Cattle Breeds: Extinction or Quasi-Extant?


Abstract
:1. Introduction
1.1. The Breed Concept
1.2. Status of Cattle Genetic Diversity
2. Materials and Methods
2.1. Cattle Genetic Resources, Status and Circumstances

{kind=link}
| Status | Local breeds | Transboundary breeds (Regional) | Transboundary breeds (International) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sweden | Uganda | Brazil | Bangladesh | Sweden | Uganda | Brazil | Bangladesh | Sweden | Uganda | Brazil | Bangladesh | |
| Critical (C) | 2 | - | 1 | - | - | - | - | - | - | - | - | - |
| Critical-maintained (CM) | 2 | - | - | - | - | - | - | - | - | - | - | - |
| Endangered (D) | - | 3 | 1 | - | - | - | - | - | - | 1 | - | |
| Endangered-maintained (DM) | 1 | - | 3 | - | - | - | - | - | - | - | - | - |
| Extinct (X) | 4 | - | 15 | 3 | - | - | - | - | - | - | - | - |
| Not at risk (NR) | 5 | 2 | 6 | 2 | - | 1 | - | 3 | 8 | 2 | 41 | 5 |
| Unknown | - | 2 | 13 | 1 | - | 1 | 1 | - | - | - | 2 | - |
| Total | 14 | 4 | 41 | 7 | - | 2 | 1 | 3 | 8 | 2 | 44 | 5 |
2.2. Data Source
2.3. Categories for Exploring Cattle Genetic Resources
- Category 1: Breeds that have been declared extinct;
- Category 2: Breeds with unknown risk status;
- Category 3: Local breeds that are declining in numbers;
- Category 4: Local breeds that are not at risk;
- Category 5: The transboundary breeds.
3. Results and Discussion
3.1. Status of Cattle Breeds
3.1.1. Category 1: Breeds That Have Been Declared Extinct
3.1.1.1. Breeds That Have Been Declared Extinct but Their Alleles Have Been Incorporated in Other Breeds
3.1.1.2. Breeds That Have Been Declared Extinct, but Their Ancestors Exist (or Unknown) in the Same/Other Region(s) with Different/Similar Breed Name
3.1.1.3. Breeds That Have Been Declared Extinct—Discontinued Experimental Animals and Unrecognized Breeds
3.1.2. Category 2: Breeds with Unknown Risk Status
3.1.3. Category 3: Local Breeds That Are Declining in Numbers
3.1.3.1. A Local Breed Is Being Upgraded with a Similar Breed and Has More or Less Lost Its Original Characteristics
