The analyses are based on model simulated anoxic volumes, mainly the reduction of anoxic volume from the no-wind scenario. The anoxic volumes are only compared at or before the peak reduction, while the post-peak recovery of anoxic volume is not covered in this paper.
3.1. Relative Anoxia Reduction by Wind Directions in Two-Day Wind Events
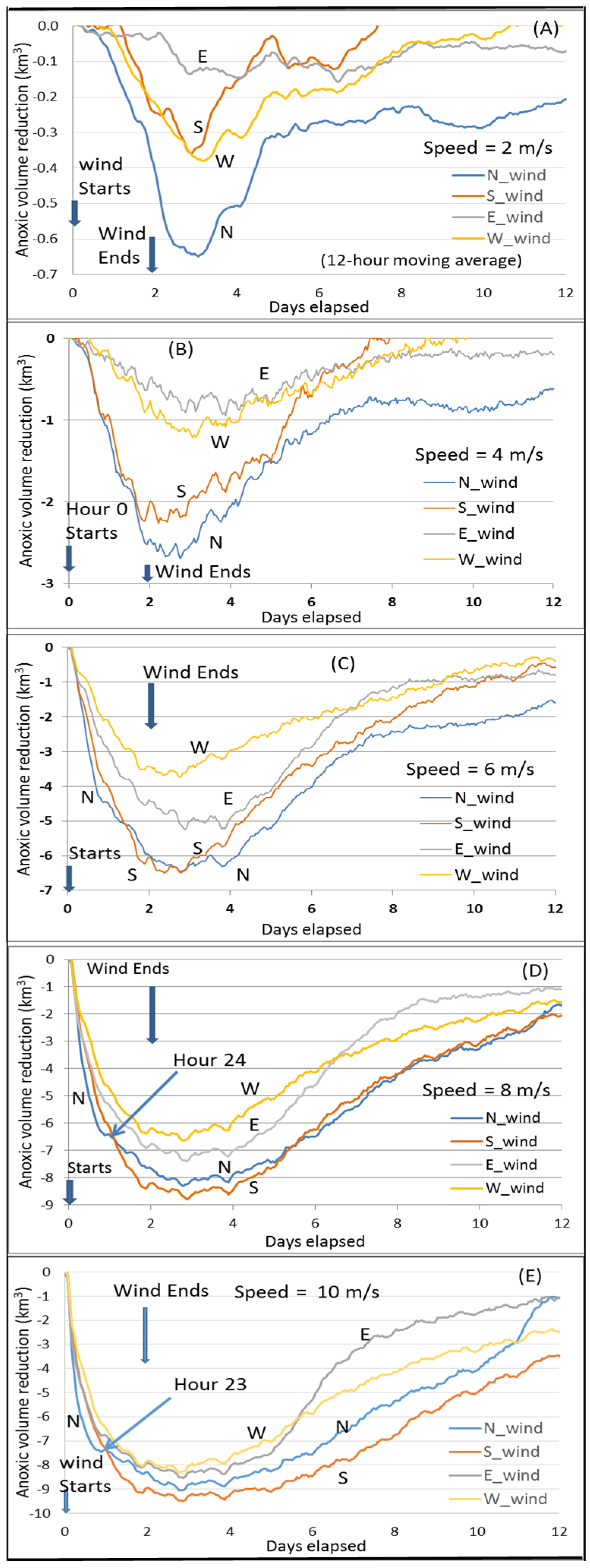
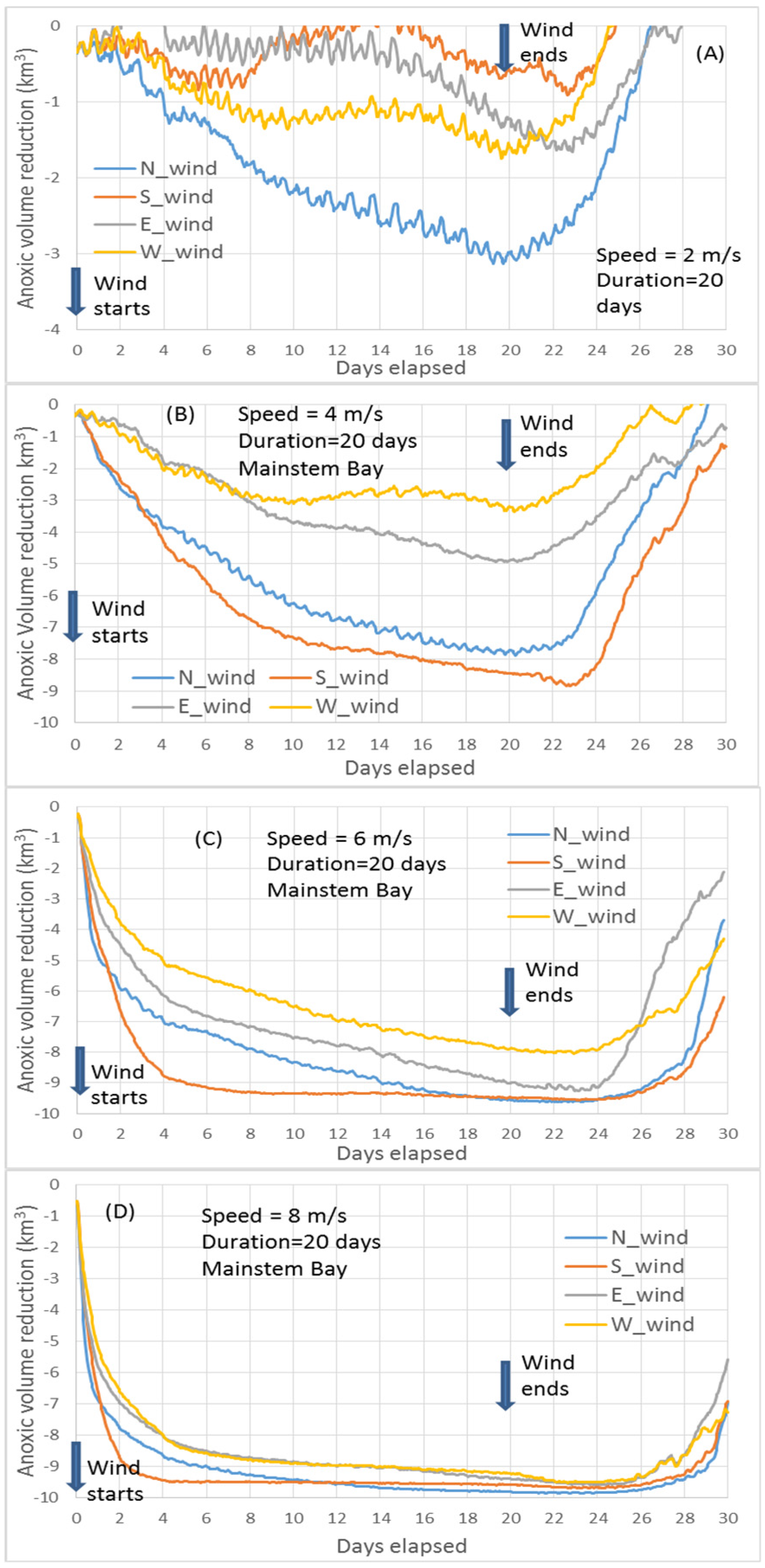
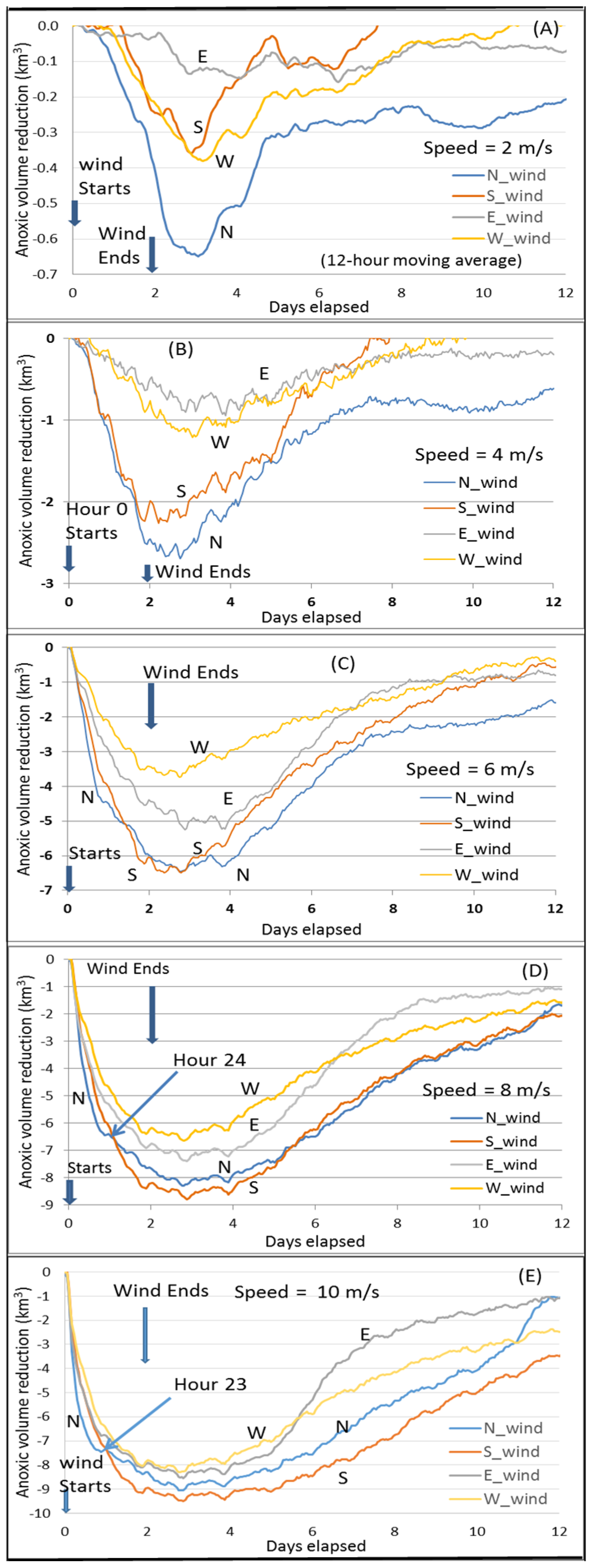
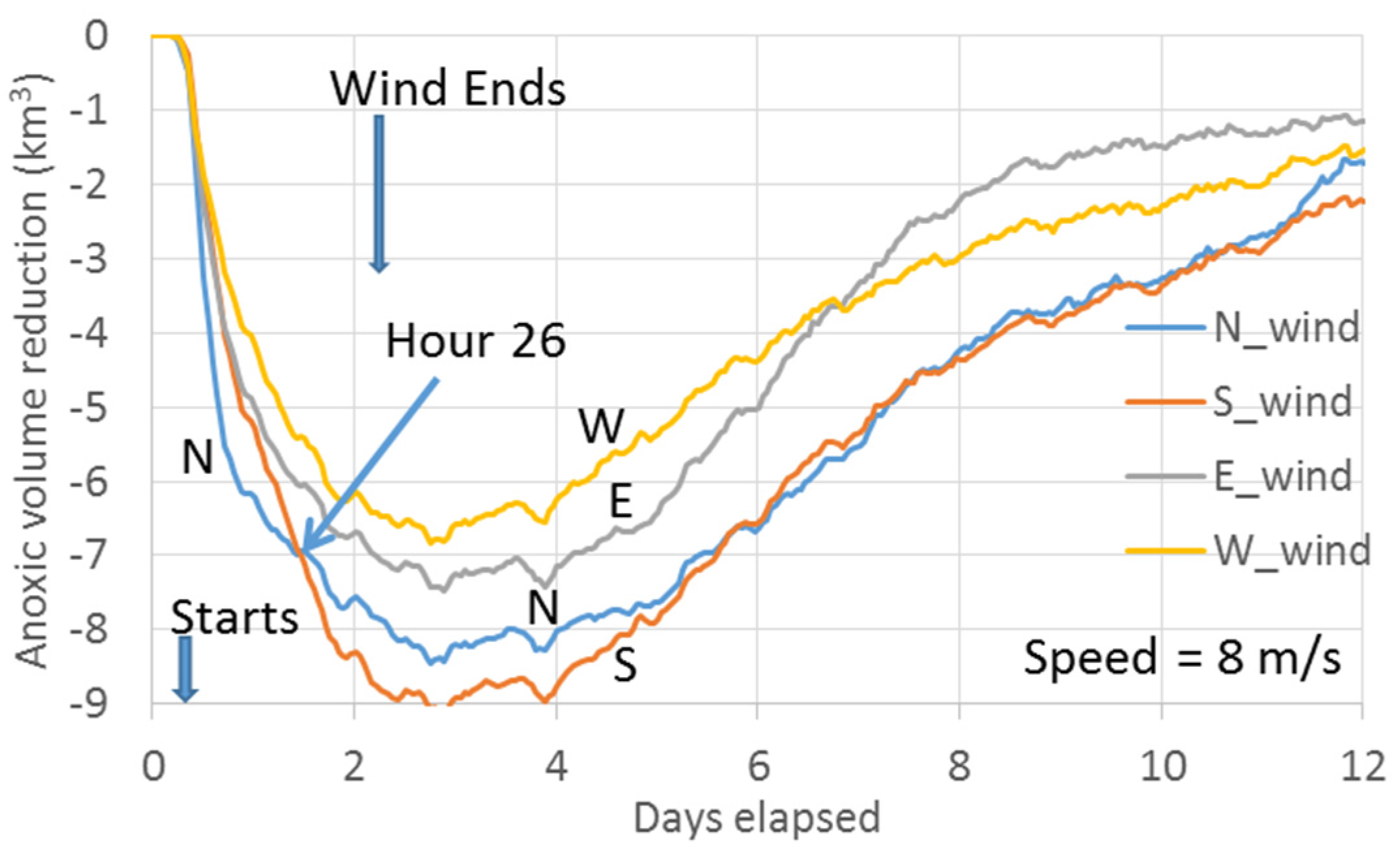
The core scenario A_8 (wind speeds of 8 m/s for two days,
Figure 4D) is the same as the key scenario used in Wang et al. [
10]. Here it is used as a reference to compare the results from other scenarios. Thus, it is useful to review the key findings by Wang et al. [
10]. The minimum point of the curves in
Figure 4D represents the maximum anoxic volume reduction. The dominant PN-type cross-channel bathymetry in the Bay’s anoxic center provides a favorable condition for the easterly wind to have a greater destratification than the westerly wind under the wind-induced cross-channel circulation [
10]. Thus, the easterly wind reduces more anoxia than the westerly wind. The direction of the southerly and northerly wind travel, respectively, against and along with the net transport direction of the surface fresher water, resulting in stronger destratification and greater reduction of anoxia by the southerly wind [
6,
7]. Notably, before Hour 24 of the wind event, northerly winds reduced anoxia more than southerly winds, also due to the effects of the PN-type bathymetry that modulates the wind-induced cross-channel circulation [
10]. The northerly wind has westward straining component as the easterly wind, and the southerly wind has eastward straining component as the westerly wind. Thus, under the modulation of the PN-type bathymetry on the wind-induced cross-channel circulation, the northerly wind promotes greater destratification. Cross-channel circulation under the simulated wind speeds could be completed in a couple of hours, while a timeline of 1–2 days is necessary for the wind-induced along-channel circulation to effectively influence the anoxic center [
