PRDM Proteins: Molecular Mechanisms in Signal Transduction and Transcriptional Regulation



Abstract
:1. Structure of PRDM Proteins and Their Alternative Gene Products
1.1. Alternative Promoters
1.2. Alternative Splicing

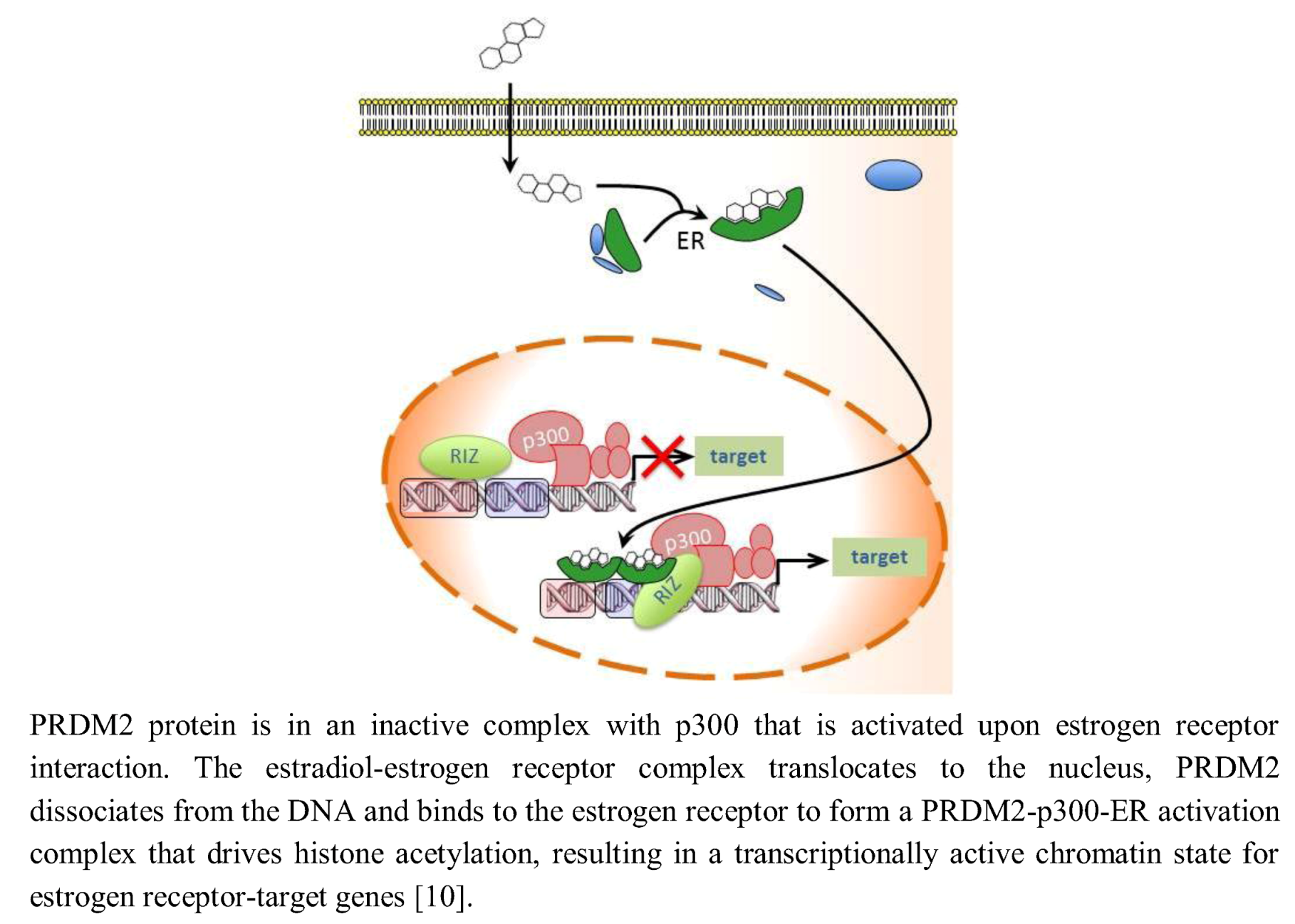{kind=link}
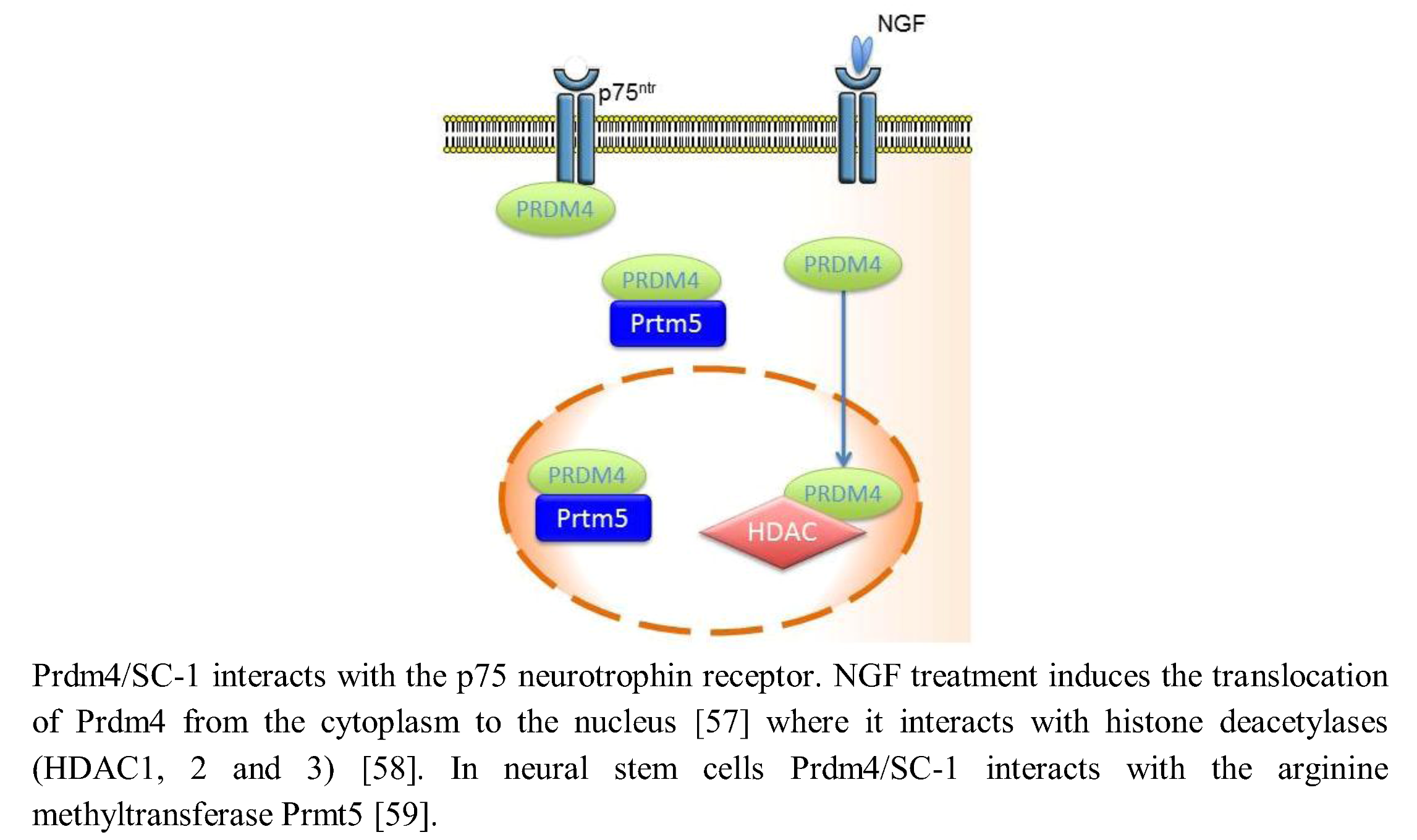{kind=link}
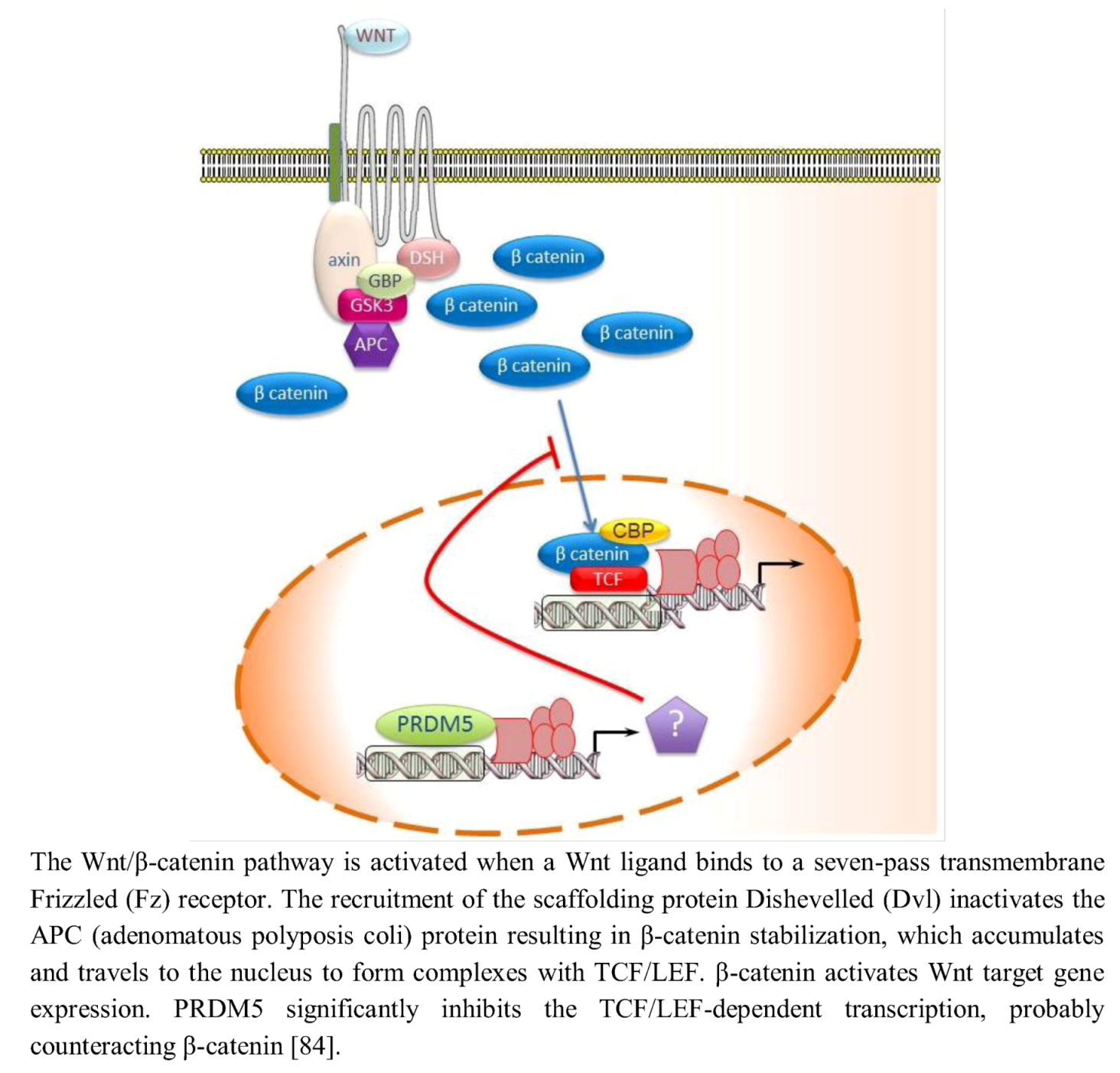{kind=link}
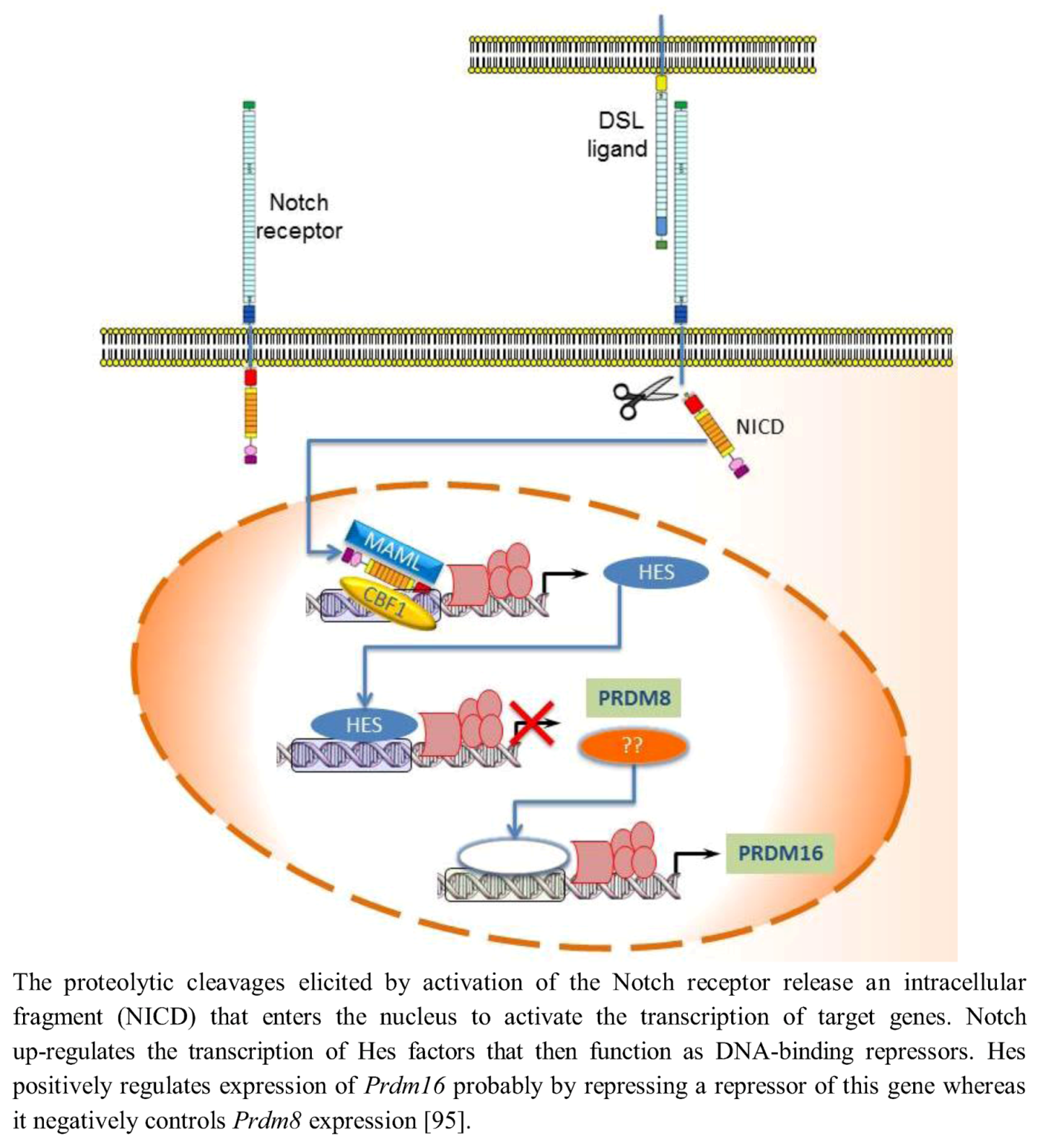{kind=link}
{kind=link}
| Human | ||||||||
|---|---|---|---|---|---|---|---|---|
| gene name | protein name | localization | molecular forms | alternative promoter (UNIPROT entry) | splicing variants (UNIPROT entry) | length (aa) | PR domain | HMT activity |
| PRDM1 (BLIMP1) | PR domain zinc finger protein 1 (BLIMP1) | nucleuscytoplasm | 3 | Isoform 1 'canonical' sequence (O75626-1) | Isoform 2 | 825 | aa 85-205 | no |
| 1-36: missing | ||||||||
| (Beta-interferon gene positive regulatory domain I-binding factor) | (O75626-2) | |||||||
| Isoform 3 | partially missing | no | ||||||
| 1-3: MLD → MEK | ||||||||
| (PR domain-containing protein 1) | 4-137: missing | |||||||
| Positive regulatory domain I-binding factor 1) | (O75626-3) | |||||||
| PRDM2 (KMT8, RIZ) | PR domain zinc finger protein 2 | nucleus | 3 | Isoform 1 (RIZ1) 'canonical' sequence (Q13029-1) | Isoform 2 (MTB-Zf) | 1,718 | aa 27-145 | H3K9 |
| (GATA-3-binding protein G3B) | 1679-1682: SYSL → RNFL | |||||||
| 1683-1718: missing * | ||||||||
| (Lysine N - methyltransferase 8) | (Q13029-2) | |||||||
| (MTB-ZF) | Isoform 3 (RIZ2) 1-201: missing (13029-3) | no | no | |||||
| (MTE-binding protein) | ||||||||
| (PR domain-containing protein 2) | ||||||||
| (RIZ, Retinoblastoma protein-interacting zinc finger protein) | ||||||||
| PRDM3/MECOM (EVI1) | MDS1 and EVI1 complex locus protein EVI1 (Ecotropic virus integration site 1 protein homolog-EVI-1) | nucleus | 6 | Isoform 1 (Evi-1a) 'canonical' sequence | 1,051 | aa 79-194 | H3K9me1 | |
| (Q03112-1) | ||||||||
| Isoform 2 (Evi-1c) (Mds1/Evi1) | ||||||||
| (Q03112-3) | ||||||||
| 1-1: M → MRSKGRARKL...APGEELLL-FM | ||||||||
| Contains an additional SET domain at positions 79-194 | ||||||||
| Isoform 3 (Mds1) | ||||||||
| (Q13465-1) | ||||||||
| Isoform 4 | ||||||||
| 1-1: M → MILDEFYNVKFCIDASQPD-VGSWLKYIRFAGCYDQHNLVACQINDQIFYRVVADIAPGEELLLFM | ||||||||
| 138-138: K → KQ | ||||||||
| (Q03112-4) | ||||||||
| Isoform 5 | ||||||||
| 672-680: missing | ||||||||
| (Q03112-5) | ||||||||
| Isoform 6 | ||||||||
| 138-138: K → KQ | ||||||||
| 672-680: missing | ||||||||
| (Q03112-6) | ||||||||
| PRDM4 (PFM1) | PR domain zinc finger protein 4 (PR domain-containing protein 4) | nucleus | 1 | 801 | aa 412-533 | no | ||
| PRDM5 (PFM2) | PR domain zinc finger protein 5 (PR domain-containing protein 5) | nucleus | 3 | Isoform 1 'canonical' sequence | 630 | aa 8-128 | no | |
| (Q9NQX1-1) | ||||||||
| Isoform 2 | ||||||||
| 218-248: missing | ||||||||
| (Q9NQX1-2) | ||||||||
| Isoform 3 (Q9NQX1-3) | ||||||||
| 101-111: EGENIFYLAVE → DKNLGP-AEWRG | ||||||||
| 112-630: missing | ||||||||
| PRDM6 (PFM3) | Putative histone-lysine N -methyltransferase PRDM6 | nucleus | 3 | Isoform 1 (Q9NQX0-3) | 595 | aa 247-369 | H4K20 | |
| 'canonical' sequence | ||||||||
| (PR domain zinc finger protein 6) | Isoform 2 (B) | |||||||
| 1-182: missing | ||||||||
| (PR domain-containing protein 6) | ||||||||
| 314-595: missing | ||||||||
| (Q9NQX0-2) | ||||||||
| Isoform 3 (A) | ||||||||
| 1-182: missing | ||||||||
| (Q9NQX0-1) | ||||||||
| PRDM7 (PFM4) | Probable histone-lysine
N
- methyltransferase PRDM7 (PR domain zinc finger protein 7) (PR domain-containing protein 7) | nucleus | 3 | Isoform 1 'canonical' sequence | 492 | aa 246-362 | no | |
| (Q9NQW5-3) | ||||||||
| Isoform 2 (B) | ||||||||
| 1-206: missing | ||||||||
| 318-377: YVNCARDDEE...RSSIEPAESL → TKARDPSMSL...RGSESGaaIF | ||||||||
| 378-492: missing | ||||||||
| (Q9NQW5-2) | ||||||||
| Isoform 3 (A) | ||||||||
| 1-206: missing | ||||||||
| 368-492: RSSIEPAESL...VKRSKKGPNS → KWGSKWKKEL...GEAPVCRKDE | ||||||||
| (Q9NQW5-1) | ||||||||
| PRDM8 (PFM5) | PR domain zinc finger protein 8 | nucleus | 2 | Isoform 1 'canonical' sequence | aa 8-135 | H3K9 | ||
| (Q9NQV8-1) | ||||||||
| Isoform 2 | ||||||||
| (PR domain-containing protein 8) | 332-334: GRG → aaL | |||||||
| 335-689: missing | ||||||||
| (Q9NQV8-2) | ||||||||
| PRDM9 ( PFM6 ) | Histone-lysine N -methyltransferase PRDM9 | nucleus | 1 | (Q9NQV7) | 894 | aa 246-362 | H3K4me3 | |
| (PR domain zinc finger protein 9; | ||||||||
| PR domain-containing protein 9) | ||||||||
| PRDM10 (KIaa 1231; PFM7; TRIS) | PR domain zinc finger protein 10 (PR domain-containing protein 10) (Tristanin) | nucleus | 6 | Isoform 3 'canonical' sequence | 1,147 | aa 206-330 | no | |
| (Q9NQV6-3) | ||||||||
| Isoform 2 | ||||||||
| 1-97: MDSKDESSHV...AYVQQDATAQ → MSAYSVPSTFA | ||||||||
| 511-514: missing | ||||||||
| 952-985: missing | ||||||||
| (Q9NQV6-2 | ||||||||
| Isoform 1 | ||||||||
| 1-97: MDSKDESSHV...AYVQQDATAQ → MSAYSVPSTFA | ||||||||
| (Q9NQV6-1) | ||||||||
| Isoform 4 | ||||||||
| 511-514: missing | ||||||||
| 984-984: I → IQVSEPTASAPSSA * | ||||||||
| (Q9NQV6-4) | ||||||||
| Isoform 5 | ||||||||
| 1-97: MDSKDESSHV...AYVQQDATAQ → MSAYSVPSTFA | ||||||||
| 984-984: I → IQVSEPTASAPSSA * | ||||||||
| (Q9NQV6-5) | ||||||||
| Isoform 6 | ||||||||
| 511-514: missing | ||||||||
| 984-984: I → IQVSEPTASAPSSA | ||||||||
| 1132-1147: TTTNGNGSSEVHITKP → AGSKVIQNEF...IVFKRISKRI * | ||||||||
| (Q9NQV6-6) | ||||||||
| PRDM11 (PFM8) | PR domain-containing protein 11 | 2 | Isoform 1 'canonical' sequence | 511 | aa 149-264 | no | ||
| (Q9NQV5-1) | ||||||||
| Isoform 2 | ||||||||
| 1-34: missing * | ||||||||