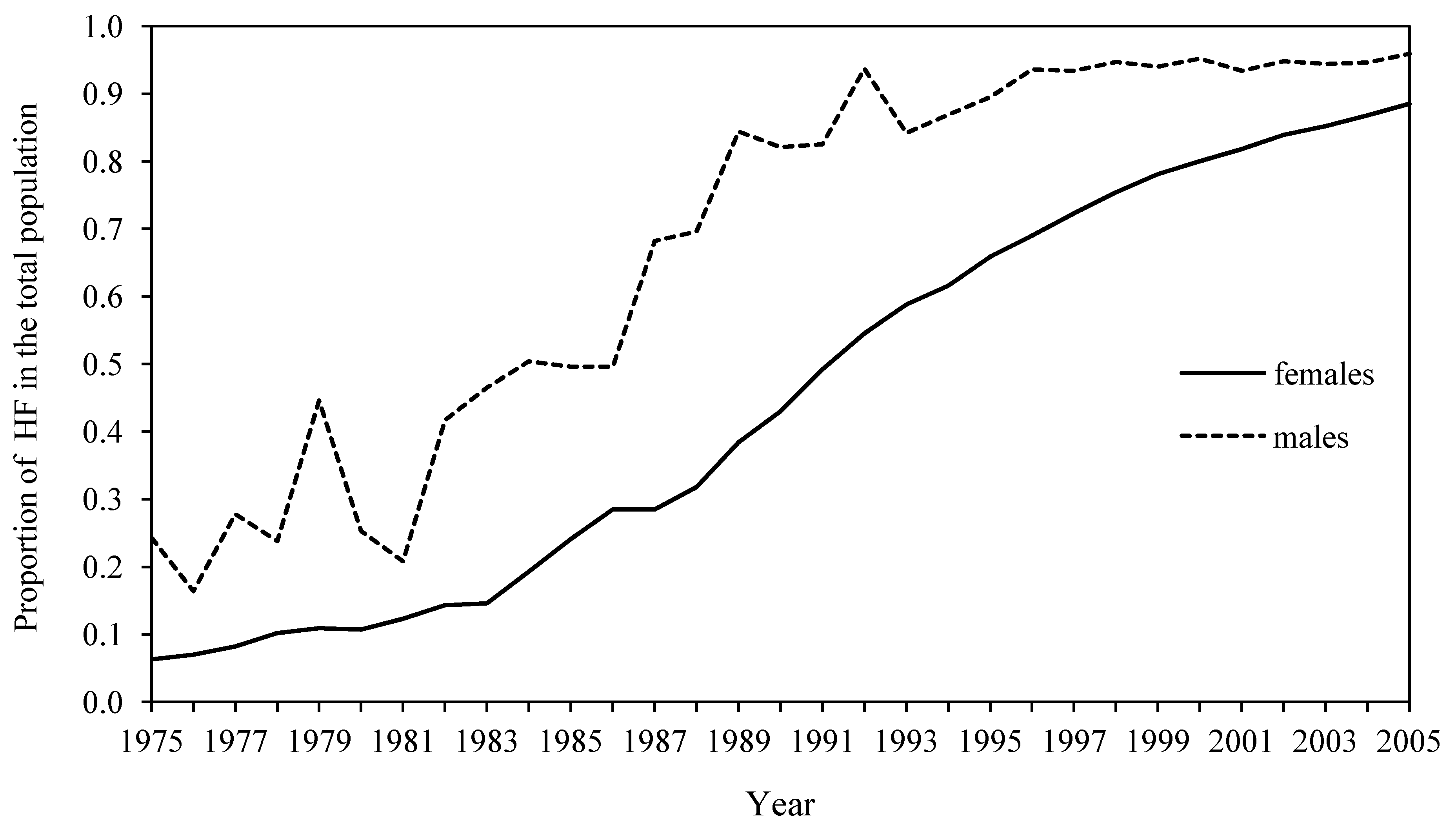
3.1.3.2. Local Breeds That Are Declining in Numbers and Further Split into Subpopulations—Not Exchanging Genetic Material, Despite Their Common Origin
3.1.3.3. Local Breeds That Are Remnants of the Landrace Breeds That Formed the More Developed Breeds
3.1.4. Category 4: Local Breeds That Are Not at Risk
3.1.5. Category 5: The Transboundary Breeds
3.2. The Fate of Extinct Cattle Breeds
4. Implications
4.1. Conservation through Utilization of Cattle Genetic Resources
4.2. Where Is the Evidence of Breed Status?
Genetic studies on cattle breeds in Sweden [26], in Brazil [22,23] and Uganda [24] clearly demonstrates similar ancestral lineages through which some of these cattle breed populations have developed. Their results also correspond well to the documented breed histories [2] and the breed survey findings in this study. There is not enough evidence however to show that there is complete assimilation of the genetic/allelic diversity or substantial loss of genetic diversity/allelic diversity in the new developed breeds from the lost breeds. This excludes the obvious systematic breed combinations in Brazil and those that have been discussed in Section 3.1.1.3. The dilemma is whether to treat breed populations that have lost their uniqueness through continuous breed combinations as extinct or not. A risk status category quasi—extant has been suggested to take care of these cattle breeds. There is an on-going ILRI project, Germplasm for Dairy Development in East Africa (DGEA), which aims at identifying appropriate germplasm and their delivery mechanisms in Uganda and Kenya. This will be done by characterizing the breed composition of crossbred dairy cows kept by smallholder farmers using high density single nucleotide polymorphism (SNP) markers. This study will be able to reveal the extent of breed substitution of local indigenous animals that were used as the base population during initial crossbreeding and the type and nature of alleles or genetic diversity of the indigenous animals that is preserved in the crossbreds.