21]. Before Hour 24, the northerly wind reduces more anoxic volume than the southerly wind, when the wind-induced cross-channel circulation plays a more important role. Following this period, however, the southerly wind-induced along-channel circulation (via downwelling from the Bay head) influences a wider area of the anoxic zone, leading to an overall greater anoxia reduction. The rest of this section will discuss anoxia reduction by winds at speeds different than the Core Scenario.
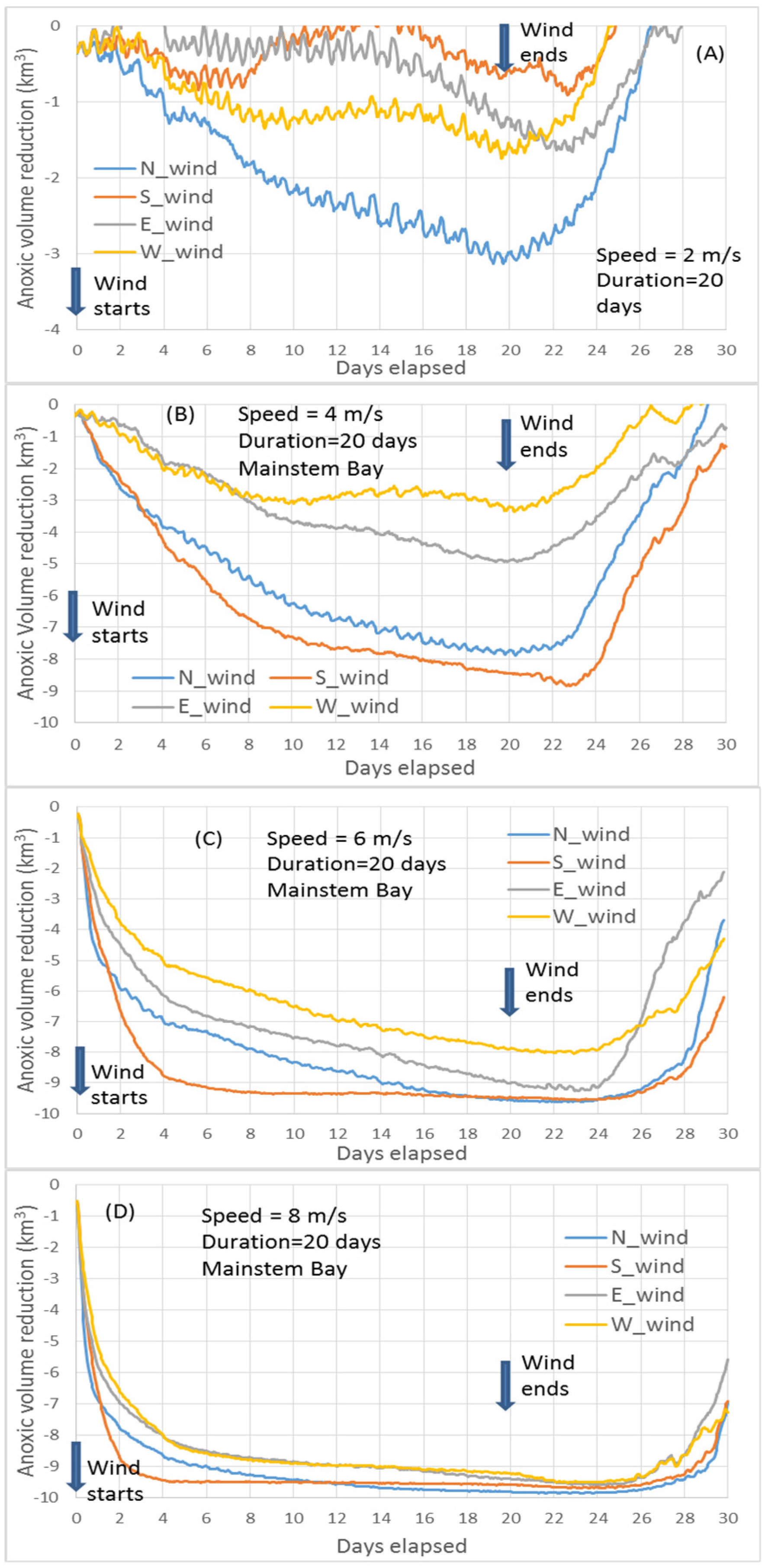
Northerly versus southerly wind. At a speed of 10 m/s, the transition point of greater anoxia reduction from northerly winds to southerly winds occurs one hour earlier (at Hour 23,
Figure 4E) than the Core Scenario (
Figure 4D), because the influence of the wind-induced along-channel circulation by the southerly wind is greater at higher speeds.
At speeds of 6 m/s, the maximum anoxic volume reduction by northerly winds becomes closer to that produced by southerly winds, and the transition to greater anoxia reduction by southerly winds is delayed to Hour 32 (
Figure 4C). This is due to a slower influence exercised upon the anoxic center by the wind-induced along-channel circulation at lower wind speeds.
At 2 or 4 m/s wind speeds, destratification is weak and the influence of wind-induced along-channel circulation by southerly winds is weak and slow. The aforementioned transition does not occur, and northerly winds reduce anoxia more than southerly winds across the entire time period (
Figure 4A,B). Again, this is primarily controlled by the wind-induced cross-channel circulation under the modulation of the PN-type cross-channel bathymetry.