| (Q9NQV5-2) | ||||||||
| PRDM12 (PFM9) | PR domain zinc finger protein 12 | nucleus | 1 | (Q9H4Q4) | 367 | aa 87-207 | no | |
| (PR domain-containing protein 12) | ||||||||
| PRDM13 ( PFM10 ) | PR domain zinc finger protein 13 | nucleus | 1 | (Q9H4Q3) | 707 | aa 1-116 | no | |
| (PR domain-containing protein 13) | ||||||||
| PRDM14 | PR domain zinc finger protein 14 | nucleus | 1 | (Q9GZV8) | 571 | aa 253-371 | no | |
| (PR domain-containing protein 14) | ||||||||
| PRDM15 (C21orf83; ZNF298) | PR domain zinc finger protein 15 | nucleus | 1 | (P57O71) | 1,507 | aa 406-529 | no | |
| (PR domain-containing protein 15)(Zinc finger protein 298) | ||||||||
| PRDM16 (KIaa 1675; MEL1; PFM13) | PR domain zinc finger protein 16 | nucleus | 4 | Isoform 1 'canonical' sequence | 1,276 | aa 83-215 | H3K9me1 | |
| (Q9HAZ2-1) | ||||||||
| Isoform 2 (MEL1L) | ||||||||
| (PR domain-containing protein 16) | 1233-1251: missing * | |||||||
| (Q9HAZ2-2) | ||||||||
| Isoform 3 | ||||||||
| 191-191: Q → QV | ||||||||
| (Transcription factor MEL1) | 868-868: missing * | |||||||
| (Q9HAZ2-3) | ||||||||
| Isoform 4 | ||||||||
| Also known as: | ||||||||
| MEL1S | ||||||||
| 1-184: missing | ||||||||
| (Q9HAZ2-4) | ||||||||
| ZNF408 (PFM14; PRDM17) | Zinc finger protein 408 | nucleus | (Q9H9D4) | 720 | ||||
| (PR domain zinc finger protein 17) | ||||||||
| Mouse | ||||||||
| gene name | protein name | localization | molecular forms | alternative promoter(UNIPROT entry) | splicing variants(UNIPROT entry) | length (aa) | PR domain | HMT activity |
| Prdm1 (Blimp1) | PR domain zinc finger protein 1(B lymphocyte-induced maturation protein 1-Blimp1) | nucleuscytoplasm | 5 | Isoform 1 'canonical' sequence(Q60636-1) | Isoform 2 | 856 | aa 118-237 | no |
| Also known as: 1A | ||||||||
| 1-47: MREAYLRCWIFSWKNVWVRP-CQRLHFKTVLLQGSLLYTALDSYSTVQ → MLDLLLEKRVGTTL | ||||||||
| (Q60636-2) | ||||||||
| Isoform 3 Also known as: 1B 1-67: missing (Q60636-3) | Isoform 4 (1C) | |||||||
| 1-47: MREAYLRCWI...TALDSYSTVQ → MTPGVPGHRTQQRPQHISALSDK-AKDCSK | ||||||||
| (Q60636-4) | ||||||||
| Isoform 5 | ||||||||
| Also known as: delta exon 7; | ||||||||
| (Beta-interferon gene positive regulatory domain I-binding factor)(PR domain-containing protein 1) | ||||||||
| 624-666: missing | ||||||||
| (Q60636-5) | ||||||||
| Prdm2 (KMT; Riz1 ; Znfpr1c1 ) | Prdm2 protein | nucleus | 1 | 1,670 | aa 34-144 | H3K9 | ||
| PRDM3 / Mecom ( Evi1 ) | MDS1 and EVI1 complex locus protein EVI1 (Ecotropic virus integration site 1 protein-EVI-1) | nucleus | 2 | Isoform 1 'canonical' sequence | 1,042 | aa 81-196 | H3K9me1 | |
| (P14404-1) | ||||||||
| Isoform 2 | ||||||||
| (Q9Z1L8-1) | ||||||||
| Prdm4 | PR domain zinc finger protein 4 | nucleus | 1 | (Q80V63) | 803 | aa 415-536 | no | |
| (PR domain-containing protein 4) | ||||||||
| Prdm5 | PR domain zinc finger protein 5 | nucleus | 1 | (Q9CXE0) | 599 | aa 8-128 | no | |
| (PR domain-containing protein 5) | ||||||||
| Prdm6 ( Gm92 ; Prism ) | Putative histone-lysine
N
-methyltransferase PRDM6 (PR domain zinc finger protein 6) (PR domain-containing protein 6) | Isoform 1 'canonical' sequence | 596 | aa 248-370 | H4K20 | |||
| (Q3UZD5-1) | ||||||||
| Isoform 2 | ||||||||
| 1-201: missing | ||||||||
| (Q3UZD5-2) | ||||||||
| Isoform 3 | ||||||||
| 1-392: missing | ||||||||
| (Q3UZD5-3) | ||||||||
| Isoform 4 | ||||||||
| 28-58: missing | ||||||||
| (Q3UZD5-4) | ||||||||
| Prdm8 | PR domain zinc finger protein 8 | nucleus | (Q8BZ97) | 687 | aa 8-135 | H3K9 | ||
| (PR domain-containing protein 8) | ||||||||
| Prdm9 ( Hst1 ; Meisetz ) | Histone-lysine N -methyltransferase PRDM9 | nucleus | 4 | Isoform 1 'canonical' (Meisetz) | 843 | aa 246-362 | H3K4me3 | |
| (Q96EQ9-1) | ||||||||
| Isoform 2 (Meisetz-S1) | ||||||||
| (Hybrid sterility protein 1) | 382-404: ELRTEIHPCLLCSLAFSSQKFLT → GGHYYDSLKKKEKREFSLRIFIF | |||||||
| (Meiosis-induced factor containing a PR/SET domain and zinc-finger motif) | 405-843: missing | |||||||
| (Q96EQ9-2) | ||||||||
| Isoform 3 (Meisetz-S2) | ||||||||
| 382-418: ELRTEIHPCLLCSLAFSSQKFL-TQHMEWNHRTEIFPG → DLFIIICKYT-VAVFRHTRRGSQILLRMVVSHHVVAGI | ||||||||
| (PR domain zinc finger protein 9) | ||||||||
| 419-843: missing | ||||||||
| (PR domain-containing protein 9) | ||||||||
| (Q96EQ9-3) | ||||||||
| Isoform 4 | ||||||||
| 1-121: missing | ||||||||
| 382-404: ELRTEIHPCLLCSLAFSSQKFLT → GGHYYDSLKKKEKREFSLRIFIF | ||||||||
| 405-843: missing | ||||||||
| (Q96EQ9-4) | ||||||||
| Prdm10 ( Gm1112 , Tris ) | PR domain zinc finger protein 10 | nucleus | 2 | Isoform 1 'canonical' sequence | 1,184 | aa 200-324 | no | |
| (Q3UTQ7-1) | ||||||||
| Isoform 2 | ||||||||
| 318-341: WYaaSYAEFVNQKIHDISEEE-RKV → QNWIHSCLPARVMIRALSY-KRILP | ||||||||
| (PR domain-containing protein 10) | ||||||||
| 342-1184: missing | ||||||||
| (Tristanin) | ||||||||
| (Q3UTQ7-2) | ||||||||
| Prdm11 | PR domain-containing protein 11 | nucleus | 1 | (A2AGX3) | 565 | aa 115-230 | no | |
| Prdm12 (Gm998) | PR domain zinc finger protein 12 | nucleus | 1 | (A2AJ77) | 365 | aa 87-207 | no | |
| (PR domain-containing protein 12) | ||||||||
| Prdm13 | PR domain zinc finger protein 13 | nucleus | 2 | Isoform 1 'canonical' sequence; | 754 | aa 5-164 | no | |
| (E9PZZ1-1) | ||||||||
| Isoform 2 | ||||||||
| (PR domain-containing protein 13) | ||||||||
| 1-48: missing | ||||||||
| (E9PZZ1-2) | ||||||||
| Prdm14 | PR domain zinc finger protein 14 | nucleus | 1 | (E9Q3T6) | 561 | aa 243-360 | no | |
| (PR domain-containing protein 14) | ||||||||
| PRDM15 (C21orf83; E130018M06Rik; ORF62; Zfp298) | PR domain containing 15 | nucleus | 1 | 1,174 | aa 76-191 | no | ||