5. Conclusions
Conflicts of Interest
Appendix
| Breed name | Local name | Transboundary name 1 | Status in DAD-IS | New status (Tick/give new sign) | Description of origin in DAD-IS and additional comments from the experts in italics |
|---|---|---|---|---|---|
| Australian Friesian Sahiwal (AFS) | Australian Friesian Sahiwal (7) | - | X | Imported AFSs were used in indiscriminate crossbreeding programme (on station and in field). A good numbers died on-station due to poor management and GE. | |
| Bangladeshi | Bengali (2) | NR | NR | Indigenous breed of Bangladesh. | |
| Chittagong Red | Austamukhi/Sundari | NR | NR | Local variety of Bangladeshi breed, Active conservation through utilization programme in existence. | |
| Dacca-Faridpur | Dhaka-Faridpur | X | X | Similar to the Hariana breed, distinctiveness disappeared after cross-breeding with Sahiwal, Sindhi and Friesian. | |
| Friesian | Holstein (165) | NR | NR | Imported from Australia; semen continually imported from Pakistan, Australia, New Zealand and Germany, 100% pure individuals are seldom found nowadays. | |
| Hariana | Hariana (5) | NR | X | Indigenous breed. | |
| Holstein | Holstein (165) | NR | NR | Imported but 100% pure individuals are seldom found nowadays. | |
| Jersey | Jersey (82) | NR | X and/or C | Imported from Australia; discarded from crossbreeding programme after 1982, remnants of Jersey inheritance in crossbreds is seen in the whole country but in a milk pocket area—Milk Vita (a national dairy cooperative) is using pure Jersey bulls in their milk-pocket area (5% of the whole country). | |
| Kamdhino | X | X | Local Bangladeshi variety, but occasionally incidence is reported in the newspaper. | ||
| Madaripur | NR | X | Variety of Bengali. | ||
| Munshiganj | X | In between C or X | Probably originated from local Bangladeshi x Red Sindhi: disappeared after crossing with Sahiwal and Friesian. Females are still seen but mature males are rarely noticed. | ||
| North Bengal Grey | - | D | Indigenous breed, mainly seen in the northern districts specially Bogra of Bangladesh. | ||
| Pabna | Shahjadpur | D | C | This variety evolved by breeding indigenous Bangladeshi cows with bulls (Hariana and Sahiwal) in early 1900s. | |
| Sahiwal | Lola/Mont-gomery/Multani/Teli | Sahiwal (30) | NR | NR | But finding 100% purebred Sahiwal is a challenge in the country. |
| Sindhi | Red Sindhi (15) | NR | C | Imported from Pakistan; discarded from crossbreeding program after 1972, but finding 100% purebred Sindhi is a challenge in the country. |
| Breed name | Local name | Transboundary name 1 | Status in DAD-IS | New status (Tick/give new sign) | Description of origin in DAD-IS and additional comments from the experts in italics |
|---|---|---|---|---|---|
| Boran | East African Short horned Zebu | Boran (11) | NR | NR | |
| Karamajong | Karamajong | - | NR | East African Zebu, similar to Toposa and Turkana. | |
| Kigezi | ? 2 | - | ? 2 | Variety of Ankole. | |
| Longhorn Ankole | Ankole/Sanga Nsagalla (Nsagara) | Ankole (6) | NR | NR | The Ankole Longhorn cattle is an intermediate Bos taurus indicus / Bos taurus taurus breed type. |
| Lugware (Lugbara) | Bahu | Lugware (2) | NR | NR | Variety of Small East African Zebu. |
| Nganda | Kyoga/Nsoga/Nyoro/Serere | NR | NR (there is a chance that the situation can change quickly because of interventions such as that promoted by the EADD project in central Uganda.) | Possibly composite of Ankole and Nkedi: The Nganda possesses characteristics which are intermediate between the Ankole and short horn Zebu breeds. | |
| Nkedi | Lango/Teso/Bukedi/Eastern province Zebu | NR | NR | Variety of Small East African Zebu. | |
| Nsagalla (Nsagara) | Bahima | Bahima (2) | NR | Not different from Ankole for all practical purposes | Variety of Ankole- This is an ecotype of the Ankole. Since it is already included, it should not stand alone here. |
