Multiple Pathways of Genome Plasticity Leading to Development of Antibiotic Resistance

{kind=link}
Abstract
:1. Introduction
2. Integrons Are Major Contributors to the Development and Dissemination of Multiple Resistances in Bacteria
2.1. Resistance Integrons
2.2. Chromosomal Integrons
3. Connections between Integron Rearrangements and Horizontal Gene Transfer
3.1. Integron Integrase Expression Is Regulated by the SOS Response
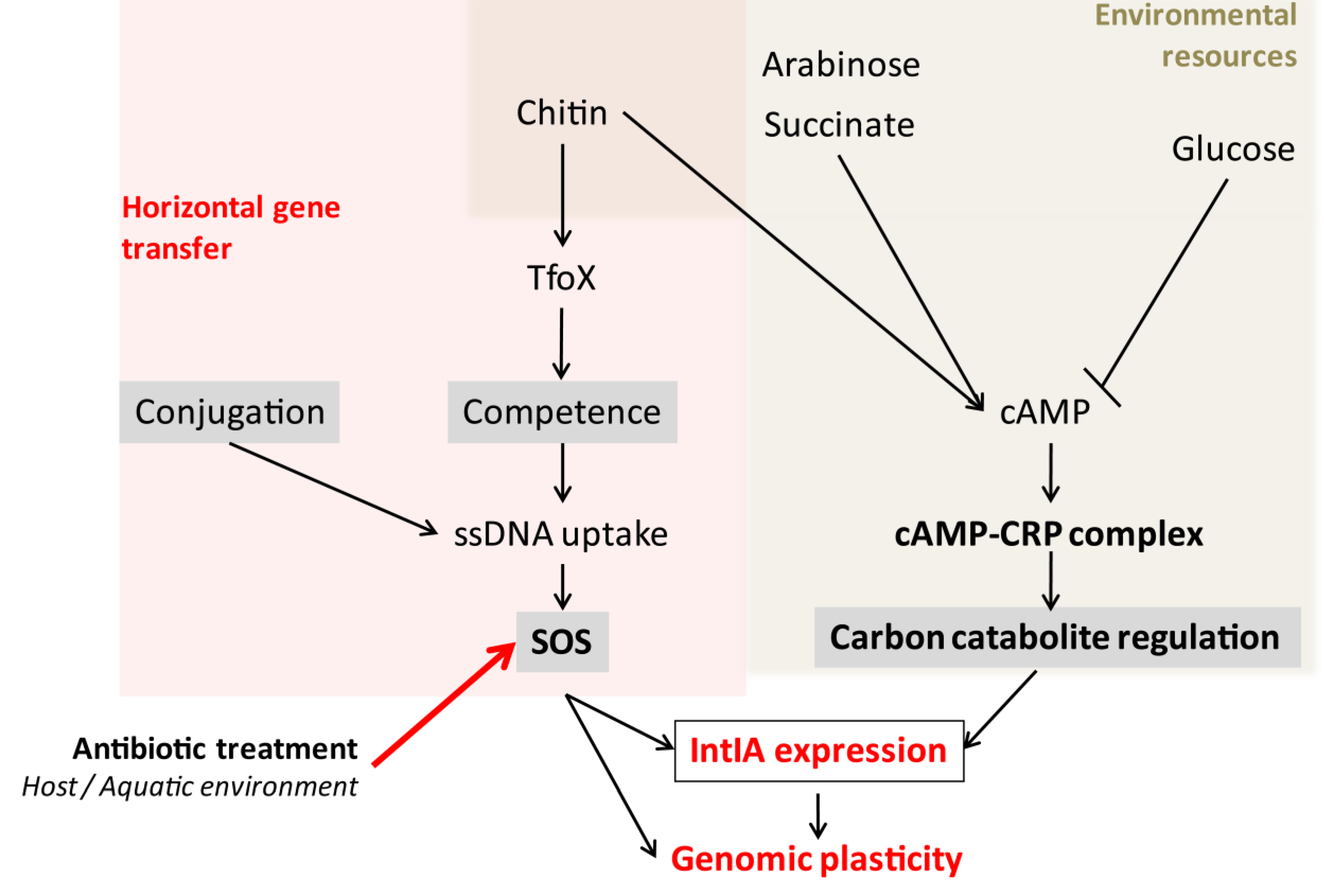
3.2. Conjugation Induces the SOS Response and Integrase Expression
3.3. Natural Transformation induces the SOS Response and Integrase Expression
3.4. Integrating Conjugative Elements, Integron Cassette Rearrangements and the SOS Response
3.4.1. ICE Transfer Induces Integron Integrase Expression
3.4.2. DNA Damaging Agents Increase ICE Plasticity
4. Integron Rearrangements and Cassette Expression are Triggered by Stress Caused by Carbon Source Limitation
4.1. Carbon Catabolite Control Regulates Natural Competence
4.2. Carbon Catabolite Control Regulates SI Integrase Expression
5. Integron Rearrangements are Triggered by Stress Caused by Exposure to Sub-Minimal Inhibitory Antibiotic Concentrations
5.1. General Biological Effects of Sub-MICs of Antibiotics
5.2. Sub-MICs of DNA Damaging Antibiotics Induce the SOS Response
5.3. Sub-MICs of Non-DNA-Damaging Antibiotics Induce the SOS Response in Various Bacterial Species
5.4. Induction of the SOS Response Promotes Acquisition, Maintenance and Spread of Antibiotic Resistances
5.4.1. Acquisition of Antibiotic Resistance
5.4.2. Spread of Antibiotic Resistance
5.4.3. Conservation of Multiple Resistances
5.5. Induction of the SOS Response Promotes the Development of Persister Cells
5.6. Linking the SOS Response, Reactive Oxygen Species Formation and Oxidative Stress Response in the Presence of Sub-MICs of Antibiotics
5.6.1. Sub-MICs of Aminoglycosides Lead to ROS Formation in V. Cholerae
5.6.2. Sub-MIC Driven ROS Formation Leads to Increased Resistance Development
5.6.3. Involvement of RpoS in the Response to Sub-MICs of Antibiotics
6. Conclusions
Acknowledgments
Conflict of Interest
References
- Collis, C.M.; Kim, M.J.; Stokes, H.W.; Hall, R.M. Binding of the purified integron DNA integrase intl1 to integron- and cassette-associated recombination sites. Mol. Microbiol. 1998, 29, 477–490. [Google Scholar] [CrossRef]
- Biskri, L.; Bouvier, M.; Guerout, A.M.; Boisnard, S.; Mazel, D. Comparative study of class 1 integron and Vibrio cholerae superintegron integrase activities. J. Bacteriol. 2005, 187, 1740–1750. [Google Scholar] [CrossRef]
- Bouvier, M.; Demarre, G.; Mazel, D. Integron cassette insertion: A recombination process involving a folded single strand substrate. EMBO J. 2005, 24, 4356–4367. [Google Scholar] [CrossRef]
- Jove, T.; Da Re, S.; Denis, F.; Mazel, D.; Ploy, M.C. Inverse correlation between promoter strength and excision activity in class 1 integrons. PLoS Genet. 2010, 6, e1000793. [Google Scholar] [CrossRef] [Green Version]
- Mazel, D. Integrons: Agents of bacterial evolution. Nat. Rev. Microbiol. 2006, 4, 608–620. [Google Scholar] [CrossRef]
- Rowe-Magnus, D.A.; Mazel, D. Resistance gene capture. Curr. Opin. Microbiol. 1999, 2, 483–488. [Google Scholar] [CrossRef]
- Martinez-Freijo, P.; Fluit, A.C.; Schmitz, F.J.; Grek, V.S.; Verhoef, J.; Jones, M.E. Class I integrons in gram-negative isolates from different european hospitals and association with decreased susceptibility to multiple antibiotic compounds. J. Antimicrob. Chemother. 1998, 42, 689–696. [Google Scholar] [CrossRef]
- Stokes, H.W.; Hall, R.M. A novel family of potentially mobile DNA elements encoding site-specific gene-integration functions: Integrons. Mol. Microbiol. 1989, 3, 1669–1683. [Google Scholar] [CrossRef]
- Liebert, C.A.; Hall, R.M.; Summers, A.O. Transposon Tn21, flagship of the floating genome. Microbiol. Mol. Biol. Rev. 1999, 63, 507–522. [Google Scholar]
- Poirel, L.; Menuteau, O.; Agoli, N.; Cattoen, C.; Nordmann, P. Outbreak of extended-spectrum beta-lactamase veb-1-producing isolates of Acinetobacter baumannii in a French hospital. J. Clin. Microbiol. 2003, 41, 3542–3547. [Google Scholar] [CrossRef]
- Skurnik, D.; Ruimy, R.; Andremont, A.; Amorin, C.; Rouquet, P.; Picard, B.; Denamur, E. Effect of human vicinity on antimicrobial resistance and integrons in animal faecal Escherichia coli. J. Antimicrob. Chemother. 2006, 57, 1215–1219. [Google Scholar] [CrossRef]
- Fluit, A.C.; Schmitz, F.J. Resistance integrons and super-integrons. Clin. Microbiol. Infect. 2004, 10, 272–288. [Google Scholar] [CrossRef]
- Rowe-Magnus, D.A.; Guerout, A.M.; Mazel, D. Bacterial resistance evolution by recruitment of super-integron gene cassettes. Mol. Microbiol. 2002, 43, 1657–1669. [Google Scholar] [CrossRef]
- Partridge, S.R.; Tsafnat, G.; Coiera, E.; Iredell, J.R. Gene cassettes and cassette arrays in mobile resistance integrons. FEMS Microbiol. Rev. 2009, 33, 757–784. [Google Scholar] [CrossRef]
- Mazel, D.; Dychinco, B.; Webb, V.A.; Davies, J. A distinctive class of integron in the Vibrio cholerae genome. Science 1998, 280, 605–608. [Google Scholar] [CrossRef]
- Rowe-Magnus, D.A.; Guerout, A.M.; Ploncard, P.; Dychinco, B.; Davies, J.; Mazel, D. The evolutionary history of chromosomal super-integrons provides an ancestry for multiresistant integrons. Proc. Natl. Acad. Sci. USA 2001, 98, 652–657. [Google Scholar]