| PRDM16 (Kiaa; 1675; Mel1) | PR domain zinc finger protein 16 | nucleus | 3 | Isoform 1 'canonical' sequence | 1,275 | aa 83-215 | H3K9me1 | |
| (A2A935-1) | ||||||||
| Isoform 2 | ||||||||
| 129-129: E → EQ | ||||||||
| (PR domain-containing protein 16) | ||||||||
| 868-868: Y → YS | ||||||||
| 1174-1176: CVE → HMQ | ||||||||
| 1177-1275: missing * | ||||||||
| (Transcription factor MEL1) | (A2A935-2) | |||||||
| Isoform 3 | ||||||||
| 868-868: Y → YS * | ||||||||
| (A2A935-3) | ||||||||
2. PRDM Proteins in Signal Transduction and Transcription Control
2.1. Nuclear Receptor Superfamily Signal Transduction

2.2. Luteinizing Hormone (LH) Signaling
2.3. Insulin-Like Growth Factor-1 (IGF-1) Signaling
2.4. NGF Signaling

2.5. Wnt/β-Catenin and BMP/SMAD Signaling

2.6. Neural Progenitor Maintenance and Differentiation
2.7. Notch Signaling
2.8. Neural Circuit Formation

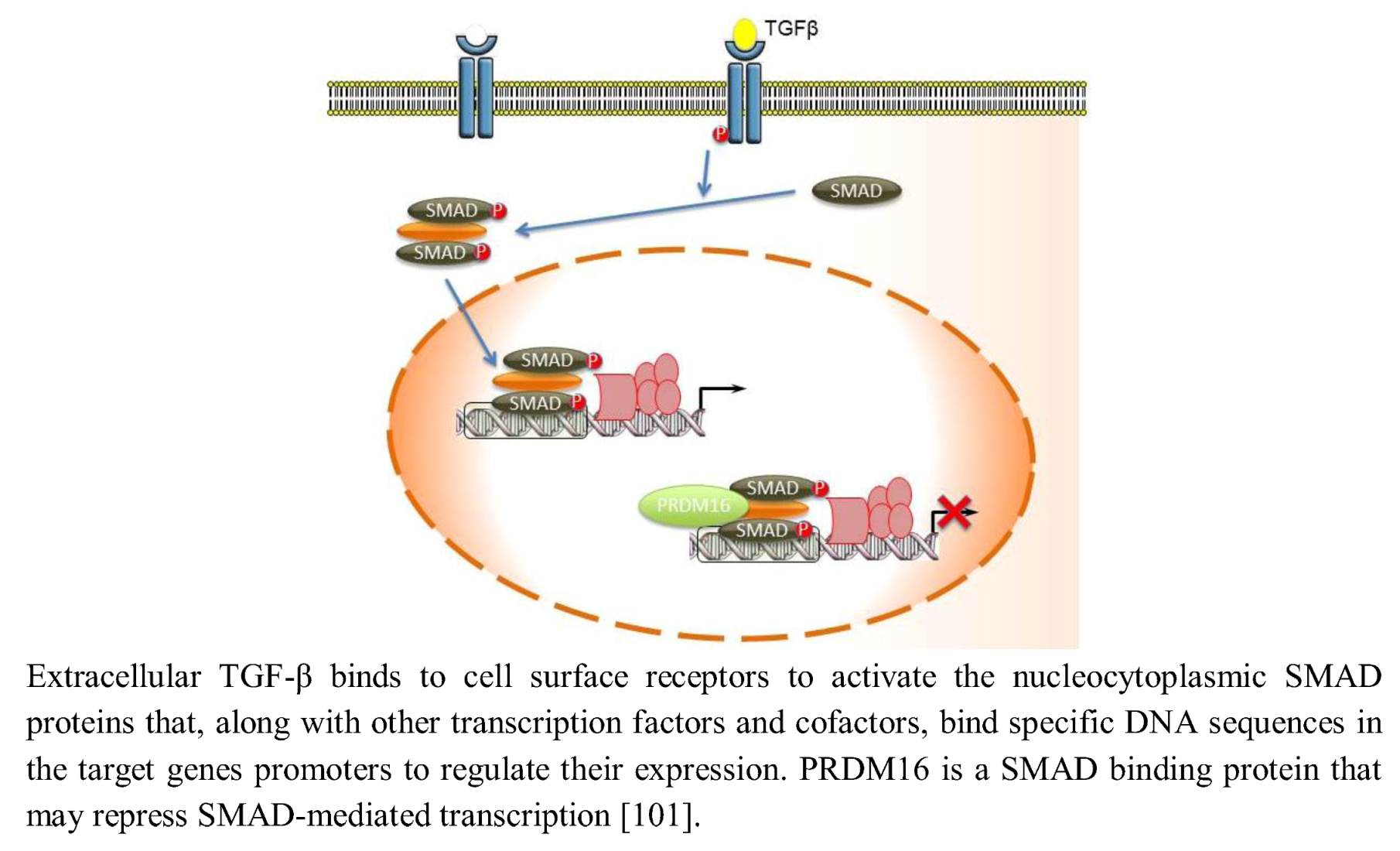
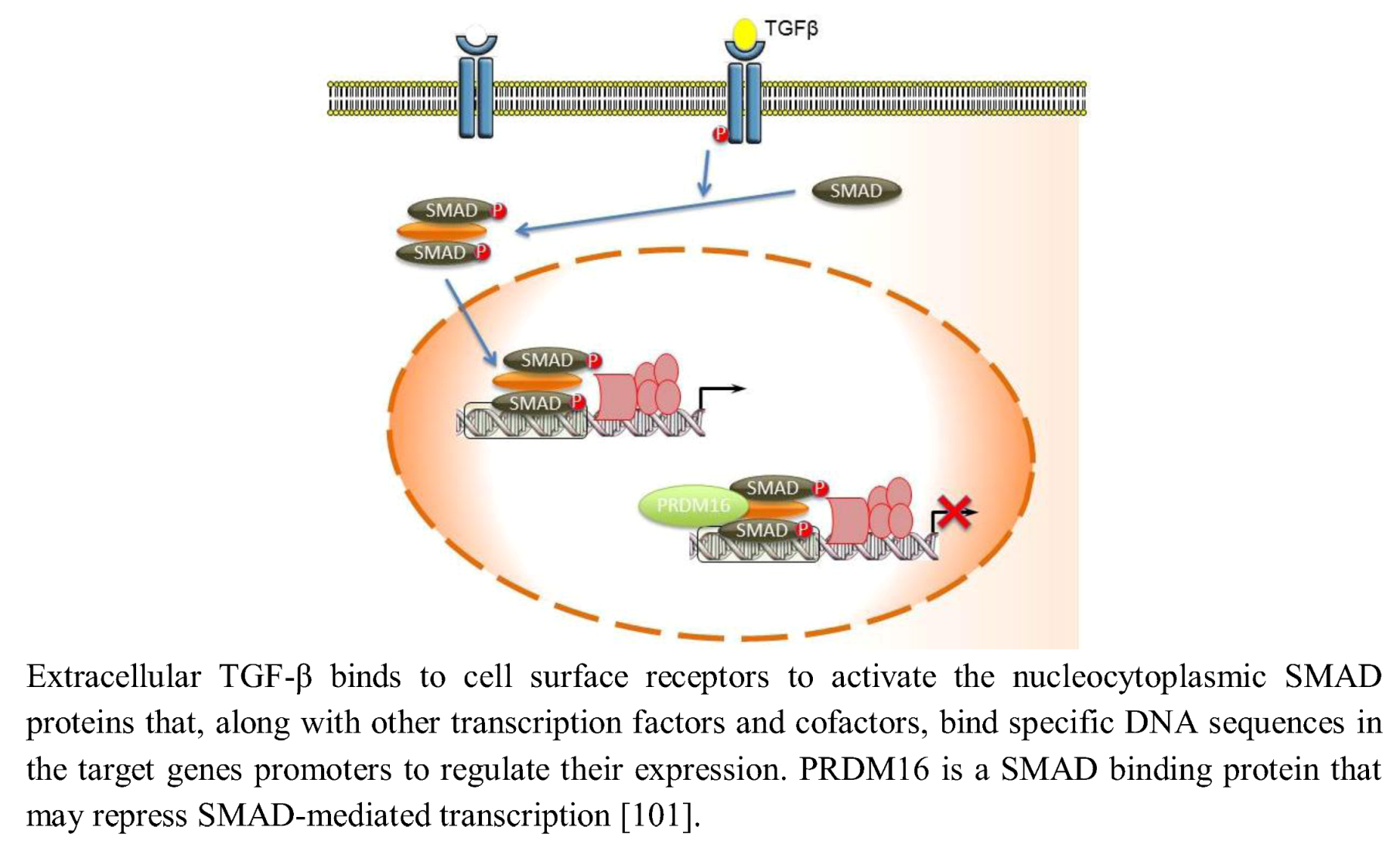
2.9. TGF-β Signaling
3. PRDM Proteins in the Host Defence
5. Conclusions and Perspectives
Acknowledgements
References and Notes
- Xiao, B.; Wilson, J.R.; Gamblin, S.J. SET domains and histone methylation. Curr. Opin. Struct. Biol. 2003, 13, 699–705. [Google Scholar] [CrossRef]
- Fumasoni, I.; Meani, N.; Rambaldi, D.; Scafetta, G.; Alcalay, M.; Ciccarelli, F. Family expansion and gene rearrangements contributed to the functional specialization of PRDM genes in vertebrates. BMC Evol. Biol. 2007, 7, 187. [Google Scholar]
- Fog, C.K.; Galli, G.G.; Lund, A.H. PRDM proteins: Important players in differentiation and disease. BioEssays 2011, 34, 50–60. [Google Scholar]
- Gyory, I.; Fejer, G.; Ghosh, N.; Seto, E.; Wright, K.L. Identification of a functionally impaired positive regulatory domain I binding factor 1 transcription repressor in myeloma cell lines. J. Immunol. 2003, 170, 3125–3133. [Google Scholar]
- Hirai, H. The transcription factor Evi-1. Int. J. Biochem. Cell Biol. 1999, 31, 1367–1371. [Google Scholar] [CrossRef]
- Liu, L.; Shao, G.; Steele-Perkins, G.; Huang, S. The retinoblastoma interacting zinc finger gene RIZ produces a PR domain-lacking product through an internal promoter. J. Biol. Chem. 1997, 272, 2984–2991. [Google Scholar] [CrossRef]
- Lahortiga, I.; Agirre, X.; Belloni, E.; Vazquez, I.; Larrayoz, M.J.; Gasparini, P.; Lo Coco, F.; Pelicci, P.G.; Calasanz, M.J.; Odero, M.D. Molecular characterization of a t(1;3)(p36;q21) in a patient with MDS. MEL1 is widely expressed in normal tissues, including bone marrow, and it is not overexpressed in the t(1;3) cells. Oncogene 2004, 23, 311–316. [Google Scholar]
- Hayashi, K.; Yoshida, K.; Matsui, Y. A histone H3 methyltransferase controls epigenetic events required for meiotic prophase. Nature 2005, 438, 374–378. [Google Scholar] [CrossRef]
- Wu, Y.; Ferguson Iii, J.E.; Wang, H.; Kelley, R.; Ren, R.; McDonough, H.; Meeker, J.; Charles, P.C.; Wang, H.; Patterson, C. PRDM6 is enriched in vascular precursors during development and inhibits endothelial cell proliferation, survival, and differentiation. J. Mol. Cell. Cardiol. 2008, 44, 47–58. [Google Scholar] [CrossRef]
- Hohenauer, T.; Moore, A.W. The Prdm family: Expanding roles in stem cells and development. Development 2012, 139, 2267–2282. [Google Scholar]
- Turner, C.A., Jr.; Mack, D.H.; Davis, M.M. Blimp-1, a novel zinc finger-containing protein that can drive the maturation of B lymphocytes into immunoglobulin-secreting cells. Cell 1994, 77, 297–306. [Google Scholar] [CrossRef]
- Mochizuki, N.; Shimizu, S.; Nagasawa, T.; Tanaka, H.; Taniwaki, M.; Yokota, J.; Morishita, K. A novel gene, MEL1, mapped to 1p36.3 is highly homologous to the MDS1/EVI1 gene and is transcriptionally activated in t(1;3)(p36;q21)-positive leukemia cells. Blood 2000, 96, 3209–3214. [Google Scholar]
- Nishikata, I.; Sasaki, H.; Iga, M.; Tateno, Y.; Imayoshi, S.; Asou, N.; Nakamura, T.; Morishita, K. A novel EVI1 gene family, MEL1, lacking a PR domain (MEL1S) is expressed mainly in t(1;3)(p36;q21)-positive AML and blocks G-CSF-induced myeloid differentiation. Blood 2003, 102, 3323–3332. [Google Scholar] [CrossRef]
- Morgan, M.A.J.; Mould, A.W.; Li, L.; Robertson, E.J.; Bikoff, E.K. Alternative splicing regulates Prdm1/Blimp-1 DNA binding activities and co-repressor interactions. Mol. Cell. Biol. 2012, 32, 3403–3413. [Google Scholar]
- Wieser, R. The oncogene and developmental regulator EVI1: Expression, biochemical properties, and biological functions. Gene 2007, 396, 346–357. [Google Scholar] [CrossRef]
- Morishita, K.; Parganas, E.; William, C.L.; Whittaker, M.H.; Drabkin, H.; Oval, J.; Taetle, R.; Valentine, M.B.; Ihle, J.N. Activation of EVI1 gene expression in human acute myelogenous leukemias by translocations spanning 300–400 kilobases on chromosome band 3q26. Proc. Natl. Acad. Sci. USA 1992, 89, 3937–3941. [Google Scholar]
- Fears, S.; Mathieu, C.; Zeleznik-Le, N.; Huang, S.; Rowley, J.D.; Nucifora, G. Intergenic splicing of MDS1 and EVI1 occurs in normal tissues as well as in myeloid leukemia and produces a new member of the PR domain family. Proc. Natl. Acad. Sci. USA 1996, 93, 1642–1647. [Google Scholar] [CrossRef]
- Siegel, D.A.; Huang, M.K.; Becker, S.F. Ectopic dendrite initiation: CNS pathogenesis as a model of CNS development. Int. J. Dev. Neurosci. 2002, 20, 373–389. [Google Scholar] [CrossRef]
- UniProt. Available online: http://www.uniprot.org/ (accessed on 05 October 2012).
- National Center for Biotechnology Information Protein Database. Available online: http://www.ncbi.nlm.nih.gov/protein/ (accessed on 05 October 2012).
- Izutsu, K.; Kurokawa, M.; Imai, Y.; Maki, K.; Mitani, K.; Hirai, H. The corepressor CtBP interacts with Evi-1 to repress transforming growth factor-β signaling. Blood 2001, 97, 2815–2822. [Google Scholar]
