogenase and glucose 6-phosphate dehydrogenase levels in Escherichia coli K-12. J. Bacteriol. 1979, 139, 1093–1096. [Google Scholar]
- Tsaneva, I.R.; Weiss, B. soxR, a locus governing a superoxide response regulon in Escherichia coli K-12. J. Bacteriol. 1990, 172, 4197–4205. [Google Scholar]
- Hanson, R.L.; Rose, C. Effects of an insertion mutation in a locus affecting pyridine nucleotide transhydrogenase (pnt::Tn5) on the growth of Escherichia coli. J. Bacteriol. 1980, 141, 401–404. [Google Scholar]
- Mailloux, R.J.; Bériault, R.; Lemire, J.; Singh, R.; Chénier, R.R.; Hamel, R.D.; Appanna, V.D. The tricarboxylic acid cycle, an ancient metabolic network with a novel twist. PLoS One 2007, 2, e690. [Google Scholar] [CrossRef]
- Johnson, P. Antioxidant enzyme expression in health and disease: Effects of exercise and hypertension. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2002, 133, 493–505. [Google Scholar] [CrossRef]
- Foster, J.W. Escherichia coli acid resistance: Tales of an amateur acidophile. Nat. Rev. Microbiol. 2004, 2, 898–907. [Google Scholar]
- Stincone, A.; Daudi, N.; Rahman, A.S.; Antczak, P.; Henderson, I.; Cole, J.; Johnson, M.D.; Lund, P.; Falciani, F. A systems biology approach sheds new light on Escherichia coli acid resistance. Nucleic Acids Res. 2011, 39, 7512–7528. [Google Scholar] [CrossRef]
- Richard, H.T.; Foster, J.W. Acid resistance in Escherichia coli. Adv. Appl. Microbiol. 2003, 52, 167–186. [Google Scholar] [CrossRef]
- Richard, H.T.; Foster, J.W. Escherichia coli glutamate- and arginine- dependent acid resistance systems increase internal pH and reverse transmembrane potential. J. Bacteriol. 2004, 86, 6032–6041. [Google Scholar] [CrossRef]
- Gong, S.; Richard, H.; Foster, J.W. YjdE (AdiC) is the arginine: Agmatine antiporter essential for arginine-dependent acid resistance in Escherichia coli. J. Bacteriol. 2003, 185, 4402–4409. [Google Scholar] [CrossRef]
- Iyer, R.; Williams, C.; Miller, C. Arginin-agmatine antiporter in extreme acid resistance in Escherichia coli. J. Bacteriol. 2003, 185, 6556–6561. [Google Scholar] [CrossRef]
- Castanie-Cornet, M.P.; Foster, J.W. Escherichia coli acid resistance: cAMP receptor protein and a 20 bp cis-acting sequence control pH and stationary phase expression of the gadA and gadBC glutamate decarboxylase genes. Microbiology 2001, 147, 709–715. [Google Scholar]
- Martin-Galiano, A.J.; Ferrandiz, M.J.; de La Campa, A.G. The promoter of the operon encoding the F0F1 ATPase of Streptococcus pneumonia is inducible by pH. Mol. Microbiol. 2001, 41, 327–338. [Google Scholar]
- Marzan, L.W.; Shimizu, K. Metabolic regulation of Escherichia coli and its phoB and phoR genes knockout mutants under phosphate and nitrogen limitations as well as at acidic condition. Microb. Cell Fact. 2011, 10, 39. [Google Scholar] [CrossRef]
- Suziedeliene, E.; Suziedelis, K.; Garbenciute, V.; Normark, S. The acid-inducible asr gene in Escherichia coli: Transcriptional control by the phoBR operon. J. Bacteriol. 1999, 181, 2084–2093. [Google Scholar]
- Kitagawa, M.; Miyakawa, M.; Matsumura, Y.; Tsuchido, T. Escherichia coli small heat shock proteins, IbpA and IbpB, protect enzymes from inactivation by heat and oxidants. Eur. J. Biochem. 2002, 269, 2907–2917. [Google Scholar] [CrossRef]
- Sørensen, H.P.; Mortensen, K.K. Soluble expression of recombinant proteins in the cytoplasm of Escherichia coli. Microb. Cell Fact. 2005, 4, 1. [Google Scholar] [CrossRef]
- Hoffmann, F.; Weber, J.; Rinas, U. Metabolic adaptation of Escherichia coli during temperature-induced recombinant protein production: 1. Readjustment of metabolic enzyme synthesis. Biotechnol. Bioeng. 2002, 80, 313–319. [Google Scholar] [CrossRef]
- Tilly, K.; Erickson, J.; Sharma, S.; Georgopoulos, C. Heat shock regulatory gene rpoH mRNA level increases after heat shock in Escherichia coli. J. Bacteriol. 1986, 168, 1155–1158. [Google Scholar]
- Tilly, K.; Spence, J.; Georgopoulos, C. Modulation of stability of the Escherichia coli heat shock regulatory factor σ32. J. Bacteriol. 1989, 171, 1585–1589. [Google Scholar]