- Wu, Y.W.; Rho, M.; Doak, T.G.; Ye, Y. Oral Spirochetes implicated in dental diseases are widespread in normal human subjects and carry extremely diverse integron gene cassettes. Appl. Environ. Microbiol. 2012, 78, 5288–5296. [Google Scholar] [CrossRef]
- Martinez, E.; Marquez, C.; Ingold, A.; Merlino, J.; Djordjevic, S.P.; Stokes, H.W.; Chowdhury, P.R. Diverse mobilized class 1 integrons are common in the chromosomes of pathogenic Pseudomonas aeruginosa clinical isolates. Antimicrob. Agents Chemother. 2012, 56, 2169–2172. [Google Scholar] [CrossRef]
- Ruiz, E.; Saenz, Y.; Zarazaga, M.; Rocha-Gracia, R.; Martinez-Martinez, L.; Arlet, G.; Torres, C. Qnr, aac(6')-ib-cr and qepA genes in Escherichia coli and Klebsiella spp.: Genetic environments and plasmid and chromosomal location. J. Antimicrob. Chemother. 2012, 67, 886–897. [Google Scholar]
- Coleman, N.; Tetu, S.; Wilson, N.; Holmes, A. An unusual integron in Treponema denticola. Microbiology 2004, 150, 3524–3526. [Google Scholar] [CrossRef]
- Gillings, M.; Boucher, Y.; Labbate, M.; Holmes, A.; Krishnan, S.; Holley, M.; Stokes, H.W. The evolution of class 1 integrons and the rise of antibiotic resistance. J. Bacteriol. 2008, 190, 5095–5100. [Google Scholar] [CrossRef]
- Le Roux, F.; Zouine, M.; Chakroun, N.; Binesse, J.; Saulnier, D.; Bouchier, C.; Zidane, N.; Ma, L.; Rusniok, C.; Lajus, A.; et al. Genome sequence of Vibrio splendidus: An abundant planctonic marine species with a large genotypic diversity. Environ. Microbiol. 2009, 11, 1959–1970. [Google Scholar] [CrossRef]
- Melano, R.; Petroni, A.; Garutti, A.; Saka, H.A.; Mange, L.; Pasteran, F.; Rapoport, M.; Rossi, A.; Galas, M. New carbenicillin-hydrolyzing beta-lactamase (carb-7) from Vibrio cholerae non-O1, non-O139 strains encoded by the vcr region of the V. cholerae genome. Antimicrob. Agents Chemother. 2002, 46, 2162–2168. [Google Scholar] [CrossRef]
- Petroni, A.; Melano, R.G.; Saka, H.A.; Garutti, A.; Mange, L.; Pasteran, F.; Rapoport, M.; Miranda, M.; Faccone, D.; Rossi, A.; et al. Carb-9, a carbenicillinase encoded in the vcr region of Vibrio cholerae non-O1, non-O139 belongs to a family of cassette-encoded beta-lactamases. Antimicrob. Agents Chemother. 2004, 48, 4042–4046. [Google Scholar] [CrossRef]
- Fonseca, E.L.; Dos Santos Freitas, F.; Vieira, V.V.; Vicente, A.C. New qnr gene cassettes associated with superintegron repeats in Vibrio cholerae O1. Emerg. Infect. Dis. 2008, 14, 1129–1131. [Google Scholar] [CrossRef]
- Gassama Sow, A.; Aidara-Kane, A.; Barraud, O.; Gatet, M.; Denis, F.; Ploy, M.C. High prevalence of trimethoprim-resistance cassettes in class 1 and 2 integrons in senegalese shigella spp isolates. J. Infect. Dev. Ctries. 2010, 4, 207–212. [Google Scholar]
- Walker, G.C. The SOS Response of Escherichia coli. In Escherichia coli and Salmonella: Cellular and Molecular Biology; Neidhardt, F.C., Curtiss, R., III, Ingraham, J.L., Lin, E.C.C., Low, K.B., Magasanik, B., Reznikoff, W.S., Riley, M., Schaechter, M., Umbarger, H.E., Eds.; American Society of Microbiology: Washington, DC, USA, 1996; pp. 1400–1416. [Google Scholar]
- Wade, J.T.; Reppas, N.B.; Church, G.M.; Struhl, K. Genomic analysis of LlexA binding reveals the permissive nature of the Escherichia coli genome and identifies unconventional target sites. Genes. Dev. 2005, 19, 2619–2630. [Google Scholar] [CrossRef]
- Guerin, E.; Cambray, G.; Sanchez-Alberola, N.; Campoy, S.; Erill, I.; Da Re, S.; Gonzalez-Zorn, B.; Barbe, J.; Ploy, M.C.; Mazel, D. The SOS response controls integron recombination. Science 2009, 324, 1034. [Google Scholar] [CrossRef]
- Cambray, G.; Sanchez-Alberola, N.; Campoy, S.; Guerin, E.; Da Re, S.; Gonzalez-Zorn, B.; Ploy, M.C.; Barbe, J.; Mazel, D.; Erill, I. Prevalence of SOS-mediated control of integron integrase expression as an adaptive trait of chromosomal and mobile integrons. Mob. DNA 2011, 2, e6. [Google Scholar] [CrossRef] [Green Version]
- Baharoglu, Z.; Krin, E.; Mazel, D. Transformation-induced SOS regulation and carbon catabolite control of the V. cholerae integron integrase: Connecting environment and genome plasticity. J. Bacteriol. 2012. [Google Scholar] [CrossRef]
- Boucher, Y.; Cordero, O.X.; Takemura, A.; Hunt, D.E.; Schliep, K.; Bapteste, E.; Lopez, P.; Tarr, C.L.; Polz, M.F. Local mobile gene pools rapidly cross species boundaries to create endemicity within global Vibrio cholerae populations. MBio 2011, 2, e00335-10. [Google Scholar]
- Baharoglu, Z.; Bikard, D.; Mazel, D. Conjugative DNA transfer induces the bacterial SOS response and promotes antibiotic resistance development through integron activation. PLoS Genet. 2010, 6, e1001165. [Google Scholar] [CrossRef]
- Matic, I.; Rayssiguier, C.; Radman, M. Interspecies gene exchange in bacteria: The role of SOS and mismatch repair systems in evolution of species. Cell 1995, 80, 507–515. [Google Scholar] [CrossRef]
- Matic, I.; Taddei, F.; Radman, M. No genetic barriers between Salmonella enterica serovar typhimurium and Escherichia coli in SOS-induced mismatch repair-deficient cells. J. Bacteriol. 2000, 182, 5922–5924. [Google Scholar] [CrossRef]
- Delmas, S.; Matic, I. Cellular response to horizontally transferred DNA in Escherichia coli is tuned by DNA repair systems. DNA Repair 2005, 4, 221–229. [Google Scholar] [CrossRef]
- Smorawinska, M.; Szuplewska, M.; Zaleski, P.; Wawrzyniak, P.; Maj, A.; Plucienniczak, A.; Bartosik, D. Mobilizable narrow host range plasmids as natural suicide vectors enabling horizontal gene transfer among distantly related bacterial species. FEMS Microbiol. Lett. 2012, 326, 76–82. [Google Scholar] [CrossRef]