- Davis, C.A.; Haberland, M.; Arnold, M.A.; Sutherland, L.B.; McDonald, O.G.; Richardson, J.A.; Childs, G.; Harris, S.; Owens, G.K.; Olson, E.N. PRISM/PRDM6, a Transcriptional Repressor That Promotes the Proliferative Gene Program in Smooth Muscle Cells. Mol. Cell. Biol. 2006, 26, 2626–2636. [Google Scholar] [CrossRef]
- Kajimura, S.; Seale, P.; Tomaru, T.; Erdjument-Bromage, H.; Cooper, M.P.; Ruas, J.L.; Chin, S.; Tempst, P.; Lazar, M.A.; Spiegelman, B.M. Regulation of the brown and white fat gene programs through a PRDM16/CtBP transcriptional complex. Genes Dev. 2008, 22, 1397–1409. [Google Scholar] [CrossRef]
- Yu, J.; Angelin-Duclos, C.; Greenwood, J.; Liao, J.; Calame, K. Transcriptional Repression by Blimp-1 (PRDI-BF1) Involves Recruitment of Histone Deacetylase. Mol. Cell. Biol. 2000, 20, 2592–2603. [Google Scholar] [CrossRef]
- Kim, K.C.; Geng, L.; Huang, S. Inactivation of a histone methyltransferase by mutations in human cancers. Cancer Res. 2003, 63, 7619–7623. [Google Scholar]
- Ancelin, K.; Lange, U.C.; Hajkova, P.; Schneider, R.; Bannister, A.J.; Kouzarides, T.; Surani, M.A. Blimp1 associates with Prmt5 and directs histone arginine methylation in mouse germ cells. Nat. Cell Biol. 2006, 8, 623–630. [Google Scholar] [CrossRef]
- Gyory, I.; Wu, J.; Fejer, G.; Seto, E.; Wright, K.L. PRDI-BF1 recruits the histone H3 methyltransferase G9a in transcriptional silencing. Nat. Immunol. 2004, 5, 299–308. [Google Scholar]
- Aranda, A.; Pascual, A. Nuclear Hormone Receptors and Gene Expression. Physiol. Rev. 2001, 81, 1269–1304. [Google Scholar]
- Medici, N.; Abbondanza, C.; Nigro, V.; Rossi, V.; Piluso, G.; Belsito, A.; Gallo, L.; Roscigno, A.; Bontempo, P.; Puca, A.A.; et al. Identification of a DNA binding protein cooperating with estrogen receptor as RIZ (retinoblastoma interacting zinc finger protein). Biochem. Biophys. Res. Commun. 1999, 264, 983–989. [Google Scholar] [CrossRef]
- Abbondanza, C.; Medici, N.; Nigro, V.; Rossi, V.; Gallo, L.; Piluso, G.; Belsito, A.; Roscigno, A.; Bontempo, P.; Puca, A.A.; et al. The retinoblastoma-interacting zinc-finger protein RIZ is a downstream effector of estrogen action. Proc. Natl. Acad. Sci. USA 2000, 97, 3130–3135. [Google Scholar]
- Gazzerro, P.; Abbondanza, C.; D’Arcangelo, A.; Rossi, M.; Medici, N.; Moncharmont, B.; Puca, G.A. Modulation of RIZ gene expression is associated to estradiol control of MCF-7 breast cancer cell proliferation. Exp. Cell Res. 2006, 312, 340–349. [Google Scholar]
- Muraosa, Y.; Takahashi, K.; Yoshizawa, M.; Shibahara, S. cDNA cloning of a novel protein containing two zinc-finger domains that may function as a transcription factor for the human heme-oxygenase-1 gene. Eur. J. Biochem. 1996, 235, 471–479. [Google Scholar]
- Xie, M.; Shao, G.; Buyse, I.; Huang, S. Transcriptional repression mediated by the PR domain zinc finger gene RIZ. J. Biol. Chem. 1997, 272, 26360–26366. [Google Scholar]
- Carling, T.; Kim, K.-C.; Yang, X.-H.; Gu, J.; Zhang, X.-K.; Huang, S. A Histone Methyltransferase Is Required for Maximal Response to Female Sex Hormones. Mol. Cell. Biol. 2004, 24, 7032–7042. [Google Scholar] [CrossRef]
- Garcia-Bassets, I.; Kwon, Y.S.; Telese, F.; Prefontaine, G.G.; Hutt, K.R.; Cheng, C.S.; Ju, B.G.; Ohgi, K.A.; Wang, J.; Escoubet-Lozach, L.; et al. Histone methylation-dependent mechanisms impose ligand dependency for gene activation by nuclear receptors. Cell 2007, 128, 505–518. [Google Scholar] [CrossRef]
- Metzger, E.; Wissmann, M.; Yin, N.; Muller, J.M.; Schneider, R.; Peters, A.H.F.M.; Gunther, T.; Buettner, R.; Schule, R. LSD1 demethylates repressive histone marks to promote androgen-receptor-dependent transcription. Nature 2005, 437, 436–439. [Google Scholar]
- Ju, B.G.; Lunyak, V.V.; Perissi, V.; Garcia-Bassets, I.; Rose, D.W.; Glass, C.K.; Rosenfeld, M.G. A topoisomerase IIbeta-mediated dsDNA break required for regulated transcription. Science 2006, 312, 1798–1802. [Google Scholar]
- Perillo, B.; Ombra, M.N.; Bertoni, A.; Cuozzo, C.; Sacchetti, S.; Sasso, A.; Chiariotti, L.; Malorni, A.; Abbondanza, C.; Avvedimento, E.V. DNA Oxidation as Triggered by H3K9me2 Demethylation Drives Estrogen-Induced Gene Expression. Science 2008, 319, 202–206. [Google Scholar] [CrossRef]
- Steele-Perkins, G.; Fang, W.; Yang, X.H.; van Gele, M.; Carling, T.; Gu, J.; Buyse, I.M.; Fletcher, J.A.; Liu, J.; Bronson, R.; et al. Tumor formation and inactivation of RIZ1, an Rb-binding member of a nuclear protein-methyltransferase superfamily. Genes Dev. 2001, 15, 2250–2262. [Google Scholar] [CrossRef]
- Buyse, I.M.; Shao, G.; Huang, S. The retinoblastoma protein binds to RIZ, a zinc-finger protein that shares an epitope with the adenovirus E1A protein. Proc. Natl. Acad. Sci. USA 1995, 92, 4467–4471. [Google Scholar]
- Shapiro, V.S.; Lee, P.; Winoto, A. Identification and cloning of the G3B cDNA encoding a 3' segment of a protein binding to GATA-3. Gene 1995, 163, 329–330. [Google Scholar] [CrossRef]
- Zhu, J.; Yamane, H.; Cote-Sierra, J.; Guo, L.; Paul, W.E. GATA-3 promotes Th2 responses through three different mechanisms: induction of Th2 cytokine production, selective growth of Th2 cells and inhibition of Th1 cell-specific factors. Cell Res. 2006, 16, 3–10. [Google Scholar] [CrossRef]
- Abbondanza, C.; de Rosa, C.; D’Arcangelo, A.; Pacifico, M.; Spizuoco, C.; Piluso, G.; di Zazzo, E.; Gazzerro, P.; Medici, N.; Moncharmont, B.; et al. Identification of a functional estrogen-responsive enhancer element in the promoter 2 of PRDM2 gene in breast cancer cell lines. J. Cell. Physiol. 2012, 227, 964–975. [Google Scholar] [CrossRef]