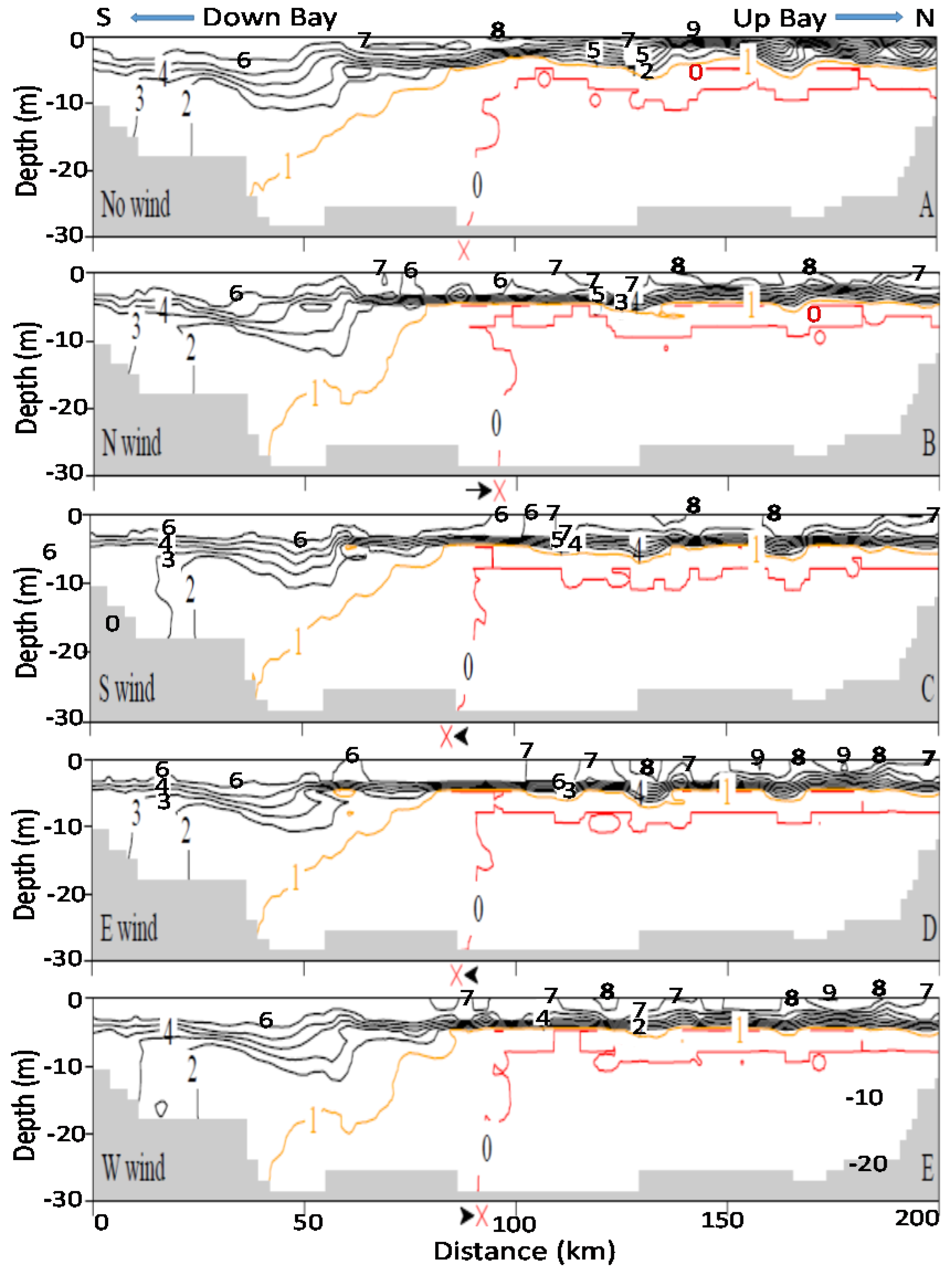
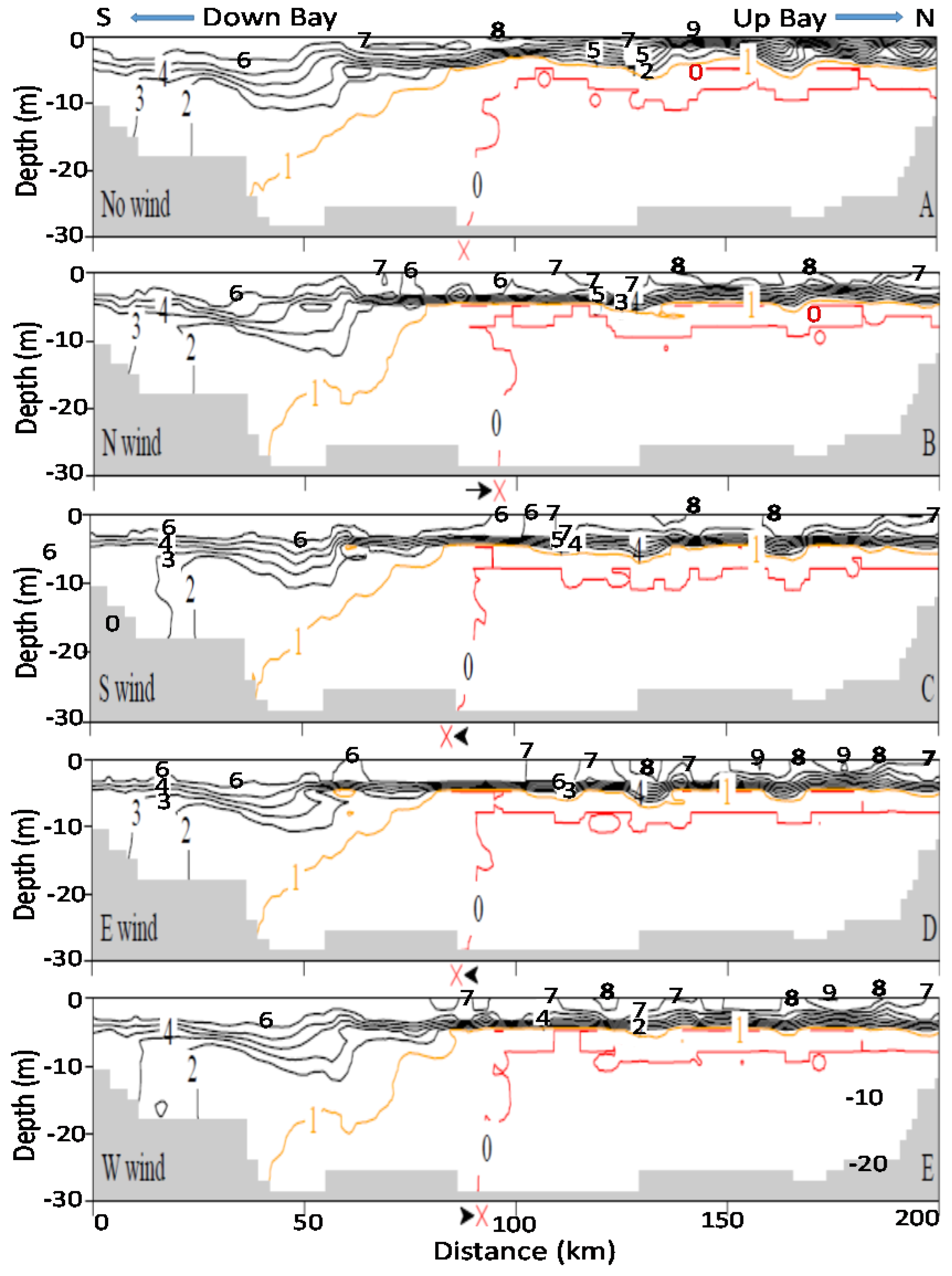
It should also be noted that the promoted up-Bay-ward bottom seawater intrusion promoted by northerly winds also plays a certain role, which can be seen in DO contours of along-channel sections in
Figure 5 and
Figure 6.
Figure 5 represents Hour 24 of the two-day wind scenario at 8 m/s, while
Figure 6 shows Hour 48 of a two-day wind scenario at 2 m/s. The symbol X by the x-axis marks the southern end of the 0 mg/L DO isopleths that intersect with the bottom bathymetry. Compared to the no-wind scenarios in
Figure 5A and
Figure 6A, the X retreats northwards for northerly winds (
Figure 5B and
Figure 6B) and extends further south in southerly winds (
Figure 5C and
Figure 6C). The role of the enhanced bottom seawater intrusion in anoxia reduction is difficult to quantitatively separate from other mechanisms. In 8 m/s wind velocities, the bulk anoxia reduction are mainly controlled by wind’s mixing and destratification, while the contribution of oxygenation by enhanced bottom seawater intrusion from northerly (or westerly) winds is relatively small (
Figure 5). In 2 m/s wind velocities, mixing or destratification is weak, the initial stratification is well maintained, and the DO isopleths in the four winds are similar to those in the no-wind condition (
Figure 6), exhibiting virtually no difference. The intrusion of oxygen-rich seawater via enhanced estuarine circulation by northerly winds caused the 0 mg/L DO isopleths to shrink and reduced overall anoxic volume. This can be further seen in
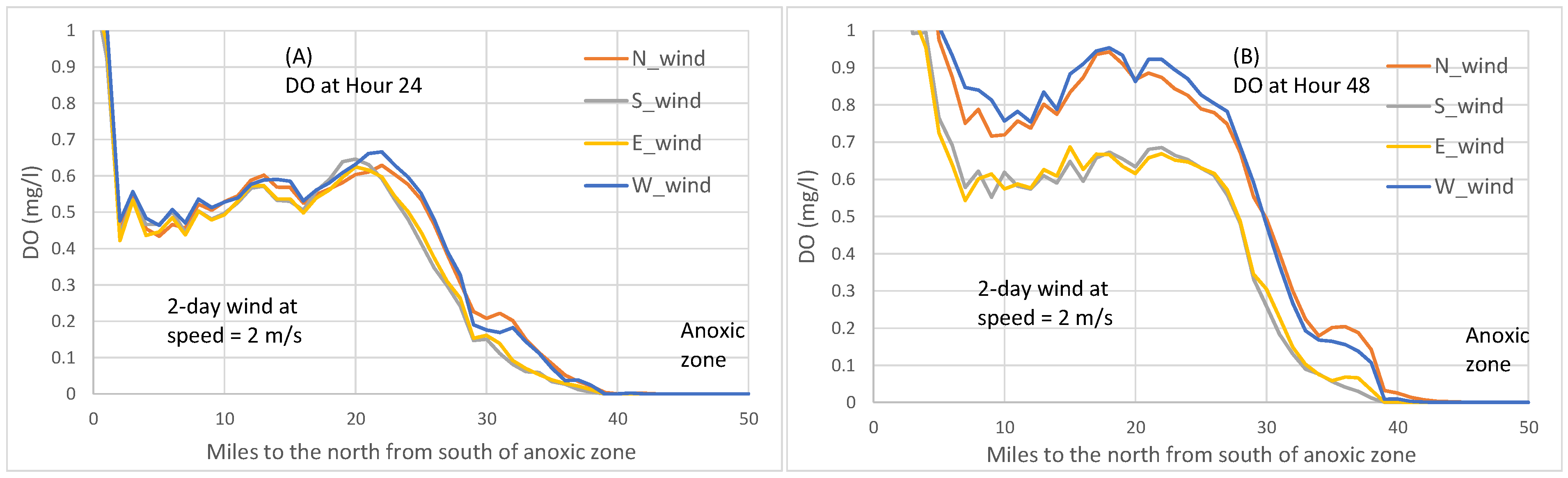
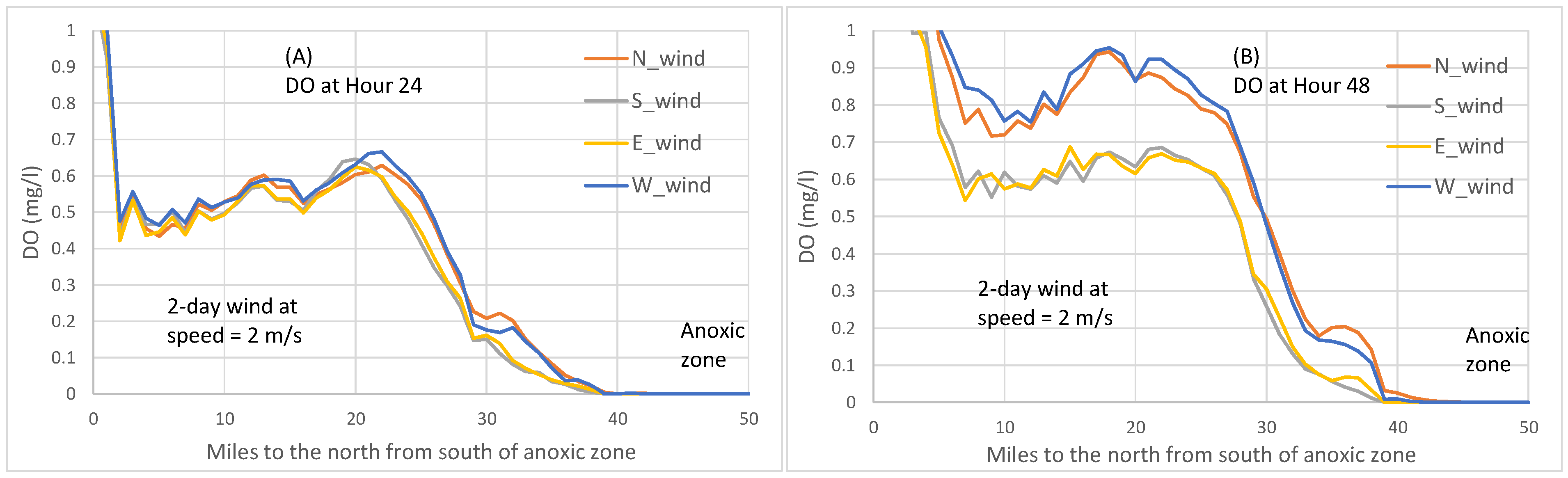
Figure 7 where bottom DO concentrations are plotted, northerly winds produce higher bottom DO than southerly winds, consistent with the anoxic volume reduction by northerly versus southerly winds shown in
Figure 4A.