- Colwell, R.R. A Global and Historical Perspective of the Genus Vibrio. In The Biology of Vibrios; Thompson, F.L., Ed.; ASM Press: Washington, DC, USA, 2006. [Google Scholar]
- Urakawa, H.; Rivera, I.N.G. Aquatic Environmen. In The Biology of Vibrios; Thompson, F.L., Ed.; ASM Press: Washington, DC, USA, 2006. [Google Scholar]
- Bryan, L.E.; Shahrabadi, M.S.; van den Elzen, H.M. Gentamicin resistance in Pseudomonas aeruginosa: R-factor-mediated resistance. Antimicrob. Agents Chemother. 1974, 6, 191–199. [Google Scholar] [CrossRef]
- Kontomichalou, P.; Mitani, M.; Clowes, R.C. Circular R-factor molecules controlling penicillinase synthesis, replicating in Escherichia coli under either relaxed or stringent control. J. Bacteriol. 1970, 104, 34–44. [Google Scholar]
- Moura, A.; Henriques, I.; Smalla, K.; Correia, A. Wastewater bacterial communities bring together broad-host range plasmids, integrons and a wide diversity of uncharacterized gene cassettes. Res. Microbiol. 2010, 161, 58–66. [Google Scholar] [CrossRef]
- Moura, A.; Oliveira, C.; Henriques, I.; Smalla, K.; Correia, A. Broad diversity of conjugative plasmids in integron-carrying bacteria from wastewater environments. FEMS Microbiol. Lett. 2012, 330, 157–164. [Google Scholar] [CrossRef]
- Stecher, B.; Denzler, R.; Maier, L.; Bernet, F.; Sanders, M.J.; Pickard, D.J.; Barthel, M.; Westendorf, A.M.; Krogfelt, K.A.; Walker, A.W.; et al. Gut inflammation can boost horizontal gene transfer between pathogenic and commensal Enterobacteriaceae. Proc. Natl. Acad. Sci. USA 2012, 109, 1269–1274. [Google Scholar] [CrossRef]
- Dubnau, D. DNA uptake in bacteria. Annu. Rev. Microbiol. 1999, 53, 217–244. [Google Scholar] [CrossRef]
- Claverys, J.P.; Martin, B.; Polard, P. The genetic transformation machinery: Composition, localization, and mechanism. FEMS Microbiol. Rev. 2009, 33, 643–656. [Google Scholar] [CrossRef]
- Yasbin, R.E.; Wilson, G.A.; Young, F.E. Transformation and transfection in lysogenic strains of Bacillus subtilis: Evidence for selective induction of prophage in competent cells. J. Bacteriol. 1975, 121, 296–304. [Google Scholar]
- Seitz, P.; Blokesch, M. Cues and regulatory pathways involved in natural competence and transformation in pathogenic and environmental gram-negative bacteria. FEMS Microbiol. Rev. 2012, 37, 336–363. [Google Scholar] [CrossRef]
- Charpentier, X.; Polard, P.; Claverys, J.P. Induction of competence for genetic transformation by antibiotics: Convergent evolution of stress responses in distant bacterial species lacking SOS? Curr. Opin. Microbiol. 2012, 15, 570–576. [Google Scholar]
- Meibom, K.L.; Blokesch, M.; Dolganov, N.A.; Wu, C.Y.; Schoolnik, G.K. Chitin induces natural competence in Vibrio cholerae. Science 2005, 310, 1824–1827. [Google Scholar] [CrossRef]
- Lo Scrudato, M.; Blokesch, M. The regulatory network of natural competence and transformation of Vibrio cholerae. PLoS Genet. 2012, 8, e1002778. [Google Scholar] [CrossRef]
- Zulty, J.J.; Barcak, G.J. Identification of a DNA transformation gene required for com101a+ expression and supertransformer phenotype in Haemophilus influenzae. Proc. Natl. Acad. Sci. USA 1995, 92, 3616–3620. [Google Scholar] [CrossRef]
- Yamamoto, S.; Morita, M.; Izumiya, H.; Watanabe, H. Chitin disaccharide (Glcnac)(2) induces natural competence in Vibrio cholerae through transcriptional and translational activation of a positive regulatory gene tfoX(vc). Gene 2010, 457, 42–49. [Google Scholar] [CrossRef]
- Karudapuram, S.; Barcak, G.J. The haemophilus influenzae dprABC genes constitute a competence-inducible operon that requires the product of the tfoX (sxy) gene for transcriptional activation. J. Bacteriol. 1997, 179, 4815–4820. [Google Scholar]
- Cameron, A.D.; Volar, M.; Bannister, L.A.; Redfield, R.J. RNA secondary structure regulates the translation of sxy and competence development in Haemophilus influenzae. Nucleic Acids Res. 2008, 36, 10–20. [Google Scholar] [CrossRef]
- Bosse, J.T.; Sinha, S.; Schippers, T.; Kroll, J.S.; Redfield, R.J.; Langford, P.R. Natural competence in strains of Actinobacillus pleuropneumoniae. FEMS Microbiol. Lett. 2009, 298, 124–130. [Google Scholar] [CrossRef]
- Redfield, R.J.; Cameron, A.D.; Qian, Q.; Hinds, J.; Ali, T.R.; Kroll, J.S.; Langford, P.R. A novel CRP-dependent regulon controls expression of competence genes in Haemophilus influenzae. J. Mol. Biol. 2005, 347, 735–747. [Google Scholar] [CrossRef]
- Meibom, K.L.; Li, X.B.; Nielsen, A.T.; Wu, C.Y.; Roseman, S.; Schoolnik, G.K. The Vibrio cholerae chitin utilization program. Proc. Natl. Acad. Sci. USA 2004, 101, 2524–2529. [Google Scholar]
- Blokesch, M.; Schoolnik, G.K. The extracellular nuclease Dns and its role in natural transformation of Vibrio cholerae. J. Bacteriol. 2008, 190, 7232–7240. [Google Scholar] [CrossRef]
- Domingues, S.; Harms, K.; Fricke, W.F.; Johnsen, P.J.; da Silva, G.J.; Nielsen, K.M. Natural transformation facilitates transfer of transposons, integrons and gene cassettes between bacterial species. PLoS Pathog. 2012, 8, e1002837. [Google Scholar] [CrossRef]
- Singletary, L.A.; Gibson, J.L.; Tanner, E.J.; McKenzie, G.J.; Lee, P.L.; Gonzalez, C.; Rosenberg, S.M. An SOS-regulated type 2 toxin-antitoxin system. J. Bacteriol. 2009, 191, 7456–7465. [Google Scholar] [CrossRef]
- Pandey, D.P.; Gerdes, K. Toxin-antitoxin loci are highly abundant in free-living but lost from host-associated prokaryotes. Nucleic Acids Res. 2005, 33, 966–976. [Google Scholar] [CrossRef]