| Breed name | Other names | Status in DAD-IS | New status (Suggested) | N 1 |
|---|---|---|---|---|
| Herrgård | X | Quasi-extant | - | |
| Skåne | X | Quasi-extant | - | |
| Småland | X | Quasi-extant | - | |
| Allmoge ko | C | ? | ? 2 | |
| Fjällnära ko | C | ? | ? 3 | |
| Bohuskulla | CM | CM | 36 | |
| Väneko | DM | DM | 187 | |
| Fjällras | ? 4 | NR | 4,423 | |
| Rödbrokig Svensk Boskap (RSB) | Red pied Swedish | X | Quasi-extant | - |
| Ringamålako | CM | CM | 119 | |
| Rödkulla | Swedish Red Poll | NR | NR | 1,746 |
| Svensk Jersey Boskap (SJB) | Swedish Jersey Cattle | NR | NR | 5,527 |
| Svensk Röd och Vit Boskap (SRB) | Swedish Red | NR | NR | 307,112 |
| Svensk Kullig Boskap (SKB) | Swedish Polled Cattle | NR | NR | 3,792 |
| Svensk Låglandsboskap (SLB) | Swedish Friesian | NR | C | 100 |
| Svensk Holstein (SLB) | Swedish Holstein | ? 4 | NR | 401,089 |
| Breed name | Other name(s) | Status in DAD-IS | New status (Tick if you agree with status in DAD-IS or give new sign) | Description of origin in DAD-IS and additional comments from the experts in italics |
|---|---|---|---|---|
| Angola | X | ? 1 | Zebu (probably African) x Curraleiro in 19th century. | |
| Aquitanica | Not a breed | - | ? 1 | Synthetic breed, originated through crossbreeding European and zebu breeds, uniting the production of the first group with the rusticity of the second. |
| Caldeano | Caracu caldeano | NR | √ | The breed is a variety of Caraçu and initially descended from the herd of the Dias family, Fazenda Recreio, Pocos de Caldas, Minais Gerais. Pure-breds are still restricted to this herd. |
| Canchim | NR | √ | The breed was formed in one of the EMBRAPA research centres and is a composite of Charolais (5/8) and Zebu (Indo-Brazilian) (3/8). The Nellore became more popular than the Indo-Brazilian and influenced also the formation of the Canchim. | |
| Cangaian | - | Locally adapted breed. | ||
| Caracu | NR | √ Numbers are increasing very fast | “Local” Bos taurus taurus. The origin of the population is reported to be the Portuguese Alem-Tejo (southern Criollo) imported in 1534. The Portuguese breed Minhota was believed to contribute most to the formation of the Caracu. | |
| China | ?1 | X | ? 1 | Southern Criollo x Indian Zebu, blood imported by Baron de Bom Ritiro 1855. |
| Crioulo do Sul | Bruxo/Legitimo/Colonia/Mineiro Southern Crioulo | X | √ | Portuguese origin. |
| Crioulo Lageano | Franqueiro | D | √ | This breed descends from animals brought by Spanish Jesuits. The Crioulo Lageano is believed to have originated from the ancient Hamiticus cattle, characterized by long horns, which were introduced into the South of Spain from the North of Africa. In Brazil, they developed exclusively by natural selection over the last four centuries. |
| Curraleiro | Corral Crioulo/Crioulo nordestino/Goias/Hard Hoof Criollo | DM | √ | This breed descends directly from the Beiroa type of Mirandesa. It seems to be that the name Pe-Duro is much more used in the north-eastern region of Brazil and considered as the main name of the breed in this region. This breed descended from cattle brought by Portuguese and Spanish colonisers. It is thought to have descended from the Mirandesa breed, which still can be found in the Spanish province of León. |
| Franqueiro | Criollo Lageano | C | CM | Descended from Southern Criollo in late 19th century. |
| Brazilian Gir | Gir | NR | ||
| Gir Mocho | Polled Gyr | NR | D | Variety of Brazilian Gir. |
| Guademar | ? | X | Cross of Curraleiro x Ongole created in 1868. | |
| Guzera | Guzerat/Azulego/Kankrej | NR | √ | Descended from Kankrej, imported from India in 1875–1964. |
| Guzera mocho | Polled Guzera | - | Not a breed | Variety of Guzera. |