- Shin, D.; Lim, S.; Seok, Y.J.; Ryu, S. Heat shock RNA polymerase (Eσ32) is involved in the transcription of mlc and Crucial for Induction of the Mlc Regulon by Glucose in Escherichia coli. J. Biol. Chem. 2001, 276, 25871–25875. [Google Scholar] [CrossRef]
- Kumari, S.; Beatty, C.M.; Browning, D.F.; Busby, S.J.; Simel, E.J.; Hovel-Miner, G.; Wolfe, A.J. Regulation of acetyl coenzyme A synthetase in Escherichia coli. J. Bacteriol. 2000, 182, 4173–4179. [Google Scholar] [CrossRef]
- Browning, D.F.; Beatty, C.M.; Wolfe, A.J.; Cole, J.A.; Busby, S.J.W. Independent regulation of the divergent Escherichia coli nrfA and acsP1 promoters by a nucleoprotein assembly at a shared regulatory region. Mol. Microbiol. 2002, 43, 687–701. [Google Scholar] [CrossRef]
- Beatty, C.M.; Browning, D.F.; Busby, S.J.W.; Wolfe, A.J. Cyclic AMP receptor protein-dependentactivation of the Escherichia coliacsP2 promoter by a synergistic class III mechanism. J. Bacteriol. 2003, 185, 5148–5157. [Google Scholar] [CrossRef]
- Browning, D.F.; Beatty, C.M.; Sanstad, E.A.; Gunn, K.E.; Busby, S.J.W.; Wolfe, A.J. Modulation of CRP-dependent transcription at the Escherichia coliacsP2 promoter by nucleoprotein complexes: Anti-activation by the nucleoid proteins FIS and IHF. Mol. Microbiol. 2004, 51, 241–254. [Google Scholar]
- Rahman, M.; Shimizu, K. Altered acetate metabolism and biomass production in several Escherichia coli mutants lacking rpoS-dependent metabolic pathway genes. Mol. Biosyst. 2008, 4, 160–169. [Google Scholar] [CrossRef]
- Privalle, C.T.; Fridovich, I. Induction of superoxide dismutase in Escherichia coli by heat shock. Proc. Natl. Acad. Sci. USA 1987, 84, 2723–2726. [Google Scholar] [CrossRef]
- Dunlop, M.J. Engineering microbes for tolerance to next-generation biofuels. Biotechnol. Biofuels. 2011, 4, 32. [Google Scholar] [CrossRef]
- Isken, S.; de Bont, J.A.M. Bacteria tolerant to organic solvents. Extremophiles 1998, 2, 229–238. [Google Scholar] [CrossRef]
- Ramos, J.L.; Duque, E.; Gallegos, M.-T.; Godoy, P.; Ramos-Gonzalez, M.I.; Rojas, A.; Teran, W.; Segura, A. Mechanisms of solvent tolerance in Gram-negative bacteria. Annu. Rev. Microbiol. 2002, 56, 743–768. [Google Scholar] [CrossRef]
- Nicolaou, S.A.; Gaida, S.M.; Papoutsakis, E.T. A comparative view of metabolite and substrate stress and tolerance in microbial bioprocessing: From biofuels and chemicals, to biocatalysis and bioremediation. Metab. Eng. 2010, 12, 307–331. [Google Scholar] [CrossRef]
- Takatsuka, Y.; Chen, C.; Nikaido, H. Mechanism of recognition of compounds of diverse structures by the multidrug efflux pump AcrB of Escherichia coli. Proc. Natl. Acad. Sci. USA 2010, 107, 6559–6656. [Google Scholar] [CrossRef]
- Ankarloo, J.; Wikman, S.; Nicholls, I.A. Escherichia coli mar and acrAB mutants display no tolerance to simple alcohols. Int. J. Mol. Sci. 2010, 11, 1403–1412. [Google Scholar] [CrossRef]
- Piper, P. The heat-shock and ethanol stress responses of yeast exhibit extensive similarity and functional overlap. FEMS Microbiol. Lett. 1995, 134, 121–127. [Google Scholar] [CrossRef]
- Rutherford, B.J.; Dahl, R.H.; Price, R.E.; Szmidt, H.L.; Benke, P.I.; Mukhopadhyay, A.; Keasling, J.D. Functional genomic study of exogenous n-butanol stress in Escherichia coli. Appl. Environ. Microbiol. 2010, 76, 1935–1945. [Google Scholar] [CrossRef]
- Tomas, C.; Beamish, J.; Papoutsakis, E. Transcriptional analysis of butanol stress and tolerance in Clostridium acetobutylicum. J. Bacteriol. 2004, 186, 2006–2018. [Google Scholar] [CrossRef]
- Fiocco, D.; Capozzi, V.; Goffin, P.; Hols, P.; Spano, G. Improved adaptation to heat, cold, and solvent tolerance in Lactobacillus plantarum. Appl. Microbiol. Biotechnol. 2007, 77, 909–915. [Google Scholar] [CrossRef]
- Reyes, L.H.; Almario, M.P.; Kao, K.C. Genomic library screens for genes involved in n-butanol tolerance in Escherichia coli. PLoS One 2011, 6, e17678. [Google Scholar] [CrossRef]