- Hochhut, B.; Lotfi, Y.; Mazel, D.; Faruque, S.M.; Woodgate, R.; Waldor, M.K. Molecular analysis of antibiotic resistance gene clusters in Vibrio cholerae O139 and O1 SXT constins. Antimicrob. Agents Chemother. 2001, 45, 2991–3000. [Google Scholar] [CrossRef]
- Wozniak, R.A.; Fouts, D.E.; Spagnoletti, M.; Colombo, M.M.; Ceccarelli, D.; Garriss, G.; Dery, C.; Burrus, V.; Waldor, M.K. Comparative ICE genomics: Insights into the evolution of the SXT/R391 family of ICEs. PLoS Genet. 2009, 5, e1000786. [Google Scholar] [CrossRef]
- Toleman, M.A.; Bennett, P.M.; Walsh, T.R. ISCR elements: Novel gene-capturing systems of the 21st century? Microbiol. Mol. Biol. Rev. 2006, 70, 296–316. [Google Scholar] [CrossRef]
- Baharoglu, Z.; Mazel, D. Vibrio cholerae triggers SOS and mutagenesis in response to a wide range of antibiotics, a route towards multi-resistance. Antimicrob. Agents Chemother. 2011, 55, 2438–2441. [Google Scholar] [CrossRef]
- Wozniak, R.A.; Waldor, M.K. Integrative and conjugative elements: Mosaic mobile genetic elements enabling dynamic lateral gene flow. Nat. Rev. Microbiol. 2010, 8, 552–563. [Google Scholar] [CrossRef]
- Garriss, G.; Burrus, V. Integrating Conjugative Elements of the SXT/R391 Family. In Bacterial Integrative Mobile Genetic Elements; Roberts, A.P., Mullany, P., Eds.; Landes Bioscience: Austin, TX, USA, 2013. [Google Scholar]
- Wozniak, R.A.; Waldor, M.K. A toxin-antitoxin system promotes the maintenance of an integrative conjugative element. PLoS Genet. 2009, 5, e1000439. [Google Scholar] [CrossRef]
- Dziewit, L.; Jazurek, M.; Drewniak, L.; Baj, J.; Bartosik, D. The SXT conjugative element and linear prophage N15 encode toxin-antitoxin-stabilizing systems homologous to the tad-ata module of the Paracoccus aminophilus plasmid pAMI2. J. Bacteriol. 2007, 189, 1983–1997. [Google Scholar] [CrossRef]
- Bordeleau, E.; Brouillette, E.; Robichaud, N.; Burrus, V. Beyond antibiotic resistance: Integrating conjugative elements of the SXT/R391 family that encode novel diguanylate cyclases participate to c-di-GMP signalling in Vibrio cholerae. Environ. Microbiol. 2009, 12, 510–523. [Google Scholar]
- Faruque, S.M.; Biswas, K.; Udden, S.M.; Ahmad, Q.S.; Sack, D.A.; Nair, G.B.; Mekalanos, J.J. Transmissibility of cholera: In vivo-formed biofilms and their relationship to infectivity and persistence in the environment. Proc. Natl. Acad. Sci. USA 2006, 103, 6350–6355. [Google Scholar]
- Butler, S.M.; Camilli, A. Both chemotaxis and net motility greatly influence the infectivity of vibrio cholerae. Proc. Natl. Acad. Sci. USA 2004, 101, 5018–5023. [Google Scholar] [CrossRef]
- Beaber, J.W.; Hochhut, B.; Waldor, M.K. SOS response promotes horizontal dissemination of antibiotic resistance genes. Nature 2004, 427, 72–74. [Google Scholar]
- Burrus, V.; Waldor, M.K. Control of SXT integration and excision. J. Bacteriol. 2003, 185, 5045–5054. [Google Scholar] [CrossRef]
- Cambray, G.; Guerout, A.M.; Mazel, D. Integrons. Annu. Rev. Genet. 2010, 44, 141–166. [Google Scholar] [CrossRef]
- Garriss, G.; Waldor, M.K.; Burrus, V. Mobile antibiotic resistance encoding elements promote their own diversity. PLoS Genet. 2009, 5, e1000775. [Google Scholar] [CrossRef]
- Garriss, G.; Poulin-Laprade, D.; Burrus, V. DNA damaging agents induce the RecA-independent homologous recombination functions of integrating conjugative elements of the SXT/R391 family. J. Bacteriol. 2013, 195, 1991–2003. [Google Scholar] [CrossRef]
- Hochhut, B.; Beaber, J.W.; Woodgate, R.; Waldor, M.K. Formation of chromosomal tandem arrays of the SXT element and R391, two conjugative chromosomally integrating elements that share an attachment site. J. Bacteriol. 2001, 183, 1124–1132. [Google Scholar] [CrossRef]
- Burrus, V.; Waldor, M.K. Formation of SXT tandem arrays and SXT-R391 hybrids. J. Bacteriol. 2004, 186, 2636–2645. [Google Scholar] [CrossRef]
- Galimand, M.; Guiyoule, A.; Gerbaud, G.; Rasoamanana, B.; Chanteau, S.; Carniel, E.; Courvalin, P. Multidrug resistance in Yersinia pestis mediated by a transferable plasmid. N. Engl. J. Med. 1997, 337, 677–680. [Google Scholar] [CrossRef]
- Lindsey, R.L.; Fedorka-Cray, P.J.; Frye, J.G.; Meinersmann, R.J. IncA/C plasmids are prevalent in multidrug-resistant Salmonella enterica isolates. Appl. Environ. Microbiol. 2009, 75, 1908–1915. [Google Scholar] [CrossRef]
- Fricke, W.F.; Welch, T.J.; McDermott, P.F.; Mammel, M.K.; LeClerc, J.E.; White, D.G.; Cebula, T.A.; Ravel, J. Comparative genomics of the IncA/C multidrug resistance plasmid family. J. Bacteriol. 2009, 191, 4750–4757. [Google Scholar] [CrossRef]
- Pan, J.C.; Ye, R.; Wang, H.Q.; Xiang, H.Q.; Zhang, W.; Yu, X.F.; Meng, D.M.; He, Z.S. Vibrio cholerae O139 multiple-drug resistance mediated by Yersinia pestis pIP1202-like conjugative plasmids. Antimicrob. Agents Chemother. 2008, 52, 3829–3836. [Google Scholar] [CrossRef]
- Welch, T.J.; Fricke, W.F.; McDermott, P.F.; White, D.G.; Rosso, M.L.; Rasko, D.A.; Mammel, M.K.; Eppinger, M.; Rosovitz, M.J.; Wagner, D.; et al. Multiple antimicrobial resistance in plague: An emerging public health risk. PLoS One 2007, 2, e309. [Google Scholar] [CrossRef]
- Carattoli, A.; Villa, L.; Poirel, L.; Bonnin, R.A.; Nordmann, P. Evolution of IncA/C blacmy-2-carrying plasmids by acquisition of the blandm-1 carbapenemase gene. Antimicrob. Agents Chemother. 2011, 56, 783–786. [Google Scholar]