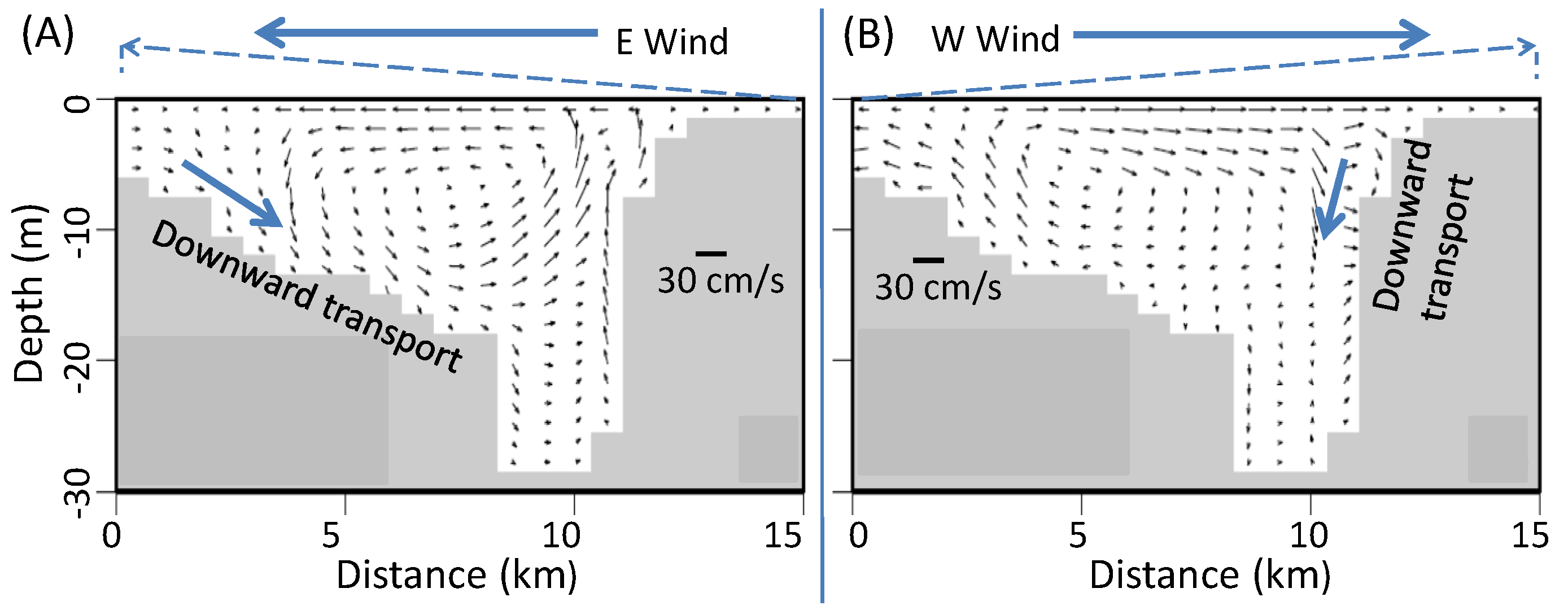
Easterly versus westerly winds. In easterly and westerly winds, the wind-induced cross-channel circulation is the dominant component that causes mixing and anoxia reduction. The PN-type cross-channel bathymetry provides favorable conditions for easterly winds over westerly winds to effect destratification by cross-channel circulation to a greater extent. Additionally, easterly winds have an up-Bay-ward component of along-channel straining, like southerly winds. Thus, at speeds of 6 m/s and greater in the two-day winds, easterly winds reduced more anoxia than westerly winds throughout the entire two-day wind event (
Figure 4C–E).
At wind speeds of 2 or 4 m/s, destratification induced by easterly and westerly winds was weak and the enhanced estuarine circulation due to westerly winds became relatively important in reducing anoxia, especially in the late wind period. Before Day 1, the hypoxia reduction was mainly controlled by the bathymetry-modulated wind-induced cross-channel circulation; therefore, easterly winds had a greater anoxia reduction than westerly winds. After about 24 h, westerly winds had a greater anoxia reduction than easterly winds, including the point of peak anoxia reduction (
Figure 4A,B), which is mainly due to the enhanced estuarine circulation by the westerly wind. The influence of bottom seawater intrusion on anoxia by westerly versus easterly winds can also be seen in
Figure 6 by the direction in which the symbol X moves. At wind speeds of 2 m/s, westerly winds caused the 0 mg/L DO isopleths to retreat northwards, reducing anoxia (
Figure 6E), and westerly winds produced higher bottom DO than easterly winds (
Figure 7). During most times after Hour 24 in the scenarios of wind speeds equal to 2 or 4 m/s, westerly winds reduced anoxia to a greater extent than easterly winds (
Figure 4A,B). Before Hour 24, easterly winds reduced more anoxic volume because this period was still primarily controlled by the bathymetry-modulated cross-channel circulation, while the enhanced estuarine circulation effected by westerly winds had not yet reached the anoxic center to a significant extent.
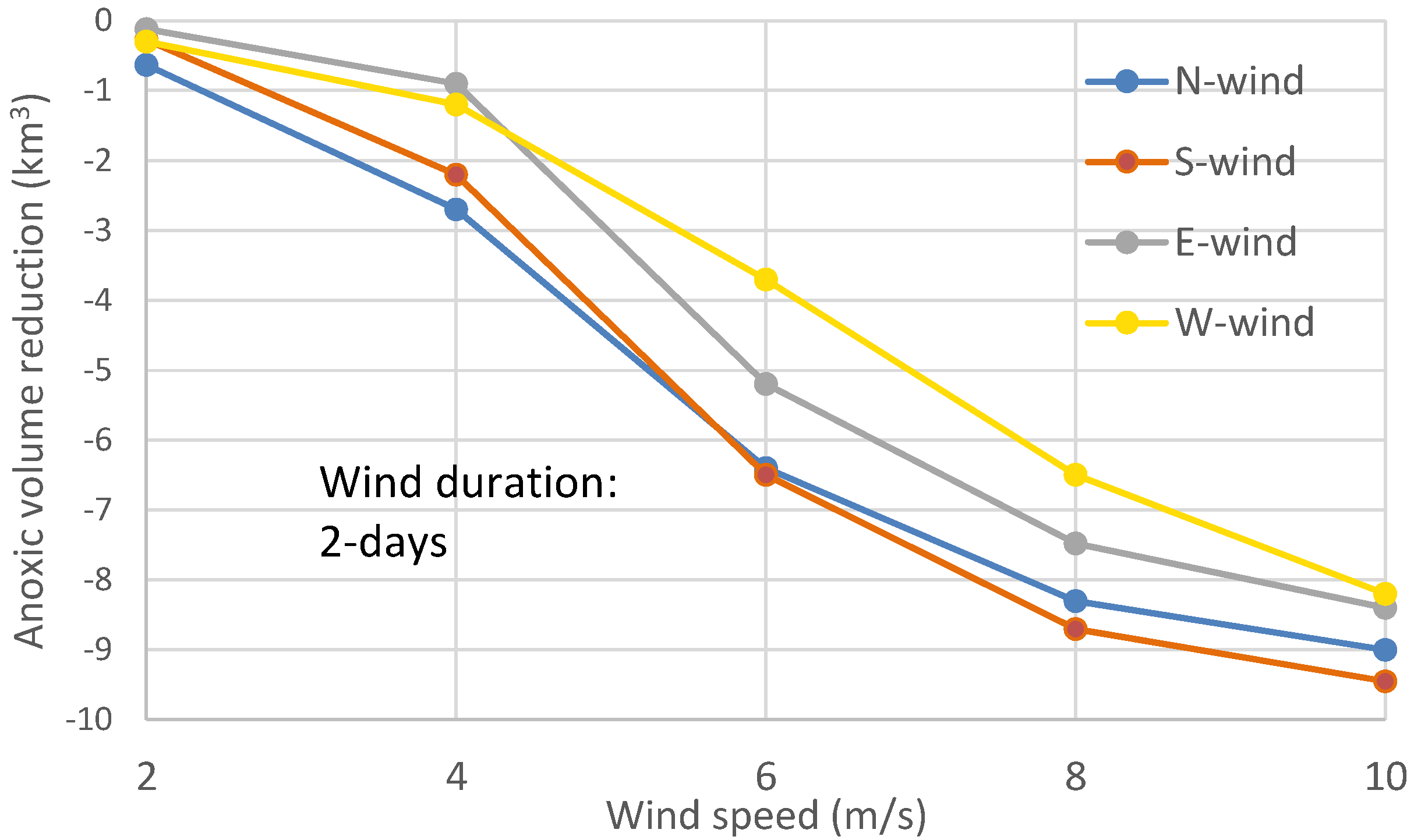
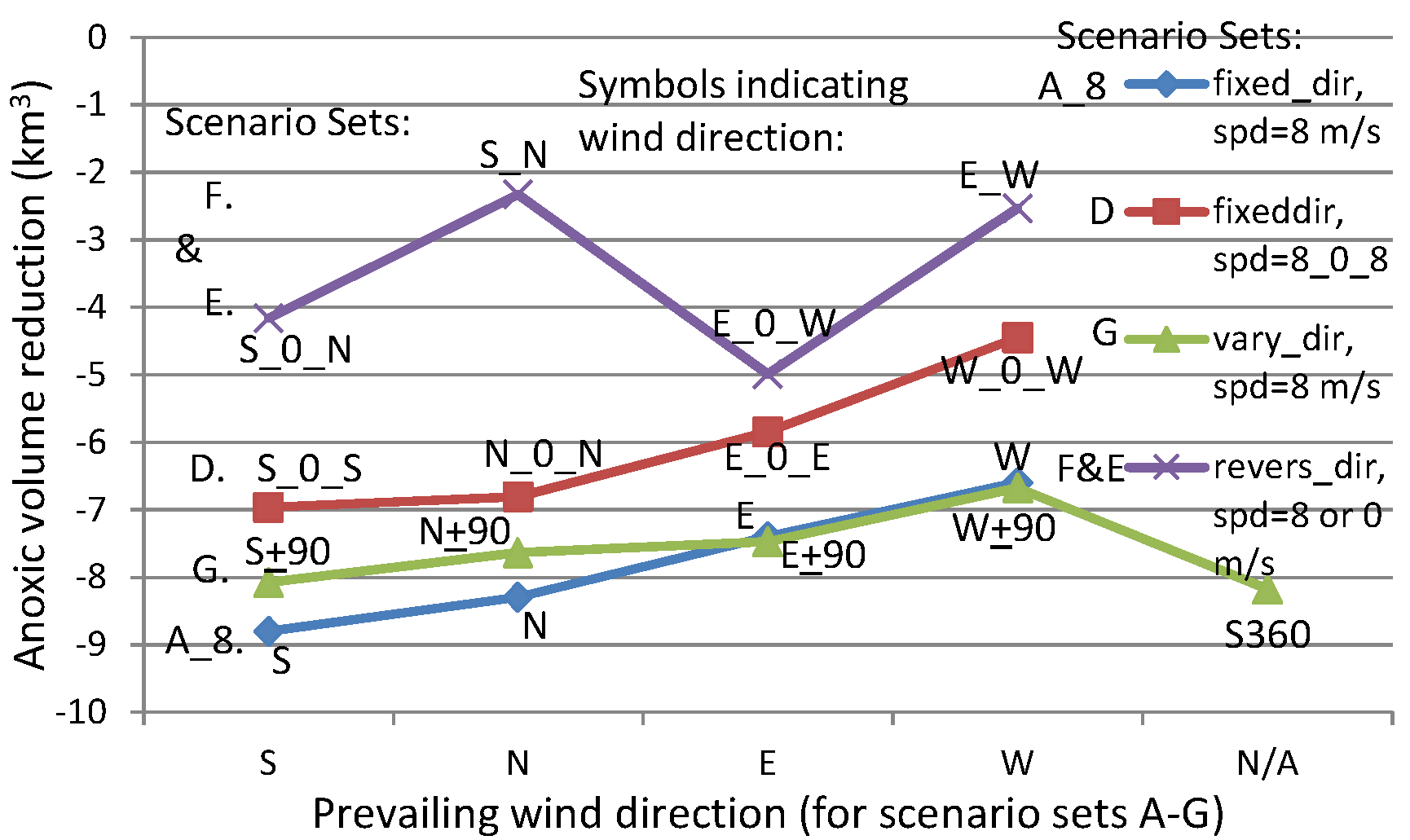
The above processes help to explain the summary figure (
Figure 8) of maximum anoxia reduction among wind directions at different speeds of the two-day winds, as seen in
Figure 4.
Assessing the influence of tide
. The stages of tide (ebb versus flood) at the moment when the wind event starts can also influence the responses of destratification to wind’s longitudinal straining [
22,
23].