| Guzolando | - | Not a breed | Synthetic breed, locally adapted breed. | |
| Ibage | Brangus | NR | √ | Composite of Nellore (3/8) and Aberdeen Angus (5/8). Synthetic breed. |
| Igarap | Nanico/Guarapueva | X | ? 1 | Iberian origin. |
| Indo-Europeu leiteiro | Dairy Indo-European | - | ? 1 | Composite of Friesian or others and Zebu. |
| Indubrasil | Indo-Brazilian/Induberaba/Indoanaxa | C Numbers decreasing | Composite of Gir, Kankrej (1910–1930) and Ongole from India (1875–1930). | |
| Irece | Crioulo leiteiro de Irece | X | ? 1 | Variety of Curraleiro. |
| Javanês | X | ? 1 | Originated by Brito Bastos, rio Formoso from one grey Zebu bull named Javanês crossed on local cows, mid-19th century. | |
| Junqueiro | Junqueira | X | D | Originated from Southern Criollo in 19th century. |
| Lavinia | DM | Not a breed | Composite of Brown Swiss (5/8) and Guzera (3/8). | |
| Malabar | X | ? 1 | Curraleiro x Indian Zebu in 19th century. | |
| Mantiqueira | Tribofe | - | Not a breed | Composite of Friesian (5/8) and Gir (3/8). |
| Mestico Leiteiro Brasileiro | Brazilian Dairy Hybrid | - | Not a breed | Composite of European (mainly Holstein) and Zebu (mainly Guzera and Gir). |
| Mocho Nacional | Caracu (polled variety)/Caracu (variedade Mocho)/Brazilian Polled | DM | √ | “Local” Bos taurus taurus. Southern Criollo type: They probably originated from Polled Criollo cattle that spontaneously appeared within the original population living in the southern Brazilian regions coming from Portugal, Spain and Asia. The official creation of the breed started in Nova Odessa, São Paulo State, in 1911. The pedigree registration started in 1939, with the establishment of the Brazilian Association of Mocho Nacional Cattle Breeders. |
| Nellore mocho | Polled Nellore | NR | √ | Variety of Nellore with Brazilian Polled blood. |
| Pantaneiro | Crioulo pantaneiro/Swamp Criollo/Tucara/Tucura | D | DM | Naturalized breed maintained in the border with Bolivia (flooded area). |
| Nilo | Pig breed! | X | | African Zebu imported and crossed with local Cattle. |
| Patuá | Pig breed! | X | | Probably had some zebu blood. |
| Pedreiro | X | ? 1 | Variety of Crioulo Lageano. | |
| Pitangueiras | - | Synthetic breed. Composite of Red Poll (5/8) and Zebu and to a lesser degree Guzera and Gir. | ||
| Polled Crioulo Pereira Camargo | Polled Criollo Lageano | D | DM | This breed probably originates from Polled Criollo cattle that spontaneously appeared within the original population in the southern Brazilian regions. Originally they were imported from Portugal, Spain and Asia. |
| Polled Gyr | Gir Mocho | - | D | In the 1940s, the Polled Gyr was formed to meet market demand, by crossing with the Mocho Nacional and Red Poll (two polled breeds). This variety continued to expand and shows the same traits and functions as the traditional Gyr. In 1976 the pedigree registration of the Polled Gyr started. |
| Quinhentão | ?1 | X | ? 1 | Cross of Ongole x Friesian and Franqueiro. |
| Riopardense | ?1 | - | ? 1 | Composite of Holstein (5/8) and Guzera (3/8). |
| Santa Gabriela | Not a breed | X | ? 1 | Discontinued experimental breed: Red Pied Friesian x (red polled Zebu x Devon-Guzerà) from 1965 on. |
| Tatu | Pig breed! | X | ? 1 | Developed from crosses of Red Sindhi or Sahiwal bulls imported in 1850. |
| Zebu leiteiro de uberada | Dairy Zebu of Uberaba. A herd of an experimental station located in Uberaba | - | Selection of Brazilian Gir (and others) at a government farm. |
References
- Darwin, C. On the Origin of Species by Mean of Natural Selection; John Murray: London, UK, 1859. [Google Scholar]
- The Food and Agriculture Organization of the United Nations (FAO), World Watch List of Domestic Animal Diversity, 3rd ed.; Scherf, B.D. (Ed.) FAO: Rome, Italy, 2000.