- Sikkema, J.; de Bont, J.A.M.; Poolman, B. Mechanisms of membrane toxicity of hydrocarbons. Microbiol. Rev. 1995, 59, 201–222. [Google Scholar]
- Holtwick, R.; Meinhardt, F.; Keweloh, H. Cis-trans isomerization of unsaturated fatty acids: Cloning and sequencing of the cti gene from Pseudomonas putida P8. Appl. Environ. Microb. 1997, 63, 4292–4297. [Google Scholar]
- Kiran, M.; Prakash, J.; Annapoorni, S.; Dube, S.; Kusano, T.; Okuyama, H.; Murata, N.; Shivaji, S. Psychrophilic Pseudomonas syringae requires transmonounsaturated fatty acid for growth at higher temperature. Extremophiles 2004, 8, 401–410. [Google Scholar] [CrossRef]
- Wu, X.; Altman, R.; Eiteman, M.A.; Altman, E. Effect of overexpressing nhaA and nhaR on sodium tolerance and lactate production in Escherichia coli. J. Biol. Eng. 2013, 7, 3. [Google Scholar] [CrossRef]
- Minty, J.J.; Lesnefsky, A.A.; Lin, F.; Chen, Y.; Zaroff, T.A.; Velose, A.B.; Xie, B.; McConnell, C.A.; Ward, R.J.; Schwartz, D.R.; et al. Evolution combined with genomic study elucidates genetic bases of isobutanol tolerance in Escherichia coli. Microb. Cell Fact. 2011, 10, 18. [Google Scholar] [CrossRef]
- Gutierrez-Rios, R.M.; Rosenblueth, D.A.; Loza, J.A.; Huerta, A.M.; Glasner, J.D.; Blattner, F.R.; Collado-Vides, J. Regulatory network of Escherichia coli: Consistency between literature knowledge and microarray profiles. Genome Res. 2003, 13, 2435–2443. [Google Scholar] [CrossRef]
- Maheswaran, M.; Forchhammer, K. Carbon-source-dependent nitrogen regulation in Escherichia coli is mediated through glutamine-dependent GlnB signaling. Microbiology 2003, 149, 2163–2172. [Google Scholar] [CrossRef]
- Quan, J.A.; Schneider, B.L.; Paulsen, I.T.; Yamada, M.; Kredich, N.M.; Saier, M.H., Jr. Regulationof carbon utilization by sulfur availability in Escherichia coli and Salmonella typhimurium. Microbiology 2002, 148, 123–131. [Google Scholar]
- De Lorenzo, V.; Herrero, M.; Giovannini, F.; Neilands, J.B. Fur (ferric uptake regulation) protein and CAP (catabolite-activator protein) modulate transcription of fur gene in Escherichia coli. Eur. J. Biochem. 1998, 173, 537–546. [Google Scholar]
- Dragosits, M.; Mozhayskiy, V.; Quinones-Soto, S.; Park, J.; Tagkopoulos, I. Evolutionary potential, cross-stress behavior and the genetic basis of acquired stress resistance in Escherichia coli. Mol. Syst. Biol. 2013, 9, 643. [Google Scholar]
- Jenkins, D.; Schultz, J.; Matin, A. Starvation-induced cross protection against heat or H2O2 challenge in Escherichia coli. J. Bacteriol. 1988, 170, 3910–3914. [Google Scholar]
- Jenkins, D.; Auger, E.; Matin, A. Role of RpoH, a heat shock regulator protein, in Escherichia coli carbon starvation protein synthesis and survival. J. Bacteriol. 1991, 173, 1992–1996. [Google Scholar]
- Gunasekera, T.; Csonka, L.; Paliy, O. Genome-wide transcriptional responses of Escherichia coli K-12 to continuous osmotic and heat stresses. J. Bacteriol. 2008, 190, 3712–3720. [Google Scholar] [CrossRef]
- White-Ziegler, C.; Um, S.; Pérez, N.; Berns, A.; Malhowski, A.; Young, S. Low temperature (23 degrees C) increases expression of biofilm-, cold-shock- and RpoS-dependent genes in Escherichia coli K-12. Microbiology 2008, 154, 148–166. [Google Scholar] [CrossRef]
- Reaves, M.L.; Rabinowitz, J.D. Metabolomics in systems microbiology Review Article. Curr. Opin. Biotechnol. 2011, 22, 17–25. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Shimizu, K. Regulation Systems of Bacteria such as Escherichia coli in Response to Nutrient Limitation and Environmental Stresses. Metabolites 2014, 4, 1-35. https://doi.org/10.3390/metabo4010001
Shimizu K. Regulation Systems of Bacteria such as Escherichia coli in Response to Nutrient Limitation and Environmental Stresses. Metabolites. 2014; 4(1):1-35. https://doi.org/10.3390/metabo4010001
Chicago/Turabian StyleShimizu, Kazuyuki. 2014. "Regulation Systems of Bacteria such as Escherichia coli in Response to Nutrient Limitation and Environmental Stresses" Metabolites 4, no. 1: 1-35. https://doi.org/10.3390/metabo4010001