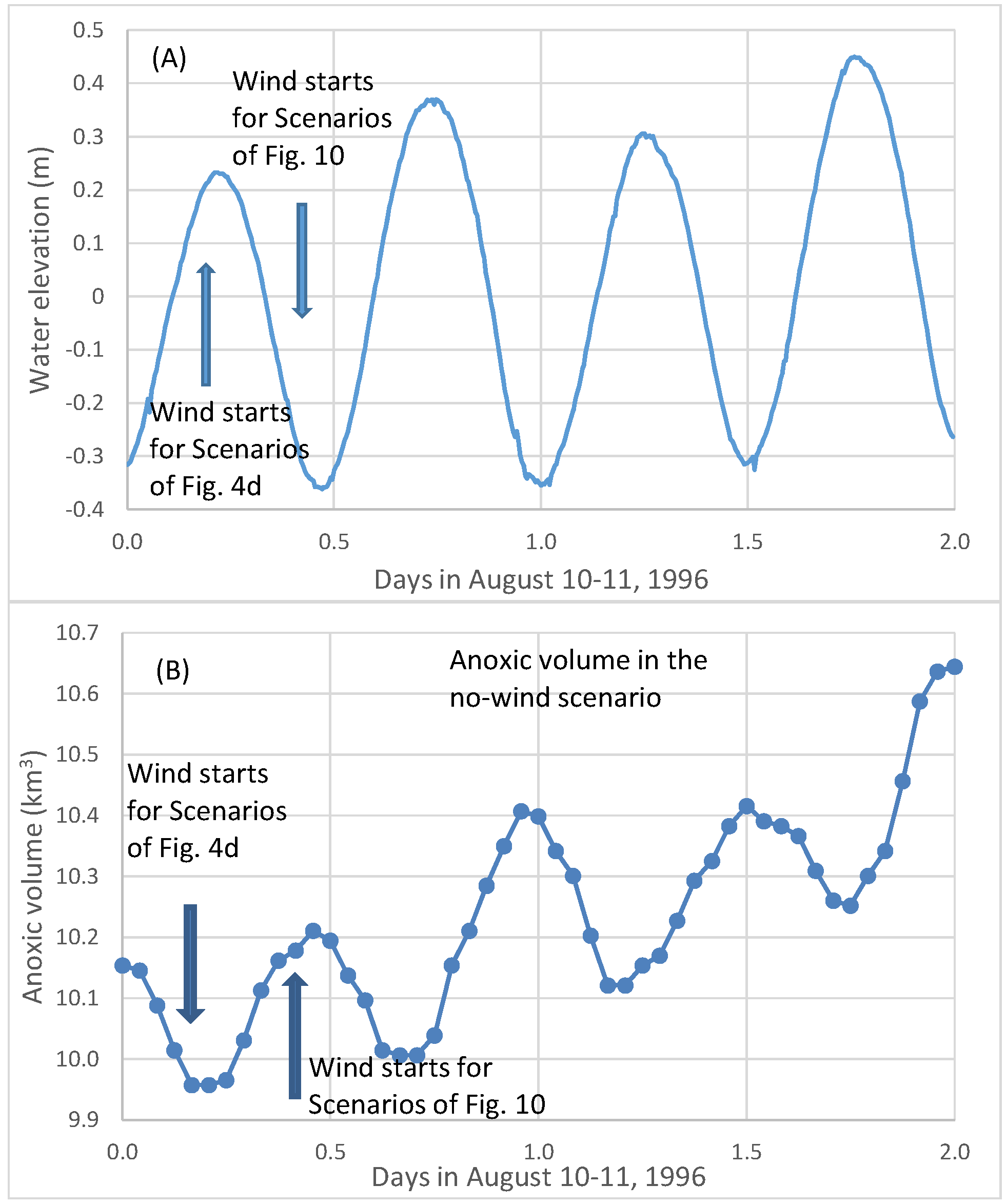
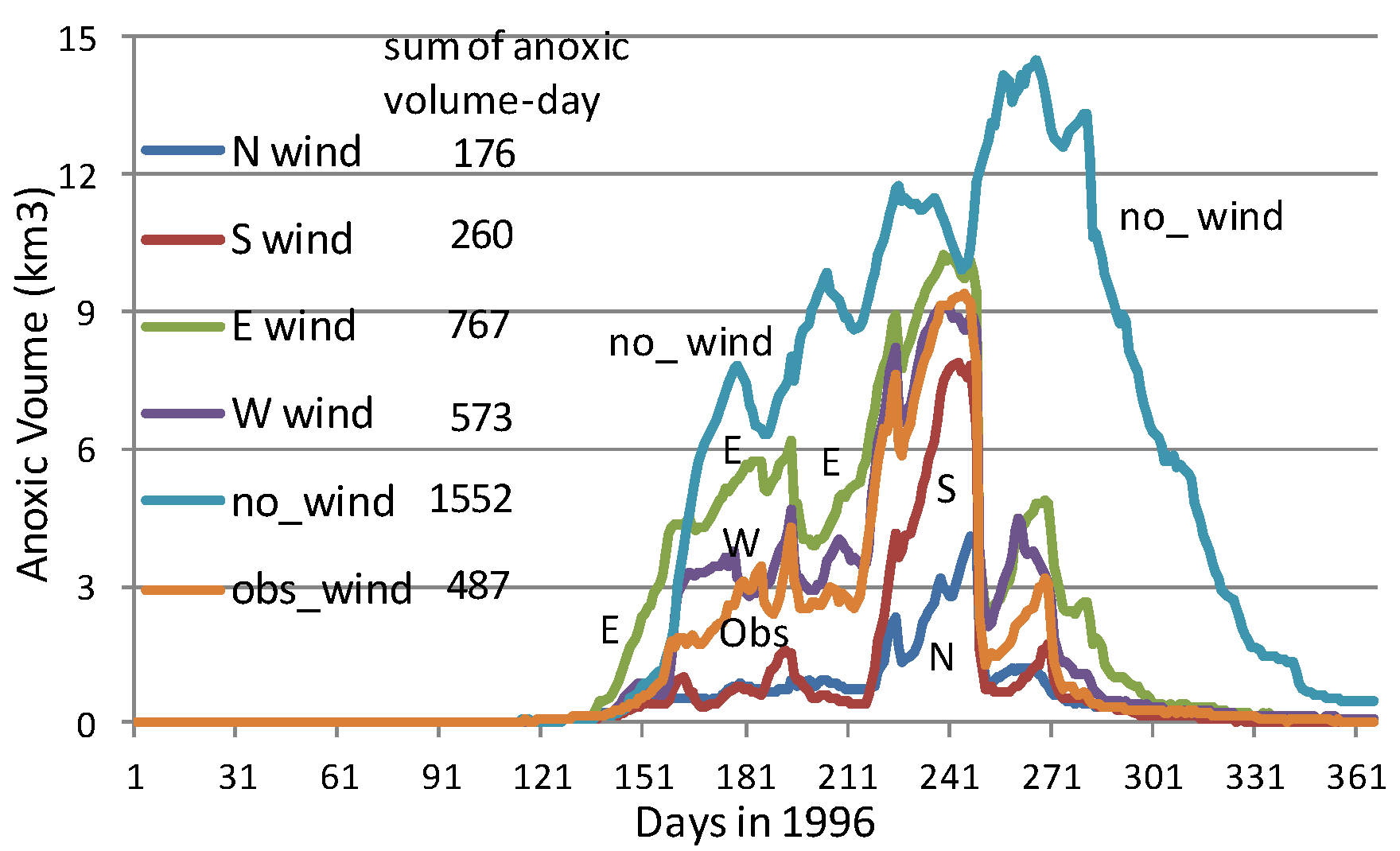
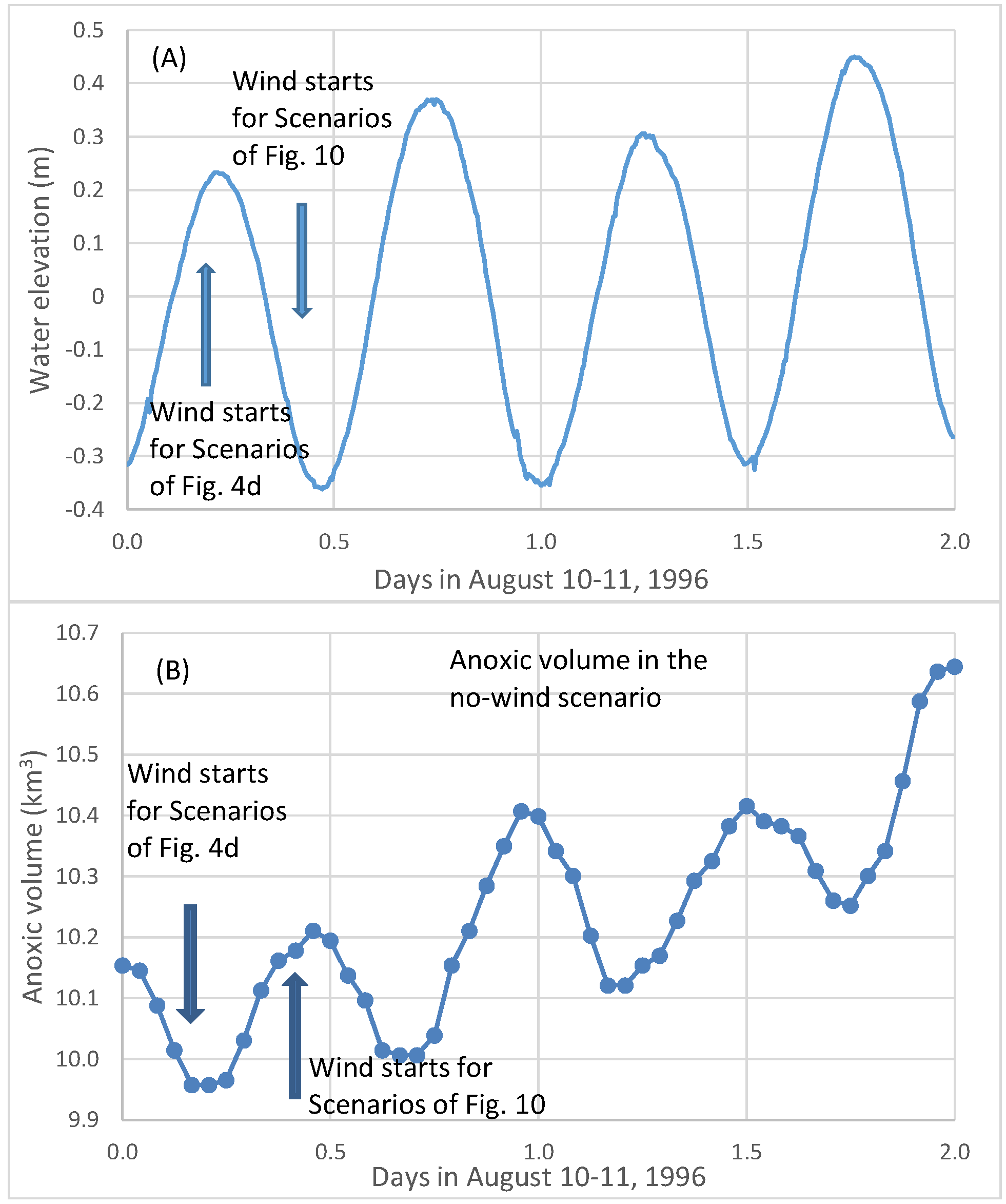
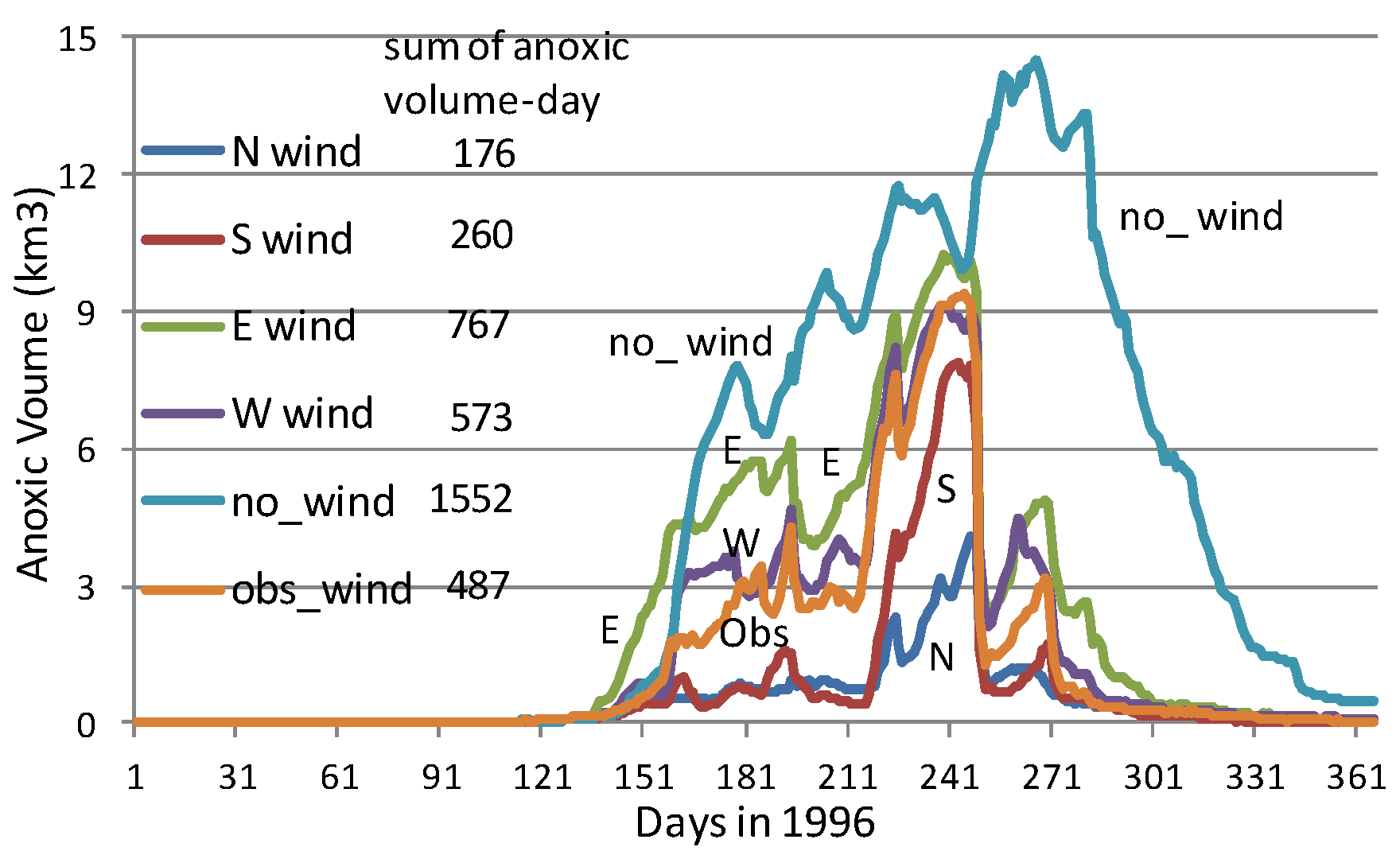
Figure 9b shows simulated hourly anoxic volume on 10 and 11 August 1996 from a no-wind scenario. The average daily anoxic volume increased from Day 1 to Day 2 due to strong oxygen consumption in early August. The two peaks and two valleys of anoxic volume in one day (
Figure 9b) were associated with the M2 tide (
Figure 9a). The peaks of the anoxic volume (lower DO) were associated with the stage of low-water after ebb tide, and the valleys of the anoxic volume (higher DO) were associated with the stage of high-water after flood tide. The flood tide brought oxygen-rich seawater to the anoxic zone and reduced anoxic volume. The influence of tides on the fluctuation of anoxic volume reached approximately 0.3 km
3 in this simulated high anoxic period.
The first arrow in
Figure 9A,B indicates the wind starting time in scenario set A_8 (
Figure 4d). It started at 4:00 a.m. on 10 August 1996, near high-tide as ebbing began at the Bay mouth. The second arrow indicates the wind’s starting time for another scenario set of 8 m/s winds, but the wind event began 6 hours later, at 10:00 a.m., near low-tide when flooding began at the Bay mouth.
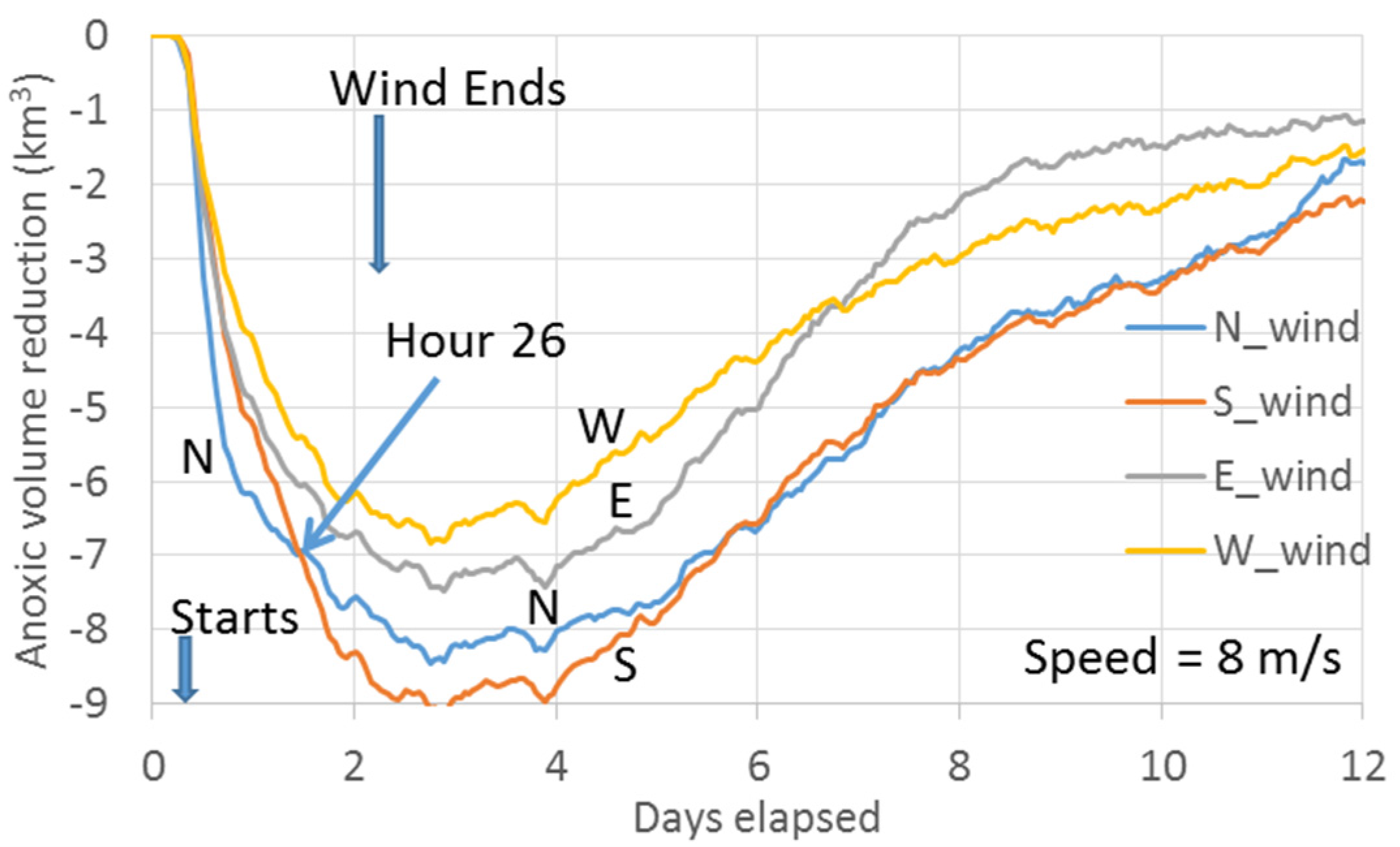
Figure 10 plots the simulated anoxic volume reductions by the latter scenario set of 8 m/s winds.
For a better comparison of
Figure 10 and