- Gandini, G.C.; Ollivier, L.; Danell, B.; Distl, O.; Georgoudis, A.; Groeneveld, E.; Martyniuk, E.; van Arendonk, J.A.M.; Wolliams, J.A. Criteria to assess the degree of endangerment of livestock breeds in Europe. Livest. Prod. Sci. 2004, 91, 173–182. [Google Scholar] [CrossRef]
- Domestic Animal Diversity Information System (DAD-IS) Home Page. Available online: http://dad.fao.org/ (accessed on 10 August 2012).
- Merriam-Webster Web Page. Breed. Available online: http://www.merriam-webster.com/dictionary/breed (accessed on 11 July 2013).
- International Livestock Research Institute (ILRI), Building a Global Research Institute; ILRI: Nairobi, Kenya, 1996.
- Köhler-Rollefson, I. Indigenous practices of animal genetic resource management and their relevance for the conservation of domestic animal diversity in developing countries. J. Anim. Breed. Genet. 1997, 114, 231–238. [Google Scholar] [CrossRef]
- FAO, Global Plan of Action for Animal Genetic Resources and the Interlaken Declaration; FAO: Rome, Italy, 2007.
- Lush, J. Animal Breeding Plans; The Iowa State College Press: Ames, IA, USA, 1943. [Google Scholar]
- MacHugh, D.E. Molecular Biogeography and Genetic Structure of Domesticated Cattle. Ph.D. Thesis, Trinity College, University of Dublin, Dublin, Ireland, March 1996. [Google Scholar]
- Hammond, K.; Leitch, H.W. Towards better management of animal genetic resources. World Anim. Rev. 1995, 84–85, 48–53. [Google Scholar]
- FAO, The Global Strategy for the Management of Farm Animal Genetic Resources; FAO: Rome, Italy, 1999.
- Tisdell, C. Socioeconomic causes of loss of animal genetic diversity: Analysis and assessment. Ecol. Econ. 2003, 45, 365–376. [Google Scholar] [CrossRef]
- FAO, The State of the World’s Animal Genetic Resources for Food and Agriculture; Rischkowsky, B.; Pilling, D. (Eds.) FAO: Rome, Italy, 2007.
- Bett, R.C.; Johansson, K.; Zonabend, E.; Malmfors, B.; Ojango, J.; Okeyo, A.M.; Philipsson, J. Trajectories of Evolution and Extinction in the Swedish Cattle Breeds. In Proceeding of the 9th World Congress Applied to Livestock Production, Leipzig, Germany, 2–6 August 2010.
- Mariante, A.S.; Albuquerque, M.S.M.; Egito, A.A.; McManus, C.; Lopes, M.A.; Paiva, S.R. Present status of the conservation of livestock genetic resources in Brazil. Livest. Sci. 2009, 120, 204–212. [Google Scholar] [CrossRef]
- Swedish Board of Agriculture, The Development of Animal Genetic Resources in Sweden 2007 and 2008; Jordbruksverket: Jönköping, Sweden, 2011.
- Bhuiyan, A.K.F. Research on Characterization, Conservation and Improvement of Red Chittagong Cattle of Bangladesh; Chaudhury Printing and Publications: Mymensingh, Bangladesh, 2008. [Google Scholar]
- Cruz, R.V.; Harasawa, H.; Lal, M.; Wu, S.; Anokhin, Y.; Punsalmaa, B.; Honda, Y.; Jafari, M.; Li, C.; Huu Ninh, N. Asia. Climate Change 2007: Impacts, Adaptation and Vulnerability. In Contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Parry, M.L., Canziani, O.F., Palutikof, J.P., van der Linden, P.J., Hanson, C.E., Eds.; Cambridge University Press: Cambridge, UK, 2007; pp. 469–506. [Google Scholar]
- FAO, Country Reports on the State of Animal Genetic Resources: Annex to The State of the World’s Animal Genetic Resources for Food and Agriculture. FAO: Roma, Italy, 2007.