- O’Sullivan, J.M.; Tan-Wong, S.M.; Morillon, A.; Lee, B.; Coles, J.; Mellor, J.; Proudfoot, N.J. Gene loops juxtapose promoters and terminators in yeast. Nat. Genet. 2004, 36, 1014–1018. [Google Scholar] [CrossRef]
- Ansari, A.; Hampsey, M. A role for the CPF 3'-end processing machinery in RNAP II-dependent gene looping. Genes Dev. 2005, 19, 2969–2978. [Google Scholar] [CrossRef]
- Bentley, D.L. Rules of engagement: Co-transcriptional recruitment of pre-mRNA processing factors. Curr. Opin. Cell Biol. 2005, 17, 251–256. [Google Scholar] [CrossRef]
- Abbondanza, C.; de Rosa, C.; Ombra, M.N.; Aceto, F.; Medici, N.; Altucci, L.; Moncharmont, B.; Puca, G.A.; Porcellini, A.; Avvedimento, E.V.; et al. Highlighting chromosome loops in DNA-picked chromatin (DPC). Epigenetics 2011, 6, 979–986. [Google Scholar] [CrossRef]
- Abbondanza, C.; de Nigris, F.; de Rosa, C.; Rossiello, R.; Puca, G.A.; Napoli, C. Silencing of YY1 downregulates RIZ1 promoter in human osteosarcoma. Oncol. Res. 2008, 17, 33–41. [Google Scholar]
- Rossi, V.; Staibano, S.; Pasquali, D.; de Rosa, C.; Mascolo, M.; Bellastella, G.; Visconti, D.; de Bellis, A.; Moncharmont, B.; de Rosa, G.; et al. Expression of RIZ1 protein (Retinoblastoma-interacting zinc-finger protein 1) in prostate cancer epithelial cells changes with cancer grade progression and is modulated in vitro by DHT and E2. J. Cell. Physiol. 2009, 221, 771–777. [Google Scholar] [CrossRef]
- TRANSFAC® 7.0 Public. 2005. Available online: http://www.gene-regulation.com/pub/databases.html/ (accessed on 17 January 2009).
- Gazzerro, P.; Bontempo, P.; Schiavone, E.M.; Abbondanza, C.; Moncharmont, B.; Armetta, I.; Medici, N.; de Simone, M.; Nola, E.; Puca, G.A.; et al. Differentiation of myeloid cell lines correlates with a selective expression of RIZ protein. Mol. Med. 2001, 7, 552–560. [Google Scholar]
- Seale, P.; Bjork, B.; Yang, W.; Kajimura, S.; Chin, S.; Kuang, S.; Scime, A.; Devarakonda, S.; Conroe, H.M.; Erdjument-Bromage, H.; et al. PRDM16 controls a brown fat/skeletal muscle switch. Nature 2008, 454, 961–968. [Google Scholar]
- Eom, G.H.; Kim, K.; Kim, S.-M.; Kee, H.J.; Kim, J.-Y.; Jin, H.M.; Kim, J.-R.; Kim, J.H.; Choe, N.; Kim, K.-B.; et al. Histone methyltransferase PRDM8 regulates mouse testis steroidogenesis. Biochem. Biophys. Res. Commun. 2009, 388, 131–136. [Google Scholar] [CrossRef]
- Pastural, E.; Takahashi, N.; Dong, W.F.; Bainbridge, M.; Hull, A.; Pearson, D.; Huang, S.; Lowsky, R.; DeCoteau, J.F.; Geyer, C.R. RIZ1 repression is associated with insulin-like growth factor-1 signaling activation in chronic myeloid leukemia cell lines. Oncogene 2006, 26, 1586–1594. [Google Scholar]
- Thoenen, H. Neurotrophins and Neuronal Plasticity. Science 1995, 270, 593–598. [Google Scholar]
- Lewin, G.R.; Barde, Y.-A. Physiology of the Neurotrophins. Annu. Rev. Neurosci. 1996, 19, 289–317. [Google Scholar] [CrossRef]
- Chittka, A.; Chao, M.V. Identification of a zinc finger protein whose subcellular distribution is regulated by serum and nerve growth factor. Proc. Natl. Acad. Sci. USA 1999, 96, 10705–10710. [Google Scholar] [CrossRef]
- Chittka, A.; Arevalo, J.C.; Rodriguez-Guzman, M.; Perez, P.; Chao, M.V.; Sendtner, M. The p75NTR-interacting protein SC1 inhibits cell cycle progression by transcriptional repression of cyclin E. J. Cell Biol. 2004, 164, 985–996. [Google Scholar] [CrossRef]
- Chittka, A.; Nitarska, J.; Grazini, U.; Richardson, W.D. Transcription Factor Positive Regulatory Domain 4 (PRDM4) recruits Protein Arginine Methyltransferase 5 (PRMT5) to mediate histone arginine methylation and control neural stem cell proliferation and differentiation. J. Biol. Chem. 2012, 287, 42995–43006. [Google Scholar] [CrossRef]
- Sato, N.; Meijer, L.; Skaltsounis, L.; Greengard, P.; Brivanlou, A.H. Maintenance of pluripotency in human and mouse embryonic stem cells through activation of Wnt signaling by a pharmacological GSK-3-specific inhibitor. Nat. Med. 2004, 10, 55–63. [Google Scholar] [CrossRef]
- Watanabe, S.; Umehara, H.; Murayama, K.; Okabe, M.; Kimura, T.; Nakano, T. Activation of Akt signaling is sufficient to maintain pluripotency in mouse and primate embryonic stem cells. Oncogene 2006, 25, 2697–2707. [Google Scholar] [CrossRef]
- Storm, M.P.; Bone, H.K.; Beck, C.G.; Bourillot, P.-Y.; Schreiber, V.; Damiano, T.; Nelson, A.; Savatier, P.; Welham, M.J. Regulation of Nanog Expression by Phosphoinositide 3-Kinase-dependent Signaling in Murine Embryonic Stem Cells. J. Biol. Chem. 2007, 282, 6265–6273. [Google Scholar]
- Lee, M.Y.; Lim, H.W.; Lee, S.H.; Han, H.J. Smad, PI3K/Akt, and Wnt-Dependent Signaling Pathways Are Involved in BMP-4-Induced ESC Self-Renewal. Stem Cells 2009, 27, 1858–1868. [Google Scholar] [CrossRef]
- Chia, N.-Y.; Chan, Y.-S.; Feng, B.; Lu, X.; Orlov, Y.L.; Moreau, D.; Kumar, P.; Yang, L.; Jiang, J.; Lau, M.-S.; et al. A genome-wide RNAi screen reveals determinants of human embryonic stem cell identity. Nature 2010, 468, 316–320. [Google Scholar]
- Bikoff, E.K.; Robertson, E.J. One PRDM is not enough for germ cell development. Nat. Genet. 2008, 40, 934. [Google Scholar] [CrossRef]
- Fujiwara, T.; Dunn, N.R.; Hogan, B.L.M. Bone morphogenetic protein 4 in the extraembryonic mesoderm is required for allantois development and the localization and survival of primordial germ cells in the mouse. Proc. Natl. Acad. Sci. USA 2001, 98, 13739–13744. [Google Scholar]
- Kimura, T.; Nakamura, T.; Murayama, K.; Umehara, H.; Yamano, N.; Watanabe, S.; Taketo, M.M.; Nakano, T. The stabilization of β-catenin leads to impaired primordial germ cell development via aberrant cell cycle progression. Dev. Biol. 2006, 300, 545–553. [Google Scholar] [CrossRef]