Figure 4d in temporal development, the origin of the x-axis in both figures is set to 4:00 a.m. on 10 August 1996. The wind events for the scenarios in
Figure 10 began 6 hours later, delineated in the graph. There is no significant difference in relative anoxia reduction among wind directions between
Figure 10 and
Figure 4d. In
Figure 10, the northerly wind reduced slightly more anoxia, and the time transitioning from greater anoxia reductions between northerly winds and southerly winds was delayed 2 hours, to Hour 26 of the wind event. Northerly winds can have certain advantages in destratification/anoxia reduction in flood tide versus ebb tide. It is difficult to determine a reference location for tidal stages that relate to wind-induced destratification in the anoxic center, because the time difference of the co-tidal lines is approximately 6–8 hours between the Bay mouth and the anoxic center in the mid-Bay and approximately 12 hours between the Bay mouth and the Bay head [
24]. The Bay mouth is used as reference location for tidal stages where the tidal changes are forced. Further analysis of this point lies beyond the scope of this work. The model experiments presented in
Figure 10 and
Figure 4d indicate that greater anoxia reduction by northerly winds than southerly winds before Hour 24 is not due to tidal stages during the wind events, and both experiments confirmed the argument of Wang et al. [
10] regarding the modulation of wind-induced cross-channel circulation by the PN-type bathymetry.