- Johnson, T.J.; Lang, K.S. IncA/C plasmids: An emerging threat to human and animal health? Mob. Genet. Elem. 2012, 2, 55–58. [Google Scholar] [CrossRef]
- Bruckner, R.; Titgemeyer, F. Carbon catabolite repression in bacteria: Choice of the carbon source and autoregulatory limitation of sugar utilization. FEMS Microbiol. Lett. 2002, 209, 141–148. [Google Scholar] [CrossRef]
- Kolb, A.; Busby, S.; Buc, H.; Garges, S.; Adhya, S. Transcriptional regulation by cAMP and its receptor protein. Annu. Rev. Biochem. 1993, 62, 749–795. [Google Scholar] [CrossRef]
- Stulke, J.; Hillen, W. Carbon catabolite repression in bacteria. Curr. Opin. Microbiol. 1999, 2, 195–201. [Google Scholar] [CrossRef]
- Cameron, A.D.; Redfield, R.J. Non-canonical crp sites control competence regulons in Escherichia coli and many other gamma-proteobacteria. Nucleic Acids Res. 2006, 34, 6001–6014. [Google Scholar] [CrossRef]
- de Crombrugghe, B.; Busby, S.; Buc, H. Cyclic AMP receptor protein: Role in transcription activation. Science 1984, 224, 831–838. [Google Scholar]
- Rojo, F. Carbon catabolite repression in Pseudomonas: Optimizing metabolic versatility and interactions with the environment. FEMS Microbiol. Rev. 2010, 34, 658–684. [Google Scholar]
- Gorke, B.; Stulke, J. Carbon catabolite repression in bacteria: Many ways to make the most out of nutrients. Nat. Rev. Microbiol. 2008, 6, 613–624. [Google Scholar] [CrossRef]
- Gosset, G.; Zhang, Z.; Nayyar, S.; Cuevas, W.A.; Saier, M.H., Jr. Transcriptome analysis of CRP-dependent catabolite control of gene expression in Escherichia coli. J. Bacteriol. 2004, 186, 3516–3524. [Google Scholar] [CrossRef]
- Liang, W.; Pascual-Montano, A.; Silva, A.J.; Benitez, J.A. The cyclic AMP receptor protein modulates quorum sensing, motility and multiple genes that affect intestinal colonization in Vibrio cholerae. Microbiology 2007, 153, 2964–2975. [Google Scholar] [CrossRef]
- Zheng, D.; Constantinidou, C.; Hobman, J.L.; Minchin, S.D. Identification of the CRP regulon using in vitro and in vivo transcriptional profiling. Nucleic Acids Res. 2004, 32, 5874–5893. [Google Scholar] [CrossRef]
- Kumar, S.; Srivastava, S. Cyclic AMP and its receptor protein are required for expression of transfer genes of conjugative plasmid f in Escherichia coli. Mol. Gen. Genet. 1983, 190, 27–34. [Google Scholar] [CrossRef]
- Skorupski, K.; Taylor, R.K. Cyclic AMP and its receptor protein negatively regulate the coordinate expression of cholera toxin and toxin-coregulated pilus in Vibrio cholerae. Proc. Natl. Acad. Sci. USA 1997, 94, 265–270. [Google Scholar] [CrossRef]
- Silva, A.J.; Benitez, J.A. Transcriptional regulation of Vibrio cholerae hemagglutinin/protease by the cyclic AMP receptor protein and RpoS. J. Bacteriol. 2004, 186, 6374–6382. [Google Scholar] [CrossRef]
- Sinha, S.; Cameron, A.D.; Redfield, R.J. Sxy induces a crp-s regulon in Escherichia coli. J. Bacteriol. 2009, 191, 5180–5195. [Google Scholar] [CrossRef]
- Sinha, S.; Mell, J.C.; Redfield, R.J. Seventeen Sxy-dependent cyclic AMP receptor protein site-regulated genes are needed for natural transformation in Haemophilus influenzae. J. Bacteriol. 2012, 194, 5245–5254. [Google Scholar] [CrossRef]
- Blokesch, M. Chitin colonization, chitin degradation and chitin-induced natural competence of Vibrio cholerae are subject to catabolite repression. Environ. Microbiol. 2012, 14, 1898–1912. [Google Scholar] [CrossRef]
- Botsford, J.L.; Harman, J.G. Cyclic AMP in prokaryotes. Microbiol. Rev. 1992, 56, 100–122. [Google Scholar]
- Liu, Y.C.; Huang, W.K.; Huang, T.S.; Kunin, C.M. Detection of antimicrobial activity in urine for epidemiologic studies of antibiotic use. J. Clin. Epidemiol. 1999, 52, 539–545. [Google Scholar] [CrossRef]
- Haggard, B.E.; Bartsch, L.D. Net changes in antibiotic concentrations downstream from an effluent discharge. J. Environ. Qual. 2009, 38, 343–352. [Google Scholar] [CrossRef]
- Fick, J.; Soderstrom, H.; Lindberg, R.H.; Phan, C.; Tysklind, M.; Larsson, D.G. Contamination of surface, ground, and drinking water from pharmaceutical production. Environ. Toxicol. Chem. 2009, 28, 2522–2527. [Google Scholar] [CrossRef]
- Kummerer, K. Antibiotics in the aquatic environment—A review—Part I. position="float" 2009, 75, 417–434. [Google Scholar] [CrossRef]
- Kummerer, K. Antibiotics in the aquatic environment—A review—Part II. position="float" 2009, 75, 435–441. [Google Scholar] [CrossRef]
- Hughes, D.; Andersson, D.I. Selection of resistance at lethal and non-lethal antibiotic concentrations. Curr. Opin. Microbiol. 2012, 15, 555–560. [Google Scholar] [CrossRef]
- Goh, E.B.; Yim, G.; Tsui, W.; McClure, J.; Surette, M.G.; Davies, J. Transcriptional modulation of bacterial gene expression by subinhibitory concentrations of antibiotics. Proc. Natl. Acad. Sci. USA 2002, 99, 17025–17030. [Google Scholar]
- Bernier, S.P.; Surette, M.G. Concentration-dependent activity of antibiotics in natural environments. Front. Microbiol. 2013, 4, e20. [Google Scholar]
- Davies, J.; Spiegelman, G.B.; Yim, G. The world of subinhibitory antibiotic concentrations. Curr. Opin. Microbiol. 2006, 9, 445–453. [Google Scholar] [CrossRef]
- Yim, G.; de la Cruz, F.; Spiegelman, G.B.; Davies, J. Transcription modulation of Salmonella enterica serovar typhimurium promoters by sub-mic levels of rifampin. J. Bacteriol. 2006, 188, 7988–7991. [Google Scholar] [CrossRef]
- Yim, G.; McClure, J.; Surette, M.G.; Davies, J.E. Modulation of Salmonella gene expression by subinhibitory concentrations of quinolones. J. Antibiot. (Tokyo) 64, 73–78.