- Kemp, C.; Willems, E.; Abdo, S.; Lambiv, L.; Leyns, L. Expression of all Wnt genes and their secreted antagonists during mouse blastocyst and postimplantation development. Dev. Dyn. 2005, 233, 1064–1075. [Google Scholar] [CrossRef]
- Ohinata, Y.; Ohta, H.; Shigeta, M.; Yamanaka, K.; Wakayama, T.; Saitou, M. A Signaling Principle for the Specification of the Germ Cell Lineage in Mice. Cell 2009, 137, 571–584. [Google Scholar] [CrossRef]
- Arnold, S.J.; Maretto, S.; Islam, A.; Bikoff, E.K.; Robertson, E.J. Dose-dependent Smad1, Smad5 and Smad8 signaling in the early mouse embryo. Dev. Biol. 2006, 296, 104–118. [Google Scholar] [CrossRef]
- Ohinata, Y.; Payer, B.; O’Carroll, D.; Ancelin, K.; Ono, Y.; Sano, M.; Barton, S.C.; Obukhanych, T.; Nussenzweig, M.; Tarakhovsky, A.; et al. Blimp1 is a critical determinant of the germ cell lineage in mice. Nature 2005, 436, 207–213. [Google Scholar]
- Surani, M.A.; Hayashi, K.; Hajkova, P. Genetic and Epigenetic Regulators of Pluripotency. Cell 2007, 128, 747–762. [Google Scholar] [CrossRef]
- Bedford, M.T.; Richard, S. Arginine Methylation: An Emerging Regulatorof Protein Function. Mol. Cell 2005, 18, 263–272. [Google Scholar] [CrossRef]
- Tee, W.W.; Pardo, M.; Theunissen, T.W.; Yu, L.; Choudhary, J.S.; Hajkova, P.; Surani, M.A. Prmt5 is essential for early mouse development and acts in the cytoplasm to maintain ES cell pluripotency. Genes Dev. 2010, 24, 2772–2777. [Google Scholar] [CrossRef]
- Seki, Y.; Yamaji, M.; Yabuta, Y.; Sano, M.; Shigeta, M.; Matsui, Y.; Saga, Y.; Tachibana, M.; Shinkai, Y.; Saitou, M. Cellular dynamics associated with the genome-wide epigenetic reprogramming in migrating primordial germ cells in mice. Development 2007, 134, 2627–2638. [Google Scholar] [CrossRef]
- Saitou, M.; Barton, S.C.; Surani, M.A. A molecular programme for the specification of germ cell fate in mice. Nature 2002, 418, 293–300. [Google Scholar] [CrossRef]
- Yabuta, Y.; Kurimoto, K.; Ohinata, Y.; Seki, Y.; Saitou, M. Gene Expression Dynamics During Germline Specification in Mice Identified by Quantitative Single-Cell Gene Expression Profiling. Biol. Reprod. 2006, 75, 705–716. [Google Scholar]
- Clark, A.T.; Bodnar, M.S.; Fox, M.; Rodriquez, R.T.; Abeyta, M.J.; Firpo, M.T.; Pera, R.A.R. Spontaneous differentiation of germ cells from human embryonic stem cells in vitro. Hum. Mol. Genet. 2004, 13, 727–739. [Google Scholar] [CrossRef]
- Toyooka, Y.; Tsunekawa, N.; Akasu, R.; Noce, T. Embryonic stem cells can form germ cells in vitro. Proc. Natl. Acad. Sci. USA 2003, 100, 11457–11462. [Google Scholar] [CrossRef]
- Yamaji, M.; Seki, Y.; Kurimoto, K.; Yabuta, Y.; Yuasa, M.; Shigeta, M.; Yamanaka, K.; Ohinata, Y.; Saitou, M. Critical function of Prdm14 for the establishment of the germ cell lineage in mice. Nat. Genet. 2008, 40, 1016–1022. [Google Scholar]
- Kurimoto, K.; Yamaji, M.; Seki, Y.; Saitou, M. Specification of the germ cell lineage in mice: A process orchestrated by the PR-domain proteins, Blimp1 and Prdm14. Cell Cycle 2008, 7, 3514–3518. [Google Scholar] [CrossRef]
- Bao, S.; Leitch, H.G.; Gillich, A.; Nichols, J.; Tang, F.; Kim, S.; Lee, C.; Zwaka, T.; Li, X.; Surani, M.A. The Germ Cell Determinant Blimp1 Is Not Required for Derivation of Pluripotent Stem Cells. Cell Stem Cell 2012, 11, 110–117. [Google Scholar] [CrossRef]
- Ying, Y.; Tao, Q. Epigenetic disruption of the WNT/beta-catenin signaling pathway in human cancers. Epigenetics 2009, 4, 307–312. [Google Scholar] [CrossRef]
- Shu, X.; Geng, H.; Li, L.; Ying, J.; Ma, C.; Wang, Y.; Poon, F.F.; Wang, X.; Ying, Y.; Yeo, W.; et al. The epigenetic modifier PRDM5 functions as a tumor suppressor through modulating WNT/β-catenin signaling and is frequently silenced in multiple tumors. PLoS One 2011, 6, e27346. [Google Scholar]
- Nguyen, L.; Besson, A.; Roberts, J.M.; Guillemot, F. Coupling cell cycle exit, neuronal differentiation and migration in cortical neurogenesis. Cell Cycle 2006, 5, 2314–2318. [Google Scholar] [CrossRef]
- Shirasaki, R.; Pfaff, S.L. Transcriptional codes and the control of neuronal identity. .Annu. Rev. Neurosci. 2002, 25, 251–281. [Google Scholar] [CrossRef]
- Bertrand, N.; Castro, D.S.; Guillemot, F. Proneural genes and the specification of neural cell types. Nat. Rev. Neurosci. 2002, 3, 517. [Google Scholar] [CrossRef]
- Ross, S.E.; Greenberg, M.E.; Stiles, C.D. Basic Helix-Loop-Helix Factors in Cortical Development. Neuron 2003, 39, 13–25. [Google Scholar] [CrossRef]
- Bramblett, D.E.; Copeland, N.G.; Jenkins, N.A.; Tsai, M.-J. BHLHB4 Is a bHLH Transcriptional Regulator in Pancreas and Brain That Marks the Dimesencephalic Boundary. Genomics 2002, 79, 402. [Google Scholar] [CrossRef]
- Joshi, P.S.; Molyneaux, B.J.; Feng, L.; Xie, X.; Macklis, J.D.; Gan, L. Bhlhb5 Regulates the Postmitotic Acquisition of Area Identities in Layers II-V of the Developing Neocortex. Neuron 2008, 60, 258. [Google Scholar] [CrossRef]
- Ross, S.E.; Mardinly, A.R.; McCord, A.E.; Zurawski, J.; Cohen, S.; Jung, C.; Hu, L.; Mok, S.I.; Shah, A.; Savner, E.M.; et al. Loss of Inhibitory Interneurons in the Dorsal Spinal Cord and Elevated Itch in Bhlhb5 Mutant Mice. Neuron 2010, 65, 886–898. [Google Scholar] [CrossRef]
- Ohtsuka, T.; Ishibashi, M.; Gradwohl, G.; Nakanishi, S.; Guillemot, F.; Kageyama, R. Hes1 and Hes5 as Notch effectors in mammalian neuronal differentiation. EMBO J. 1999, 18, 2196–2207. [Google Scholar] [CrossRef]
- Nieto, M.; Schuurmans, C.; Britz, O.; Guillemot, F. Neural bHLH Genes Control the Neuronal versus Glial Fate Decision in Cortical Progenitors. Neuron 2001, 29, 401–413. [Google Scholar] [CrossRef]