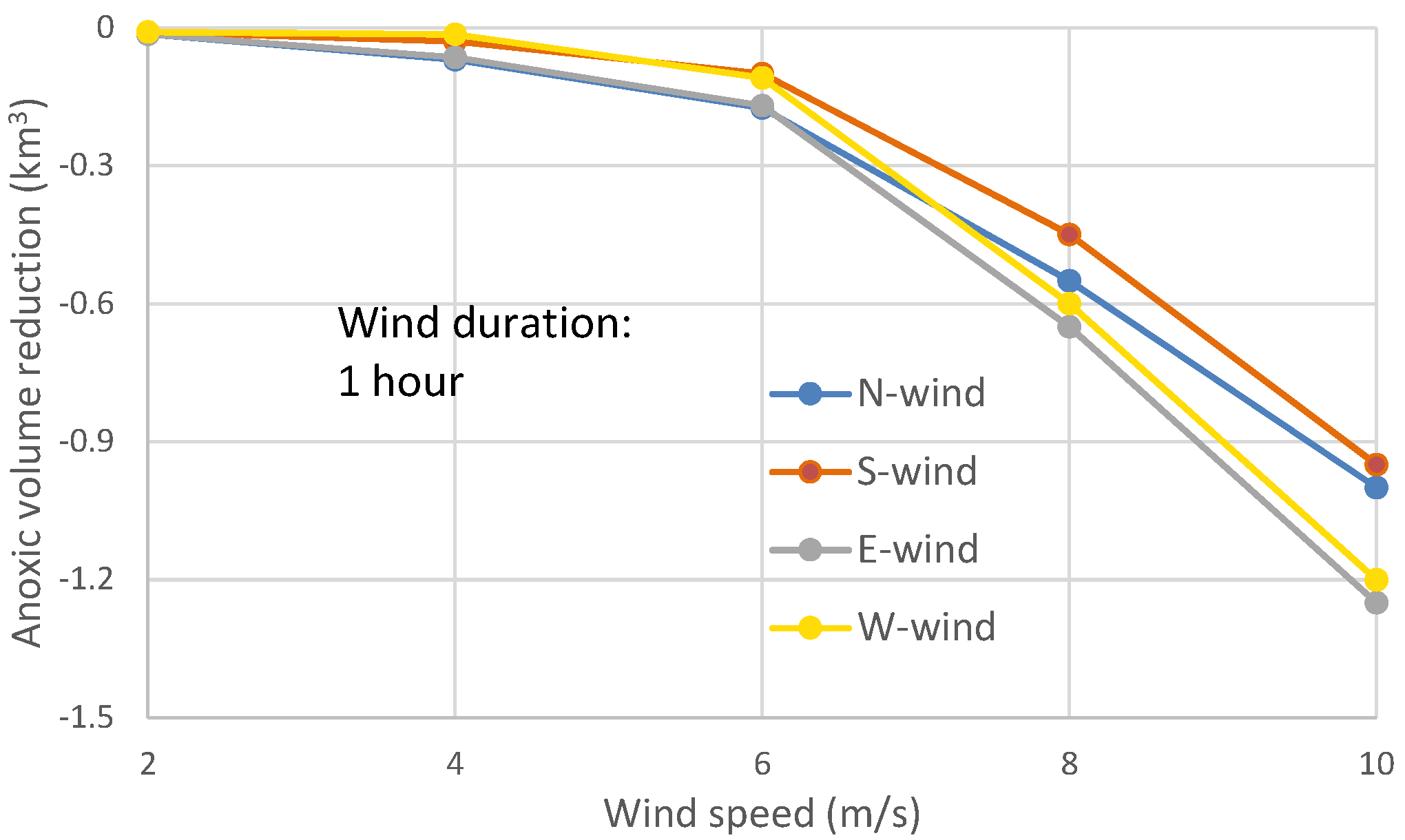
3.2. Relative Anoxia Reduction by Wind Directions in One-Hour Wind Events
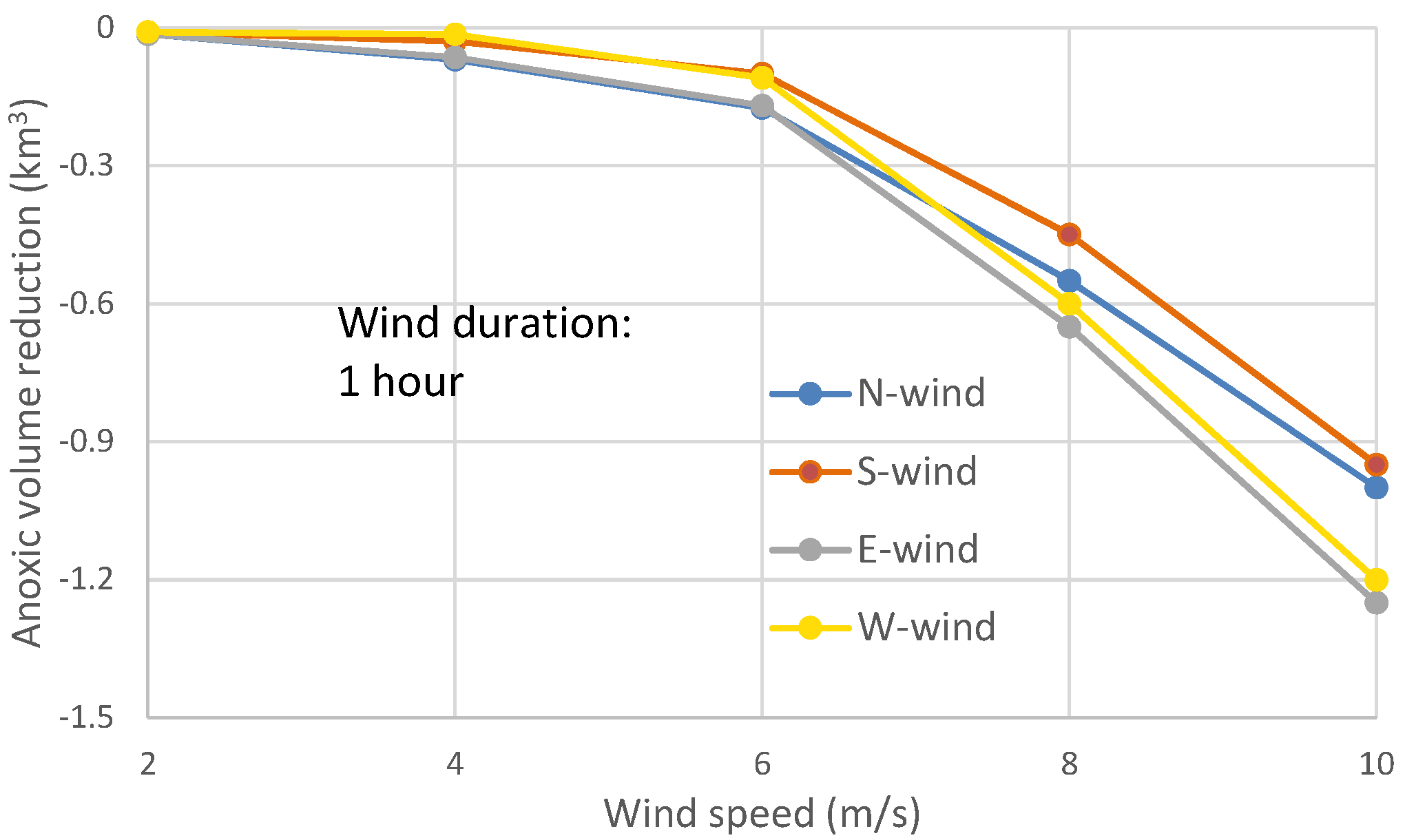
Although there rarely exists a continuous calm period for a few days with only one hour of wind, it is worthwhile to conduct model experiments to assess the response of anoxia to one-hour wind events. In the model experiments of one-hour winds, the post-wind recovery of anoxia appeared soon after the wind event stopped. The peak anoxia reduction occurred about 4–5 h after the end of the wind event for wind speeds at 4–8 m/s, and occurred sooner at lower wind speeds. The maximum reduction of anoxic volume was less than 0.3 km
3 at wind speeds of 6 m/s or lower (
Figure 11), near the same magnitude of the tidal influence on anoxic volume (
Figure 9). The differences in anoxia reduction among wind directions in wind speeds less than 4 m/s were not prominent, most are less than 0.1 km
3.
The wind-induced along-channel circulation in one-hour wind events did not significantly influence the center of anoxia, while the cross-channel circulation became more important in reducing the peak anoxic volume (
Figure 11). The modulation of the PN-type bathymetry caused easterly winds to reduce more anoxia than westerly winds, and northerly winds to reduce more anoxia than southerly winds.
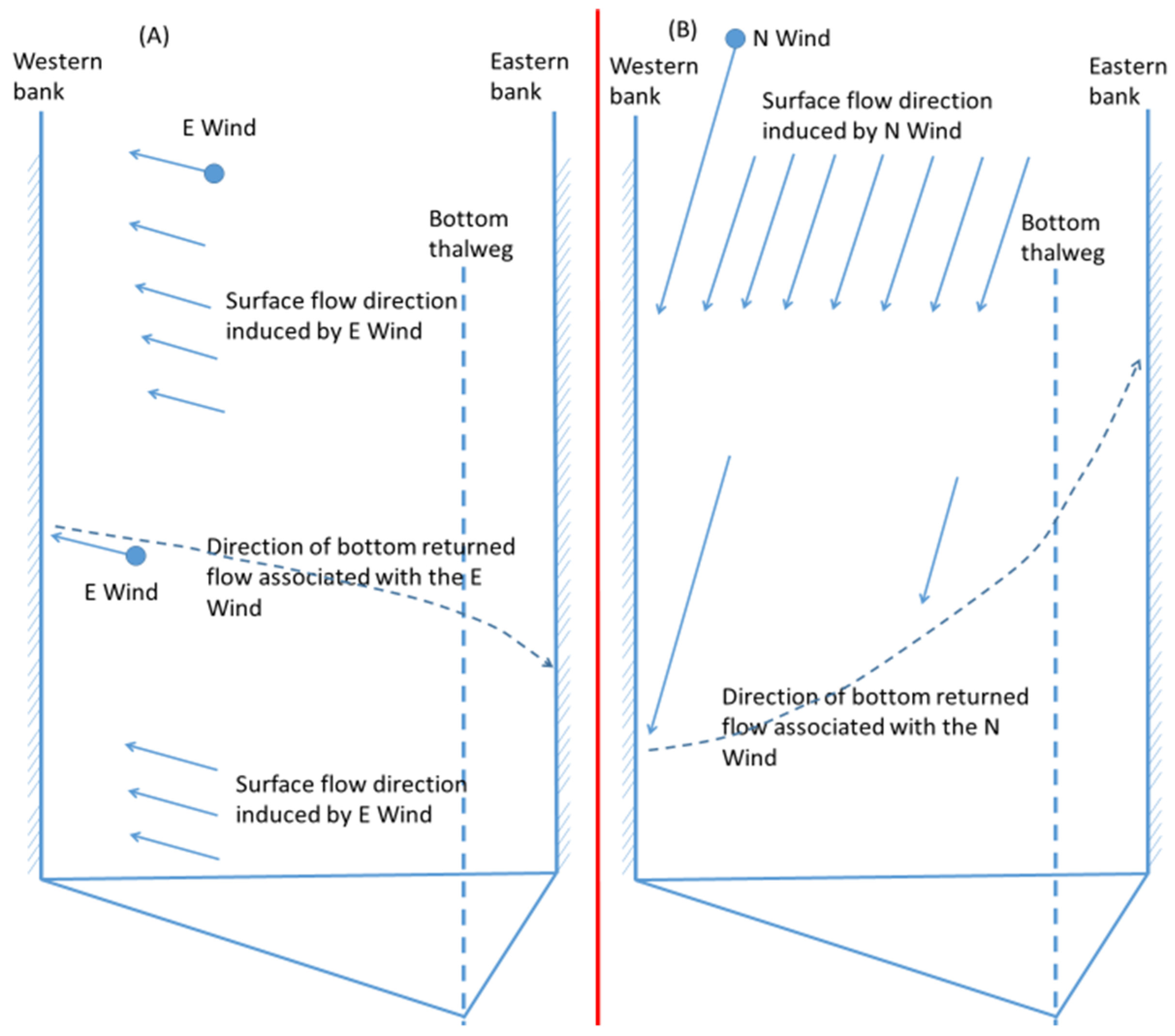
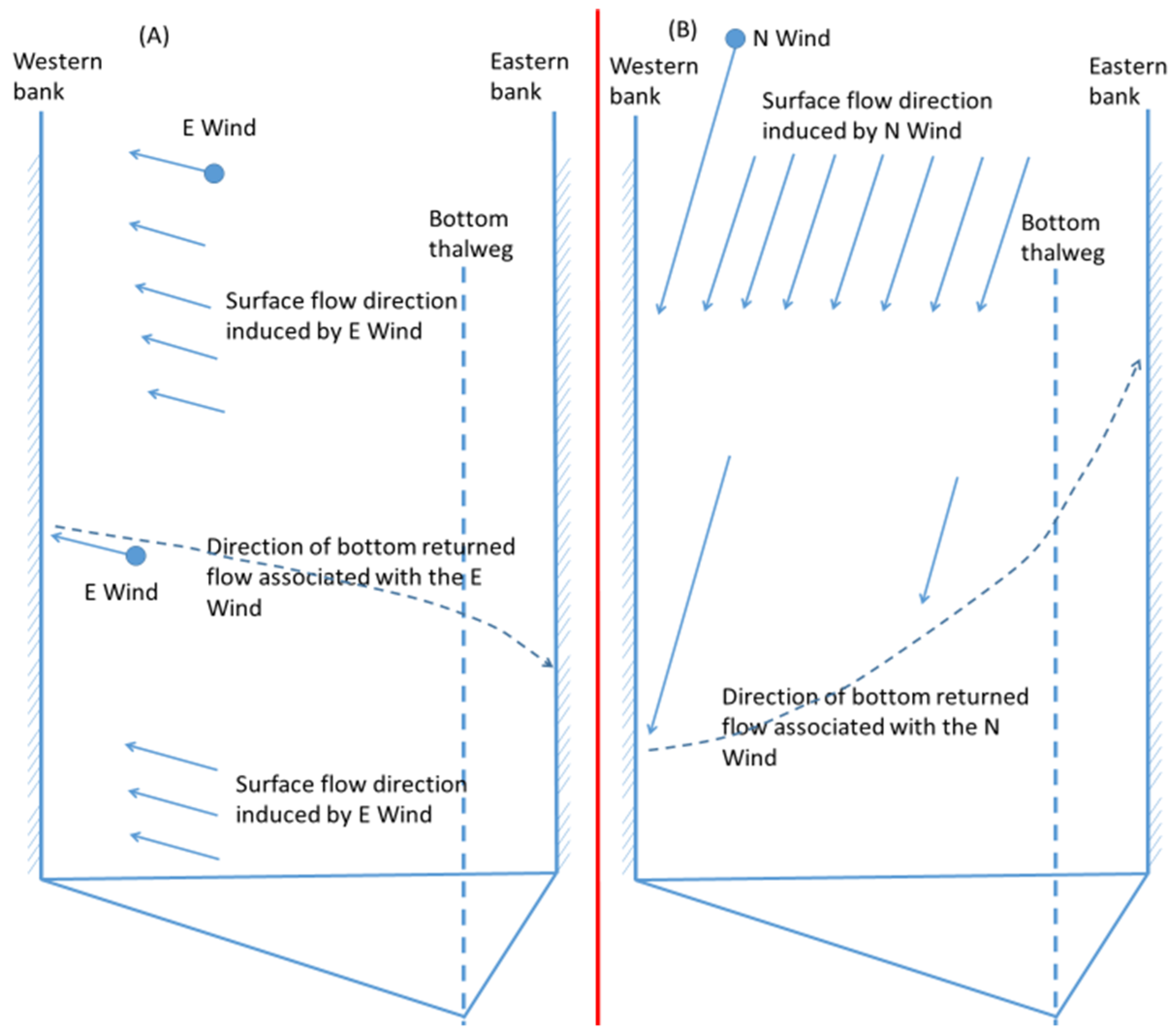
In two-day wind events, northerly and southerly winds reduced more anoxia than easterly and westerly winds, but in most cases of one-hour winds easterly (or westerly) winds reduced more anoxia than northerly (or southerly) winds, as explained in the following using
Figure 12. In
Figure 12, the dashed arrow approximates the returned flow along the bed from the downwind shore. The wind-induced cross-channel circulation can complete its cycle in an hour, and the deepest bottom was influenced to a certain degree, to a greater extent by easterly winds than by northerly winds, because the travel distance of the returned bottom current was shorter in easterly winds. Thus, in the one-hour wind events easterly winds had greater anoxia reduction than northerly winds. However, if the wind event continued for several hours, the wind-induced cross-channel circulation by northerly and easterly winds would involve several cycles. Because the bottom was constantly influenced by the wind-induced circulation, and not by travel distances, the anoxia reduction by wind-induced cross-channel circulation by northerly and easterly winds became similar. The northerly wind had stronger direct wind mixing (due to a longer fetch) and stronger longitudinal straining than the easterly wind, and the influence of longitudinal straining was stronger later in the wind period than the first hour of the wind event. Overall, northerly winds caused greater destratification and anoxia reduction than easterly winds in the later period of two-day wind events. Similarly, westerly winds reduced more anoxia than southerly winds if the wind event only lasted for one hour. This phenomenon is more prominent for greater wind speeds (8 or 10 m/s).