- Tsui, W.H.; Yim, G.; Wang, H.H.; McClure, J.E.; Surette, M.G.; Davies, J. Dual effects of Mls antibiotics: Transcriptional modulation and interactions on the ribosome. Chem. Biol. 2004, 11, 1307–1316. [Google Scholar] [CrossRef]
- Linares, J.F.; Gustafsson, I.; Baquero, F.; Martinez, J.L. Antibiotics as intermicrobial signaling agents instead of weapons. Proc. Natl. Acad. Sci. USA 2006, 103, 19484–19489. [Google Scholar] [CrossRef]
- Kaplan, J.B. Antibiotic-induced biofilm formation. Int. J. Artif. Organs 2011, 34, 737–751. [Google Scholar] [CrossRef]
- Kaplan, J.B.; Jabbouri, S.; Sadovskaya, I. Extracellular DNA-dependent biofilm formation by Staphylococcus epidermidis rp62a in response to subminimal inhibitory concentrations of antibiotics. Res. Microbiol. 2011, 162, 535–541. [Google Scholar] [CrossRef]
- Haddadin, R.N.; Saleh, S.; Al-Adham, I.S.; Buultjens, T.E.; Collier, P.J. The effect of subminimal inhibitory concentrations of antibiotics on virulence factors expressed by Staphylococcus aureus biofilms. J. Appl. Microbiol. 2010, 108, 1281–1291. [Google Scholar] [CrossRef]
- Hoffman, L.R.; D'Argenio, D.A.; MacCoss, M.J.; Zhang, Z.; Jones, R.A.; Miller, S.I. Aminoglycoside antibiotics induce bacterial biofilm formation. Nature 2005, 436, 1171–1175. [Google Scholar]
- Rogers, S.A.; Huigens, R.W., 3rd; Cavanagh, J.; Melander, C. Synergistic effects between conventional antibiotics and 2-aminoimidazole-derived antibiofilm agents. Antimicrob. Agents Chemother 2010, 54, 2112–2118. [Google Scholar] [CrossRef]
- Bernier, S.P.; Lebeaux, D.; Defrancesco, A.S.; Valomon, A.; Soubigou, G.; Coppee, J.Y.; Ghigo, J.M.; Beloin, C. Starvation, together with the SOS response, mediates high biofilm-specific tolerance to the fluoroquinolone ofloxacin. PLoS Genet. 2013, 9, e1003144. [Google Scholar] [CrossRef]
- Mueller, R.S.; Dill, B.D.; Pan, C.; Belnap, C.P.; Thomas, B.C.; VerBerkmoes, N.C.; Hettich, R.L.; Banfield, J.F. Proteome changes in the initial bacterial colonist during ecological succession in an acid mine drainage biofilm community. Environ. Microbiol. 2011, 13, 2279–2292. [Google Scholar] [CrossRef]
- Gotoh, H.; Kasaraneni, N.; Devineni, N.; Dallo, S.F.; Weitao, T. SOS involvement in stress-inducible biofilm formation. Biofouling 2010, 26, 603–611. [Google Scholar] [CrossRef]
- Molin, S.; Tolker-Nielsen, T. Gene transfer occurs with enhanced efficiency in biofilms and induces enhanced stabilisation of the biofilm structure. Curr. Opin. Biotechnol. 2003, 14, 255–261. [Google Scholar] [CrossRef]
- Hennequin, C.; Aumeran, C.; Robin, F.; Traore, O.; Forestier, C. Antibiotic resistance and plasmid transfer capacity in biofilm formed with a CTX-M-15-producing Klebsiella pneumoniae isolate. J. Antimicrob. Chemother. 2012, 67, 2123–2130. [Google Scholar] [CrossRef]
- Ysern, P.; Clerch, B.; Castano, M.; Gibert, I.; Barbe, J.; Llagostera, M. Induction of SOS genes in Escherichia coli and mutagenesis in Salmonella typhimurium by fluoroquinolones. Mutagenesis 1990, 5, 63–66. [Google Scholar] [CrossRef]
- Miller, C.; Thomsen, L.E.; Gaggero, C.; Mosseri, R.; Ingmer, H.; Cohen, S.N. SOS response induction by beta-lactams and bacterial defense against antibiotic lethality. Science 2004, 305, 1629–1631. [Google Scholar] [CrossRef]
- Perez-Capilla, T.; Baquero, M.R.; Gomez-Gomez, J.M.; Ionel, A.; Martin, S.; Blazquez, J. SOS-independent induction of dinB transcription by beta-lactam-mediated inhibition of cell wall synthesis in Escherichia coli. J. Bacteriol. 2005, 187, 1515–1518. [Google Scholar] [CrossRef]
- Shaw, K.J.; Miller, N.; Liu, X.; Lerner, D.; Wan, J.; Bittner, A.; Morrow, B.J. Comparison of the changes in global gene expression of Escherichia coli induced by four bactericidal agents. J. Mol. Microbiol. Biotechnol. 2003, 5, 105–122. [Google Scholar] [CrossRef]
- Mesak, L.R.; Davies, J. Phenotypic changes in ciprofloxacin-resistant Staphylococcus aureus. Res. Microbiol. 2009, 160, 785–791. [Google Scholar] [CrossRef]
- Mesak, L.R.; Miao, V.; Davies, J. Effects of subinhibitory concentrations of antibiotics on SOS and DNA repair gene expression in Staphylococcus aureus. Antimicrob. Agents Chemother. 2008, 52, 3394–3397. [Google Scholar] [CrossRef]
- Lopez, E.; Blazquez, J. Effect of subinhibitory concentrations of antibiotics on intrachromosomal homologous recombination in Escherichia coli. Antimicrob. Agents Chemother. 2009, 53, 3411–3415. [Google Scholar] [CrossRef]
- Cirz, R.T.; Romesberg, F.E. Induction and inhibition of ciprofloxacin resistance-conferring mutations in hypermutator bacteria. Antimicrob. Agents Chemother. 2006, 50, 220–225. [Google Scholar] [CrossRef]
- Didier, J.P.; Villet, R.; Huggler, E.; Lew, D.P.; Hooper, D.C.; Kelley, W.L.; Vaudaux, P. Impact of ciprofloxacin exposure on Staphylococcus aureus genomic alterations linked with emergence of rifampin resistance. Antimicrob. Agents Chemother. 2011, 55, 1946–1952. [Google Scholar] [CrossRef]
- Cohen, S.E.; Walker, G.C. The transcription elongation factor NusA is required for stress-induced mutagenesis in Escherichia coli. Curr. Biol. 2010, 20, 80–85. [Google Scholar] [CrossRef] [Green Version]
- Baharoglu, Z.; Krin, E.; Mazel, D. RpoS plays a central role in the SOS induction by sub-lethal aminoglycoside concentrations in Vibrio cholerae. PLoS Genet. 2013, 9, e1003421. [Google Scholar] [CrossRef]