- Mariante, A.S.; Egito, A.A. Animal genetic resources in Brazil: Result of five centuries of natural selection. Theriogenology 2002, 57, 223–235. [Google Scholar] [CrossRef]
- Primo, A.T. The Iberian Cattle in the Americas. In Reunião Anual da Sociedade Brasileira de Zootecnia, Anais; Sociedade Brasileira de Zootecnia: Rio de Janeiro, Brazil, 1993; pp. 183–199. [Google Scholar]
- Serrano, G.M.; Egito, A.A.; McManus, C.; Mariante, A. Genetic Diversity and Population Structure of Brazilian Native Bovine Breeds Based on RAPD Markers. In Proceedings of the 7th World Congress on Genetics Applied to Livestock Production, Montpellier, France, 2002.
- Ndumu, D.B.; Baumung, R.; Hanotte, O.; Wurzinger, M.; Okeyo, M.; Jianlin, H.; Kibogo, H; Sölkner, J. Genetic and morphological characterisation of the Ankole Longhorn cattle in the African Great Lakes region. Genet Sel. Evol. 2008, 40, 467–490. [Google Scholar]
- Swedish Dairy Association Home Page. Available online: http://www.svenskmjolk.se/ (accessed on 23 February 2010).
- Tapio, I.; Värv, S.; Bennewitz, J.; Maleviciute, J.; Fimland, E.; Grislis, Z.; Meuwissen, T.H.E.; Miceikiene, I.; Olsaker, I.; Viinalass, H.; et al. Prioritization for conservation of Northern European cattle breeds based on analysis of microsatellite data. Conserv. Biol. 2006, 20, 1768–1779. [Google Scholar] [CrossRef]
- Hammack, S.P. Texas Adapted Genetic Strategies for Beef Cattle VI: Creating Breeds; AgriLIFE Extension Texas A&M: College Station, Texas, USA, 2009. [Google Scholar]
- Toro, M.; Mäki-Tanila, A. Genomics Reveals Domestication History and Facilitates Breed Development. In Utilisation and Conservation of Farm Animal Genetic Resources; Oldenbroek, K., Ed.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2007; pp. 75–102. [Google Scholar]
- Oldenbroek, K. Utilisation and Conservation of Farm Animal Genetic Resources; Wageningen Academic Publishers: Wageningen, The Netherlands, 2007. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Bett, R.C.; Okeyo, M.A.; Malmfors, B.; Johansson, K.; Agaba, M.; Kugonza, D.R.; Bhuiyan, A.K.F.H.; Filho, A.E.V.; Mariante, A.S.; Mujibi, F.D.; et al. Cattle Breeds: Extinction or Quasi-Extant? Resources 2013, 2, 335-357. https://doi.org/10.3390/resources2030335
Bett RC, Okeyo MA, Malmfors B, Johansson K, Agaba M, Kugonza DR, Bhuiyan AKFH, Filho AEV, Mariante AS, Mujibi FD, et al. Cattle Breeds: Extinction or Quasi-Extant? Resources. 2013; 2(3):335-357. https://doi.org/10.3390/resources2030335
Chicago/Turabian StyleBett, Rawlynce C., Mwai A. Okeyo, Birgitta Malmfors, Kjell Johansson, Morris Agaba, Donald R. Kugonza, A.K.F.H. Bhuiyan, Anibal E. Vercesi Filho, Arthur S. Mariante, Fidalis D. Mujibi, and et al. 2013. "Cattle Breeds: Extinction or Quasi-Extant?" Resources 2, no. 3: 335-357. https://doi.org/10.3390/resources2030335
APA StyleBett, R. C., Okeyo, M. A., Malmfors, B., Johansson, K., Agaba, M., Kugonza, D. R., Bhuiyan, A. K. F. H., Filho, A. E. V., Mariante, A. S., Mujibi, F. D., & Philipsson, J. (2013). Cattle Breeds: Extinction or Quasi-Extant? Resources, 2(3), 335-357. https://doi.org/10.3390/resources2030335