In summary, the peak anoxia reductions between opposite wind directions in the one-hour wind events (
Figure 11) are comparable to the anoxia reduction in the first 2–6 h of two-day wind events (
Figure 4) that are mainly controlled by wind-induced cross-channel circulation and wind induced direct mixing. These are different from the peak anoxia reductions in two-day wind scenarios (
Figure 8), since the latter generates more influence by wind-induced along-channel circulation. Generally, in one-hour wind events, the northerly winds effected a greater anoxia reduction than southerly winds, easterly winds had greater anoxia reduction than westerly winds (
Figure 11), and easterly (or westerly) winds had greater anoxia reduction than northerly (or southerly) winds.
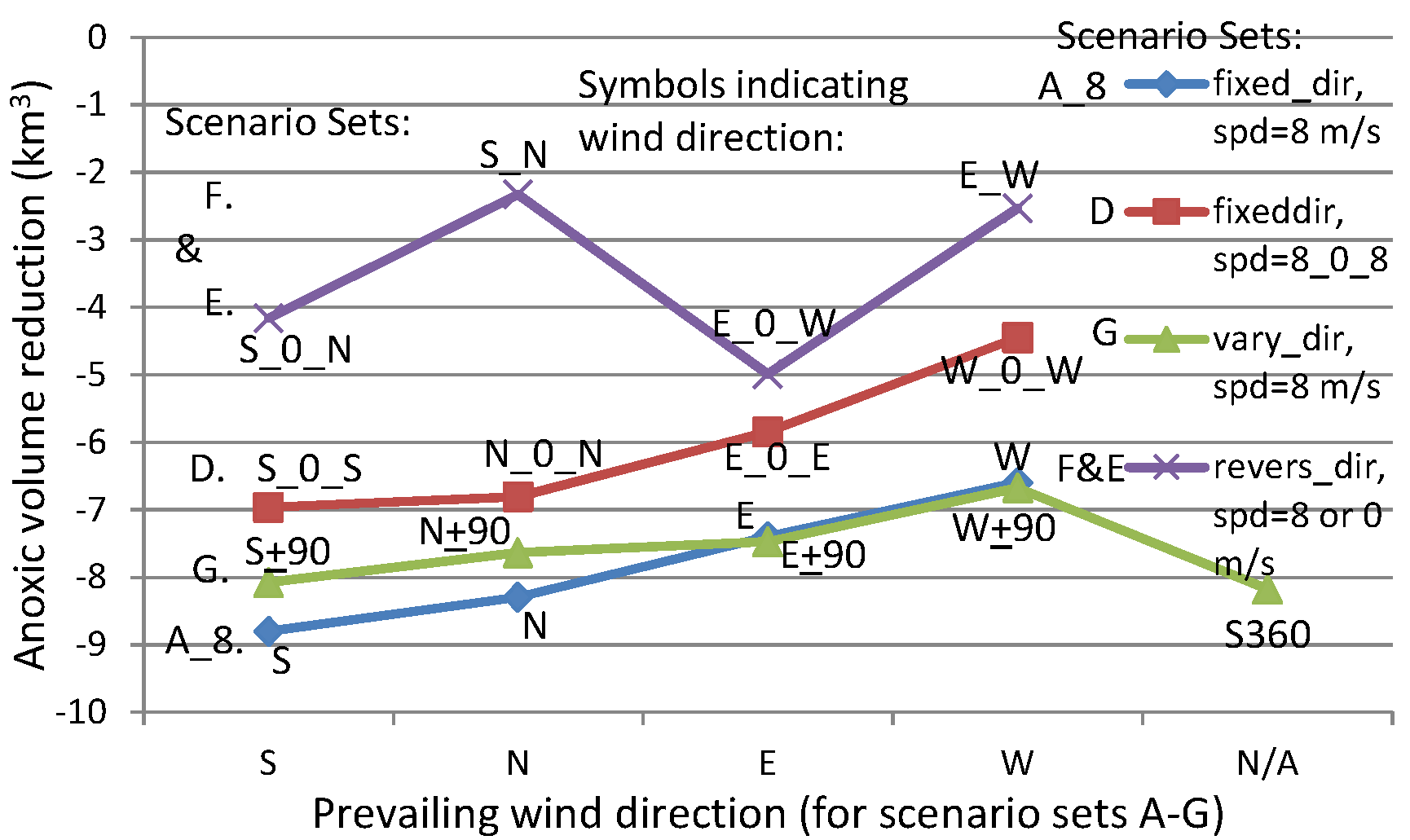
3.5. Winds of Hourly Reversing Directions: Scenario Sets F and E
Scenario S_N of Scenario Set F switches between southerly and northerly wind directions every hour, and is used to compare the southerly and northerly winds of Scenario A_8 (
Figure 15). Similarly, Scenario E_W is used to compare the easterly and westerly winds of Scenario A_8. All have speeds of 8 m/s over a two-day period. The frequent switching seemed to negate the effects of the preceding wind direction. The anoxia reduction by Scenario E_W was 4–5 km
3 less than the reduction by constant westerly or easterly winds, and the anoxia reduction by Scenario S_N was 6–7 km
3 less than the reduction by constant northerly or southerly winds (
Figure 15). With intermittent no wind between reversing wind directions (e.g., Scenario S_0_N or E_0_W) the reduction of anoxic volume was approximately 2 km
3 more than the S_N or E_W scenarios. Nevertheless, Scenario S_0_N and E_0_W still had a reduction in anoxic volume of 3–4 km
3 less than the constant unidirectional southerly or northerly winds and easterly or westerly winds, respectively.
The anoxic volume reduction by Scenario E_0_W lies in between reductions modeled in Scenarios E_0_E and W_0_W. Scenario E_0_E had more frequent easterly winds than the other two scenarios, and yielded more anoxia reduction. The intermittent no wind between switching directions in Scenario E_0_W weakened the cancellation of anoxia reduction processes by the two switching wind directions, therefore, still had a greater anoxia reduction than Scenario W_0_W. Cross-channel circulation was important in destratification by easterly and westerly winds. The widths of cross channel around the anoxic center were narrow. The wind’s effect could be effectively realized within the one-hour period of calm. The next phase of wind in opposite direction generated the next round of reduction. Because of more frequencies in easterly wind, Scenario E_0_W had a greater anoxia reduction than Scenario W_0_W. While, Scenario E_W had no calm period between changing directions, significantly reducing destratification.
Compared to Scenario N_0_N and S_0_S, Scenario S_0_N had an approximately 3 km3 lesser reduction of anoxic volume. The wind-induced circulation of along-channel transport by northerly or southerly winds could not be completed within a single hour. Reversing wind directions weakened the actions of the preceding wind direction. Therefore, Scenario S_0_N had weaker anoxia reduction than both Scenarios S_0_S and N_0_N. Scenario S_N switched wind directions between southerly and northerly each hour without a calm period, and yielded even weaker anoxia reduction. This model experiment confirms that wind-induced along-channel circulation affects the anoxic center much more slowly than wind-induced cross-channel circulation.
3.6. Winds with Rotating Directions at Fixed Speeds of 8 m/s: Scenario G
A comparison of anoxia reduction among scenarios with ± 90 degrees rotation about a central direction every 4 h (Scenario Set G) and the Core Scenario was also completed. For a scenario rotating about a southerly or northerly central direction (S ± 90 or N ± 90), the anoxic volume reduction was about 0.6 km
3 less than that caused by the winds of fixed southerly or northerly direction (
Figure 15). This can be attributed to the addition of easterly and westerly winds (
Table 2), which had shorter fetch and generated weaker destratification than southerly or northerly winds. For scenarios rotating about an easterly or westerly central direction (i.e., E ± 90 or W ± 90), anoxia reductions were slightly increased than the corresponding easterly (E) wind or westerly (W) wind of Core Scenario A_8, because of the inclusion of southerly and northerly winds.
Scenario S_360 had a decreased reduction in anoxia than the wind in a constant southerly direction did, as expected. Surprisingly, it also reduced more anoxia than Scenario S ± 90 did, and the causes are unclear. It is hypothesized that a continuous rotating direction is more effective in destratification and anoxia reduction than a rotation backwards along an already traversed path that could weaken the anoxia reduction effects of a prior wind direction.
These scenarios indicate that turbulence induced by winds of gradual changing directions at a similar speed generally can continue to weaken stratification. In many cases, directional change of 90 degrees does not significantly cancel out wind induced circulations. It is more likely that conditions in the Chesapeake Bay often lie somewhere in between scenarios G and D. For time scales of a few days, in most cases a prevailing southerly wind would cause greater anoxia reduction than a prevailing northerly wind, and would also hold true for prevailing easterly versus westerly winds. However, for month-long prevailing unidirectional wind the relative anoxia reductions between opposite wind directions can exhibit different anoxia reduction patterns, which will be discussed in the next section.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}