- Hocquet, D.; Llanes, C.; Thouverez, M.; Kulasekara, H.D.; Bertrand, X.; Plesiat, P.; Mazel, D.; Miller, S.I. Evidence for induction of integron-based antibiotic resistance by the SOS response in a clinical setting. PLoS Pathog. 2012, 8, e1002778. [Google Scholar] [CrossRef] [Green Version]
- Erill, I.; Campoy, S.; Barbe, J. Aeons of distress: An evolutionary perspective on the bacterial SOS response. FEMS Microbiol. Rev. 2007, 31, 637–656. [Google Scholar] [CrossRef]
- Gutierrez, A.; Laureti, L.; Crussard, S.; Abida, H.; Rodríguez Rojas, A.; Blázquez, J.; Baharoglu, Z.; Mazel, D.; Darfeuille, F.; Vogel, J.; et al. β-lactam antibiotics promote mutagenesis via RpoS-mediated replication fidelity reduction. Nat. Commun. 2013, 4, e1610. [Google Scholar] [CrossRef]
- Denamur, E.; Tenaillon, O.; Deschamps, C.; Skurnik, D.; Ronco, E.; Gaillard, J.L.; Picard, B.; Branger, C.; Matic, I. Intermediate mutation frequencies favor evolution of multidrug resistance in Escherichia coli. Genetics 2005, 171, 825–827. [Google Scholar] [CrossRef]
- Da Re, S.; Garnier, F.; Guerin, E.; Campoy, S.; Denis, F.; Ploy, M.C. The SOS response promotes qnrB quinolone-resistance determinant expression. EMBO Rep. 2009, 10, 929–933. [Google Scholar] [CrossRef]
- Ubeda, C.; Maiques, E.; Knecht, E.; Lasa, I.; Novick, R.P.; Penades, J.R. Antibiotic-induced SOS response promotes horizontal dissemination of pathogenicity island-encoded virulence factors in Staphylococci. Mol. Microbiol. 2005, 56, 836–844. [Google Scholar] [CrossRef]
- Maiques, E.; Ubeda, C.; Campoy, S.; Salvador, N.; Lasa, I.; Novick, R.P.; Barbe, J.; Penades, J.R. Beta-lactam antibiotics induce the SOS response and horizontal transfer of virulence factors in Staphylococcus aureus. J. Bacteriol. 2006, 188, 2726–2729. [Google Scholar] [CrossRef]
- Prudhomme, M.; Attaiech, L.; Sanchez, G.; Martin, B.; Claverys, J.P. Antibiotic stress induces genetic transformability in the human pathogen Streptococcus pneumoniae. Science 2006, 313, 89–92. [Google Scholar] [CrossRef]
- Gullberg, E.; Cao, S.; Berg, O.G.; Ilback, C.; Sandegren, L.; Hughes, D.; Andersson, D.I. Selection of resistant bacteria at very low antibiotic concentrations. PLoS Pathog. 2011, 7, e1002158. [Google Scholar] [CrossRef]
- Gustafsson, I.; Sjolund, M.; Torell, E.; Johannesson, M.; Engstrand, L.; Cars, O.; Andersson, D.I. Bacteria with increased mutation frequency and antibiotic resistance are enriched in the commensal flora of patients with high antibiotic usage. J. Antimicrob. Chemother. 2003, 52, 645–650. [Google Scholar] [CrossRef]
- Liu, A.; Fong, A.; Becket, E.; Yuan, J.; Tamae, C.; Medrano, L.; Maiz, M.; Wahba, C.; Lee, C.; Lee, K.; et al. Selective advantage of resistant strains at trace levels of antibiotics: A simple and ultrasensitive color test for detection of antibiotics and genotoxic agents. Antimicrob. Agents Chemother. 2011, 55, 1204–1210. [Google Scholar] [CrossRef]
- Dorr, T.; Lewis, K.; Vulic, M. SOS response induces persistence to fluoroquinolones in Escherichia coli. PLoS Genet. 2009, 5, e1000760. [Google Scholar] [CrossRef]
- Dorr, T.; Vulic, M.; Lewis, K. Ciprofloxacin causes persister formation by inducing the TisB toxin in Escherichia coli. PLoS Biol. 2010, 8, e1000317. [Google Scholar] [CrossRef]
- Lewis, K. Persister cells. Annu. Rev. Microbiol. 2010, 64, 357–372. [Google Scholar] [CrossRef]
- Guerout, A.M.; Iqbal, N.; Mine, N.; Ducos-Galand, M.; van Melderen, L.; Mazel, D. Characterization of the phd-doc and ccd toxin-antitoxin cassettes from Vibrio superintegrons. J. Bacteriol. 2013, 195, 2270–2283. [Google Scholar] [CrossRef]
- Kohanski, M.A.; DePristo, M.A.; Collins, J.J. Sublethal antibiotic treatment leads to multidrug resistance via radical-induced mutagenesis. Mol. Cell 2010, 37, 311–320. [Google Scholar]
- Mcbride, T.J.; Preston, B.D.; Loeb, L.A. Mutagenic spectrum resulting from DNA damage by oxygen radicals. Biochemistry 1991, 30, 207–213. [Google Scholar] [CrossRef]
- Nunoshiba, T.; Obata, F.; Boss, A.C.; Oikawa, S.; Mori, T.; Kawanishi, S.; Yamamoto, E. Role of iron and superoxide for generation of hydroxyl radical, oxidative DNA lesions, and mutagenesis in Escherichia coli. J. Biol. Chem. 1999, 274, 34832–34837. [Google Scholar]
- Fraud, S.; Poole, K. Oxidative stress induction of the mexXY multidrug efflux genes and promotion of aminoglycoside resistance development in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2011, 55, 1068–1074. [Google Scholar] [CrossRef]
- Aiassa, V.; Barnes, A.I.; Smania, A.M.; Albesa, I. Sublethal ciprofloxacin treatment leads to resistance via antioxidant systems in Proteus mirabilis. FEMS Microbiol. Lett. 2012, 327, 25–32. [Google Scholar] [CrossRef]
- Dwyer, D.J.; Kohanski, M.A.; Collins, J.J. Role of reactive oxygen species in antibiotic action and resistance. Curr. Opin. Microbiol. 2009, 12, 482–489. [Google Scholar] [CrossRef]
- Dwyer, D.J.; Kohanski, M.A.; Hayete, B.; Collins, J.J. Gyrase inhibitors induce an oxidative damage cellular death pathway in Escherichia coli. Mol. Syst. Biol. 2007, 3, e91. [Google Scholar]
- Wholey, W.Y.; Jakob, U. Hsp33 confers bleach resistance by protecting elongation factor Tu against oxidative degradation in Vibrio cholerae. Mol. Microbiol. 2012, 83, 981–991. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, D.; Joshi-Datar, A.; Lepine, F.; Bauerle, E.; Olakanmi, O.; Beer, K.; McKay, G.; Siehnel, R.; Schafhauser, J.; Wang, Y.; et al. Active starvation responses mediate antibiotic tolerance in biofilms and nutrient-limited bacteria. Science 2011, 334, 982–986. [Google Scholar] [CrossRef]