- Endo, K.; Karim, M.R.; Taniguchi, H.; Krejci, A.; Kinameri, E.; Siebert, M.; Ito, K.; Bray, S.J.; Moore, A.W. Chromatin modification of Notch targets in olfactory receptor neuron diversification. Nat. Neurosci. 2011, 15, 224–233. [Google Scholar] [CrossRef]
- Kinameri, E.; Inoue, T.; Aruga, J.; Imayoshi, I.; Kageyama, R.; Shimogori, T.; Moore, A.W. Prdm Proto-Oncogene Transcription Factor Family Expression and Interaction with the Notch-Hes Pathway in Mouse Neurogenesis. PLoS One 2008, 3, e3859. [Google Scholar]
- Ross, S.E.; McCord, A.E.; Jung, C.; Atan, D.; Mok, S.I.; Hemberg, M.; Kim, T.-K.; Salogiannis, J.; Hu, L.; Cohen, S.; et al. Bhlhb5 and Prdm8 Form a Repressor Complex Involved in Neuronal Circuit Assembly. Neuron 2012, 73, 292–303. [Google Scholar] [CrossRef]
- Ito, Y.; Yeo, J.Y.; Chytil, A.; Han, J.; Bringas, P.; Nakajima, A.; Shuler, C.F.; Moses, H.L.; Chai, Y. Conditional inactivation of Tgfbr2 in cranial neural crest causes cleft palate and calvaria defects. Development 2003, 130, 5269–5280. [Google Scholar]
- Pisano, M.M.; Mukhopadhyay, P.; Greene, R.M. Molecular fingerprinting of TGFß-treated embryonic maxillary mesenchymal cells. Orthodontics Craniofacial Res. 2003, 6, 194. [Google Scholar] [CrossRef]
- D’Angelo, M.; Greene, R.M. Transforming growth factor-beta modulation of glycosaminoglycan production by mesenchymal cells of the developing murine secondary palate. Dev. Biol. 1991, 145, 374–378. [Google Scholar] [CrossRef]
- Mukhopadhyay, P.; Greene, R.M.; Pisano, M.M. Expression profiling of transforming growth factor β superfamily genes in developing orofacial tissue. Birth Defects Res. Part A 2006, 76, 528–543. [Google Scholar] [CrossRef]
- Warner, D.R.; Horn, K.H.; Mudd, L.; Webb, C.L.; Greene, R.M.; Pisano, M.M. PRDM16/MEL1: A novel Smad binding protein expressed in murine embryonic orofacial tissue. Biochim. Biophys. Acta 2007, 1773, 814–820. [Google Scholar]
- Alliston, T.; Ko, T.C.; Cao, Y.; Liang, Y.-Y.; Feng, X.-H.; Chang, C.; Derynck, R. Repression of Bone Morphogenetic Protein and Activin-inducible Transcription by Evi-1. J. Biol. Chem. 2005, 280, 24227–24237. [Google Scholar]
- Bjork, B.C.; Turbe-Doan, A.; Prysak, M.; Herron, B.J.; Beier, D.R. Prdm16 is required for normal palatogenesis in mice. Hum. Mol. Genet. 2010, 19, 774–789. [Google Scholar]
- Sato, T.; Goyama, S.; Nitta, E.; Takeshita, M.; Yoshimi, M.; Nakagawa, M.; Kawazu, M.; Ichikawa, M.; Kurokawa, M. Evi-1 promotes para-aortic splanchnopleural hematopoiesis through up-regulation of GATA-2 and repression of TGF-b signaling. Cancer Sci. 2008, 99, 1407–1413. [Google Scholar]
- Takahata, M.; Inoue, Y.; Tsuda, H.; Imoto, I.; Koinuma, D.; Hayashi, M.; Ichikura, T.; Yamori, T.; Nagasaki, K.; Yoshida, M.; et al. SKI and MEL1 Cooperate to Inhibit Transforming Growth Factor-β Signal in Gastric Cancer Cells. J. Biol. Chem. 2009, 284, 3334–3344. [Google Scholar]
- Jakobsen, L.P.; Borup, R.; Vestergaard, J.; Larsen, L.A.; Lage, K.; Maroun, L.L.; Kjaer, I.; Niemann, C.U.; Andersen, M.; Knudsen, M.A.; et al. Expression analyses of human cleft palate tissue suggest a role for osteopontin and immune related factors in palatal development. Exp. Mol. Med. 2009, 41, 77–85. [Google Scholar] [CrossRef]
- Warner, D.R.; Mukhopadhyay, P.; Webb, C.L.; Greene, R.M.; Pisano, M.M. Chromatin immunoprecipitation-promoter microarray identification of genes regulated by PRDM16 in murine embryonic palate mesenchymal cells. Exp. Biol. Med. (Maywood) 2012, 237, 387–394. [Google Scholar] [CrossRef]
- Ren, B.; Chee, K.J.; Kim, T.H.; Maniatis, T. PRDI-BF1/Blimp-1 repression is mediated by corepressors of the Groucho family of proteins. Genes Dev. 1999, 13, 125–137. [Google Scholar] [CrossRef]
- Diehl, S.A.; Schmidlin, H.; Nagasawa, M.; Blom, B.; Spits, H. IL-6 Triggers IL-21 production by human CD4+ T cells to drive STAT3-dependent plasma cell differentiation in B cells. Immunol. Cell Biol. 2012, 90, 208–211. [Google Scholar]
- Kallies, A.; Hawkins, E.D.; Belz, G.T.; Metcalf, D.; Hommel, M.; Corcoran, L.M.; Hodgkin, P.D.; Nutt, S.L. Transcriptional repressor Blimp-1 is essential for T cell homeostasis and self-tolerance. Nat. Immunol. 2006, 7, 466–474. [Google Scholar]
- Martins, G.A.; Cimmino, L.; Shapiro-Shelef, M.; Szabolcs, M.; Herron, A.; Magnusdottir, E.; Calame, K. Transcriptional repressor Blimp-1 regulates T cell homeostasis and function. Nat. Immunol. 2006, 7, 457–465. [Google Scholar] [CrossRef]
- Gong, D.; Malek, T.R. Cytokine-Dependent Blimp-1 Expression in Activated T Cells Inhibits IL-2 Production. J. Immunol. 2007, 178, 242–252. [Google Scholar]
- Wang, L.; van Panhuys, N.; Hu-Li, J.; Kim, S.; Le Gros, G.; Min, B. Blimp-1 Induced by IL-4 Plays a Critical Role in Suppressing IL-2 Production in Activated CD4 T Cells. J. Immunol. 2008, 181, 5249–5256. [Google Scholar]
- Thiele, S.; Wittmann, J.; Jäck, H.-M.; Pahl, A. miR-9 enhances IL-2 production in activated human CD4+ T cells by repressing Blimp-1. Eur. J. Immunol. 2012, 42, 2100–2108. [Google Scholar] [CrossRef]
- Yu, D.; Rao, S.; Tsai, L.M.; Lee, S.K.; He, Y.; Sutcliffe, E.L.; Srivastava, M.; Linterman, M.; Zheng, L.; Simpson, N.; et al. The Transcriptional Repressor Bcl-6 Directs T Follicular Helper Cell Lineage Commitment. Immunity 2009, 31, 457–468. [Google Scholar] [CrossRef]
- Basso, K.; Dalla-Favera, R. BCL6: Master regulator of the germinal center reaction and key oncogene in B cell lymphomagenesis. Adv. Immunol. 2010, 105, 193–210. [Google Scholar] [CrossRef]
- Oestreich, K.J.; Huang, A.C.; Weinmann, A.S. The lineage-defining factors T-bet and Bcl-6 collaborate to regulate Th1 gene expression patterns. J. Exp. Med. 2011, 208, 1001–1013. [Google Scholar] [CrossRef]