- Shatalin, K.; Shatalina, E.; Mironov, A.; Nudler, E. H2S: A universal defense against antibiotics in bacteria. Science 2011, 334, 986–990. [Google Scholar] [CrossRef]
- Belenky, P.; Collins, J.J. Microbiology. Antioxidant strategies to tolerate antibiotics. Science 2011, 334, 915–916. [Google Scholar]
- Allen, K.J.; Griffiths, M.W. Impact of hydroxyl- and superoxide anion-based oxidative stress on logarithmic and stationary phase Escherichia coli O157:H7 stress and virulence gene expression. Food Microbiol. 2012, 29, 141–147. [Google Scholar] [CrossRef]
- Merrikh, H.; Ferrazzoli, A.E.; Bougdour, A.; Olivier-Mason, A.; Lovett, S.T. A DNA damage response in Escherichia coli involving the alternative sigma factor, RpoS. Proc. Natl. Acad. Sci. USA 2009, 106, 611–616. [Google Scholar]
- Merrikh, H.; Ferrazzoli, A.E.; Lovett, S.T. Growth phase and (p)ppGpp control of IraD, a regulator of RpoS stability, in Escherichia coli. J. Bacteriol. 2009, 191, 7436–7446. [Google Scholar] [CrossRef]
- Battesti, A.; Tsegaye, Y.M.; Packer, D.G.; Majdalani, N.; Gottesman, S. H-NS regulation of IraD and IraM anti-adaptors for control of RpoS degradation. J. Bacteriol. 2012, 194, 2470–2478. [Google Scholar] [CrossRef]
- Hengge-Aronis, R. Signal transduction and regulatory mechanisms involved in control of the sigma(S) (RpoS) subunit of RNA polymerase. Microbiol. Mol. Biol. Rev. 2002, 66, 373–395. [Google Scholar] [CrossRef]
- Barth, E.; Gora, K.V.; Gebendorfer, K.M.; Settele, F.; Jakob, U.; Winter, J. Interplay of cellular cAMP levels, {sigma}S activity and oxidative stress resistance in Escherichia coli. Microbiology 2009, 155, 1680–1689. [Google Scholar] [CrossRef]
- Daly, M.J. A new perspective on radiation resistance based on Deinococcus radiodurans. Nat. Rev. Microbiol. 2009, 7, 237–245. [Google Scholar] [CrossRef]
- Henle, E.S.; Linn, S. Formation, prevention, and repair of DNA damage by iron/hydrogen peroxid. J. Biol. Chem. 1997, 272, 19095–19098. [Google Scholar] [CrossRef]
- Keyer, K.; Imlay, J.A. Superoxide accelerates DNA damage by elevating free-iron levels. Proc. Natl. Acad. Sci. USA 1996, 93, 13635–13640. [Google Scholar] [CrossRef]
- Hansen, S.; Lewis, K.; Vulic, M. Role of global regulators and nucleotide metabolism in antibiotic tolerance in Escherichia coli. Antimicrob. Agents Chemother. 2008, 52, 2718–2726. [Google Scholar] [CrossRef]
- Miyazaki, R.; Minoia, M.; Pradervand, N.; Sulser, S.; Reinhard, F.; van der Meer, J.R. Cellular variability of RpoS expression underlies subpopulation activation of an integrative and conjugative element. PLoS Genet. 2012, 8, e1002818. [Google Scholar] [CrossRef]
- Santos-Zavaleta, A.; Gama-Castro, S.; Perez-Rueda, E. A comparative genome analysis of the RpoS sigmulon shows a high diversity of responses and origins. Microbiology 2011, 157, 1393–1401. [Google Scholar] [CrossRef]
- Chiang, S.M.; Schellhorn, H.E. Evolution of the RpoS regulon: Origin of RpoS and the conservation of RpoS-dependent regulation in bacteria. J. Mol. Evol. 2010, 70, 557–571. [Google Scholar] [CrossRef]
- King, T.; Seeto, S.; Ferenci, T. Genotype-by-environment interactions influencing the emergence of RpoS mutations in Escherichia coli populations. Genetics 2006, 172, 2071–2079. [Google Scholar] [CrossRef]
- Zambrano, M.M.; Siegele, D.A.; Almiron, M.; Tormo, A.; Kolter, R. Microbial competition: Escherichia coli mutants that take over stationary phase cultures. Science 1993, 259, 1757–1760. [Google Scholar]
- Zhang, Y.; Shi, C.; Yu, J.; Ren, J.; Sun, D. RpoS regulates a novel type of plasmid DNA transfer in Escherichia coli. PLoS One 2012, 7, e33514. [Google Scholar] [CrossRef]
- Joelsson, A.; Kan, B.; Zhu, J. Quorum sensing enhances the stress response in Vibrio cholerae. Appl. Environ. Microbiol. 2007, 73, 3742–3746. [Google Scholar] [CrossRef]
- Allison, K.R.; Brynildsen, M.P.; Collins, J.J. Metabolite-enabled eradication of bacterial persisters by aminoglycosides. Nature 2011, 473, 216–220. [Google Scholar] [CrossRef]
- Farha, M.A.; Brown, E.D. Discovery of antibiotic adjuvants. Nat. Biotechnol. 2013, 31, 120–122. [Google Scholar] [CrossRef]
- Brynildsen, M.P.; Winkler, J.A.; Spina, C.S.; Macdonald, I.C.; Collins, J.J. Potentiating antibacterial activity by predictably enhancing endogenous microbial ROS production. Nat. Biotechnol. 2013, 31, 160–165. [Google Scholar] [CrossRef]
- Lu, T.K.; Collins, J.J. Engineered bacteriophage targeting gene networks as adjuvants for antibiotic therapy. Proc. Natl. Acad. Sci. USA 2009, 106, 4629–4634. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Baharoglu, Z.; Garriss, G.; Mazel, D. Multiple Pathways of Genome Plasticity Leading to Development of Antibiotic Resistance. Antibiotics 2013, 2, 288-315. https://doi.org/10.3390/antibiotics2020288
Baharoglu Z, Garriss G, Mazel D. Multiple Pathways of Genome Plasticity Leading to Development of Antibiotic Resistance. Antibiotics. 2013; 2(2):288-315. https://doi.org/10.3390/antibiotics2020288
Chicago/Turabian StyleBaharoglu, Zeynep, Geneviève Garriss, and Didier Mazel. 2013. "Multiple Pathways of Genome Plasticity Leading to Development of Antibiotic Resistance" Antibiotics 2, no. 2: 288-315. https://doi.org/10.3390/antibiotics2020288