- Oestreich, K.J.; Mohn, S.E.; Weinmann, A.S. Molecular mechanisms that control the expression and activity of Bcl-6 in TH1 cells to regulate flexibility with a TFH-like gene profile. Nat. Immunol. 2012, 13, 405–411. [Google Scholar]
- Cretney, E.; Xin, A.; Shi, W.; Minnich, M.; Masson, F.; Miasari, M.; Belz, G.T.; Smyth, G.K.; Busslinger, M.; Nutt, S.L.; et al. The transcription factors Blimp-1 and IRF4 jointly control the differentiation and function of effector regulatory T cells. Nat. Immunol. 2011, 12, 304–311. [Google Scholar]
- Zheng, Y.; Josefowicz, S.Z.; Kas, A.; Chu, T.-T.; Gavin, M.A.; Rudensky, A.Y. Genome-wide analysis of Foxp3 target genes in developing and mature regulatory T cells. Nature 2007, 445, 936–940. [Google Scholar]
- Zheng, Y.; Chaudhry, A.; Kas, A.; deRoos, P.; Kim, J.M.; Chu, T.-T.; Corcoran, L.; Treuting, P.; Klein, U.; Rudensky, A.Y. Regulatory T-cell suppressor program co-opts transcription factor IRF4 to control TH2 responses. Nature 2009, 458, 351–356. [Google Scholar]
- Kwon, H.; Thierry-Mieg, D.; Thierry-Mieg, J.; Kim, H.-P.; Oh, J.; Tunyaplin, C.; Carotta, S.; Donovan, C.E.; Goldman, M.L.; Tailor, P.; et al. Analysis of Interleukin-21-Induced Prdm1 Gene Regulation Reveals Functional Cooperation of STAT3 and IRF4 Transcription Factors. Immunity 2009, 31, 941–952. [Google Scholar] [CrossRef]
- Sciammas, R.; Shaffer, A.L.; Schatz, J.H.; Zhao, H.; Staudt, L.M.; Singh, H. Graded Expression of Interferon Regulatory Factor-4 Coordinates Isotype Switching with Plasma Cell Differentiation. Immunity 2006, 25, 225–236. [Google Scholar] [CrossRef]
- Noman, A.S.M.; Koide, N.; Iftakhar-E-Khuda, I.; Dagvadorj, J.; Tumurkhuu, G.; Naiki, Y.; Komatsu, T.; Yoshida, T.; Yokochi, T. Retinoblastoma protein-interacting zinc finger 1, a tumor suppressor, augments lipopolysaccharide-induced proinflammatory cytokine production via enhancing nuclear factor-κB activation. Cell. Immunol. 2010, 264, 114–118. [Google Scholar] [CrossRef]
- Shadat, N.M.A.; Koide, N.; Khuda, I.I.E.; Dagvadorj, J.; Tumurkhuu, G.; Naiki, Y.; Komatsu, T.; Yoshida, T.; Yokochi, T. Retinoblastoma Protein-Interacting Zinc Finger 1 (RIZ1) Regulates the Proliferation of Monocytic Leukemia Cells via Activation of p53. Cancer Invest. 2010, 28, 806–812. [Google Scholar] [CrossRef]
- Duan, Z.; Horwitz, M. Gfi-1 Oncoproteins in Hematopoiesis. Hematology 2003, 8, 339–344. [Google Scholar] [CrossRef]
- Hock, H.; Hamblen, M.J.; Rooke, H.M.; Traver, D.; Bronson, R.T.; Cameron, S.; Orkin, S.H. Intrinsic Requirement for Zinc Finger Transcription Factor Gfi-1 in Neutrophil Differentiation. Immunity 2003, 18, 109–120. [Google Scholar] [CrossRef]
- Karsunky, H.; Zeng, H.; Schmidt, T.; Zevnik, B.; Kluge, R.; Schmid, K.W.; Duhrsen, U.; Moroy, T. Inflammatory reactions and severe neutropenia in mice lacking the transcriptional repressor Gfi1. Nat. Genet. 2002, 30, 295–300. [Google Scholar] [CrossRef]
- Person, R.E.; Li, F.-Q.; Duan, Z.; Benson, K.F.; Wechsler, J.; Papadaki, H.A.; Eliopoulos, G.; Kaufman, C.; Bertolone, S.J.; Nakamoto, B.; et al. Mutations in proto-oncogene GFI1 cause human neutropenia and target ELA2. Nat. Genet. 2003, 34, 308–312. [Google Scholar] [CrossRef]
- Duan, Z.; Person, R.E.; Lee, H.-H.; Huang, S.; Donadieu, J.; Badolato, R.; Grimes, H.L.; Papayannopoulou, T.; Horwitz, M.S. Epigenetic Regulation of Protein-Coding and MicroRNA Genes by the Gfi1-Interacting Tumor Suppressor PRDM5. Mol. Cell. Biol. 2007, 27, 6889–6902. [Google Scholar]
- Ségurel, L.; Leffler, E.M.; Przeworski, M. The Case of the Fickle Fingers: How the PRDM9 Zinc Finger Protein Specifies Meiotic Recombination Hotspots in Humans. PLoS Biol. 2011, 9, e1001211. [Google Scholar] [CrossRef]
- Loyola, A.; Bonaldi, T.; Roche, D.; Imhof, A.; Almouzni, G. PTMs on H3 Variants before Chromatin Assembly Potentiate Their Final Epigenetic State. Mol. Cell 2006, 24, 309–316. [Google Scholar] [CrossRef]
- Pinheiro, I.; Margueron, R.; Shukeir, N.; Eisold, M.; Fritzsch, C.; Richter, F.M.; Mittler, G.; Genoud, C.; Goyama, S.; Kurokawa, M.; et al. Prdm3 and Prdm16 are H3K9me1 Methyltransferases Required for Mammalian Heterochromatin Integrity. Cell 2012, 150, 948–960. [Google Scholar] [CrossRef]
- Olsen, B.R.; Reginato, A.M.; Wang, W. Bone development. Annu. Rev. Cell Dev. Biol. 2000, 16, 191–220. [Google Scholar] [CrossRef]
- Galli, G.G.; Honnens de Lichtenberg, K.; Carrara, M.; Hans, W.; Wuelling, M.; Mentz, B.; Multhaupt, H.A.; Fog, C.K.; Jensen, K.T.; Rappsilber, J.; et al. Prdm5 Regulates Collagen Gene Transcription by Association with RNA Polymerase II in Developing Bone. PLoS Genet. 2012, 8, e1002711. [Google Scholar] [CrossRef]
- Shing, D.C.; Trubia, M.; Marchesi, F.; Radaelli, E.; Belloni, E.; Tapinassi, C.; Scanziani, E.; Mecucci, C.; Crescenzi, B.; Lahortiga, I.; et al. Overexpression of sPRDM16 coupled with loss of p53 induces myeloid leukemias in mice. J. Clin. Invest. 2007, 117, 3696–3707. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Di Zazzo, E.; De Rosa, C.; Abbondanza, C.; Moncharmont, B. PRDM Proteins: Molecular Mechanisms in Signal Transduction and Transcriptional Regulation. Biology 2013, 2, 107-141. https://doi.org/10.3390/biology2010107
Di Zazzo E, De Rosa C, Abbondanza C, Moncharmont B. PRDM Proteins: Molecular Mechanisms in Signal Transduction and Transcriptional Regulation. Biology. 2013; 2(1):107-141. https://doi.org/10.3390/biology2010107
Chicago/Turabian StyleDi Zazzo, Erika, Caterina De Rosa, Ciro Abbondanza, and Bruno Moncharmont. 2013. "PRDM Proteins: Molecular Mechanisms in Signal Transduction and Transcriptional Regulation" Biology 2, no. 1: 107-141. https://doi.org/10.3390/biology2010107